
Effects of Dendropanax morbiferus Leaf Extract on Sleep Parameters in Invertebrate and Vertebrate Models

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plant Material and Preparation of Extracts
2.3. Analysis of Antioxidant Capacity
2.4. Analysis of Polyphenol Components
2.5. Drosophila Melanogaster Stocks
2.6. Analysis of Sleep Behavior
2.7. Locomotor Activity Analysis
2.8. Analysis of Gene Expression via Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.9. Animals
2.10. Pentobarbital-Induced Sleep Test
2.11. Electroencephalogram (EEG) Analysis
2.12. Enzyme-Linked Immunosorbent Assay (ELISA)
2.13. Malondialdehyde (MDA) Assay
2.14. Statistical Analysis
3. Results
3.1. Antioxidant Activity and Polyphenol Components of DE
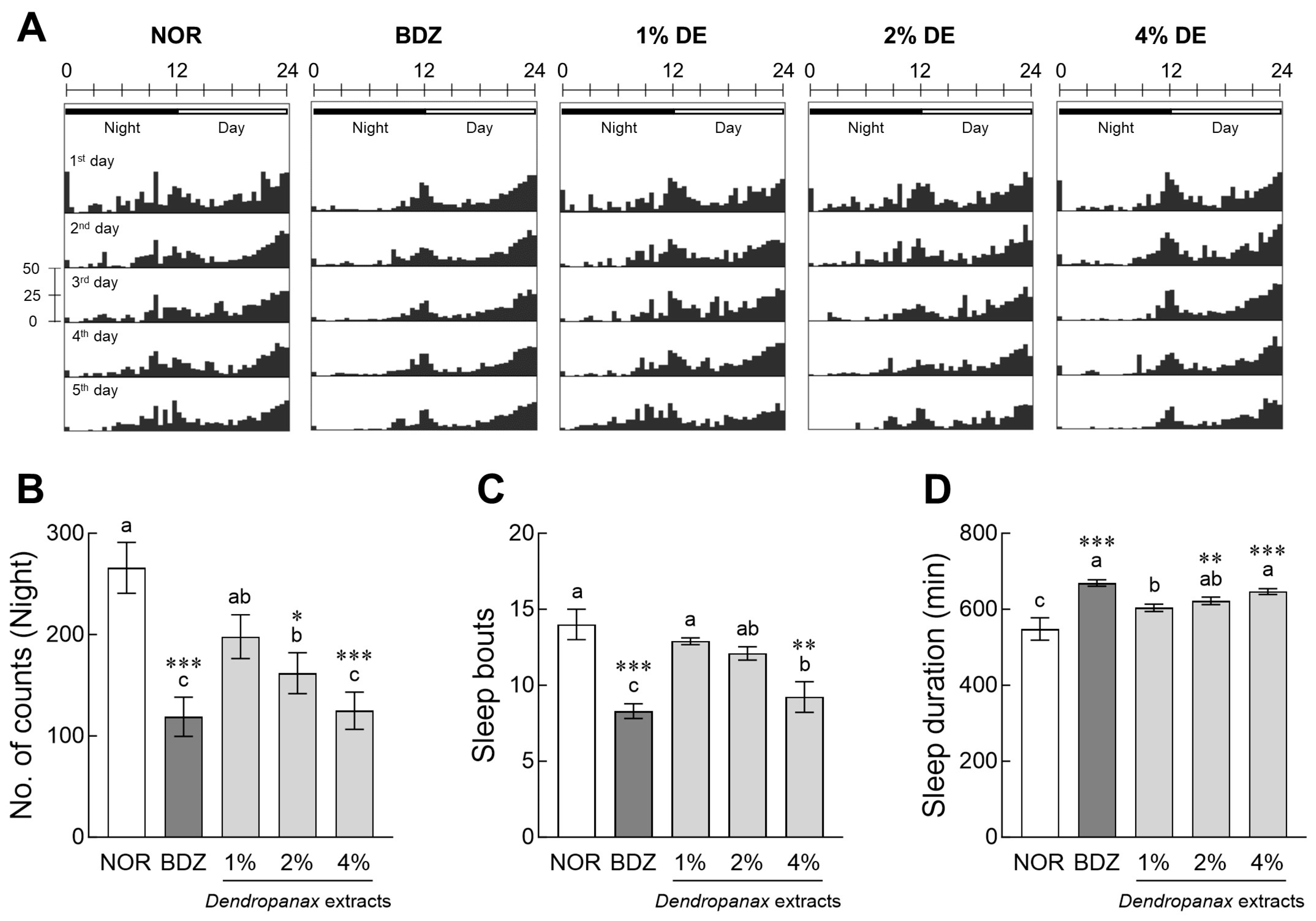
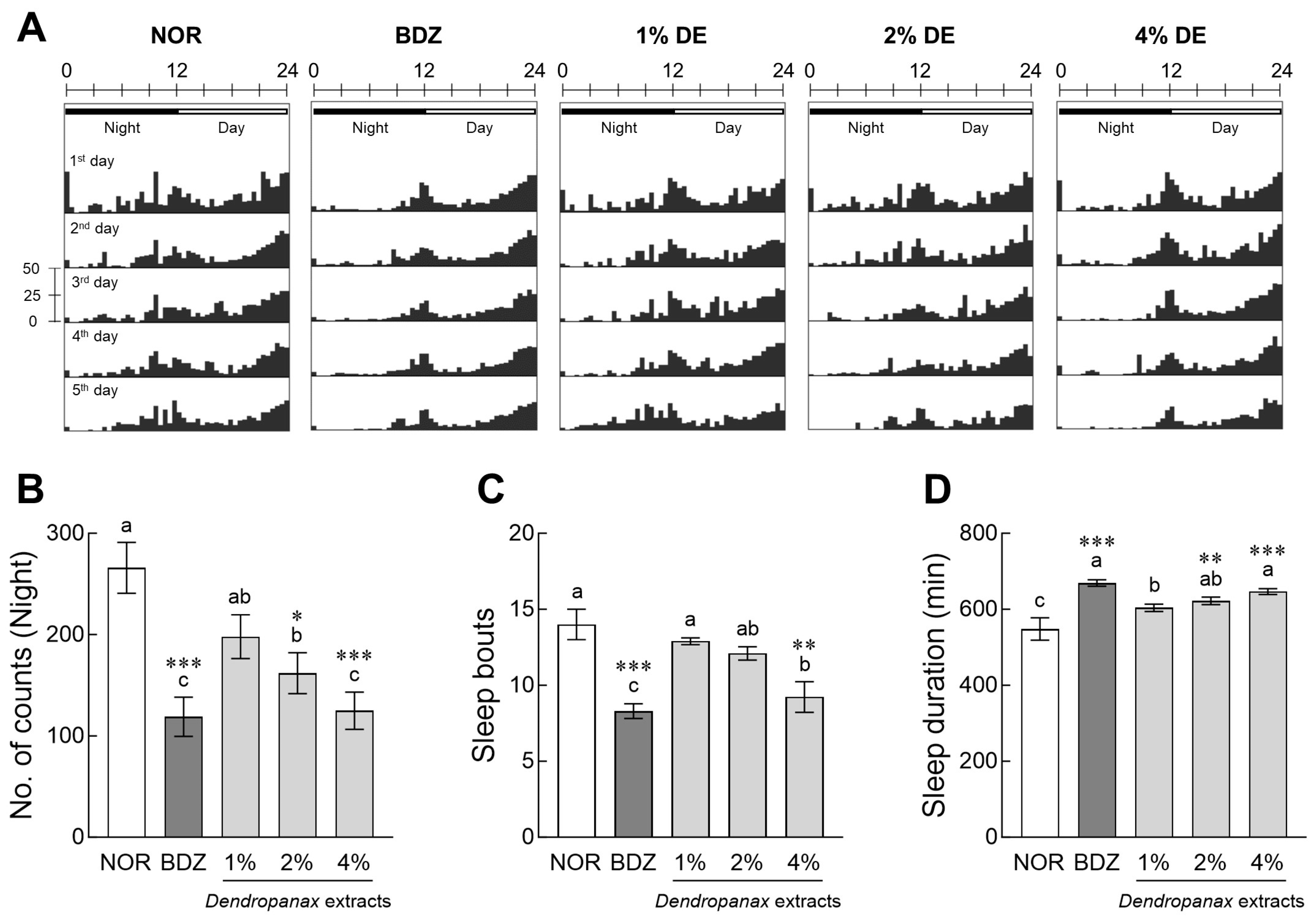
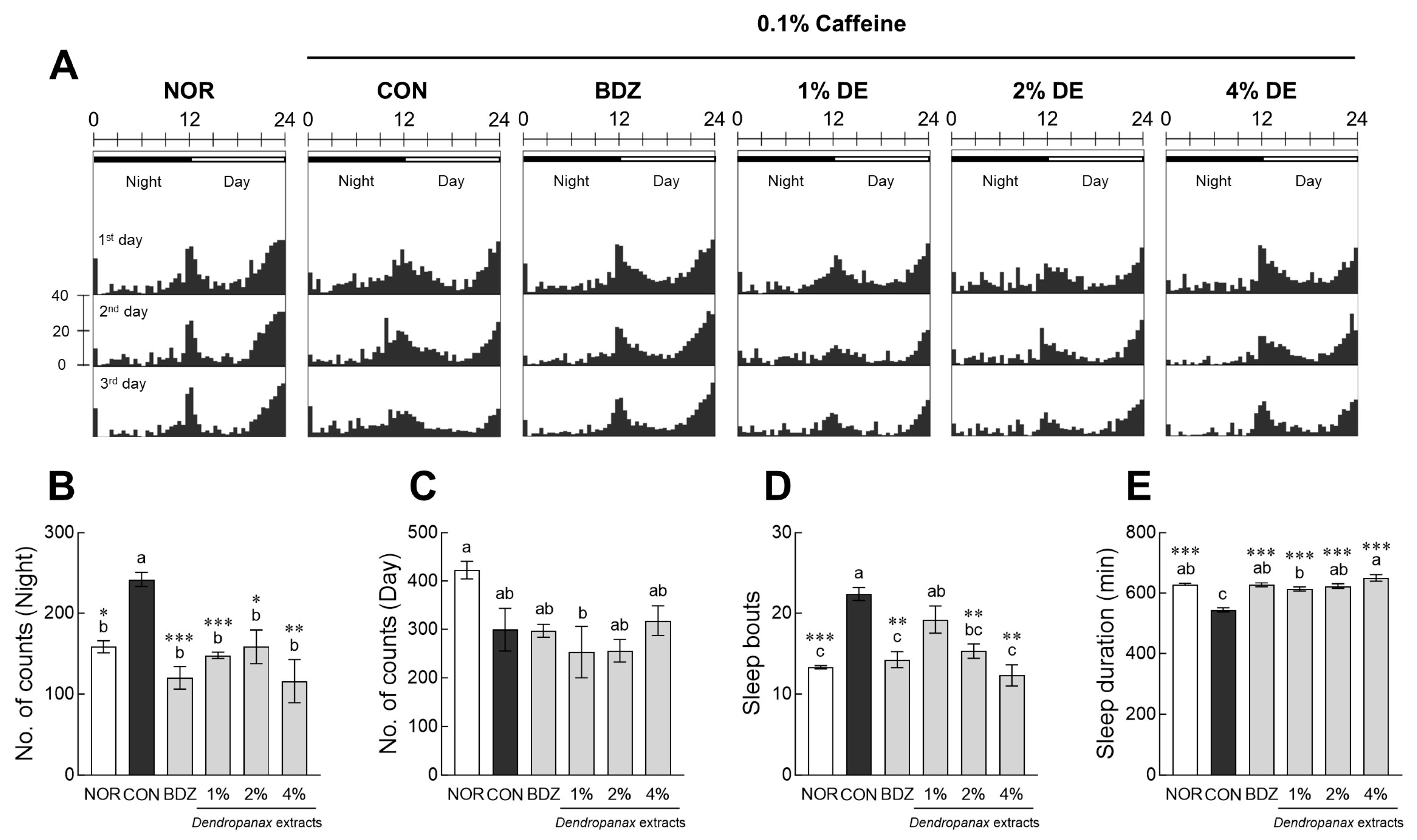
3.2. Evaluation of Sleep Activity of DE in Drosophila
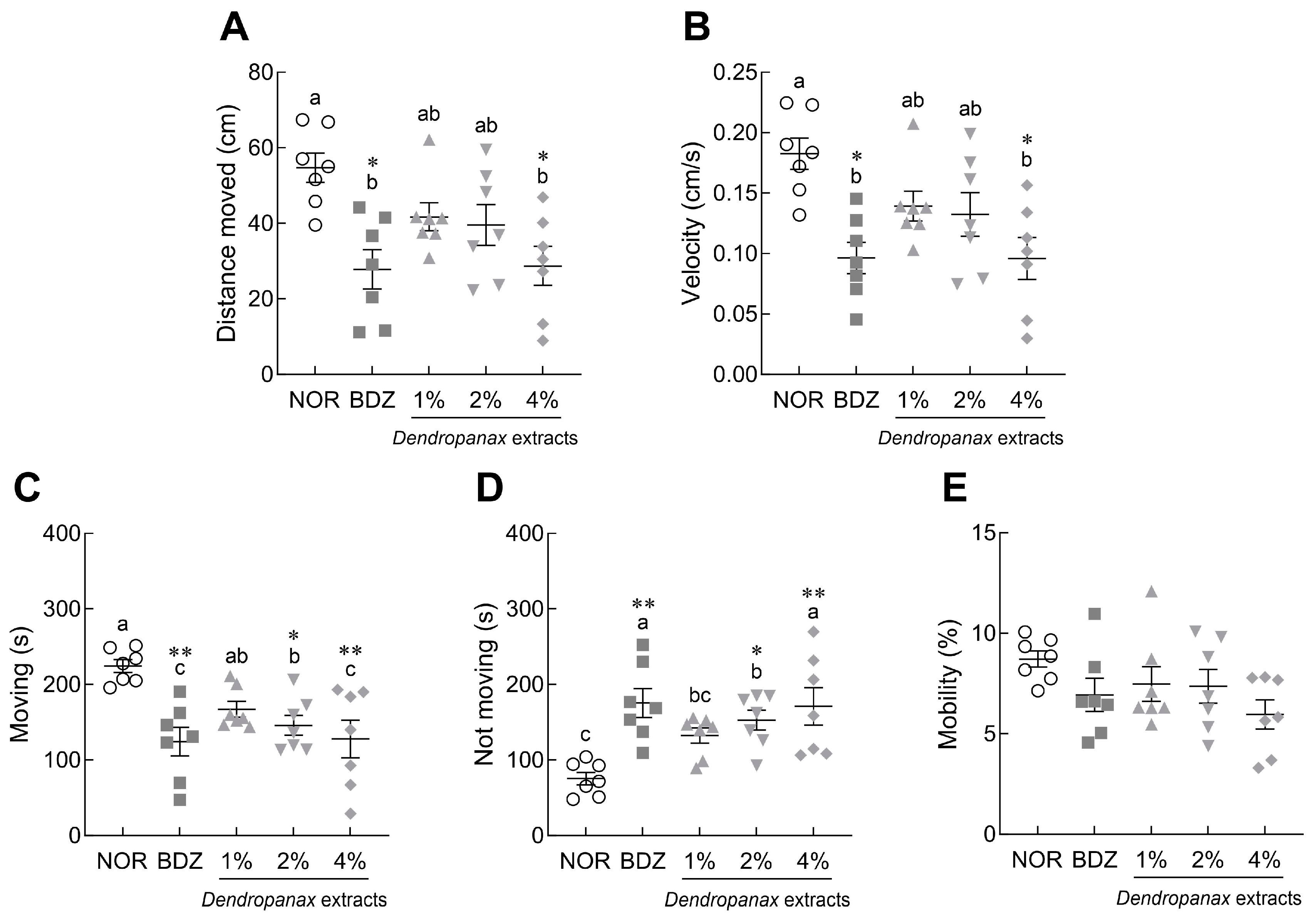
3.3. Effects of DE on Locomotor Activity in Drosophila
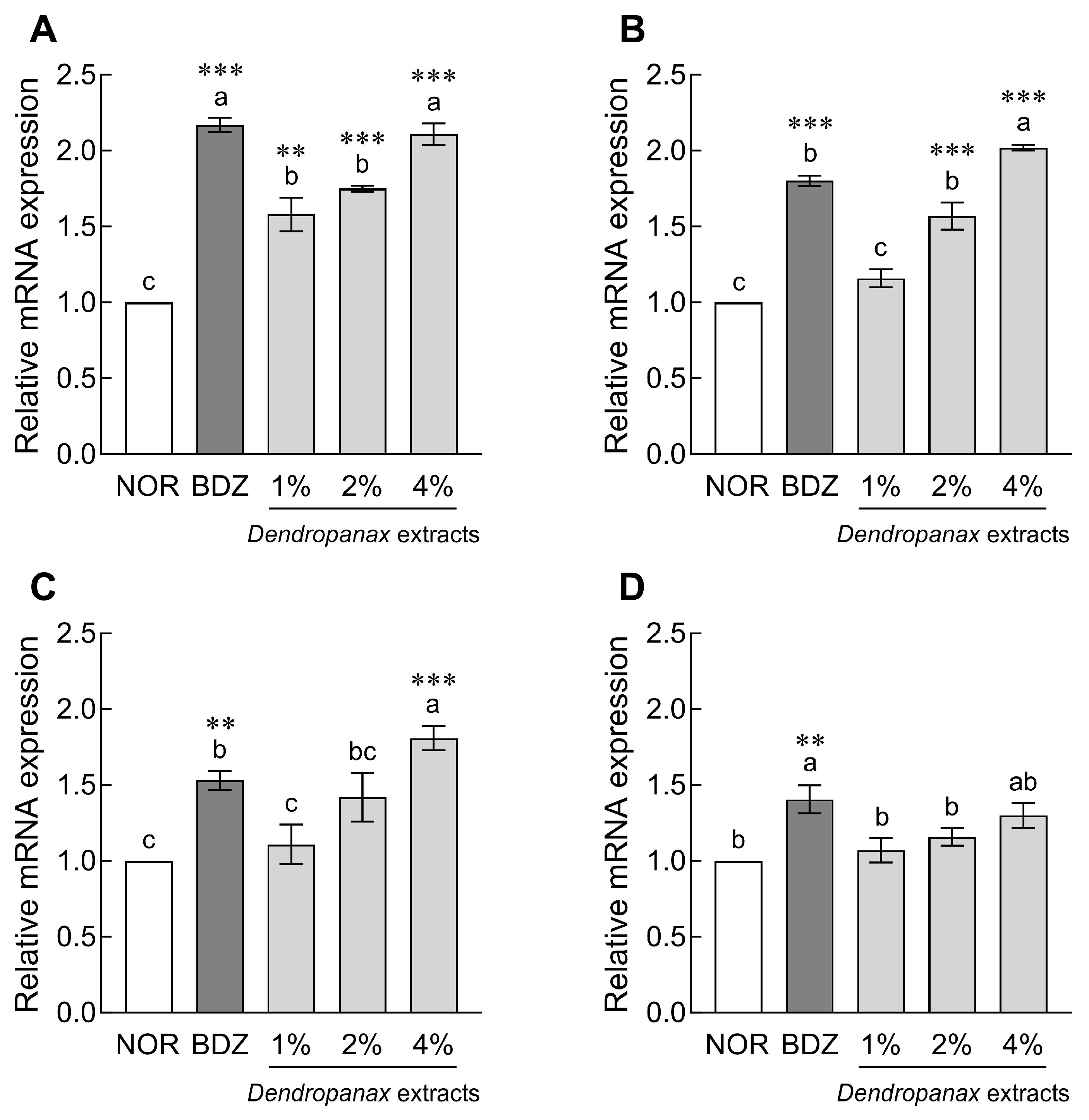
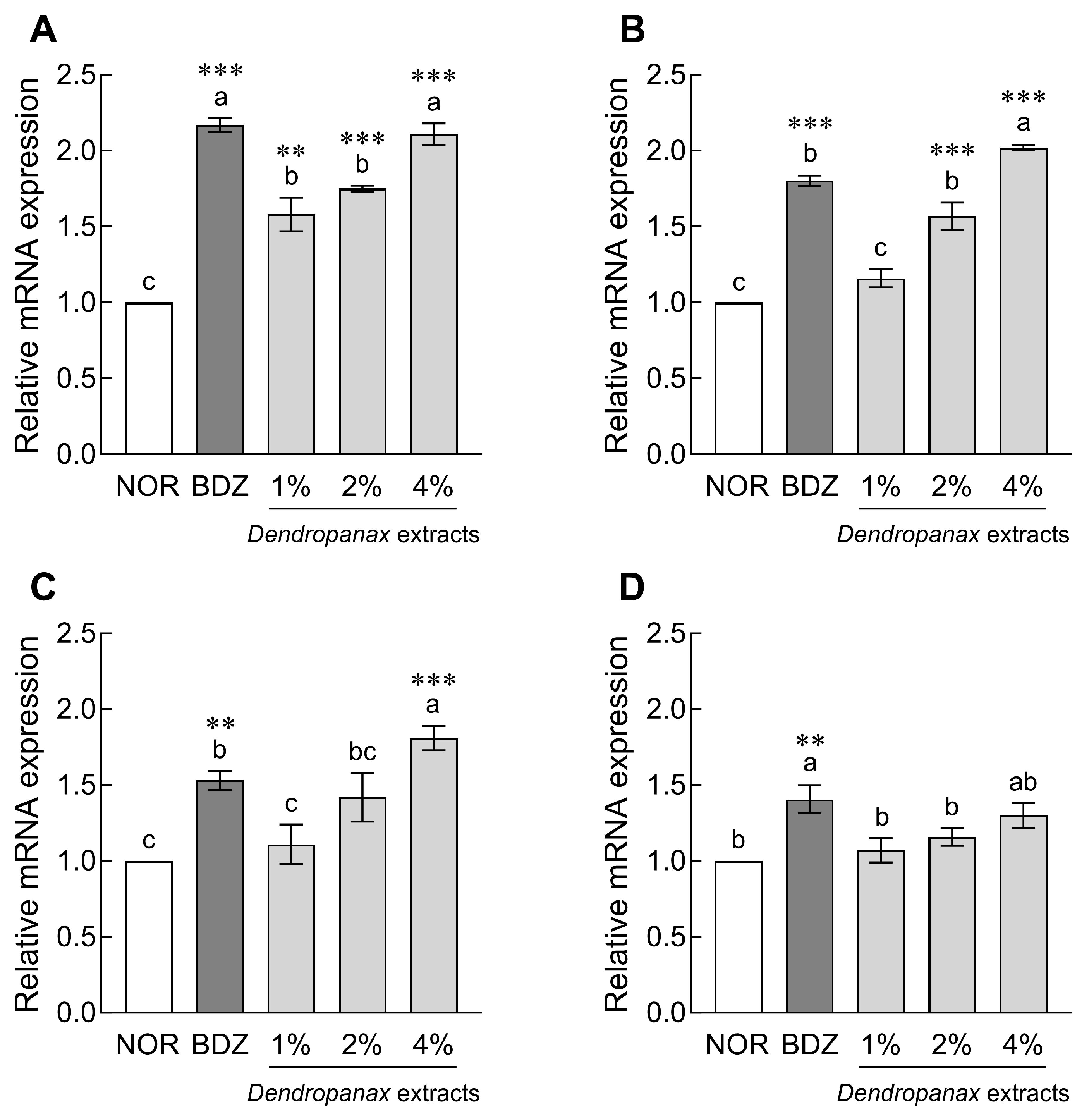
3.4. Effect of DE on mRNA Expression of Sleep-Related Receptors in Drosophila
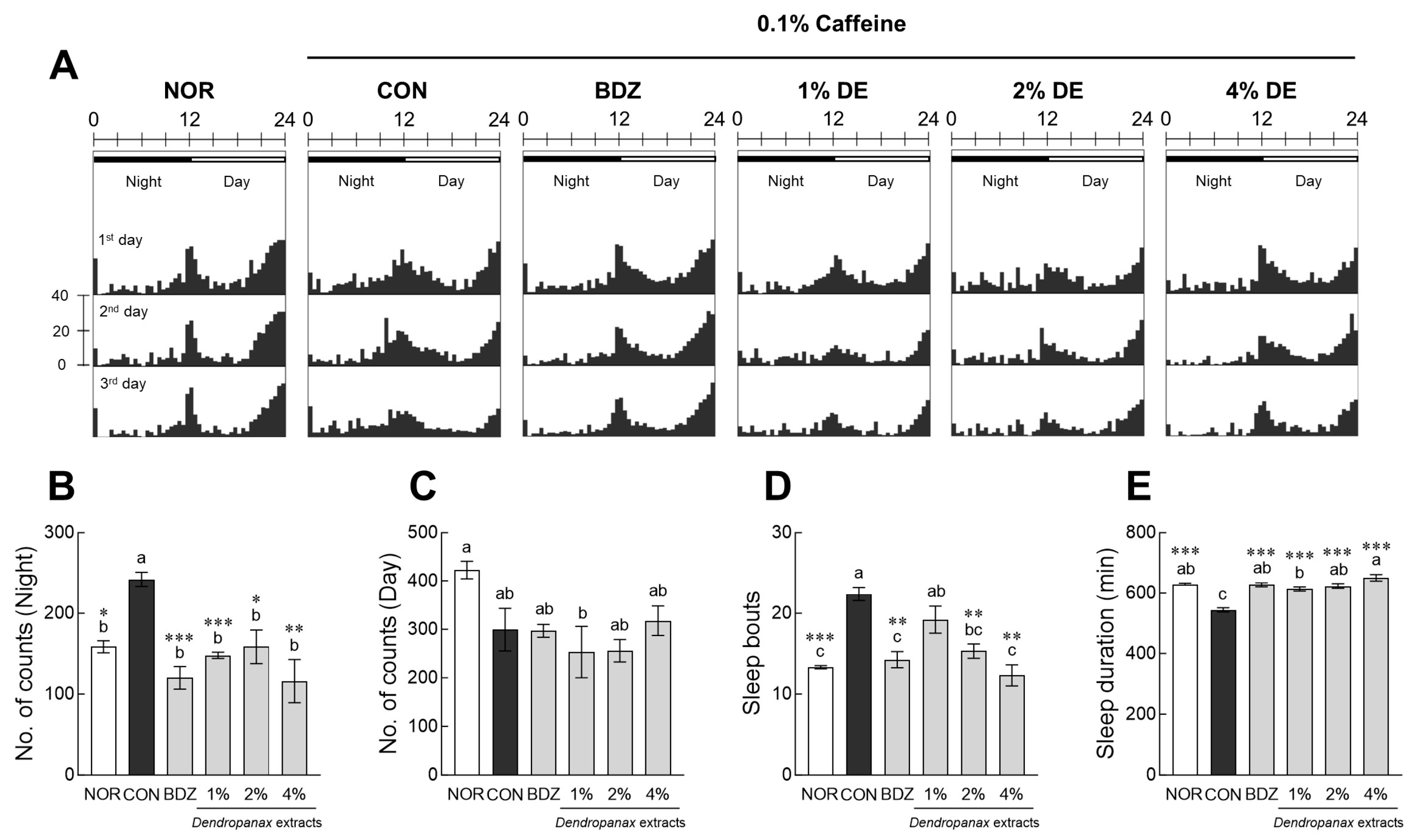
3.5. Effects of DE on Sleep Activity and Antioxidant Enzymes Expression in Insomnia-Induced Drosophila Model
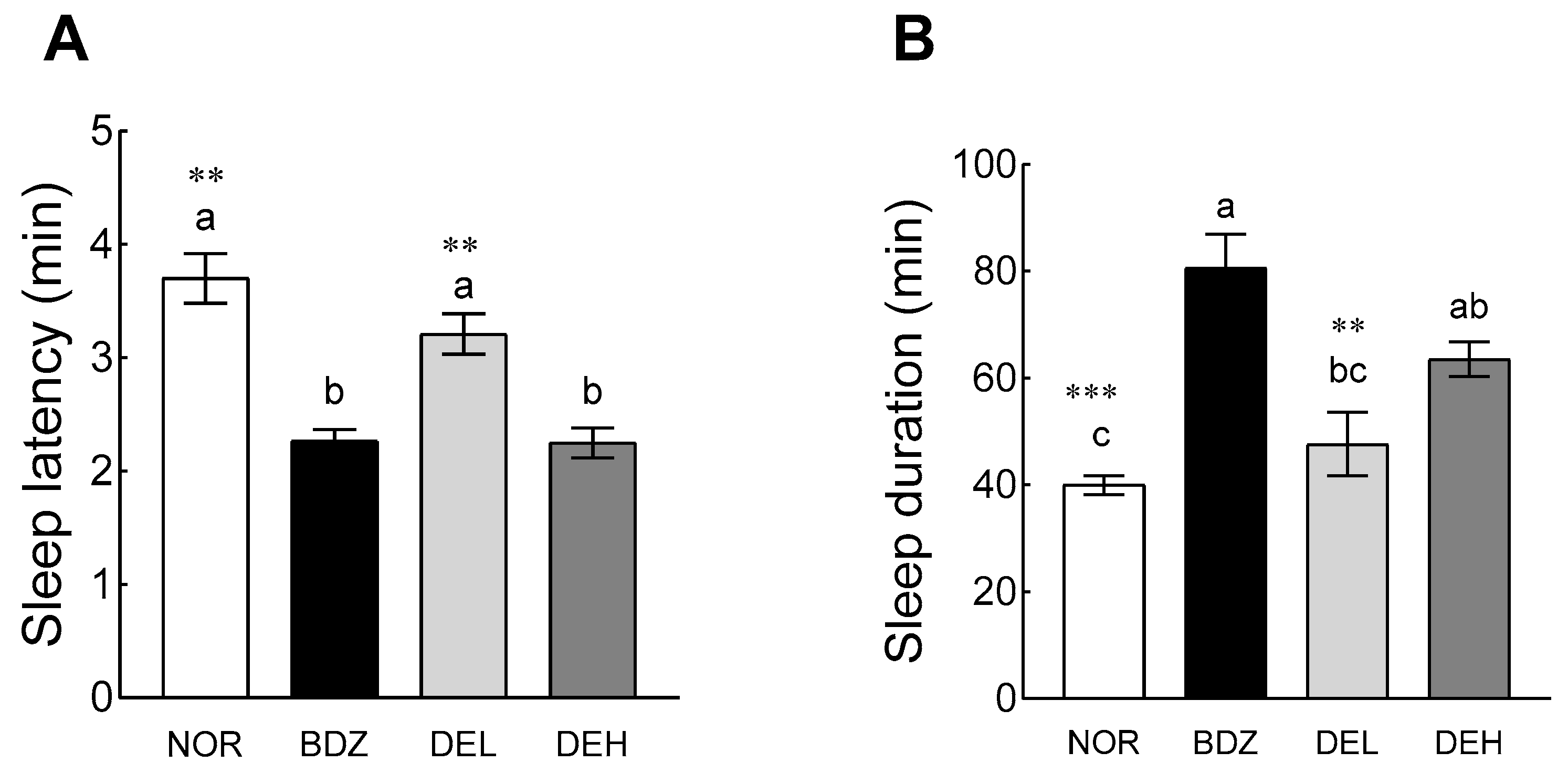
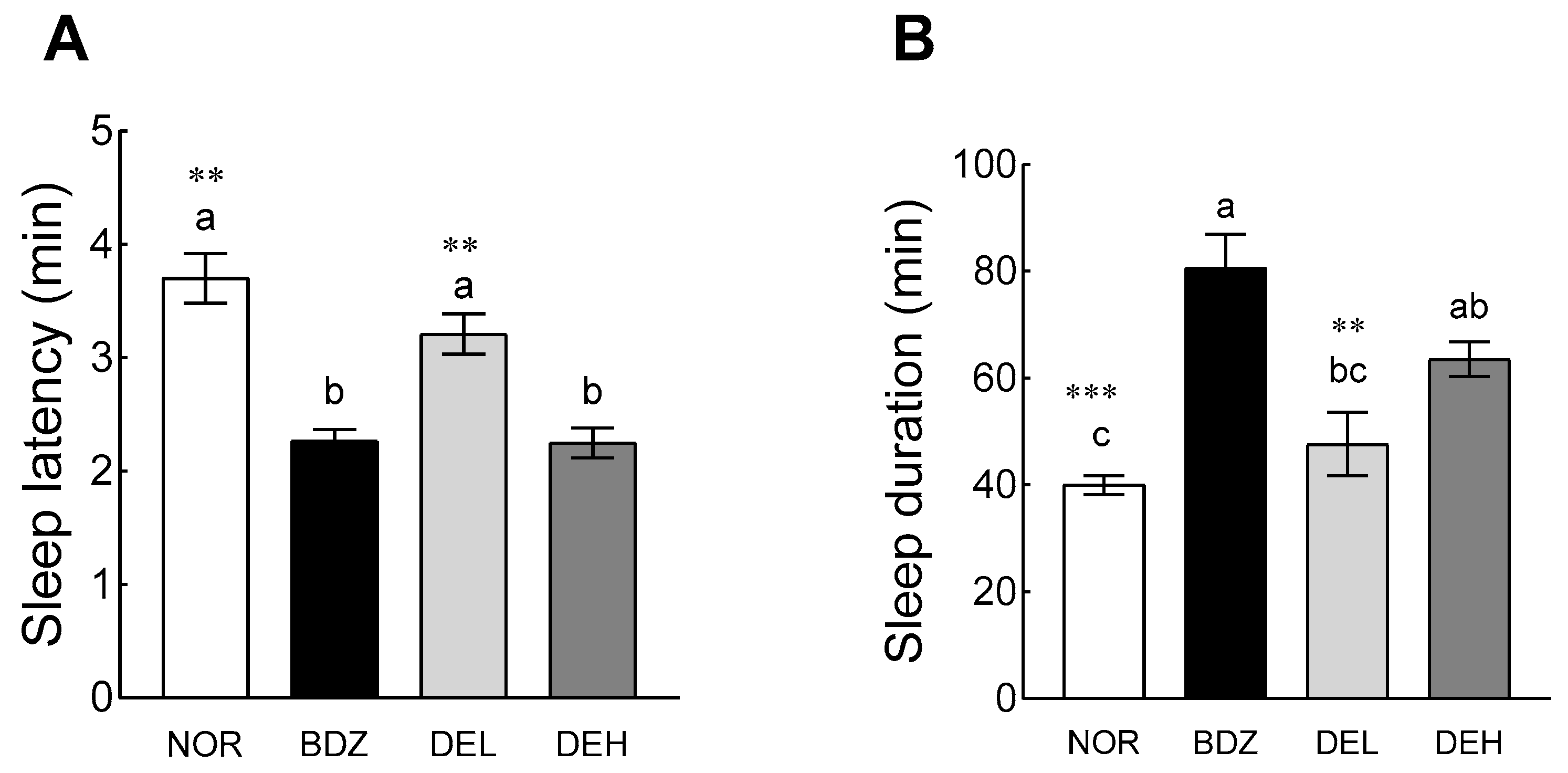
3.6. Pentobarbital-Induced Sleep Test
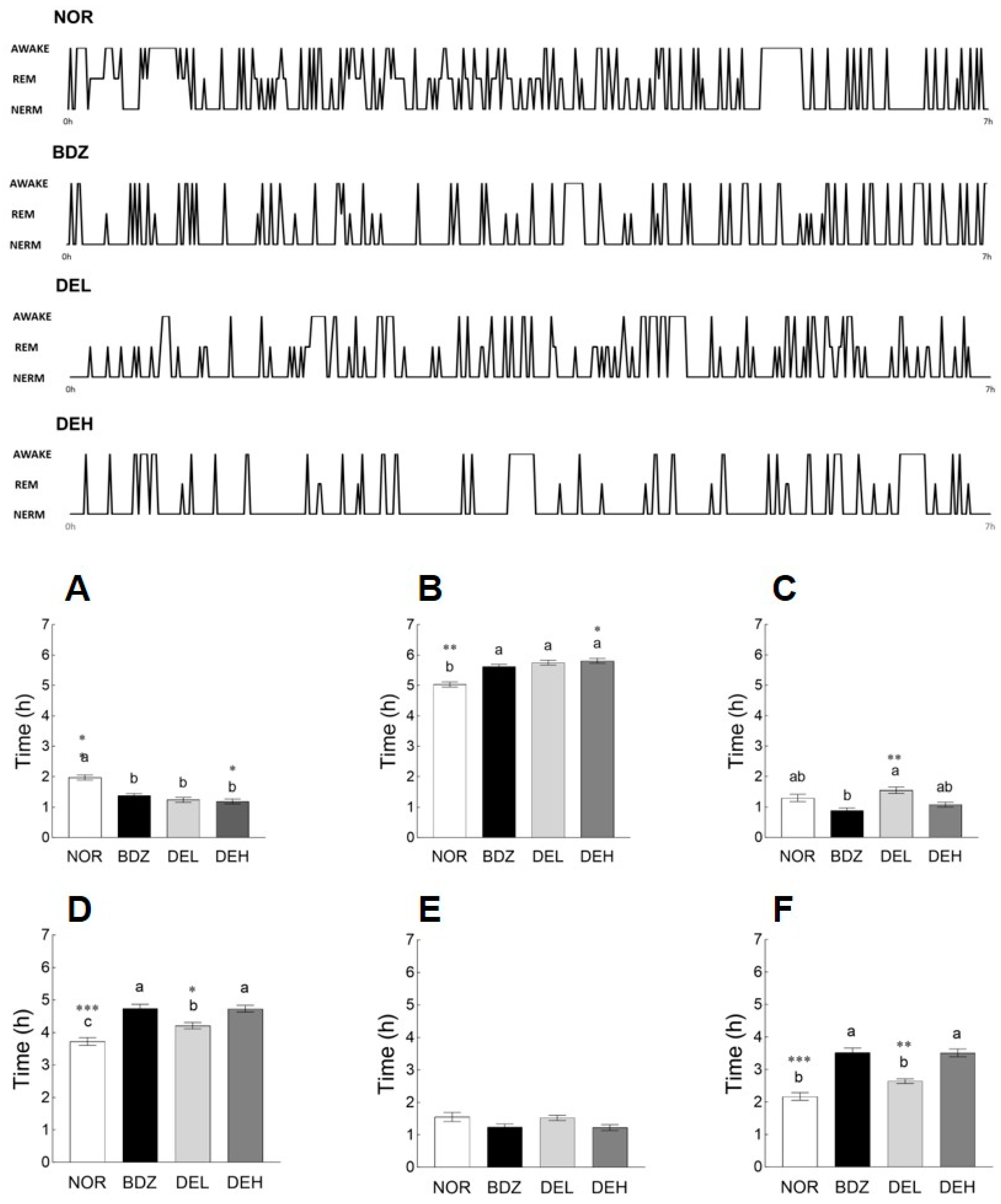
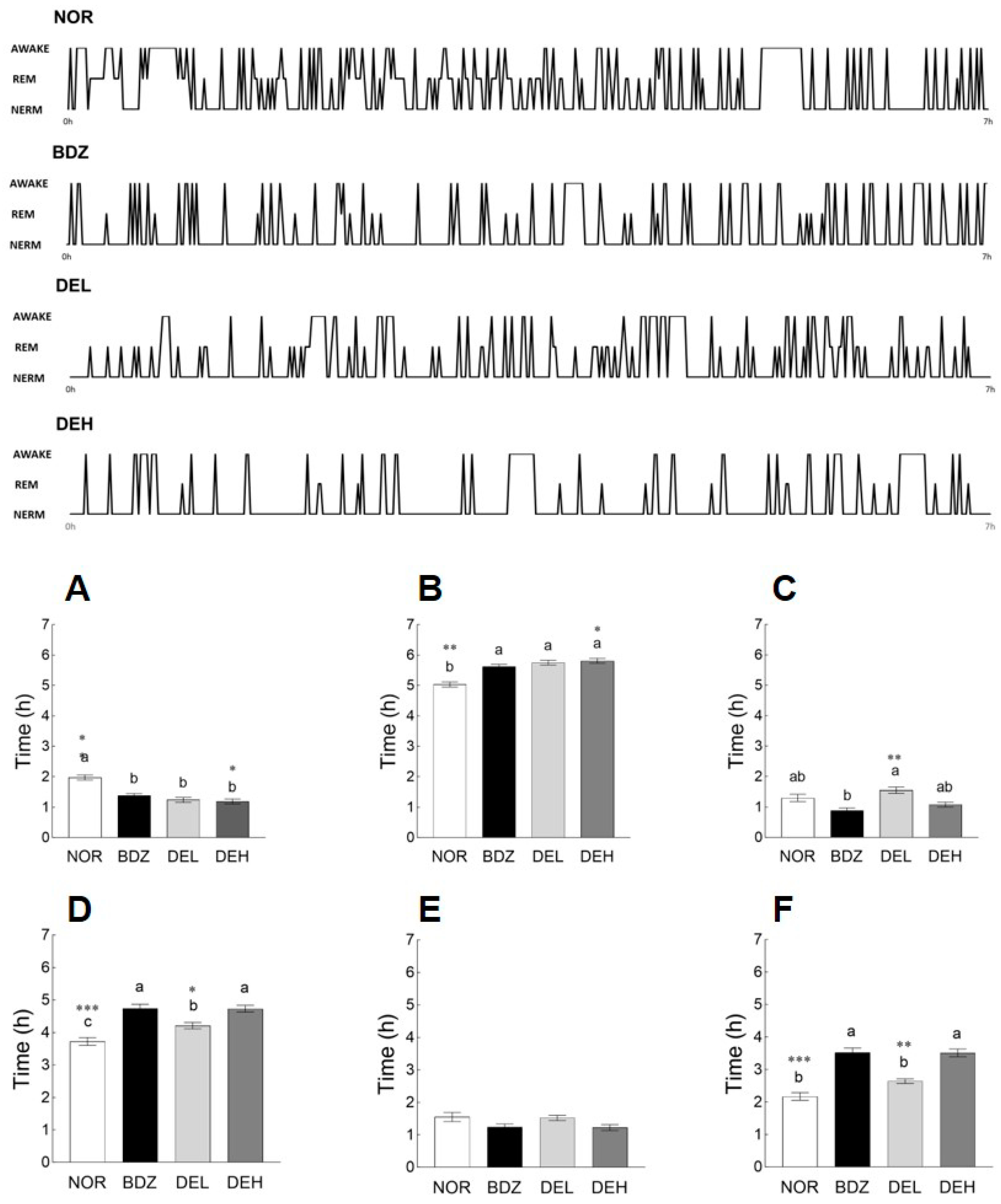
3.7. EEG Pattern Recording
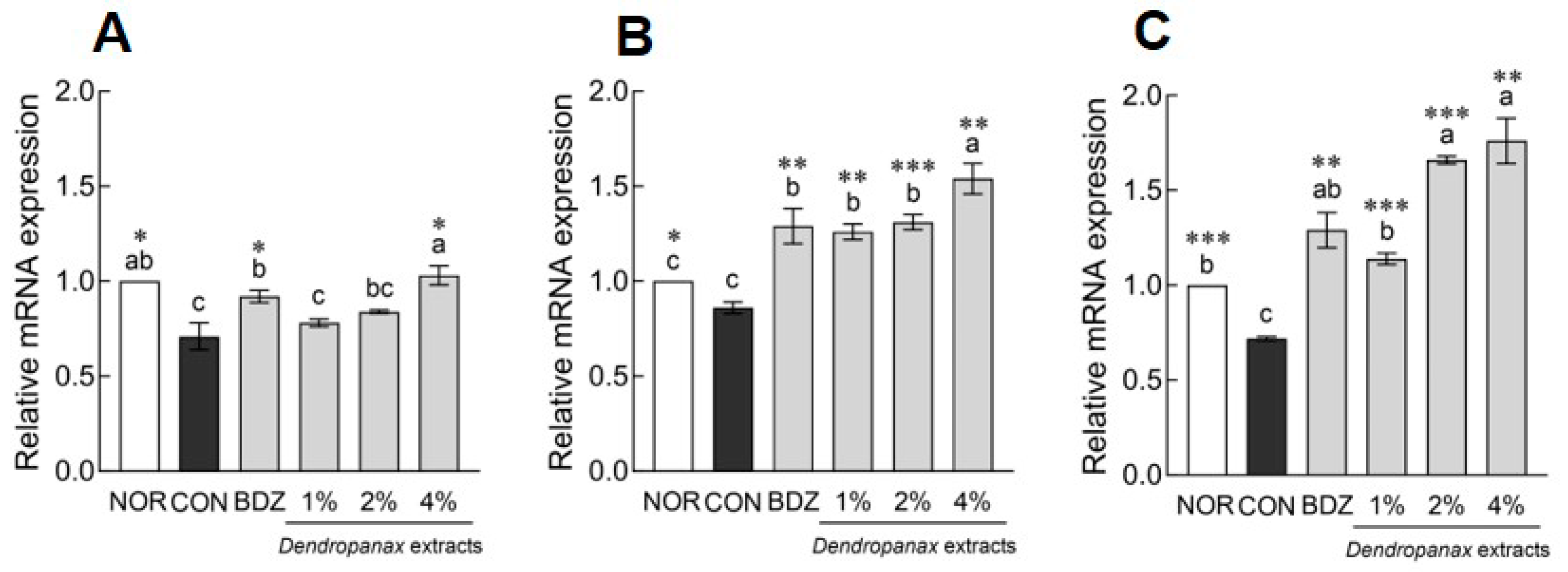
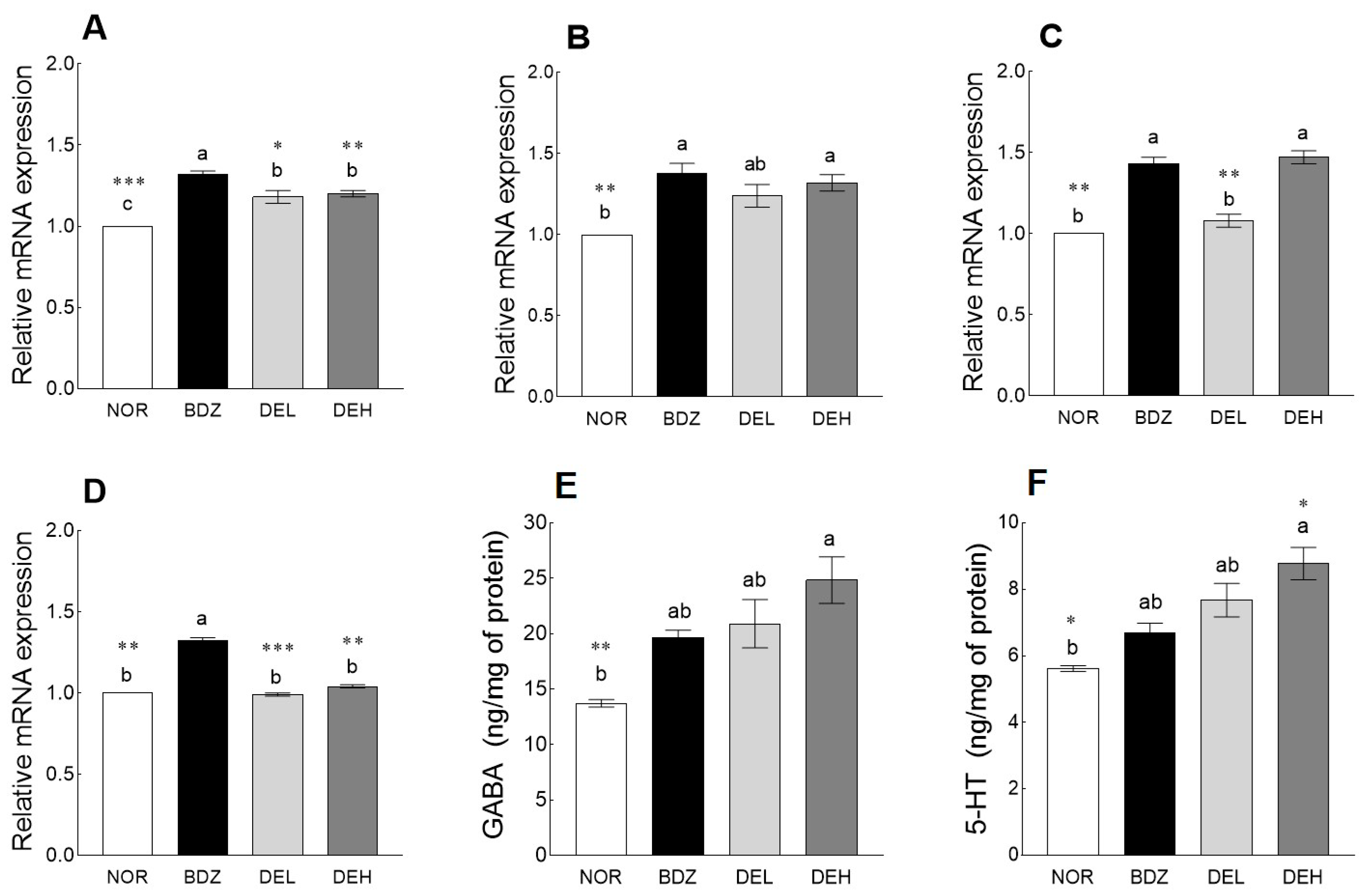
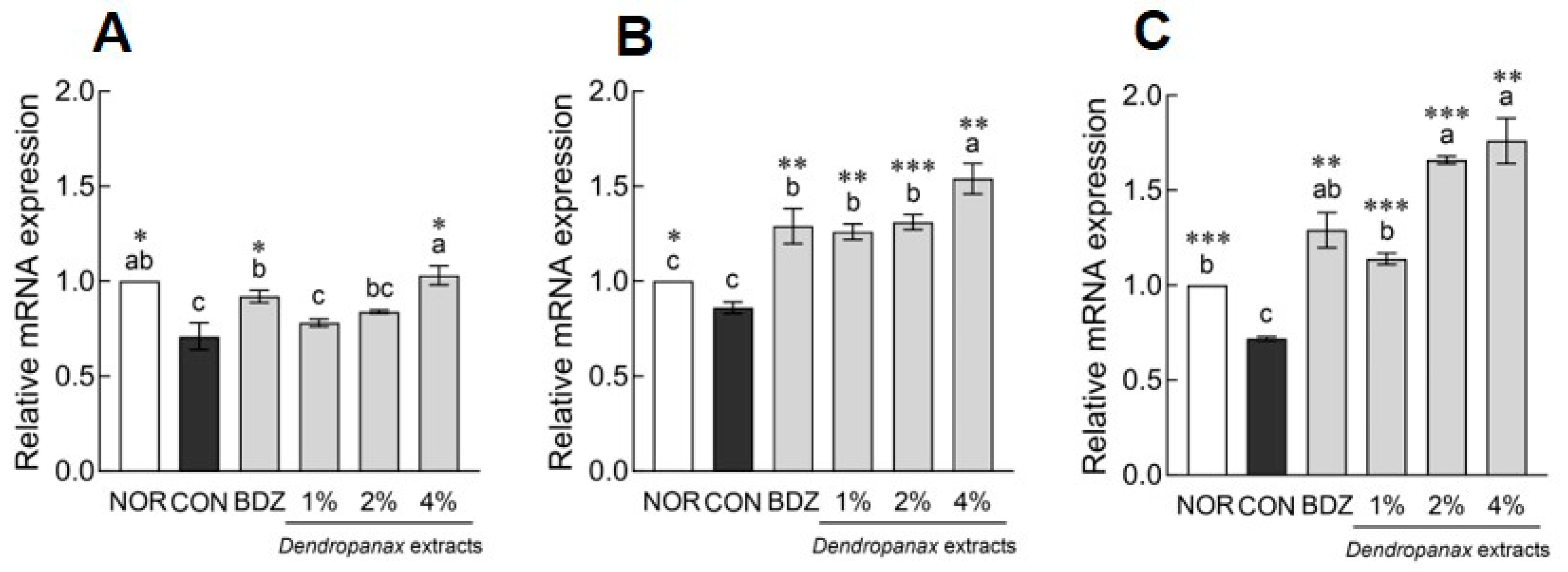
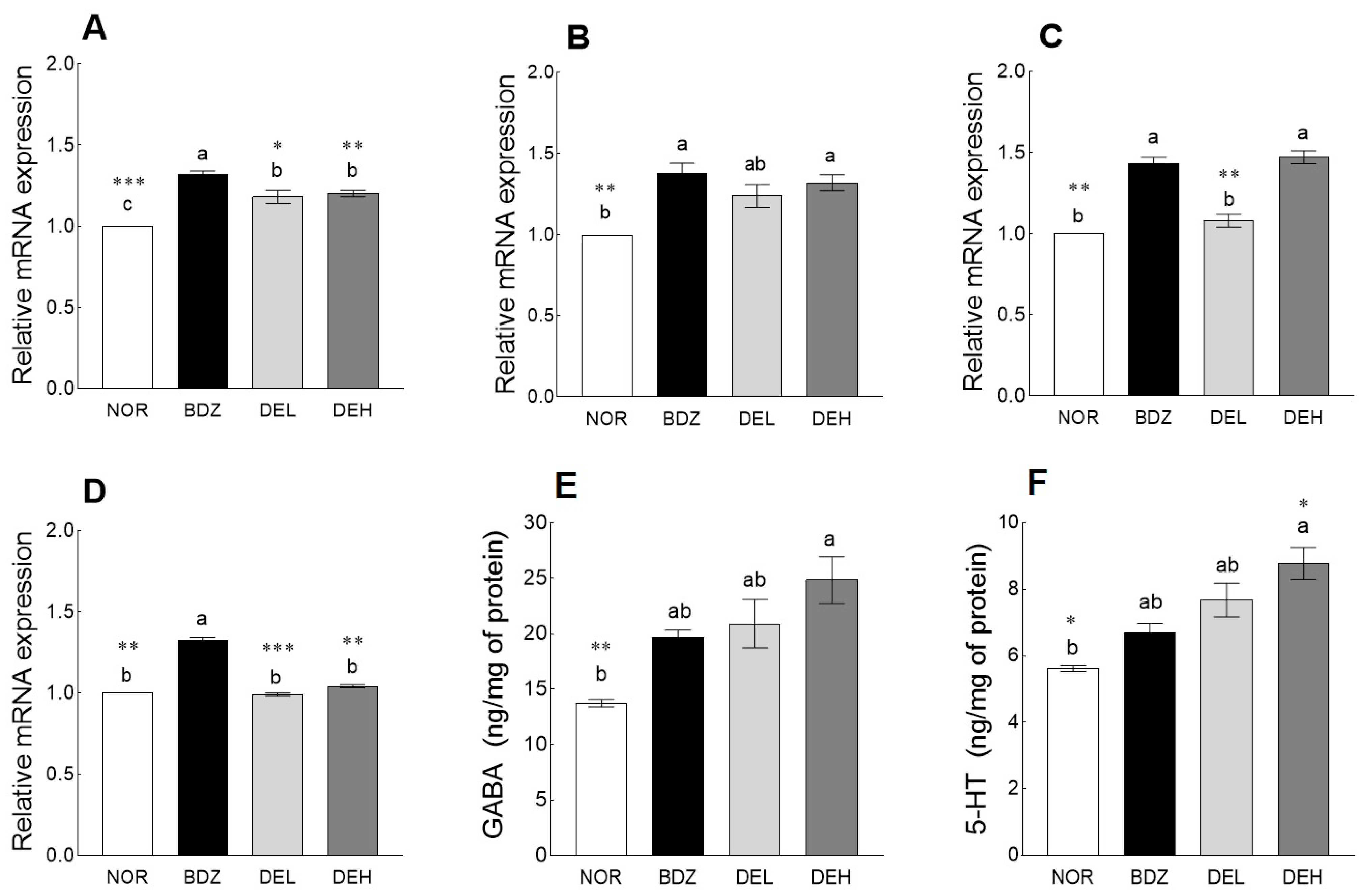
3.8. Effects of DE on mRNA Levels and Neurotransmitter Content in ICR Mice
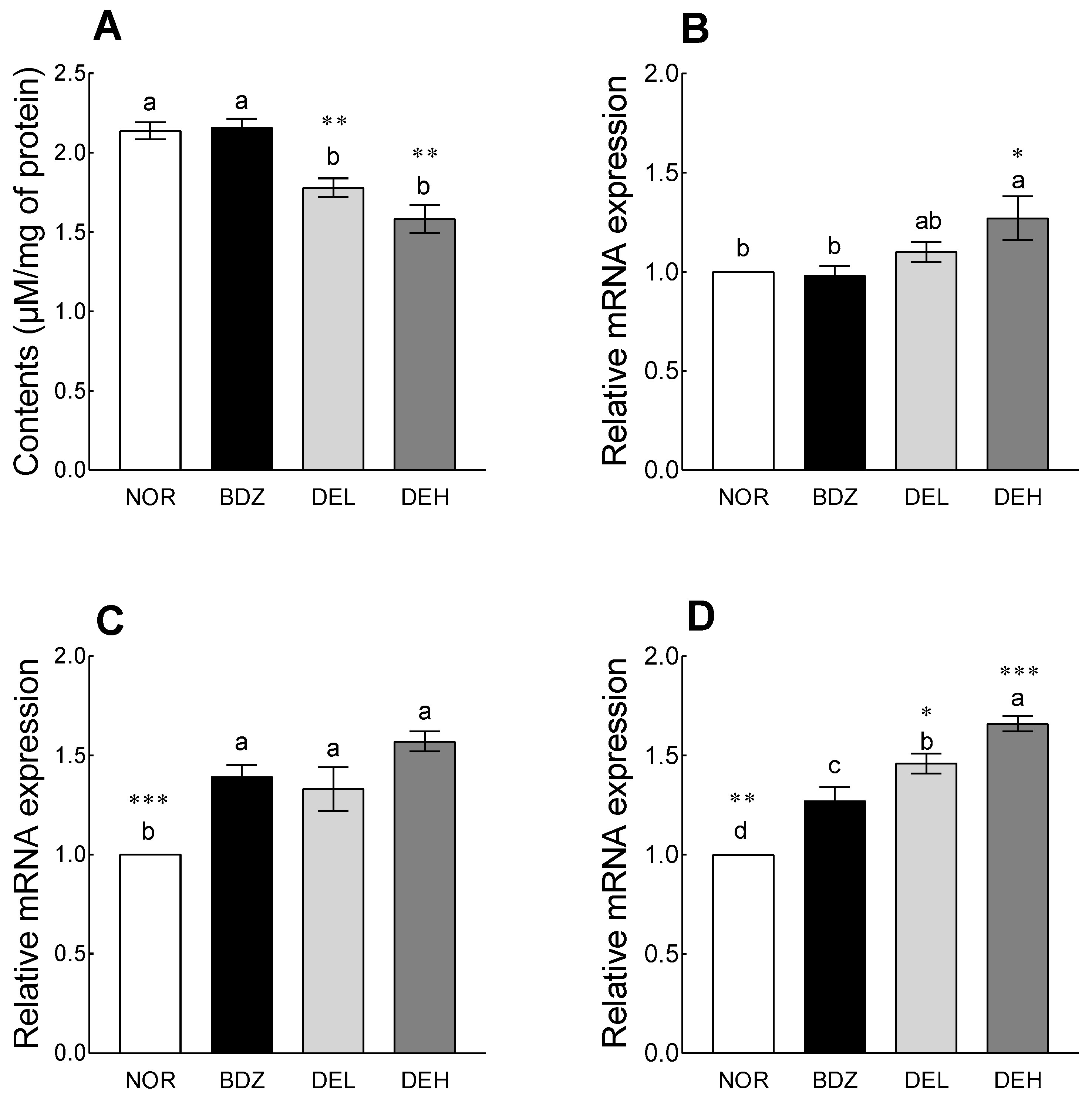
3.9. Effects of DE on MDA Content and Antioxidant Enzyme-Related mRNA Expression in ICR Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Cheng, L.; Liu, Y.; Wu, Z.; Weng, P. The intestinal microbiota links tea polyphenols with the regulation of mood and sleep to improve immunity. Food Rev. Int. 2023, 39, 1485–1498. [Google Scholar] [CrossRef]
