

Anserine and Carnosine Induce HSP70-Dependent H2S Formation in Endothelial Cells and Murine Kidney

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Hsp70-Knockout MCEC
2.3. Maximal H2S Production Capacity
2.4. Anserine and Carnosine Concentrations
2.5. Western Blotting
2.6. Carnosinase Activity
2.7. Mice
2.8. Statistical Analysis
3. Results
3.1. Dipeptide-Induced H2S Formation in Tubular Epithelial and Endothelial Cells
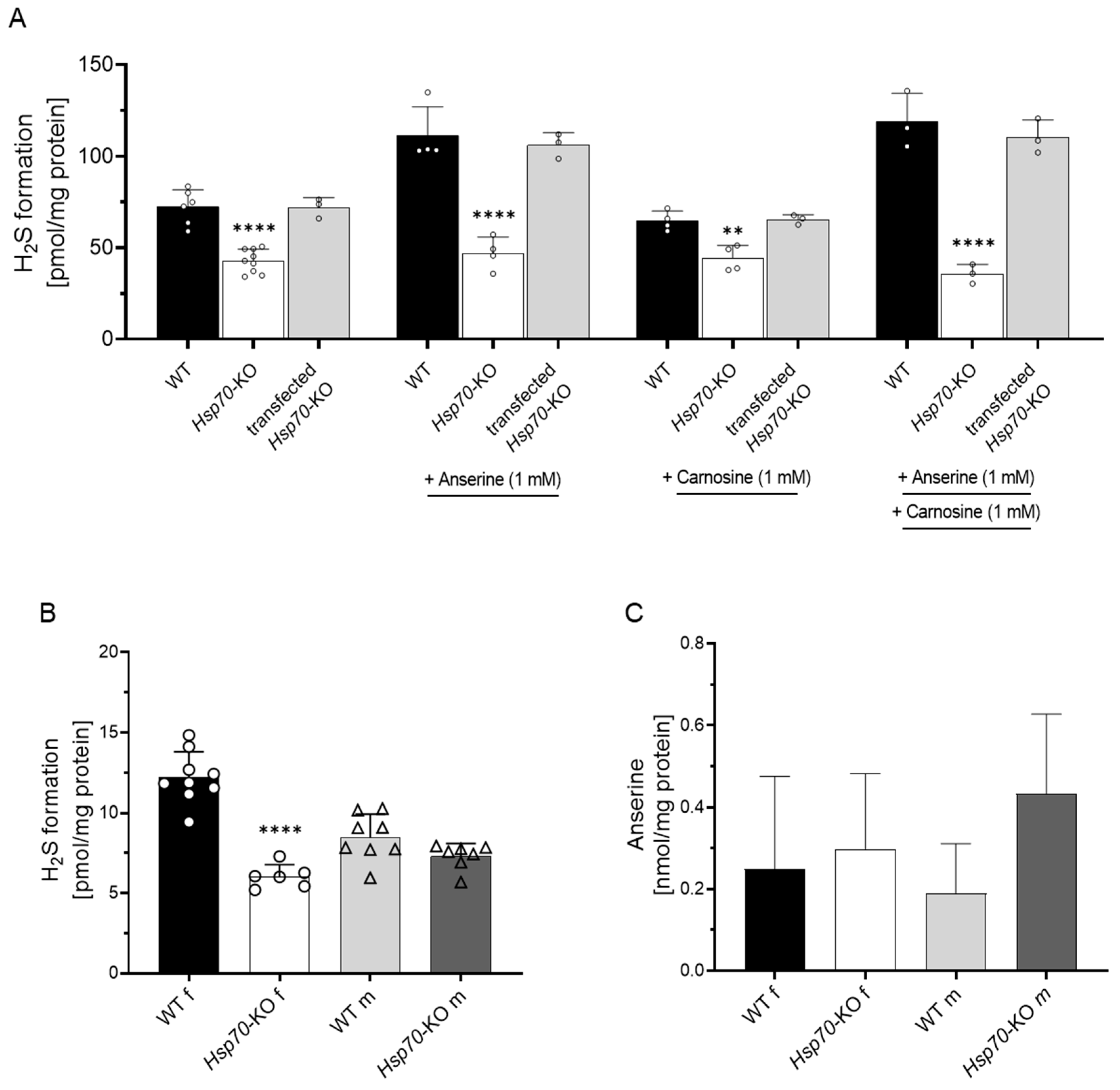
3.2. Anserine-Induced H2S Formation Depends on HSP70
3.3. H2S Formation in Murine Kidney Tissue
3.4. Kidney Anserine Abundance and H2S Formation
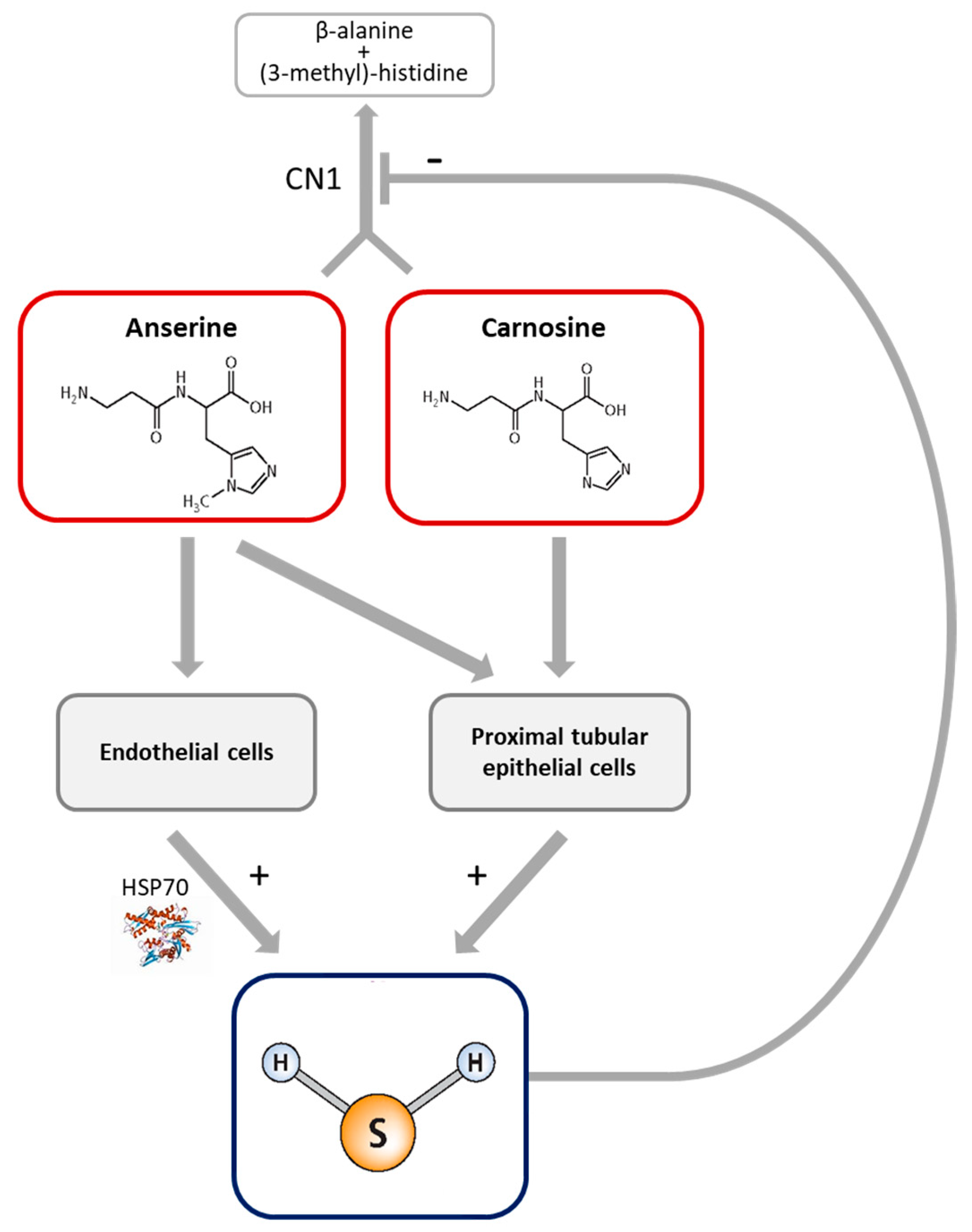
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kwiatkowski, S.; Kiersztan, A.; Drozak, J. Biosynthesis of carnosine and related dipeptides in vertebrates. Curr. Protein Pept. Sci. 2018, 19, 771–789. [Google Scholar] [CrossRef] [PubMed]
- Teufel, M.; Saudek, V.; Ledig, J.P.; Bernhardt, A.; Boularand, S.; Carreau, A.; Cairns, N.J.; Carter, C.; Cowley, D.J.; Duverger, D.; et al. Sequence identification and characterization of human carnosinase and a closely related non-specific dipeptidase. J. Biol. Chem. 2003, 278, 6251–6531. [Google Scholar] [CrossRef] [PubMed]
- Janssen, B.; Hohenadel, D.; Brinkkoetter, P.; Peters, V.; Rind, N.; Fischer, C.; Rychlik, I.; Cerna, M.; Romzova, M.; de Heer, E.; et al. Carnosine as a protective factor in diabetic nephropathy: Association with a leucine repeat of the carnosinase gene CNDP1. Diabetes 2005, 54, 2320–2327. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Liu, X.Q.; Zhang, S.Q.; Qi, X.M.; Zhang, Q.; Yard, B.; Wu, Y.G. Correlation between serum carnosinase concentration and renal damage in diabetic nephropathy patients. Amino Acids 2021, 53, 687–700. [Google Scholar] [CrossRef]
- Peters, V.; Yard, B.; Schmitt, C.P. Carnosine and diabetic nephropathy. Curr. Med. Chem. 2019, 27, 1801–1812. [Google Scholar] [CrossRef]
- Kilis-Pstrusinska, K. Carnosine and Kidney Diseases: What We Currently Know? Curr. Med. Chem. 2020, 27, 1764–1781. [Google Scholar] [CrossRef]
- Matthews, J.J.; Dolan, E.; Swinton, P.A.; Santos, L.; Artioli, G.G.; Turner, M.D.; Elliott-Sale, K.J.; Sale, C. Effect of Carnosine or β-Alanine Supplementation on Markers of Glycemic Control and Insulin Resistance in Humans and Animals: A Systematic Review and Meta-analysis. Adv. Nutr. 2021, 12, 2216–2231. [Google Scholar] [CrossRef]
