


Smoking Induces a Decline in Semen Quality and the Activation of Stress Response Pathways in Sperm

 ,
,  ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Semen Sample Collection, Analysis, and Protein Extraction
2.3. Slot Blot Analysis
2.4. Statistical Analysis
3. Results
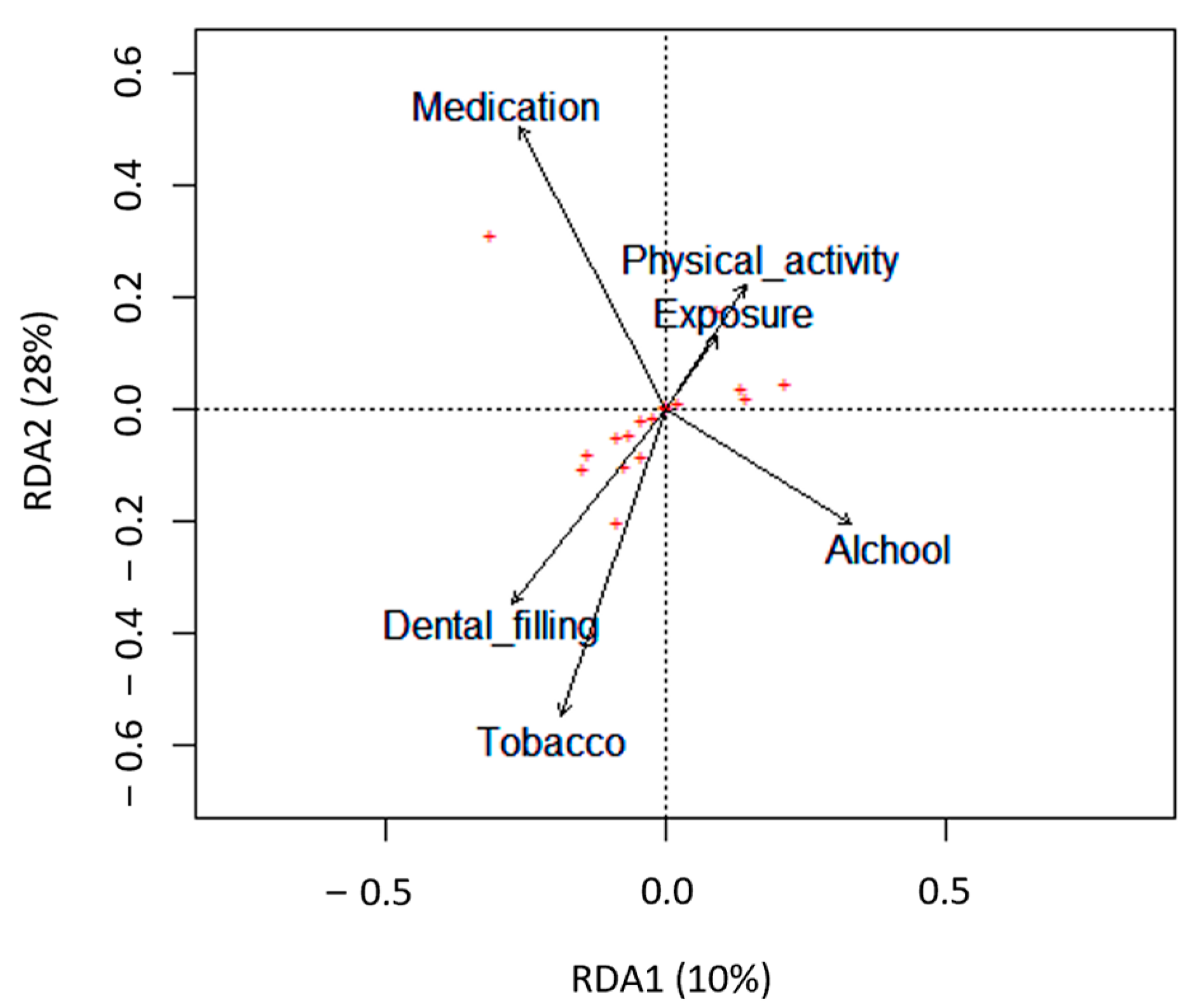
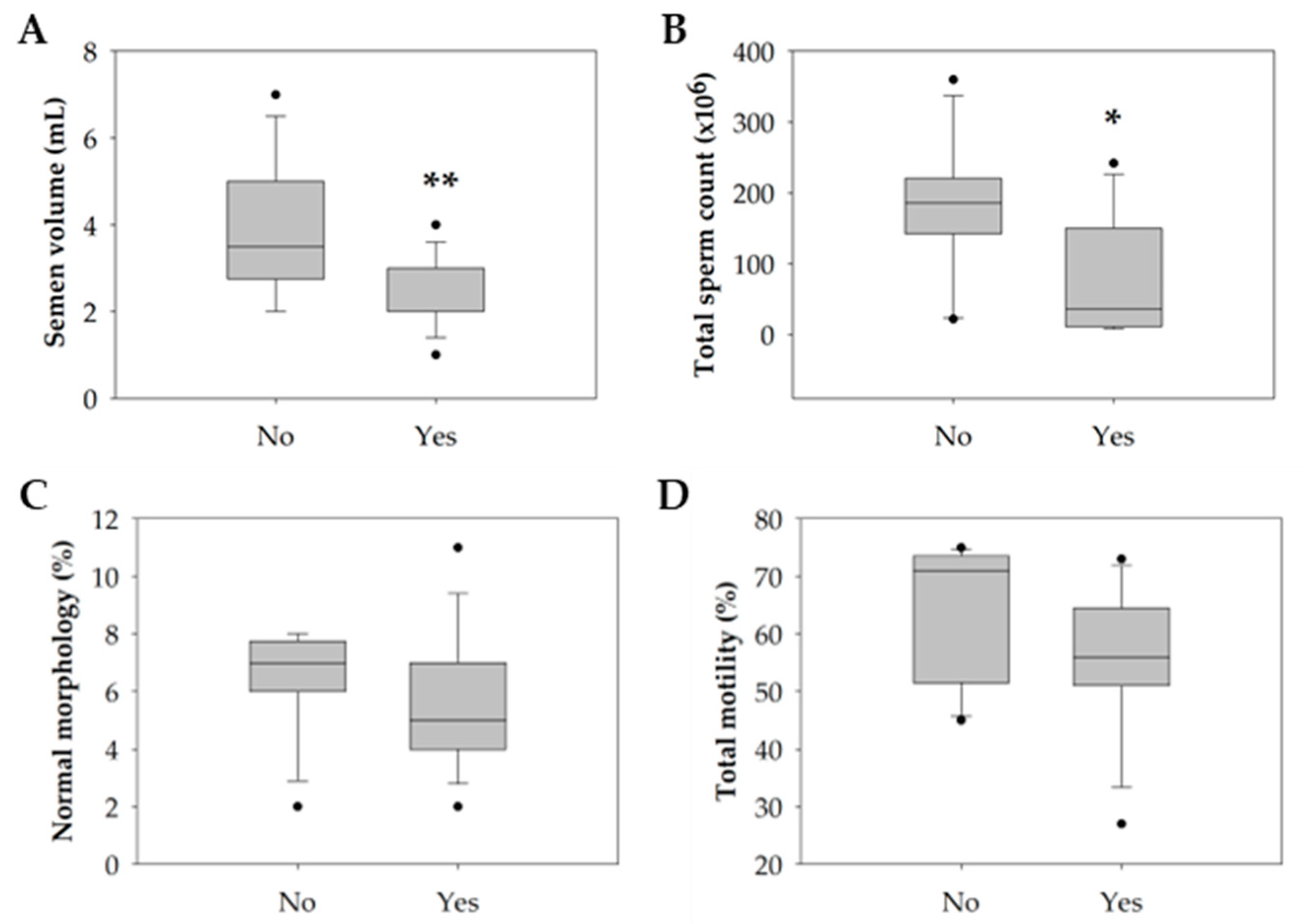
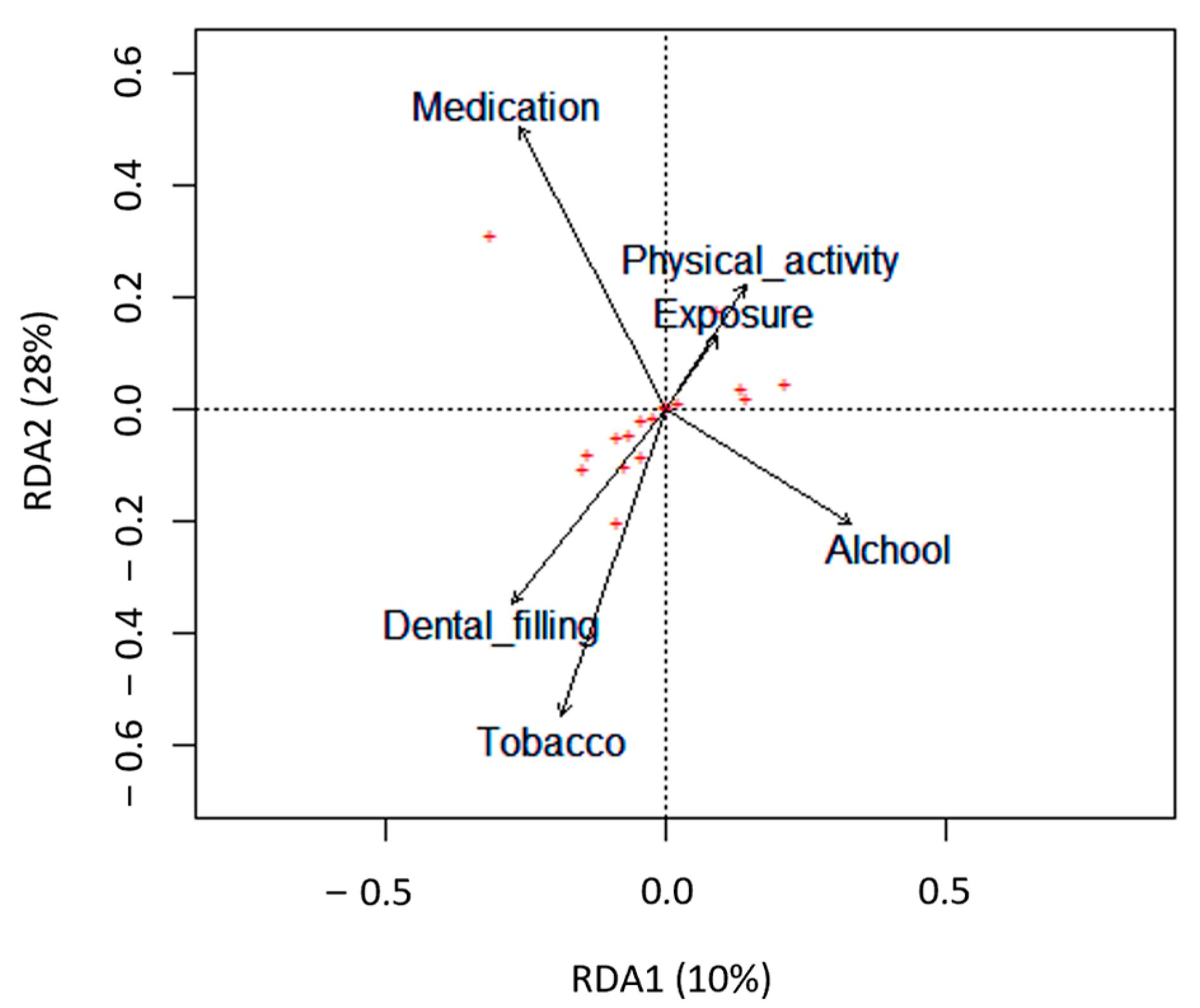
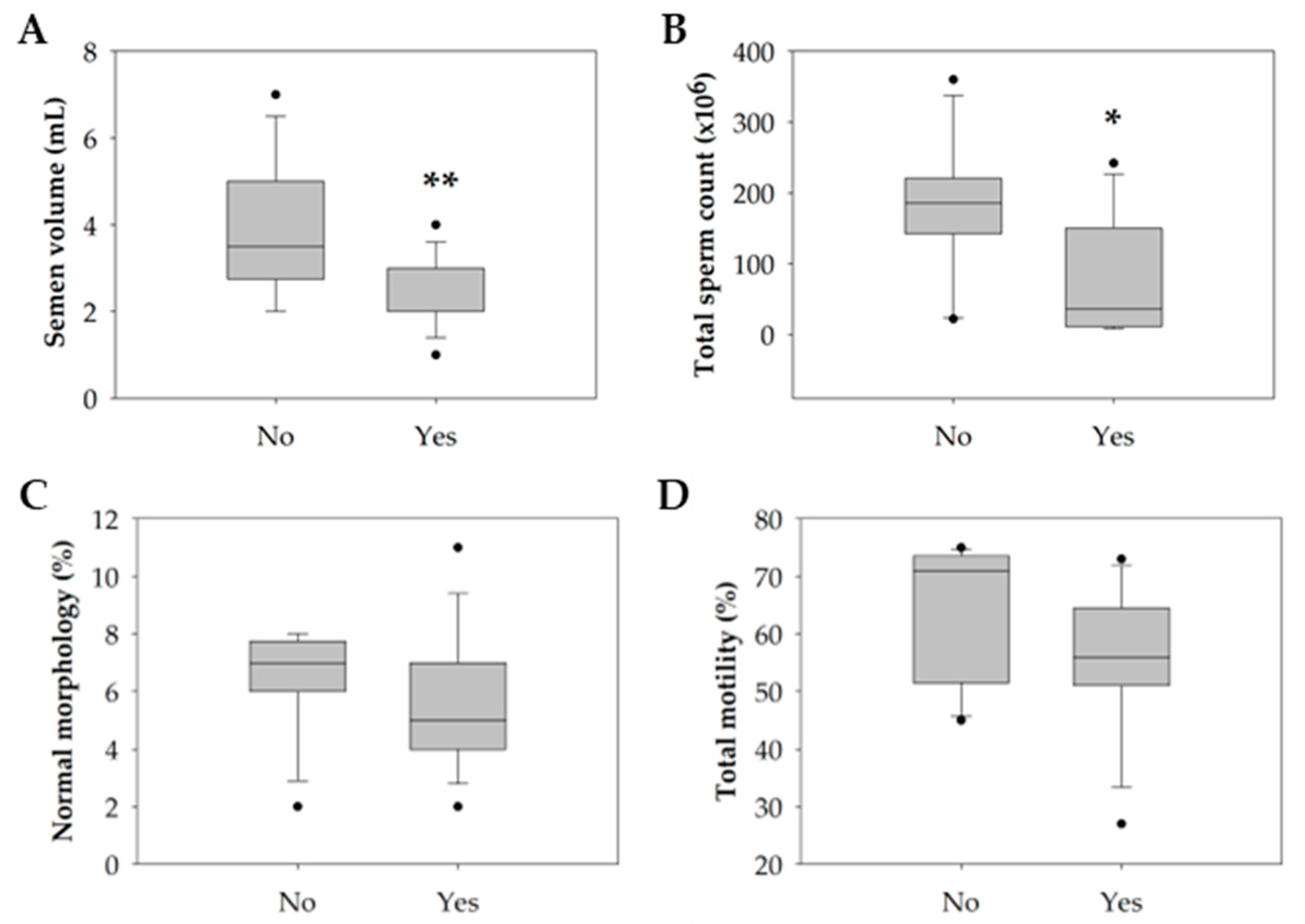
3.1. Impact of Lifestyle on the Conventional Semen Quality Parameters
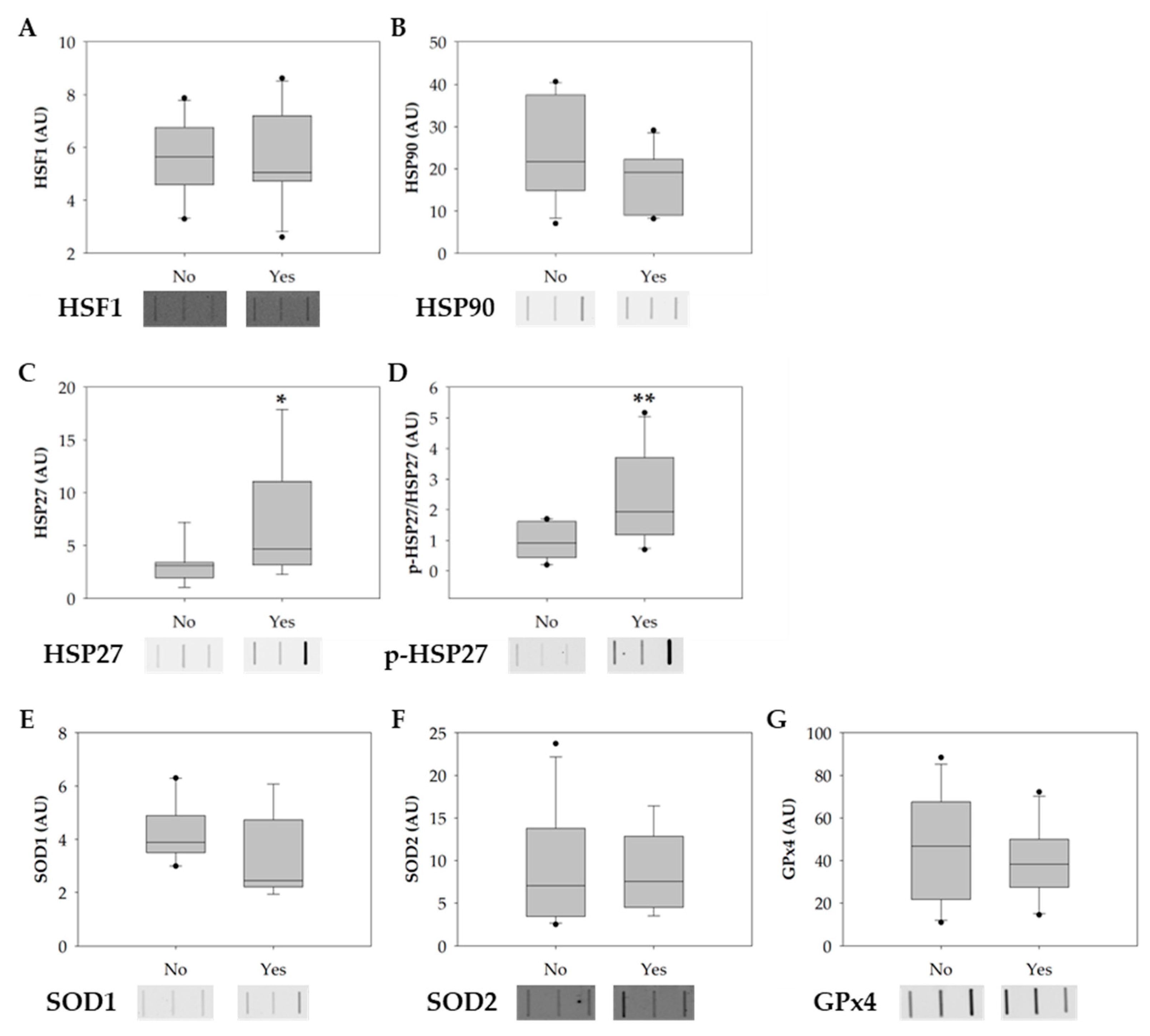
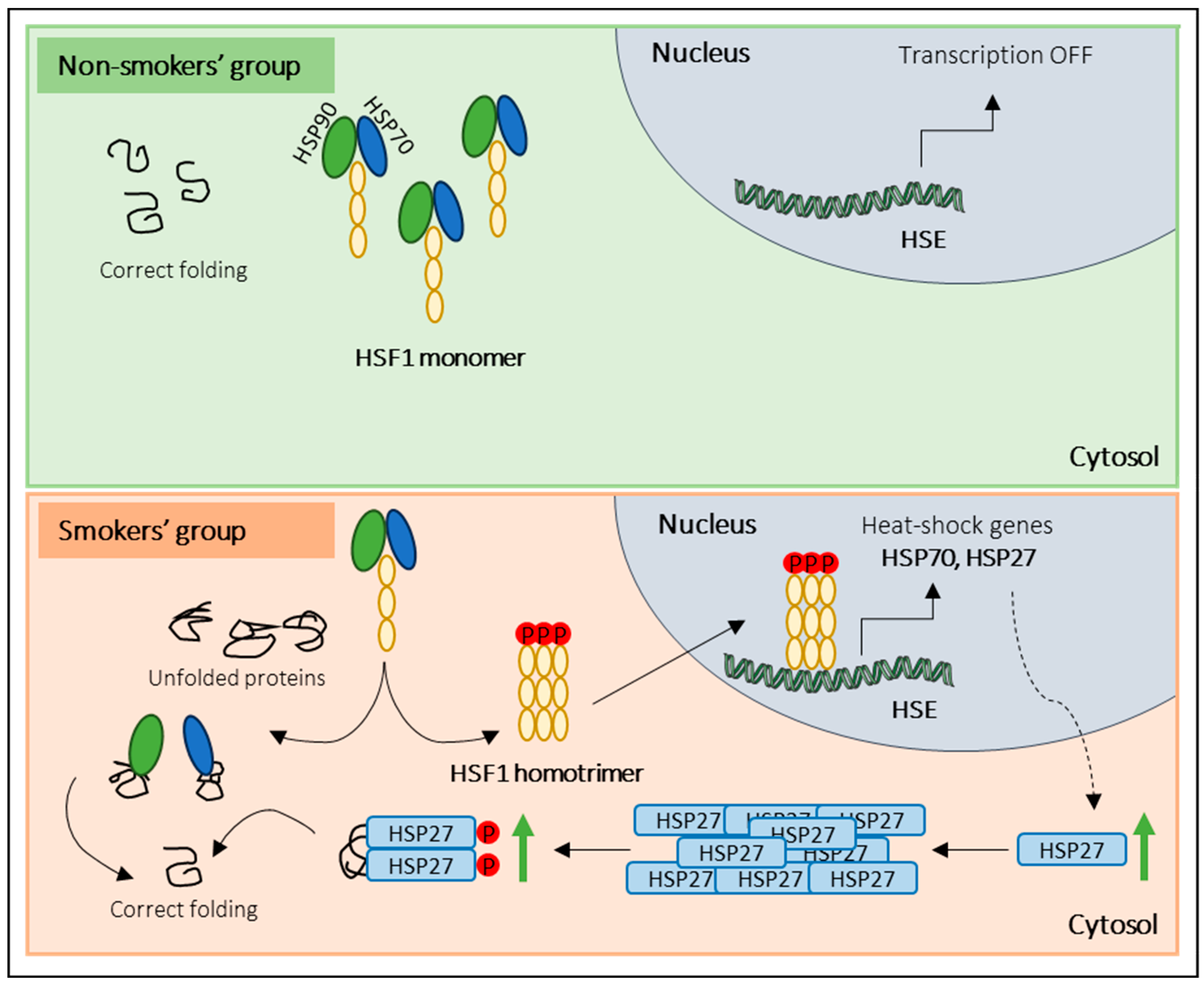
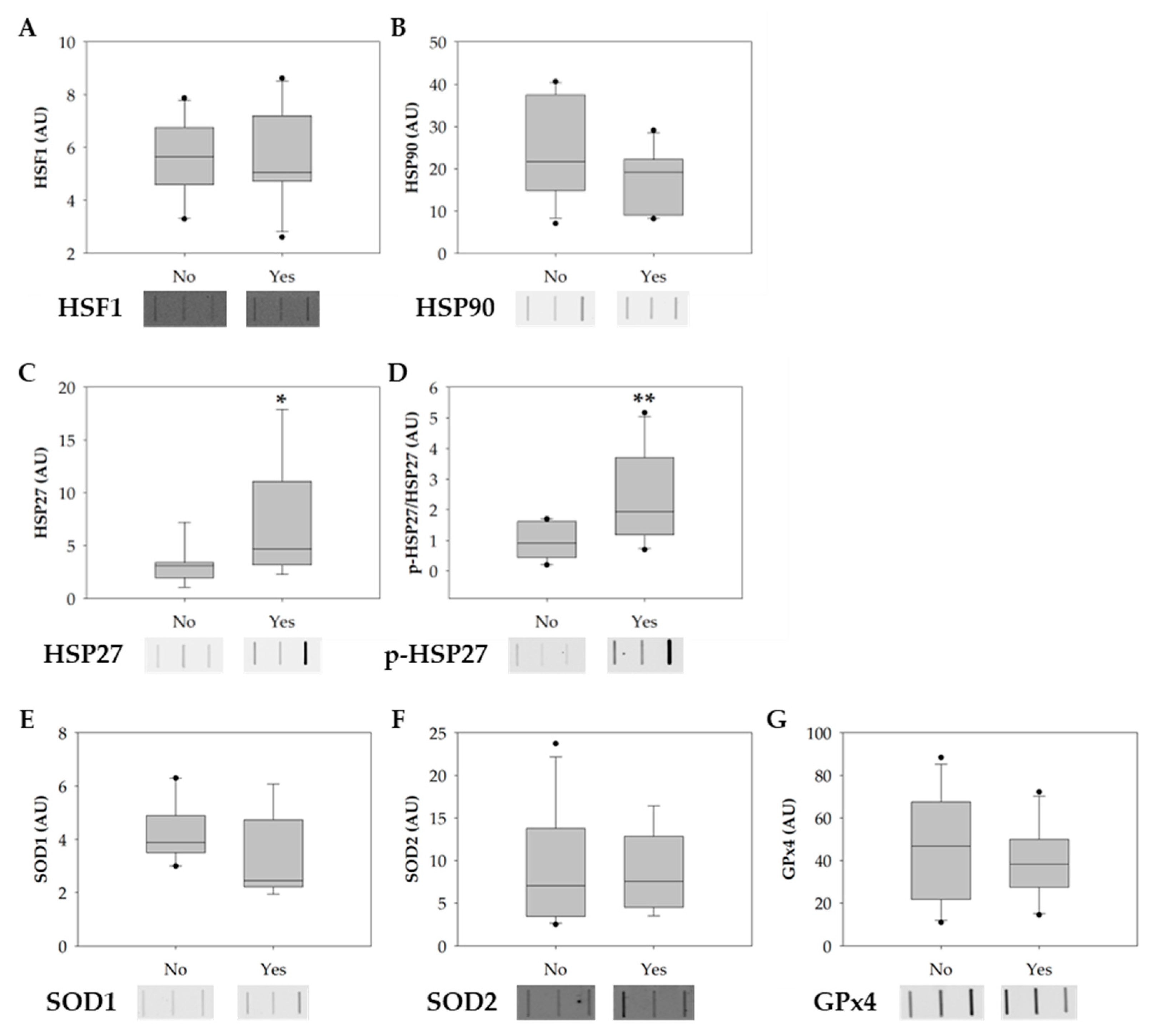
3.2. Smoking Increases the Phosphorylation of HSP27
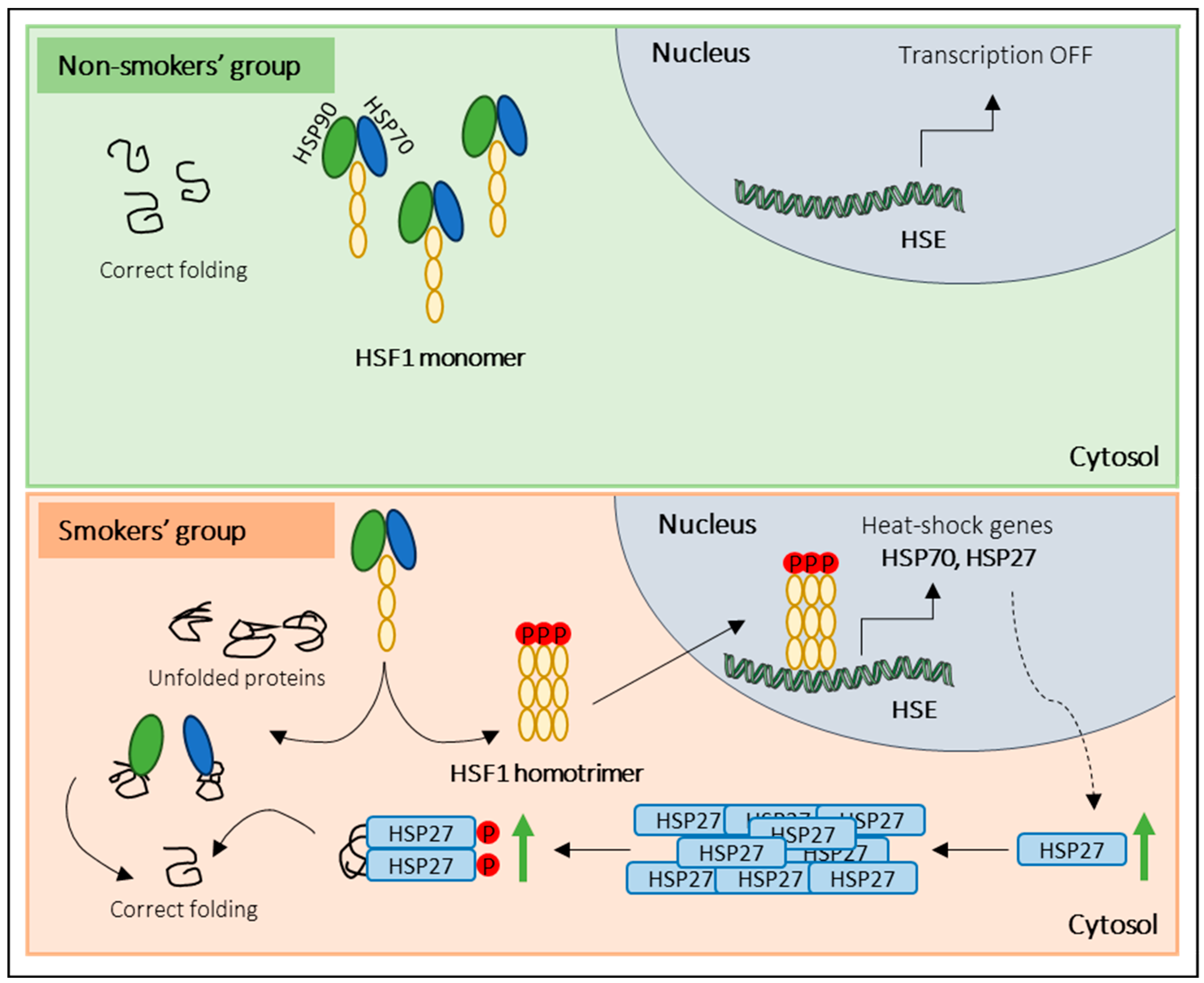
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vander Borght, M.; Wyns, C. Fertility and Infertility: Definition and Epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Infertility Prevalence Estimates, 1990–2021; World Health Organization: Geneva, Switzerland, 2023. [Google Scholar]
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.-L.L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male Infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Esteves, S.C.; Humaidan, P. Towards Infertility Care on Equal Terms: A Prime Time for Male Infertility. Reprod. Biomed. Online 2023, 47, 11–14. [Google Scholar] [CrossRef]
- Henriques, M.C.; Loureiro, S.; Fardilha, M.; Herdeiro, M.T. Exposure to Mercury and Human Reproductive Health: A Systematic Review. Reprod. Toxicol. 2019, 85, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.; Silva, J.V.; Santos, M.A.S.; Fardilha, M. Fighting Bisphenol A-Induced Male Infertility: The Power of Antioxidants. Antioxidants 2021, 10, 289. [Google Scholar] [CrossRef]