- Tewari, D.; Sah, A.N.; Bawari, S.; Nabavi, S.F.; Dehpour, A.R.; Shirooie, S.; Braidy, N.; Fiebich, B.L.; Vacca, R.A.; Nabavi, S.M. Role of Nitric Oxide in Neurodegeneration: Function, Regulation, and Inhibition. Curr. Neuropharmacol. 2021, 19, 114–126. [Google Scholar] [CrossRef]
- Riemann, D.; Spiegelhalder, K.; Feige, B.; Voderholzer, U.; Berger, M.; Perlis, M.; Nissen, C. The hyperarousal model of insomnia: A review of the concept and its evidence. Sleep Med. Rev. 2010, 14, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Sher, P.K.; Study, R.E.; Mazzetta, J.; Barker, J.L.; Nelson, P.G. Depression of Benzodiazepine Binding and Diazepam Potentiation of Gaba-Mediated Inhibition after Chronic Exposure of Spinal-Cord Cultures to Diazepam. Brain Res. 1983, 268, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Bent, S.; Padula, A.; Moore, D.; Patterson, M.; Mehling, W. Valerian for sleep: A systematic review and meta-analysis. Am. J. Med. 2006, 119, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Unno, K.; Noda, S.; Kawasaki, Y.; Yamada, H.; Morita, A.; Iguchi, K.; Nakamura, Y. Reduced Stress and Improved Sleep Quality Caused by Green Tea Are Associated with a Reduced Caffeine Content. Nutrients 2017, 9, 777. [Google Scholar] [CrossRef]
- Park, C.W.; Hong, K.B.; Suh, H.J.; Ahn, Y. Sleep-promoting activity of amylase-treated Ashwagandha (Withania somnifera L. Dunal) root extract via GABA receptors. J. Food Drug Anal. 2023, 31, 278. [Google Scholar] [CrossRef]
- Um, M.Y.; Yoon, M.; Lee, J.; Jung, J.; Cho, S. A Novel Potent Sleep-Promoting Effect of Turmeric: Turmeric Increases Non-Rapid Eye Movement Sleep in Mice Via Histamine H1Receptor Blockade. Mol. Nutr. Food Res. 2021, 65, 2100100. [Google Scholar] [CrossRef]
- Ahn, Y.; Kim, S.; Park, C.; Kim, J.E.; Suh, H.J.; Jo, K. Sleep-promoting activity of lotus (Nelumbo nucifera) rhizome water extract via GABA(A) receptors. Pharm. Biol. 2022, 60, 1341–1348. [Google Scholar] [CrossRef]
- MacFarlane, J.; Morin, C.M.; Montplaisir, J. Hypnotics in Insomnia: The Experience of Zolpidem. Clin. Ther. 2014, 36, 1676–1701. [Google Scholar] [CrossRef]
- Everson, C.A.; Laatsch, C.D.; Hogg, N. Antioxidant defense responses to sleep loss and sleep recovery. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R374–R383. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, A.; Dor, Y.K.; Nambara, K.; Pollina, E.A.; Lin, C.D.; Greenberg, M.E.; Rogulja, D. Sleep Loss Can Cause Death through Accumulation of Reactive Oxygen Species in the Gut. Cell 2020, 181, 1307–1328.e15. [Google Scholar] [CrossRef]