- Caruso, G. Unveiling the Hidden Therapeutic Potential of Carnosine, a Molecule with a Multimodal Mechanism of Action: A Position Paper. Molecules 2022, 27, 3303. [Google Scholar] [CrossRef]
- Oppermann, H.; Elsel, S.; Birkemeyer, C.; Meixensberger, J.; Gaunitz, F. Erythrocytes Prevent Degradation of Carnosine by Human Serum Carnosinase. Int. J. Mol. Sci. 2021, 22, 12802. [Google Scholar] [CrossRef]
- Holeček, M. Histidine in Health and Disease: Metabolism, Physiological Importance, and Use as a Supplement. Nutrients 2020, 12, 848. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; de Courten, B.; Regazzoni, L.; Gilardoni, E.; Ferrario, G.; Baron, G.; Altomare, A.; D’Amato, A.; Vistoli, G.; Carini, M. Understanding the antioxidant and carbonyl sequestering activity of carnosine: Direct and indirect mechanisms. Free Radic. Res. 2021, 55, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Weigand, T.; Singler, B.; Fleming, T.; Nawroth, P.; Klika, K.D.; Thiel, C.; Baelde, H.; Garbade, S.F.; Wagner, A.H.; Hecker, M.; et al. Carnosine Catalyzes the Formation of the Oligo/Polymeric Products of Methylglyoxal. Cell Physiol. Biochem. 2018, 46, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Scuto, M.; Trovato Salinaro, A.; Modafferi, S.; Polimeni, A.; Pfeffer, T.; Weigand, T.; Calabrese, V.; Schmitt, C.P.; Peters, V. Carnosine Activates Cellular Stress Response in Podocytes and Reduces Glycative and Lipoperoxidative Stress. Biomedicines 2020, 8, 177. [Google Scholar] [CrossRef]
- Peters, V.; Calabrese, V.; Forsberg, E.; Volk, N.; Fleming, T.; Baelde, H.; Weigand, T.; Thiel, C.; Trovato, A.; Scuto, M.; et al. Protective Actions of Anserine Under Diabetic Conditions. Int. J. Mol. Sci. 2018, 19, 2751. [Google Scholar] [CrossRef]
- Chebotareva, N.; Bobkova, I.; Shilov, E. Heat shock proteins and kidney disease: Perspectives of HSP therapy. Cell Stress Chaperones 2017, 22, 319–343. [Google Scholar] [CrossRef]
- Calabrese, V.; Scuto, M.; Salinaro, A.T.; Dionisio, G.; Modafferi, S.; Ontario, M.L.; Greco, V.; Sciuto, S.; Schmitt, C.P.; Calabrese, E.J.; et al. Hydrogen Sulfide and Carnosine: Modulation of Oxidative Stress and Inflammation in Kidney and Brain Axis. Antioxidants 2020, 9, 1303. [Google Scholar] [CrossRef]