- Silva, J.V.; Santiago, J.; Matos, B.; Henriques, M.C.; Patrício, D.; Martins, A.D.; Duarte, J.A.; Ferreira, R.; Alves, M.G.; Oliveira, P.; et al. Effects of Age and Lifelong Moderate-Intensity Exercise Training on Rats’ Testicular Function. Int. J. Mol. Sci. 2022, 23, 11619. [Google Scholar] [CrossRef]
- Chen, T.; Belladelli, F.; Del Giudice, F.; Eisenberg, M.L. Male Fertility as a Marker for Health. Reprod. Biomed. Online 2022, 44, 131–144. [Google Scholar] [CrossRef]
- Nwonuma, C.O.; Osemwegie, O.O.; Irokanulo, E.O.; Alejolowo, O.O.; Kayode, O.T.; Olaolu, T.D.; Ada, A.S.; Rotimi, D.E.; Maimako, R.F.; Adedayo, A.S.; et al. Comparative Effects of Combined Use of Alcohol with Cannabis and Tobacco on Testicular Function in Rats. Toxicol. Res. 2021, 10, 761–770. [Google Scholar] [CrossRef]
- Vivarelli, F.; Canistro, D.; Cirillo, S.; Cardenia, V.; Rodriguez-Estrada, M.T.; Paolini, M. Impairment of Testicular Function in Electronic Cigarette (e-Cig, e-Cigs) Exposed Rats under Low-Voltage and Nicotine-Free Conditions. Life Sci. 2019, 228, 53–65. [Google Scholar] [CrossRef]
- Qi, X.; Guan, Q.; Zhang, W.; Huang, X.; Yu, C. The Time-Dependent Adverse Effects of a High-Fat Diet on Sperm Parameters. Adv. Clin. Exp. Med. 2023, 32, 889–900. [Google Scholar] [CrossRef]
- Leisegang, K.; Dutta, S. Do Lifestyle Practices Impede Male Fertility? Andrologia 2021, 53, e13595. [Google Scholar] [CrossRef] [PubMed]
- Saygın, H.; Korgalı, E.; Koç, T.; Doğan, K. The Effect of Smoking and Electronic Cigarettes on Rat Testicles. Rev. Int. Andrología 2023, 21, 100365. [Google Scholar] [CrossRef] [PubMed]