- Balakrishnan, R.; Cho, D.Y.; Su-Kim, I.; Choi, D.K. Dendropanax morbiferus and Other Species from the Genus Dendropanax: Therapeutic Potential of Its Traditional Uses, Phytochemistry, and Pharmacology. Antioxidants 2020, 9, 962. [Google Scholar] [CrossRef]
- Jung, G.H.; Kim, S.H.; Woo, J.S.; Yoo, E.S.; Lee, J.H.; Han, S.H.; Jung, S.H.; Kim, S.M.; Kim, E.G.; Shin, M.J.; et al. Apoptosis Induction by Dendropanax morbiferus Leaves Extract in Human Breast Cancer Cells SK-BR-3 through MAPK Pathway. J. Life Sci. 2021, 31, 827–833. [Google Scholar]
- Kim, J.Y.; Yoon, J.Y.; Sugiura, Y.; Lee, S.K.; Park, J.D.; Song, G.J.; Yang, H.J. Dendropanax morbiferus leaf extract facilitates oligodendrocyte development. R. Soc. Open Sci. 2019, 6, 190266. [Google Scholar] [CrossRef] [PubMed]
- Hyun, T.K.; Ko, Y.J.; Kim, E.H.; Chung, I.M.; Kim, J.S. Anti-inflammatory activity and phenolic composition of Dendropanax morbifera leaf extracts. Ind. Crops Prod. 2015, 74, 263–270. [Google Scholar] [CrossRef]
- Park, S.Y.; Karthivashan, G.; Ko, H.M.; Cho, D.Y.; Kim, J.; Cho, D.J.; Ganesan, P.; Su-Kim, I.; Choi, D.K. Aqueous Extract of Dendropanax morbiferus Leaves Effectively Alleviated Neuroinflammation and Behavioral Impediments in MPTP-Induced Parkinson’s Mouse Model. Oxid. Med. Cell. Longev. 2018, 2018, 3175214. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J.; Kwon, E.B.; Ryu, H.W.; Lee, S.; Lee, J.W.; Kim, D.Y.; Lee, M.K.; Oh, S.R.; Lee, H.S.; Lee, S.U.; et al. Polyacetylene From Dendropanax morbifera Alleviates Diet-Induced Obesity and Hepatic Steatosis by Activating AMPK Signaling Pathway. Front. Pharmacol. 2018, 9, 537. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef]
- Ispiryan, A.; Viskelis, J.; Viskelis, P.; Urbonaviciene, D.; Raudone, L. Biochemical and Antioxidant Profiling of Raspberry Plant Parts for Sustainable Processing. Plants 2023, 12, 2424. [Google Scholar] [CrossRef]