- Bellini, S.; Barutta, F.; Mastrocola, R.; Imperatore, L.; Bruno, G.; Gruden, G. Heat Shock Proteins in Vascular Diabetic Complications: Review and Future Perspective. Int. J. Mol. Sci. 2017, 18, 2709. [Google Scholar] [CrossRef]
- Calvert, J.W.; Jha, S.; Gundewar, S.; Elrod, J.W.; Ramachandran, A.; Pattillo, C.B.; Kevil, C.G.; Lefer, D.J. Hydrogen sulfide mediates cardioprotection through Nrf2 signaling. Circ. Res. 2009, 105, 365–374. [Google Scholar] [CrossRef]
- Peleli, M.; Zampas, P.; Papapetropoulos, A. Hydrogen Sulfide and the Kidney: Physiological Roles, Contribution to Pathophysiology, and Therapeutic Potential. Antioxid. Redox Signal. 2022, 36, 220–243. [Google Scholar] [CrossRef]
- Kabil, O.; Banerjee, R. Enzymology of H2S biogenesis, decay and signaling. Antioxid. Redox Signal. 2014, 20, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Li, H.; Li, S.; Yang, G. Regulation of cystathionine gamma-lyase/H₂S system and its pathological implication. Front. Biosci. 2014, 19, 1355–1369. [Google Scholar] [CrossRef] [PubMed]
- Beck, K.F.; Pfeilschifter, J. The Pathophysiology of H(2)S in Renal Glomerular Diseases. Biomolecules 2022, 12, 207. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Q.; Xue, N.; Chen, J.; Shen, Z.; Cui, X.; Fang, Y.; Ding, X. Low Plasma Hydrogen Sulfide Is Associated with Impaired Renal Function and Cardiac Dysfunction. Am. J. Nephrol. 2018, 47, 361–371. [Google Scholar] [CrossRef]
- Ngowi, E.E.; Sarfraz, M.; Afzal, A.; Khan, N.H.; Khattak, S.; Zhang, X.; Li, T.; Duan, S.F.; Ji, X.Y.; Wu, D.D. Roles of Hydrogen Sulfide Donors in Common Kidney Diseases. Front. Pharmacol. 2020, 11, 564281. [Google Scholar] [CrossRef]
- Koning, A.M.; Frenay, A.R.; Leuvenink, H.G.; van Goor, H. Hydrogen sulfide in renal physiology, disease and transplantation--the smell of renal protection. Nitric Oxide Biol. Chem. 2015, 46, 37–49. [Google Scholar] [CrossRef]
- de Koning, M.L.; van Dorp, P.; Assa, S.; Hartman, M.H.; Voskuil, M.; Anthonio, R.L.; Veen, D.; Pundziute-Do Prado, G.; Leiner, T.; van Goor, H.; et al. Rationale and Design of the Groningen Intervention Study for the Preservation of Cardiac Function with Sodium Thiosulfate after St-segment Elevation Myocardial Infarction (GIPS-IV) trial. Am. Heart J. 2022, 243, 167–176. [Google Scholar] [CrossRef]
- Lu, P.H.; Chuo, H.E.; Kuo, K.L.; Liao, J.F.; Lu, P.H. Clinical Efficacy and Safety of Sodium Thiosulfate in the Treatment of Uremic Pruritus: A Meta-Analysis of Randomized Controlled Trials. Toxins 2021, 13, 769. [Google Scholar] [CrossRef]
