
Exploitation of Olive (Olea europaea L.) Seed Proteins as Upgraded Source of Bioactive Peptides with Multifunctional Properties: Focus on Antioxidant and Dipeptidyl-Dipeptidase—IV Inhibitory Activities, and Glucagon-like Peptide 1 Improved Modulation

 ,
,  , , ,
, , ,  ,
,  ,
,  ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical and Samples
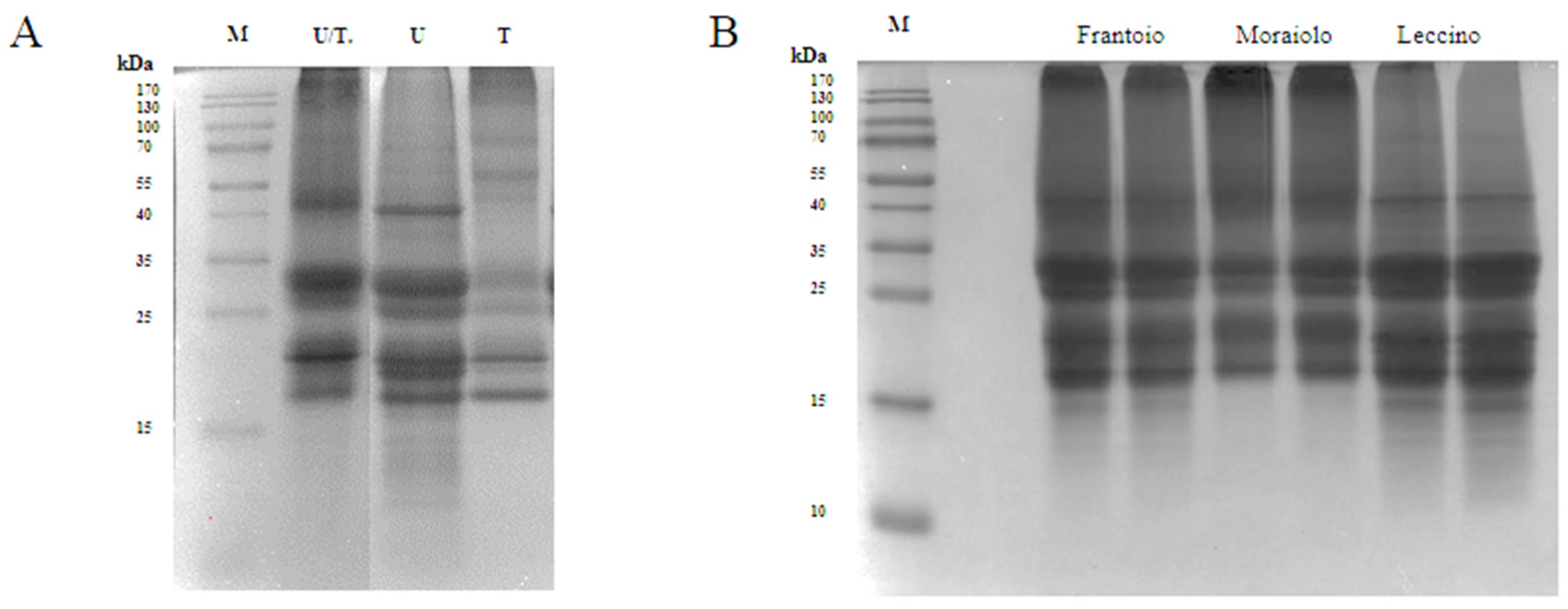
2.2. Protein Extraction from Olive Seed
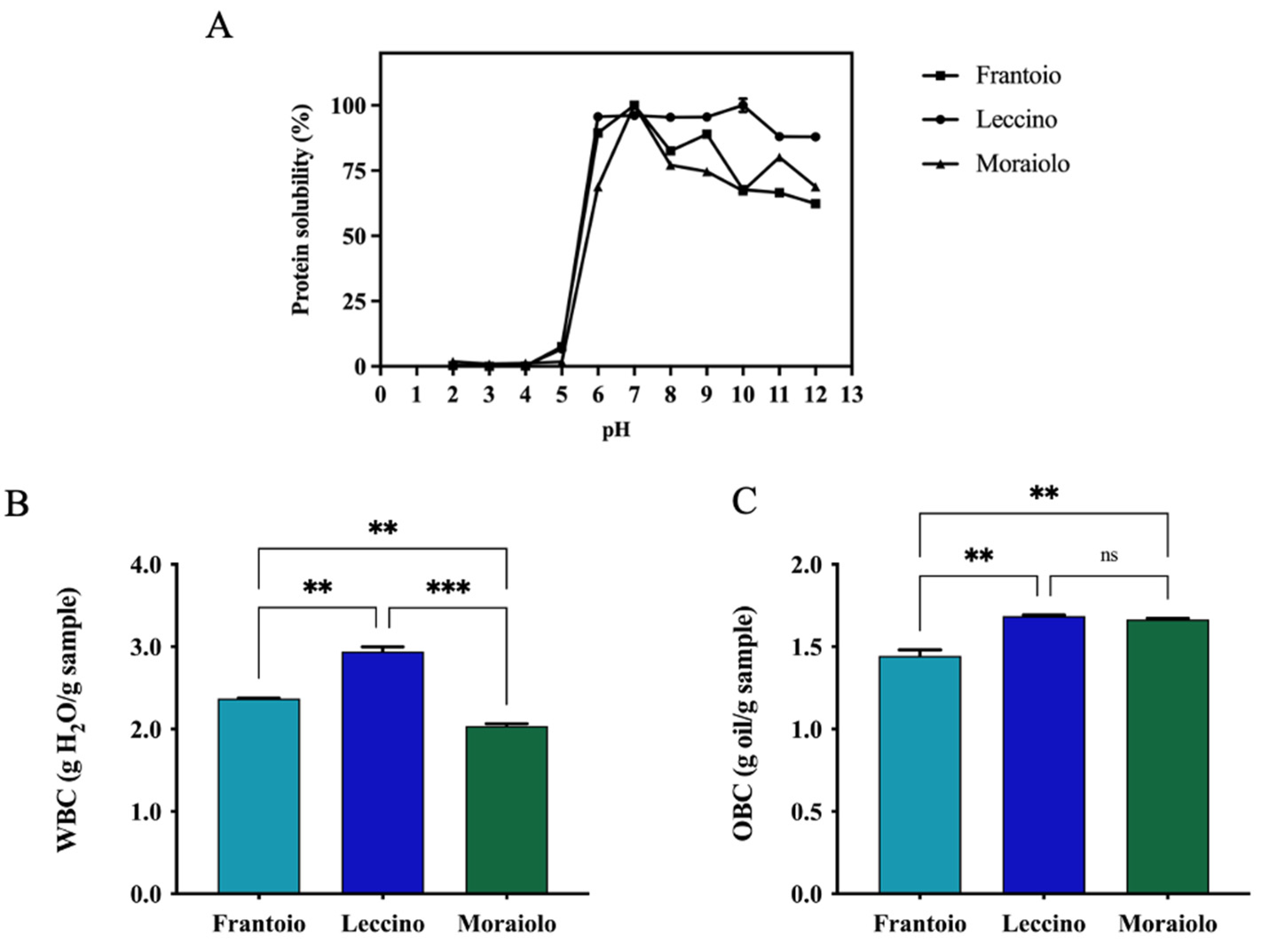
2.3. Protein Solubility (PS), Water-Binding Capacity (WBC), and Oil-Binding Capacity (OBC)