- Budin, S.B.; Kho, J.H.; Lee, J.H.; Ramalingam, A.; Jubaidi, F.F.; Latif, E.S.; Zainalabidin, S.; Taib, I.S.; Mohamed, J. Low-Dose Nicotine Exposure Induced the Oxidative Damage of Reproductive Organs and Altered the Sperm Characteristics of Adolescent Male Rats. Malays. J. Med. Sci. 2017, 24, 50–57. [Google Scholar] [CrossRef]
- Amor, H.; Hammadeh, M.E.; Mohd, I.; Jankowski, P.M. Impact of Heavy Alcohol Consumption and Cigarette Smoking on Sperm DNA Integrity. Andrologia 2022, 54, e14434. [Google Scholar] [CrossRef] [PubMed]
- Amor, H.; Zeyad, A.; Hammadeh, M.E. Tobacco Smoking and Its Impact on the Expression Level of Sperm Nuclear Protein Genes: H2BFWT, TNP1, TNP2, PRM1 and PRM2. Andrologia 2021, 53, e13964. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.-B.; Wang, Z.-X.; Qiao, Z.-D. The Hazardous Effects of Tobacco Smoking on Male Fertility. Asian J. Androl. 2015, 17, 954. [Google Scholar] [CrossRef] [PubMed]
- Bisconti, M.; Simon, J.-F.; Grassi, S.; Leroy, B.; Martinet, B.; Arcolia, V.; Isachenko, V.; Hennebert, E. Influence of Risk Factors for Male Infertility on Sperm Protein Composition. Int. J. Mol. Sci. 2021, 22, 13164. [Google Scholar] [CrossRef]
- Antoniassi, M.P.; Intasqui, P.; Camargo, M.; Zylbersztejn, D.S.; Carvalho, V.M.; Cardozo, K.H.M.; Bertolla, R.P. Analysis of the Functional Aspects and Seminal Plasma Proteomic Profile of Sperm from Smokers. BJU Int. 2016, 118, 814–822. [Google Scholar] [CrossRef]
- Saito, A.; Imaizumi, K. Unfolded Protein Response-Dependent Communication and Contact among Endoplasmic Reticulum, Mitochondria, and Plasma Membrane. Int. J. Mol. Sci. 2018, 19, 3215. [Google Scholar] [CrossRef]
- Doultsinos, D.; Avril, T.; Lhomond, S.; Dejeans, N.; Guédat, P.; Chevet, E. Control of the Unfolded Protein Response in Health and Disease. SLAS Discov. 2017, 22, 787–800. [Google Scholar] [CrossRef]