- Merz, T.; McCook, O.; Brucker, C.; Waller, C.; Calzia, E.; Radermacher, P.; Datzmann, T. H(2)S in Critical Illness-A New Horizon for Sodium Thiosulfate? Biomolecules 2022, 12, 543. [Google Scholar] [CrossRef]
- Zhou, X.; Feng, Y.; Zhan, Z.; Chen, J. Hydrogen sulfide alleviates diabetic nephropathy in a streptozotocin-induced diabetic rat model. J. Biol. Chem. 2014, 289, 28827–28834. [Google Scholar] [CrossRef]
- Ahn, Y.J.; Lee, Y.J.; Lee, J.; Lee, D.; Park, H.K.; Lee, G.J. Colorimetric detection of endogenous hydrogen sulfide production in living cells. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 177, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Weigand, T.; Colbatzky, F.; Pfeffer, T.; Garbade, S.F.; Klingbeil, K.; Colbatzky, F.; Becker, M.; Zemva, J.; Bulkescher, R.; Schürfeld, R.; et al. A Global Cndp1-Knock-Out Selectively Increases Renal Carnosine and Anserine Concentrations in an Age- and Gender-Specific Manner in Mice. Int. J. Mol. Sci. 2020, 21, 4887. [Google Scholar] [CrossRef] [PubMed]
- Elbarbary, N.S.; Ismail, E.A.R.; El-Naggar, A.R.; Hamouda, M.H.; El-Hamamsy, M. The effect of 12 weeks carnosine supplementation on renal functional integrity and oxidative stress in pediatric patients with diabetic nephropathy: A randomized placebo-controlled trial. Pediatr. Diabetes 2017, 19, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Siriwattanasit, N.; Satirapoj, B.; Supasyndh, O. Effect of Oral carnosine supplementation on urinary TGF-β in diabetic nephropathy: A randomized controlled trial. BMC Nephrol. 2021, 22, 236. [Google Scholar] [CrossRef]
- Menon, K.; Marquina, C.; Liew, D.; Mousa, A.; de Courten, B. Histidine-containing dipeptides reduce central obesity and improve glycaemic outcomes: A systematic review and meta-analysis of randomized controlled trials. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2020, 21, e12975. [Google Scholar] [CrossRef]
- Ji, K.; Xue, L.; Cheng, J.; Bai, Y. Preconditioning of H2S inhalation protects against cerebral ischemia/reperfusion injury by induction of HSP70 through PI3K/Akt/Nrf2 pathway. Brain Res. Bull. 2016, 121, 68–74. [Google Scholar] [CrossRef]