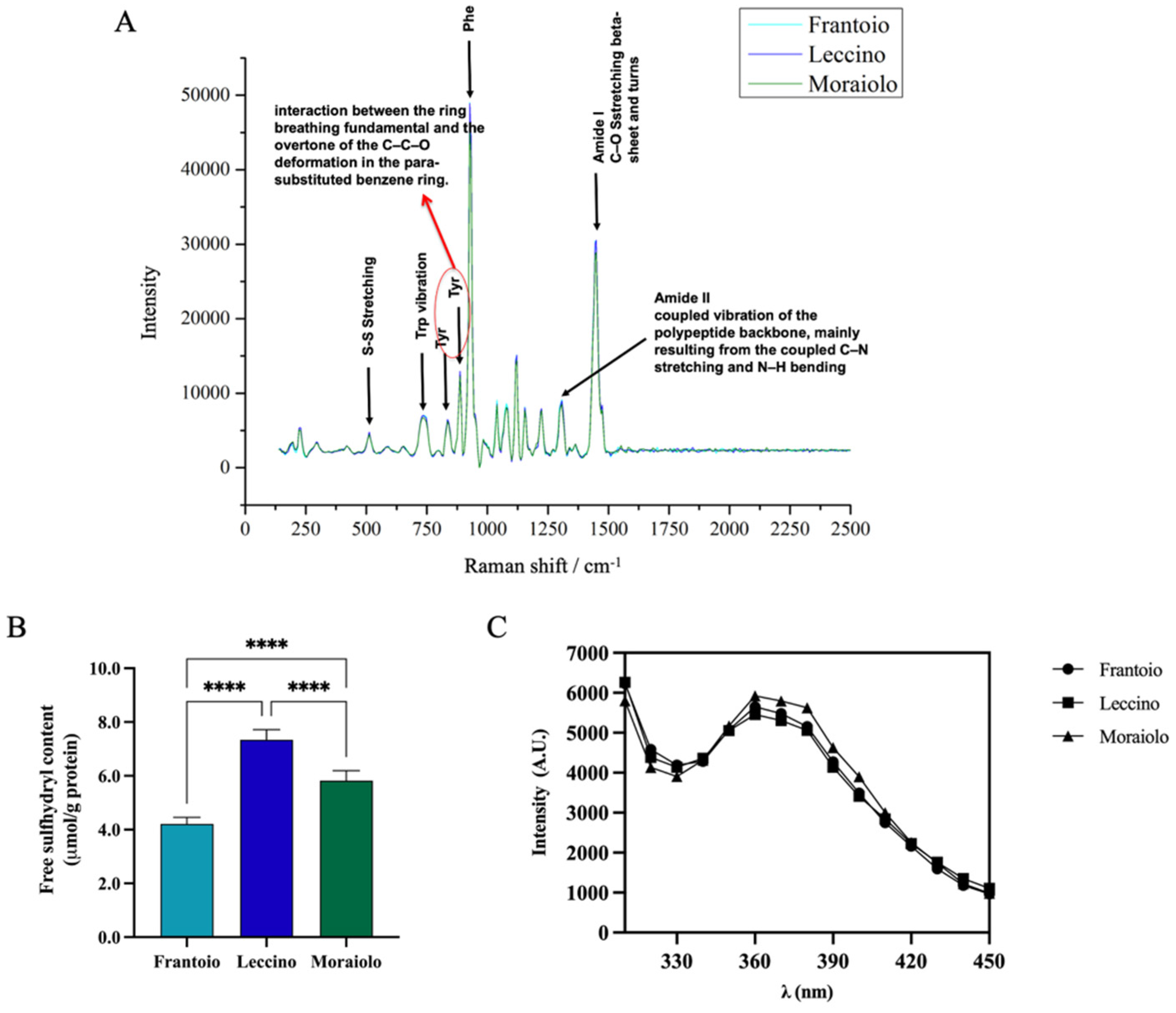
2.4. Free-Sulfhydryl Group Determination
2.5. Intrinsic Fluorescence Spectroscopy
2.6. Raman Spectroscopy
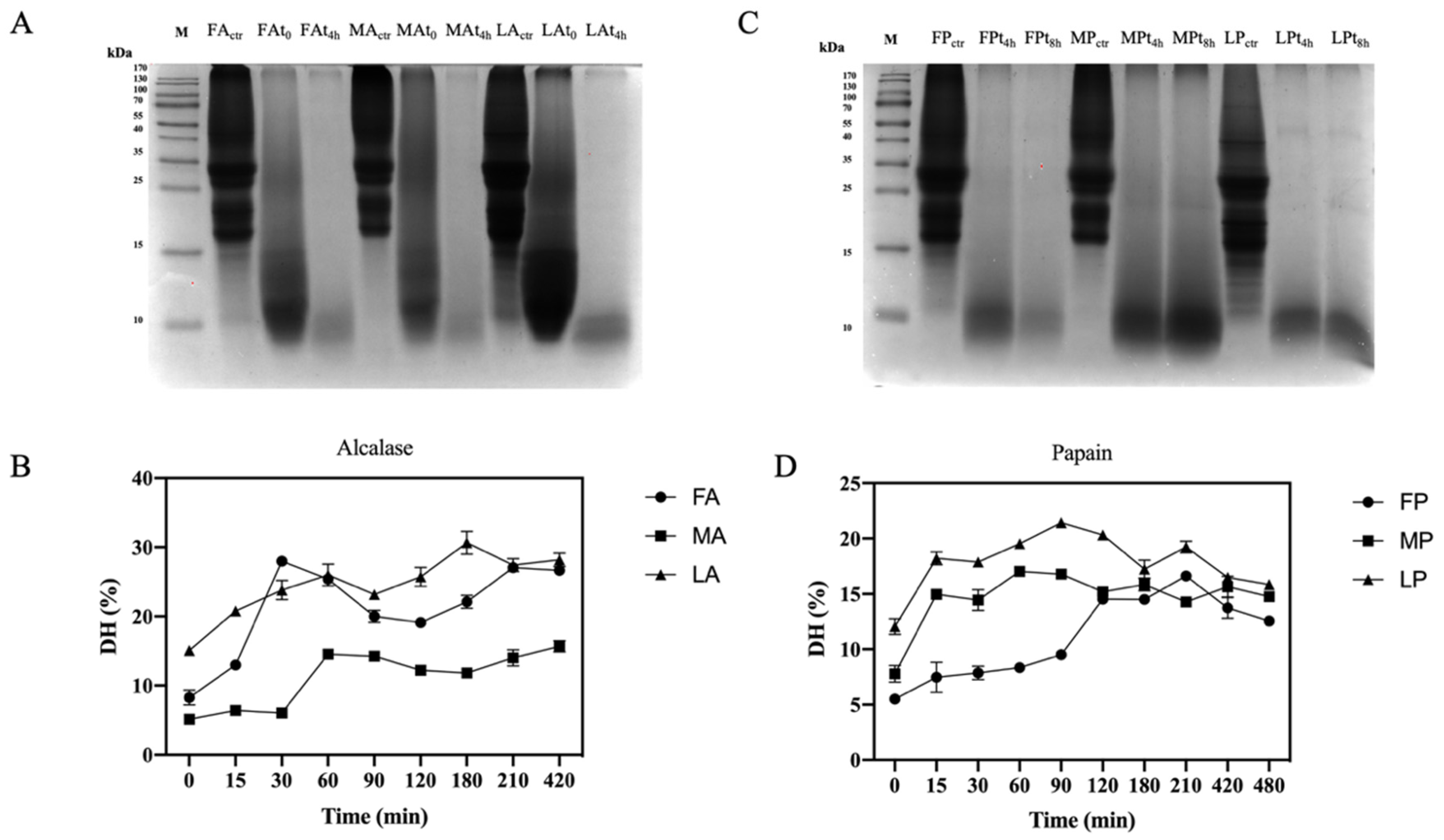
2.7. Olive Seed Protein Hydrolysis for Releasing Bioactive peptides
2.8. Short Peptide Purification and Analysis by High-Performance Liquid Chromatography–High-Resolution Mass Spectrometry
2.9. Profile of Potential Biological Activities and Peptide Ranking
2.10. Cell Culture