- Lindholm, D.; Korhonen, L.; Eriksson, O.; Kõks, S. Recent Insights into the Role of Unfolded Protein Response in ER Stress in Health and Disease. Front. Cell Dev. Biol. 2017, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, M.W.; Nargund, A.M.; Haynes, C.M. Signaling the Mitochondrial Unfolded Protein Response. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2013, 1833, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Almanza, A.; Carlesso, A.; Chintha, C.; Creedican, S.; Doultsinos, D.; Leuzzi, B.; Luís, A.; McCarthy, N.; Montibeller, L.; More, S.; et al. Endoplasmic Reticulum Stress Signalling—From Basic Mechanisms to Clinical Applications. FEBS J. 2019, 286, 241–278. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.; Silva, J.V.; Fardilha, M. First Insights on the Presence of the Unfolded Protein Response in Human Spermatozoa. Int. J. Mol. Sci. 2019, 20, 5518. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Wan, L. Associations between Smoking Status and Infertility: A Cross-Sectional Analysis among USA Women Aged 18-45 Years. Front. Endocrinol. 2023, 14, 1140739. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Rosa-Villagrán, L.; Barrera, N.; Montes, J.; Riso, C.; Sapiro, R. Decline of Semen Quality over the Last 30 Years in Uruguay. Basic Clin. Androl. 2021, 31, 8. [Google Scholar] [CrossRef]
- Mishra, P.; Negi, M.P.S.; Srivastava, M.; Singh, K.; Rajender, S. Decline in Seminal Quality in Indian Men over the Last 37 Years. Reprod. Biol. Endocrinol. 2018, 16, 103. [Google Scholar] [CrossRef]
- Durairajanayagam, D. Lifestyle Causes of Male Infertility. Arab J. Urol. 2018, 16, 10–20. [Google Scholar] [CrossRef]
- Li, Y.; Lin, H.; Li, Y.; Cao, J. Association between Socio-Psycho-Behavioral Factors and Male Semen Quality: Systematic Review and Meta-Analyses. Fertil. Steril. 2011, 95, 116–123. [Google Scholar] [CrossRef]