- Han, S.H.; Ahn, Y.; Lee, H.J.; Suh, H.J.; Jo, K. Antioxidant and Immunostimulatory Activities of a Submerged Culture of Cordyceps sinensis Using Spent Coffee. Foods 2021, 10, 1697. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.; Han, S.H.; Kim, M.G.; Hong, K.B.; Kim, W.J.; Suh, H.J.; Jo, K. Anti-depressant effects of ethanol extract from Cannabis sativa (hemp) seed in chlorpromazine-induced Drosophila melanogaster depression model. Pharm. Biol. 2021, 59, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Jeong, A.H.; Hwang, J.; Jo, K.; Kim, S.; Ahn, Y.; Suh, H.J.; Choi, H.S. Fermented Gamma Aminobutyric Acid Improves Sleep Behaviors in Fruit Flies and Rodent Models. Int. J. Mol. Sci. 2021, 22, 3537. [Google Scholar] [CrossRef]
- Fernandez-Mendoza, J.; Vgontzas, A.N. Insomnia and its impact on physical and mental health. Curr. Psychiatry Rep. 2013, 15, 418. [Google Scholar] [CrossRef] [PubMed]
- Beltran Gonzalez, A.N.; Lopez Pazos, M.I.; Calvo, D.J. Reactive Oxygen Species in the Regulation of the GABA Mediated Inhibitory Neurotransmission. Neuroscience 2020, 439, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Grabska-Kobylecka, I.; Kaczmarek-Bak, J.; Figlus, M.; Prymont-Przyminska, A.; Zwolinska, A.; Sarniak, A.; Wlodarczyk, A.; Glabinski, A.; Nowak, D. The Presence of Caffeic Acid in Cerebrospinal Fluid: Evidence That Dietary Polyphenols Can Cross the Blood-Brain Barrier in Humans. Nutrients 2020, 12, 1531. [Google Scholar] [CrossRef]
- Eom, T.; Kim, K.C.; Kim, J.-S. Dendropanax morbifera leaf polyphenolic compounds: Optimal extraction using the response surface method and their protective effects against alcohol-induced liver damage. Antioxidants 2020, 9, 120. [Google Scholar] [CrossRef]
- Javed, H.; Khan, M.M.; Ahmad, A.; Vaibhav, K.; Ahmad, M.E.; Khan, A.; Ashafaq, M.; Islam, F.; Siddiqui, M.S.; Safhi, M.M.; et al. Rutin prevents cognitive impairments by ameliorating oxidative stress and neuroinflammation in rat model of sporadic dementia of Alzheimer type. Neuroscience 2012, 210, 340–352. [Google Scholar] [CrossRef]
- Lee, T.K.; Kang, I.J.; Kim, B.; Sim, H.J.; Kim, D.W.; Ahn, J.H.; Lee, J.C.; Ryoo, S.; Shin, M.C.; Cho, J.H.; et al. Experimental Pretreatment with Chlorogenic Acid Prevents Transient Ischemia-Induced Cognitive Decline and Neuronal Damage in the Hippocampus through Anti-Oxidative and Anti-Inflammatory Effects. Molecules 2020, 25, 3578. [Google Scholar] [CrossRef]