- Lavilla, C.J.; Billacura, M.P.; Hanna, K.; Boocock, D.J.; Coveney, C.; Miles, A.K.; Foulds, G.A.; Murphy, A.; Tan, A.; Jackisch, L.; et al. Carnosine protects stimulus-secretion coupling through prevention of protein carbonyl adduction events in cells under metabolic stress. Free Radic. Biol. Med. 2021, 175, 65–79. [Google Scholar] [CrossRef]
- Saadah, L.M.; Deiab, G.I.A.; Al-Balas, Q.; Basheti, I.A. Carnosine to Combat Novel Coronavirus (nCoV): Molecular Docking and Modeling to Cocrystallized Host Angiotensin-Converting Enzyme 2 (ACE2) and Viral Spike Protein. Molecules 2020, 25, 5605. [Google Scholar] [CrossRef]
- Nakagawa, K.; Ueno, A.; Nishikawa, Y. Interactions between carnosine and captopril on free radical scavenging activity and angiotensin-converting enzyme activity in vitro. Yakugaku Zasshi 2006, 126, 37–42. [Google Scholar] [CrossRef]
- Calabrese, V.; Cornelius, C.; Mancuso, C.; Pennisi, G.; Calafato, S.; Bellia, F.; Bates, T.E.; Giuffrida Stella, A.M.; Schapira, T.; Dinkova Kostova, A.T.; et al. Cellular stress response: A novel target for chemoprevention and nutritional neuroprotection in aging, neurodegenerative disorders and longevity. Neurochem. Res. 2008, 33, 2444–2471. [Google Scholar] [CrossRef]
- Caruso, G.; Privitera, A.; Antunes, B.M.; Lazzarino, G.; Lunte, S.M.; Aldini, G.; Caraci, F. The Therapeutic Potential of Carnosine as an Antidote against Drug-Induced Cardiotoxicity and Neurotoxicity: Focus on Nrf2 Pathway. Molecules 2022, 27, 4452. [Google Scholar] [CrossRef] [PubMed]
- Fresta, C.G.; Fidilio, A.; Lazzarino, G.; Musso, N.; Grasso, M.; Merlo, S.; Amorini, A.M.; Bucolo, C.; Tavazzi, B.; Lazzarino, G.; et al. Modulation of Pro-Oxidant and Pro-Inflammatory Activities of M1 Macrophages by the Natural Dipeptide Carnosine. Int. J. Mol. Sci. 2020, 21, 776. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.M.; Chen, X.; Cheng, K.; Shi, Q.; Peng, K. Anserine and glucosamine supplementation attenuates the levels of inflammatory markers in rats with rheumatoid arthritis. AMB Express 2020, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Sakano, T.; Egusa, A.S.; Kawauchi, Y.; Wu, J.; Nishimura, T.; Nakao, N.; Kuramoto, A.; Kawashima, T.; Shiotani, S.; Okada, Y.; et al. Pharmacokinetics and tissue distribution of orally administrated imidazole dipeptides in carnosine synthase gene knockout mice. Biosci. Biotechnol. Biochem. 2022, 86, 1276–1285. [Google Scholar] [CrossRef]