- Bundhun, P.K.; Janoo, G.; Bhurtu, A.; Teeluck, A.R.; Soogund, M.Z.S.; Pursun, M.; Huang, F. Tobacco Smoking and Semen Quality in Infertile Males: A Systematic Review and Meta-Analysis. BMC Public Health 2019, 19, 36. [Google Scholar] [CrossRef]
- Rodprasert, W.; Toppari, J.; Virtanen, H.E. Environmental Toxicants and Male Fertility. Best Pract. Res. Clin. Obstet. Gynaecol. 2023, 86, 102298. [Google Scholar] [CrossRef] [PubMed]
- Asare-Anane, H.; Bannison, S.B.; Ofori, E.K.; Ateko, R.O.; Bawah, A.T.; Amanquah, S.D.; Oppong, S.Y.; Gandau, B.B.N.; Ziem, J.B. Tobacco Smoking Is Associated with Decreased Semen Quality. Reprod. Health 2016, 13, 90. [Google Scholar] [CrossRef] [PubMed]
- De Brucker, S.; Drakopoulos, P.; Dhooghe, E.; De Geeter, J.; Uvin, V.; Santos-Ribeiro, S.; Michielsen, D.; Tournaye, H.; De Brucker, M. The Effect of Cigarette Smoking on the Semen Parameters of Infertile Men. Gynecol. Endocrinol. 2020, 36, 1127–1130. [Google Scholar] [CrossRef]
- Sharma, R.; Harlev, A.; Agarwal, A.; Esteves, S.C. Cigarette Smoking and Semen Quality: A New Meta-Analysis Examining the Effect of the 2010 World Health Organization Laboratory Methods for the Examination of Human Semen. Eur. Urol. 2016, 70, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Kulaksiz, D.; Toprak, T.; Tokat, E.; Yilmaz, M.; Ramazanoglu, M.A.; Garayev, A.; Sulukaya, M.; Degirmentepe, R.B.; Allahverdiyev, E.; Gul, M.; et al. Sperm Concentration and Semen Volume Increase after Smoking Cessation in Infertile Men. Int. J. Impot. Res. 2022, 34, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Lotti, F.; Corona, G.; Vitale, P.; Maseroli, E.; Rossi, M.; Fino, M.G.; Maggi, M. Current Smoking Is Associated with Lower Seminal Vesicles and Ejaculate Volume, despite Higher Testosterone Levels, in Male Subjects of Infertile Couples. Hum. Reprod. 2015, 30, 590–602. [Google Scholar] [CrossRef]