- Hermawati, E.; Arfian, N.; Mustofa, M.; Partadiredja, G. Chlorogenic acid ameliorates memory loss and hippocampal cell death after transient global ischemia. Eur. J. Neurosci. 2020, 51, 651–669. [Google Scholar] [CrossRef]
- Ki, Y.; Lim, C. Sleep-promoting effects of threonine link amino acid metabolism in Drosophila neuron to GABAergic control of sleep drive. Elife 2019, 8, e40593. [Google Scholar] [CrossRef]
- Catterson, J.H.; Knowles-Barley, S.; James, K.; Heck, M.M.; Harmar, A.J.; Hartley, P.S. Dietary modulation of Drosophila sleep-wake behaviour. PLoS ONE 2010, 5, e12062. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; Ahn, Y.; Cho, H.J.; Kwak, W.K.; Jo, K.; Suh, H.J. Chemical compositions and sleep-promoting activities of hop (Humulus lupulus L.) varieties. J. Food Sci. 2023, 88, 2217–2228. [Google Scholar] [CrossRef]
- Ly, S.; Pack, A.I.; Naidoo, N. The neurobiological basis of sleep: Insights from Drosophila. Neurosci. Biobehav. Rev. 2018, 87, 67–86. [Google Scholar] [CrossRef]
- Yuan, Q.; Joiner, W.J.; Sehgal, A. A sleep-promoting role for the Drosophila serotonin receptor 1A. Curr. Biol. 2006, 16, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.N.; Ho, K.; Crocker, A.; Yue, Z.; Koh, K.; Sehgal, A. The effects of caffeine on sleep in Drosophila require PKA activity, but not the adenosine receptor. J. Neurosci. 2009, 29, 11029–11037. [Google Scholar] [CrossRef] [PubMed]
- Inoue, E.; Suzuki, T.; Nakayama, T.; Yoshimura, T.; Sudo, K.; Shimizu, Y.; Iwaki, Y.; Kawasaki, H.; Ishida, N. Novel hypnotics of Japanese traditional herbal medicines to caffeine-induced insomnia in Drosophila by using Newly-developed automated sleep and rhythm analysis system (AutoCircaS). Gene 2022, 846, 146852. [Google Scholar] [CrossRef]
- Yi, P.L.; Tsai, C.H.; Chen, Y.C.; Chang, F.C. Gamma-aminobutyric acid (GABA) receptor mediates suanzaorentang, a traditional Chinese herb remedy, -induced sleep alteration. J. Biomed. Sci. 2007, 14, 285–297. [Google Scholar] [CrossRef]
- Hong, K.B.; Park, Y.; Suh, H.J. Two combined amino acids promote sleep activity in caffeine-induced sleepless model systems. Nutr. Res. Pract. 2018, 12, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Asbah, A.; Ummussaadah, U.; Parenden, N.; Putri, A.S.W.; Rosa, R.A.; Rumata, N.R.; Emran, T.B.; Dhama, K.; Nainu, F. Pharmacological Effect of Caffeine on Drosophila melanogaster: A Proof-of-Concept in vivo Study for Nootropic Investigation. Arch. Razi Inst. 2021, 76, 1645–1654. [Google Scholar] [CrossRef]