- Wang-Eckhardt, L.; Becker, I.; Wang, Y.; Yuan, J.; Eckhardt, M. Absence of endogenous carnosine synthesis does not increase protein carbonylation and advanced lipoxidation end products in brain, kidney or muscle. Amino Acids 2022, 54, 1013–1023. [Google Scholar] [CrossRef]
- Peters, V.; Lanthaler, B.; Amberger, A.; Fleming, T.; Forsberg, E.; Hecker, M.; Wagner, A.H.; Yue, W.W.; Hoffmann, G.F.; Nawroth, P.; et al. Carnosine metabolism in diabetes is altered by reactive metabolites. Amino Acids 2015, 47, 2367–2376. [Google Scholar] [CrossRef]
- Zhang, S.; Albrecht, T.; Rodriguez-Niño, A.; Qiu, J.; Schnuelle, P.; Peters, V.; Schmitt, C.P.; van den Born, J.; Bakker, S.J.L.; Lammert, A.; et al. Carnosinase concentration, activity, and CNDP1 genotype in patients with type 2 diabetes with and without nephropathy. Amino Acids 2019, 51, 611–617. [Google Scholar] [CrossRef]
- Peters, V.; Jansen, E.E.; Jakobs, C.; Riedl, E.; Janssen, B.; Yard, B.A.; Wedel, J.; Hoffmann, G.F.; Zschocke, J.; Gotthardt, D.; et al. Anserine inhibits carnosine degradation but in human serum carnosinase (CN1) is not correlated with histidine dipeptide concentration. Clin. Chim Acta 2011, 412, 263–267. [Google Scholar] [CrossRef]
- Peters, V.; Schmitt, C.P.; Weigand, T.; Klingbeil, K.; Thiel, C.; van den Berg, A.; Calabrese, V.; Nawroth, P.; Fleming, T.; Forsberg, E.; et al. Allosteric inhibition of carnosinase (CN1) by inducing a conformational shift. J. Enzym. Inhib Med. Chem 2017, 32, 1102–1110. [Google Scholar] [CrossRef]
- Teoh, J.P.; Li, X.; Simoncini, T.; Zhu, D.; Fu, X. Estrogen-Mediated Gaseous Signaling Molecules in Cardiovascular Disease. Trends Endocrinol. Metab. TEM 2020, 31, 773–784. [Google Scholar] [CrossRef]
- de Koning, M.L.Y.; Assa, S.; Maagdenberg, C.G.; van Veldhuisen, D.J.; Pasch, A.; van Goor, H.; Lipsic, E.; van der Harst, P. Safety and Tolerability of Sodium Thiosulfate in Patients with an Acute Coronary Syndrome Undergoing Coronary Angiography: A Dose-Escalation Safety Pilot Study (SAFE-ACS). J. Interv. Cardiol. 2020, 2020, 6014915. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Yang, Y.; Wei, S.; Huang, X.; Peng, Z.; Ke, X.; Zeng, Z.; Song, Y. Hydrogen Sulfide Protects Against High Glucose-Induced Human Umbilical Vein Endothelial Cell Injury Through Activating PI3K/Akt/eNOS Pathway. Drug Des. Dev. Ther. 2020, 14, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liao, S.; Pan, Z.; Jiang, S.; Fan, J.; Yu, S.; Xue, L.; Yang, J.; Ma, S.; Liu, T.; et al. Hydrogen sulfide alleviates particulate matter-induced emphysema and airway inflammation by suppressing ferroptosis. Free Radic. Biol. Med. 2022, 186, 1–16. [Google Scholar] [CrossRef]