- Harlev, A.; Agarwal, A.; Gunes, S.O.; Shetty, A.; du Plessis, S.S. Smoking and Male Infertility: An Evidence-Based Review. World J. Mens. Health 2015, 33, 143. [Google Scholar] [CrossRef]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative Stress and Male Infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef]
- Viloria, T.; Meseguer, M.; Martínez-Conejero, J.A.; O’Connor, J.E.; Remohí, J.; Pellicer, A.; Garrido, N. Cigarette Smoking Affects Specific Sperm Oxidative Defenses but Does Not Cause Oxidative DNA Damage in Infertile Men. Fertil. Steril. 2010, 94, 631–637. [Google Scholar] [CrossRef]
- Yu, B.; Chen, J.; Liu, D.; Zhou, H.; Xiao, W.; Xia, X.; Huang, Z. Cigarette Smoking Is Associated with Human Semen Quality in Synergy with Functional NRF2 Polymorphisms. Biol. Reprod. 2013, 89, 5. [Google Scholar] [CrossRef]
- Kiziler, A.R.; Aydemir, B.; Onaran, I.; Alici, B.; Ozkara, H.; Gulyasar, T.; Akyolcu, M.C. High Levels of Cadmium and Lead in Seminal Fluid and Blood of Smoking Men Are Associated with High Oxidative Stress and Damage in Infertile Subjects. Biol. Trace Elem. Res. 2007, 120, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Elshal, M.F.; El-Sayed, I.H.; Elsaied, M.A.; El-Masry, S.A.; Kumosani, T.A. Sperm Head Defects and Disturbances in Spermatozoal Chromatin and DNA Integrities in Idiopathic Infertile Subjects: Association with Cigarette Smoking. Clin. Biochem. 