- Liang, Y.; Huang, R.; Chen, Y.; Zhong, J.; Deng, J.; Wang, Z.; Wu, Z.; Li, M.; Wang, H.; Sun, Y. Study on the Sleep-Improvement Effects of Hemerocallis citrina Baroni in Drosophila melanogaster and Targeted Screening to Identify Its Active Components and Mechanism. Foods 2021, 10, 883. [Google Scholar] [CrossRef]
- Fernández, S.P.; Wasowski, C.; Loscalzo, L.M.; Granger, R.E.; Johnston, G.A.; Paladini, A.C.; Marder, M. Central nervous system depressant action of flavonoid glycosides. Eur. J. Pharmacol. 2006, 539, 168–176. [Google Scholar] [CrossRef]
- Park, I.; Ochiai, R.; Ogata, H.; Kayaba, M.; Hari, S.; Hibi, M.; Katsuragi, Y.; Satoh, M.; Tokuyama, K. Effects of subacute ingestion of chlorogenic acids on sleep architecture and energy metabolism through activity of the autonomic nervous system: A randomised, placebo-controlled, double-blinded cross-over trial. Br. J. Nutr. 2017, 117, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Su, X.; Kang, Y.; Si, J.; Wang, L.; Li, X.; Ma, K. Effect and mechanism of chlorogenic acid on cognitive dysfunction in mice by lipopolysaccharide-induced neuroinflammation. Front. Immunol. 2023, 14, 1178188. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, L.; Chen, B.; Fang, Y.; Lin, W.; Zhang, T.; Feng, X.; Tao, X.; Wu, Y.; Fu, X.; et al. Chlorogenic acid exerts neuroprotective effect against hypoxia-ischemia brain injury in neonatal rats by activating Sirt1 to regulate the Nrf2-NF-κB signaling pathway. Cell Commun. Signal. 2022, 20, 84. [Google Scholar] [CrossRef]
- Hibi, M. Potential of Polyphenols for Improving Sleep: A Preliminary Results from Review of Human Clinical Trials and Mechanistic Insights. Nutrients 2023, 15, 1257. [Google Scholar] [CrossRef]
- Rebas, E.; Rzajew, J.; Radzik, T.; Zylinska, L. Neuroprotective Polyphenols: A Modulatory Action on Neurotransmitter Pathways. Curr. Neuropharmacol. 2020, 18, 431–445. [Google Scholar] [CrossRef]