- Zhao, H.; Qiu, Y.; Wu, Y.; Sun, H.; Gao, S. Protective Effects of GYY4137 on Renal Ischaemia/Reperfusion Injury through Nrf2-Mediated Antioxidant Defence. Kidney Blood Press. Res. 2021, 46, 257–265. [Google Scholar] [CrossRef]
- Uyy, E.; Suica, V.I.; Boteanu, R.M.; Safciuc, F.; Cerveanu-Hogas, A.; Ivan, L.; Stavaru, C.; Simionescu, M.; Antohe, F. Diabetic nephropathy associates with deregulation of enzymes involved in kidney sulphur metabolism. J. Cell. Mol. Med. 2020, 24, 12131–12140. [Google Scholar] [CrossRef]
- Rodriguez-Niño, A.; Pastene, D.O.; Hettler, S.A.; Qiu, J.; Albrecht, T.; Vajpayee, S.; Perciaccante, R.; Gretz, N.; Bakker, S.J.L.; Krämer, B.K.; et al. Influence of carnosine and carnosinase-1 on diabetes-induced afferent arteriole vasodilation: Implications for glomerular hemodynamics. Am. J. Physiol. Ren. Physiol. 2022, 323, F69–F80. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Q.; Jiang, L.; Lei, L.; Nie, Z.Y.; Zhu, W.; Wang, S.; Zeng, H.X.; Zhang, S.Q.; Zhang, Q.; Yard, B.; et al. Carnosine alleviates diabetic nephropathy by targeting GNMT, a key enzyme mediating renal inflammation and fibrosis. Clin. Sci. 2020, 134, 3175–3193. [Google Scholar] [CrossRef] [PubMed]
- Everaert, I.; Van der Stede, T.; Stautemas, J.; Hanssens, M.; van Aanhold, C.; Baelde, H.; Vanhaecke, L.; Derave, W. Oral anserine supplementation does not attenuate type-2 diabetes or diabetic nephropathy in BTBR ob/ob mice. Amino Acids 2021, 53, 1269–1277. [Google Scholar] [CrossRef]
- Drozak, J.; Chrobok, L.; Poleszak, O.; Jagielski, A.K.; Derlacz, R. Molecular identification of carnosine N-methyltransferase as chicken histamine N-methyltransferase-like protein (hnmt-like). PLoS ONE 2013, 8, e64805. [Google Scholar] [CrossRef]
- Peters, V.; Schmitt, C.P.; Zschocke, J.; Gross, M.L.; Brismar, K.; Forsberg, E. Carnosine treatment largely prevents alterations of renal carnosine metabolism in diabetic mice. Amino Acids 2012, 42, 2411–2416. [Google Scholar] [CrossRef]