2009, 42, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Ou, Z.; Wen, Q.; Deng, Y.; Yu, Y.; Chen, Z.; Sun, L. Cigarette Smoking Is Associated with High Level of Ferroptosis in Seminal Plasma and Affects Semen Quality. Reprod. Biol. Endocrinol. 2020, 18, 55. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.; Santos, M.A.S.; Fardilha, M.; Silva, J.V. Stress Response Pathways in the Male Germ Cells and Gametes. Mol. Hum. Reprod. 2020, 26, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Widlak, W.; Vydra, N.; Malusecka, E.; Dudaladava, V.; Winiarski, B.; Ścieglińska, D.; Widlak, P. Heat Shock Transcription Factor 1 Down-Regulates Spermatocyte-Specific 70 KDa Heat Shock Protein Expression Prior to the Induction of Apoptosis in Mouse Testes. Genes Cells 2007, 12, 487–499. [Google Scholar] [CrossRef]
- Lampros, M.; Vlachos, N.; Voulgaris, S.; Alexiou, G.A. The Role of Hsp27 in Chemotherapy Resistance. Biomedicines 2022, 10, 897. [Google Scholar] [CrossRef]
- Shan, R.; Liu, N.; Yan, Y.; Liu, B. Apoptosis, Autophagy and Atherosclerosis: Relationships and the Role of Hsp27. Pharmacol. Res. 2021, 166, 105169. [Google Scholar] [CrossRef]
- Gur, Y.; Breitbart, H. Mammalian Sperm Translate Nuclear-Encoded Proteins by Mitochondrial-Type Ribosomes. Genes Dev. 2006, 20, 411–416. [Google Scholar] [CrossRef]
- Zhu, Z.; Umehara, T.; Okazaki, T.; Goto, M.; Fujita, Y.; Hoque, S.A.M.; Kawai, T.; Zeng, W.; Shimada, M. Gene Expression and Protein Synthesis in Mitochondria Enhance the Duration of High-Speed Linear Motility in Boar Sperm. Front. Physiol. 2019, 10, 252. [Google Scholar] [CrossRef]
 ) represent the different municipalities of residence of the studied population.
) represent the different municipalities of residence of the studied population.
) represent the different municipalities of residence of the studied population.