- Hernandez-Leon, A.; González-Trujano, M.E.; Fernández-Guasti, A. The anxiolytic-like effect of rutin in rats involves GABAA receptors in the basolateral amygdala. Behav. Pharmacol. 2017, 28, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Hernández, E.; González-Trujano, M.E.; Terrazas, T.; Herrera Santoyo, J.; Guevara-Fefer, P. Anxiolytic and sedative-like effects of flavonoids from Tilia americana var. mexicana: GABAergic and serotonergic participation. Salud Ment. 2016, 39, 37–46. [Google Scholar] [CrossRef]
- Foudah, A.I.; Alqarni, M.H.; Alam, A.; Devi, S.; Salkini, M.A.; Alam, P. Rutin Improves Anxiety and Reserpine-Induced Depression in Rats. Molecules 2022, 27, 7313. [Google Scholar] [CrossRef] [PubMed]
- Yaşar, S.N.; Can, O.D.; Oztürk, N.; Sagratini, G.; Ricciutelli, M.; Vittori, S.; Maggi, F. Central nervous system activities of Hypericum origanifolium extract via GABAergic and opioidergic mechanisms. Phytother. Res. 2013, 27, 877–884. [Google Scholar] [CrossRef]
- Shinomiya, K.; Omichi, J.; Ohnishi, R.; Ito, H.; Yoshida, T.; Kamei, C. Effects of chlorogenic acid and its metabolites on the sleep-wakefulness cycle in rats. Eur. J. Pharmacol. 2004, 504, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, R.; Tomonobu, K.; Ikushima, I. Effect of chlorogenic acids on fatigue and sleep in healthy males: A randomized, double-blind, placebo-controlled, crossover study. Food Sci. Nutr. 2018, 6, 2530–2536. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, W.; Li, Q.; Li, Y.; Yan, Y.; Huang, F.; Wu, X.; Zhou, Q.; Shu, X.; Ruan, Z. Dietary chlorogenic acid regulates gut microbiota, serum-free amino acids and colonic serotonin levels in growing pigs. Int. J. Food Sci. Nutr. 2018, 69, 566–573. [Google Scholar] [CrossRef]
- Wu, J.; Chen, H.; Li, H.; Tang, Y.; Yang, L.; Cao, S.; Qin, D. Antidepressant Potential of Chlorogenic Acid-Enriched Extract from Eucommia ulmoides Oliver Bark with Neuron Protection and Promotion of Serotonin Release through Enhancing Synapsin I Expression. Molecules 2016, 21, 260. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Liu, C.; Bao, H.; Li, S.; Huang, Z.; Gu, D.; Xiong, L.; Miao, L. Estimation of gingival crevicular fluid oxidative stress markers in school-aged children and teenagers with insufficient sleep. BMC Oral Health 2022, 22, 616. [Google Scholar] [CrossRef] [PubMed]