- Luo, Y.; Zuo, Y.; Shi, G.; Xiang, H.; Gu, H. Progress on the reaction-based methods for detection of endogenous hydrogen sulfide. Anal. Bioanal. Chem. 2022, 414, 2809–2839. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 9 | Age of Mice (Weeks) | Gender | Anserine (nmol/mg) | Carnosine (nmol/mg) | H2S Formation (pmol/mg) |
|---|---|---|---|---|---|
| WT | 23–25 | f | 0.8 ± 0.4 | <0.1 nmol | 10.74 ± 1.21 |
| WT | 23–25 | m | 0.7 ± 0.5 | <0.1 nmol | 9.44 ± 1.78 |
| Cndp1-KO | 23–25 | f | 3.0 ± 1.3 | 3.8 ± 2.36 | 21.33 ± 4.46 a |
| Cndp1-KO | 23–25 | m | 1.8 ± 0.8 a | 0.9 ± 0.47 | 9.62 ± 1.79 |
| WT | 47–51 | f | 0.3 ± 0.04 | <0.1 nmol | 10.48 ± 2.20 |
| WT | 47–51 | m | 0.3 ± 0.2 | <0.1 nmol | 9.88 ± 1.11 |
| Cndp1-KO | 47–51 | f | 3.1 ± 1.3 a | 1.81 ± 0.72 | 29.22 ± 6.24 a |
| Cndp1-KO | 47–51 | m | 1.1 ± 0.3 a | 0.32 ± 0.19 | 6.38 ± 1.14 a |
| STZ WT | 23–25 | f | 0.7 ± 0.4 | <0.1 nmol | 13.20 ± 4.62 |
| STZ WT | 23–25 | m | 0.8 ± 0.4 | <0.1 nmol | 13.43 ± 1.67 b |
| STZ Cndp1-KO | 23–25 | f | 1.0 ± 0.4 b | 0.80 ± 0.29 b | 13.84 ± 2.53 b |
| STZ Cndp1-KO | 23–25 | m | 1.2 ± 0.5 a,b | 0.68 ± 0.31 | 8.38 ± 3.21 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wetzel, C.; Pfeffer, T.; Bulkescher, R.; Zemva, J.; Modafferi, S.; Polimeni, A.; Salinaro, A.T.; Calabrese, V.; Schmitt, C.P.; Peters, V. Anserine and Carnosine Induce HSP70-Dependent H2S Formation in Endothelial Cells and Murine Kidney. Antioxidants 2023, 12, 66. https://doi.org/10.3390/antiox12010066
Wetzel C, Pfeffer T, Bulkescher R, Zemva J, Modafferi S, Polimeni A, Salinaro AT, Calabrese V, Schmitt CP, Peters V. Anserine and Carnosine Induce HSP70-Dependent H2S Formation in Endothelial Cells and Murine Kidney. Antioxidants. 2023; 12(1):66. https://doi.org/10.3390/antiox12010066
Chicago/Turabian StyleWetzel, Charlotte, Tilman Pfeffer, Ruben Bulkescher, Johanna Zemva, Sergio Modafferi, Alessandra Polimeni, Angela Trovato Salinaro, Vittorio Calabrese, Claus Peter Schmitt, and Verena Peters. 2023. "Anserine and Carnosine Induce HSP70-Dependent H2S Formation in Endothelial Cells and Murine Kidney" Antioxidants 12, no. 1: 66. https://doi.org/10.3390/antiox12010066
APA StyleWetzel, C., Pfeffer, T., Bulkescher, R., Zemva, J., Modafferi, S., Polimeni, A., Salinaro, A. T., Calabrese, V., Schmitt, C. P., & Peters, V. (2023). Anserine and Carnosine Induce HSP70-Dependent H2S Formation in Endothelial Cells and Murine Kidney. Antioxidants, 12(1), 66. https://doi.org/10.3390/antiox12010066