) represent the different municipalities of residence of the studied population.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Host | Dilution | Supplier | Cat. No |
|---|---|---|---|---|
| SOD1 | Mouse | 1:1000 | Millipore | MABC684 |
| SOD2 | Rabbit | 1:1000 | Abcam | Ab13533 |
| HSP27 | Mouse | 1:1000 | Santa Cruz Biotechnology | sc-13132 |
| p-HSP27 (Ser82) | Mouse | 1:1000 | Santa Cruz Biotechnology | sc-166693 |
| GPx4 | Rabbit | 1:1000 | Millipore | ABC269 |
| HSF1 | Mouse | 1:500 | Santa Cruz Biotechnology | sc-17757 |
| HSP90 | Mouse | 1:2000 | ProteinTech | 13171-1-AP |
| Parameter | N | |
|---|---|---|
| Age (years) | 32.5 ± 7.4 | 30 |
| BMI (kg/m2) | 24.3 ± 4.0 | 30 |
| Education | ||
| Primary | 10 | 3 |
| High School | 63 | 19 |
| University | 27 | 8 |
| Employment Situation | ||
| Employed | 90 | 27 |
| Unemployed | 3 | 1 |
| Student | 7 | 2 |
| Exposure to chemicals at work | ||
| No | 47 | 14 |
| Yes | 53 | 16 |
| Smoking status | ||
| No | 53 | 16 |
| Yes | 47 | 14 |
| Alcohol consumption | ||
| No | 47 | 14 |
| Yes | 53 | 16 |
| Physical activity | ||
| No | 43 | 13 |
| Yes | 57 | 17 |
| Semen parameters | ||
| Sexual abstinence (days) | 4 ± 1 | 29 |
| Volume (mL) | 3.1 ± 1.4 | 30 |
| Concentration (×106/mL) | 47 ± 36 | 28 |
| Total spermatozoa number (×106) | 133 ± 110 | 28 |
| Total motility (Progressive and non-progressive, %) | 59 ± 2 | 29 |
| Progressive motility (%) | 42 ± 15 | 29 |
| Non-progressive motility (%) | 17 ± 9 | 29 |
| Immotile (%) | 41 ± 13 | 29 |
| Normal morphology (%) | 6 ± 2 | 28 |
| Head defects (%) | 86 ± 5 | 28 |
| Midpiece defects (%) | 50 ± 10 | 28 |
| Tail defects (%) | 26 ± 7 | 28 |
| Teratozoospermia index | 1.72 ± 0.15 | 28 |
| Consumed Items | Never | 1 to 3 Times per Month | 1 to 3 Times per Week | 4 to 6 Times per Week | Daily |
|---|---|---|---|---|---|
| Dairy products | 3 | 17 | 23 | 23 | 33 |
| Coffee | 3 | 3 | 10 | 3 | 77 |
| Tea | 27 | 43 | 20 | 3 | 7 |
| Eggs | 3 | 27 | 63 | 7 | 0 |
| Fresh vegetables | 3 | 0 | 40 | 43 | 13 |
| Canned vegetables | 27 | 30 | 30 | 13 | 0 |
| Fresh fruit or fruit juices | 0 | 10 | 23 | 27 | 40 |
| Canned fruit | 53 | 43 | 3 | 0 | 0 |
| Refrigerants | 23 | 30 | 17 | 7 | 23 |
| Organic food | 17 | 30 | 20 | 23 | 10 |
| Fast-food | 10 | 70 | 17 | 0 | 0 |
| Poultry | 0 | 0 | 63 | 33 | 3 |
| Meat | 0 | 23 | 50 | 23 | 3 |
| Fresh or frozen fish | 0 | 20 | 63 | 17 | 0 |
| Canned fish | 40 | 37 | 23 | 0 | 0 |
| Shellfish | 17 | 63 | 20 | 0 | 0 |
| Grilled food | 3 | 33 | 47 | 13 | 3 |
| Smoked food | 3 | 53 | 30 | 7 | 7 |
| Bottled water | 10 | 3 | 3 | 7 | 77 |
| Parameter | Non-Smokers (N = 16) | Smokers (N = 14) | p-Value |
|---|---|---|---|
| Age (years) | 31.7 ± 7.9 | 33.4 ± 6.7 | 0.545 |
| BMI (kg/m2) | 25.2 ± 4.9 | 23.2 ± 2.5 | 0.184 |
| Semen quality parameters | |||
| Volume (mL) | 3.7 ± 1.5 | 2.3 ± 0.9 | 0.004 ** |
| Concentration (×106/mL) | 54.5 ± 34.4 | 38.5 ± 36.1 | 0.242 |
| Total spermatozoa count (×106) | 181.9 ± 113.0 | 84.3 ± 85.1 | 0.044 * |
| Total motility (Progressive and non-progressive, %) | 62.6 ± 13.9 | 55.8 ± 11.6 | 0.093 |
| Progressive motility (%) | 46.2 ± 14.4 | 38.3 ± 15.0 | 0.159 |
| Non-progressive motility (%) | 17.1 ± 7.7 | 17.5 ± 10.6 | 0.813 |
| Immotile (%) | 37.4 ± 13.9 | 44.2 ± 11.6 | 0.093 |
| Normal morphology (%) | 6.4 ± 1.6 | 5.4 ± 2.2 | 0.069 |
| Head defects (%) | 85.2 ± 5.5 | 86.3 ± 4.8 | 0.210 |
| Midpiece defects (%) | 50.7 ± 9.8 | 50.1 ± 10.2 | 0.881 |
| Tail defects (%) | 25.5 ± 7.6 | 25.6 ± 6.0 | 0.913 |
| Teratozoospermia index | 1.72 ± 0.17 | 1.72 ± 0.13 | 0.891 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henriques, M.C.; Santiago, J.; Patrício, A.; Herdeiro, M.T.; Loureiro, S.; Fardilha, M. Smoking Induces a Decline in Semen Quality and the Activation of Stress Response Pathways in Sperm. Antioxidants 2023, 12, 1828. https://doi.org/10.3390/antiox12101828
Henriques MC, Santiago J, Patrício A, Herdeiro MT, Loureiro S, Fardilha M. Smoking Induces a Decline in Semen Quality and the Activation of Stress Response Pathways in Sperm. Antioxidants. 2023; 12(10):1828. https://doi.org/10.3390/antiox12101828
Chicago/Turabian StyleHenriques, Magda Carvalho, Joana Santiago, António Patrício, Maria Teresa Herdeiro, Susana Loureiro, and Margarida Fardilha. 2023. "Smoking Induces a Decline in Semen Quality and the Activation of Stress Response Pathways in Sperm" Antioxidants 12, no. 10: 1828. https://doi.org/10.3390/antiox12101828
APA StyleHenriques, M. C., Santiago, J., Patrício, A., Herdeiro, M. T., Loureiro, S., & Fardilha, M. (2023). Smoking Induces a Decline in Semen Quality and the Activation of Stress Response Pathways in Sperm. Antioxidants, 12(10), 1828. https://doi.org/10.3390/antiox12101828