- D’Almeida, V.; Lobo, L.L.; Hipólide, D.C.; de Oliveira, A.C.; Nobrega, J.N.; Tufik, S. Sleep deprivation induces brain region-specific decreases in glutathione levels. Neuroreport 1998, 9, 2853–2856. [Google Scholar] [CrossRef]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a Potent Antioxidant: Implications for Neurodegenerative Disorders. Oxid. Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef]
- Zeng, L.; Xiang, R.; Fu, C.; Qu, Z.; Liu, C. The Regulatory effect of chlorogenic acid on gut-brain function and its mechanism: A systematic review. Biomed. Pharmacother. 2022, 149, 112831. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ABTS (IC50, mg/mL) | DPPH (IC50, mg/mL) | |
|---|---|---|
| D. morbiferus extract | 0.70 ± 0.02 | 0.32 ± 0.02 |
| Rutin | 0.23 ± 0.01 | 0.11 ± 0.01 |
| Chlorogenic acid | 0.13 ± 0.00 | 0.08 ± 0.00 |
| Compound | Gallic Acid | 3,4-Dihydroxybenzoic Acid | Rutin | Chlorogenic Acid | Caffeic Acid | p-Coumaric Acid | Trans Ferulic Acid | Quercetin | Total Polyphenol |
|---|---|---|---|---|---|---|---|---|---|
| Contents (μg/mg) | 0.15 ± 0.00 | 2.64 ± 0.00 | 35.78 ± 0.01 | 59.22 ± 0.02 | 0.43 ± 0.01 | 0.36 ± 0.00 | 0.12 ± 0.00 | 0.17 ± 0.00 | 98.88 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, K.; Ahn, Y.; Cheon, G.Y.; Suh, H.J.; Cho, Y.J.; Park, S.-S.; Hong, K.-B. Effects of Dendropanax morbiferus Leaf Extract on Sleep Parameters in Invertebrate and Vertebrate Models. Antioxidants 2023, 12, 1890. https://doi.org/10.3390/antiox12101890
Ko K, Ahn Y, Cheon GY, Suh HJ, Cho YJ, Park S-S, Hong K-B. Effects of Dendropanax morbiferus Leaf Extract on Sleep Parameters in Invertebrate and Vertebrate Models. Antioxidants. 2023; 12(10):1890. https://doi.org/10.3390/antiox12101890
Chicago/Turabian StyleKo, Kayoung, Yejin Ahn, Ga Yeong Cheon, Hyung Joo Suh, Yun Jae Cho, Sung-Soo Park, and Ki-Bae Hong. 2023. "Effects of Dendropanax morbiferus Leaf Extract on Sleep Parameters in Invertebrate and Vertebrate Models" Antioxidants 12, no. 10: 1890. https://doi.org/10.3390/antiox12101890
APA StyleKo, K., Ahn, Y., Cheon, G. Y., Suh, H. J., Cho, Y. J., Park, S.-S., & Hong, K.-B. (2023). Effects of Dendropanax morbiferus Leaf Extract on Sleep Parameters in Invertebrate and Vertebrate Models. Antioxidants, 12(10), 1890. https://doi.org/10.3390/antiox12101890