2.11. 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) Assay
2.12. Antioxidant Activity of Olive Seed Hydrolysates
2.12.1. Diphenyl-2-Picrylhydrazyl Radical (DPPH) Assay
2.12.2. 2,2′-Azino-Bis(3-Ethylbenzothiazoline-6-Sulfonic Acid) Diammonium Salt Assay
2.12.3. FRAP Assay
2.12.4. Fluorometric Intracellular ROS Assay
2.12.5. Lipid Peroxidation (MDA) Assay
2.13. Antidiabetic Activity of Olive Seed Hydrolysates
2.13.1. In Vitro Measurement of the DPP-IV Inhibitory Activity
2.13.2. Evaluation of the Inhibitory Effect of Olive Seed Hydrolysates on Cellular DPP-IV Activity
2.13.3. Evaluation of the GLP-1 Stability and Secretion at Cellular Level
2.14. Statically Analysis
3. Results
3.1. Extraction of Olive Seed Proteins
3.2. Structural Properties Characterization: Raman Spectroscopy, Free-Sulfhydryl Group (SH) Content, and Intrinsic Fluorescence (IF)
3.3. Protein Solubility (PS), Water-Binding Capacity (WBC), and Oil-Biding Capacity (OBC) of Extracted Proteins
3.4. Production of Olive Seed Protein Hydrolysates Using Alcalase and Papain
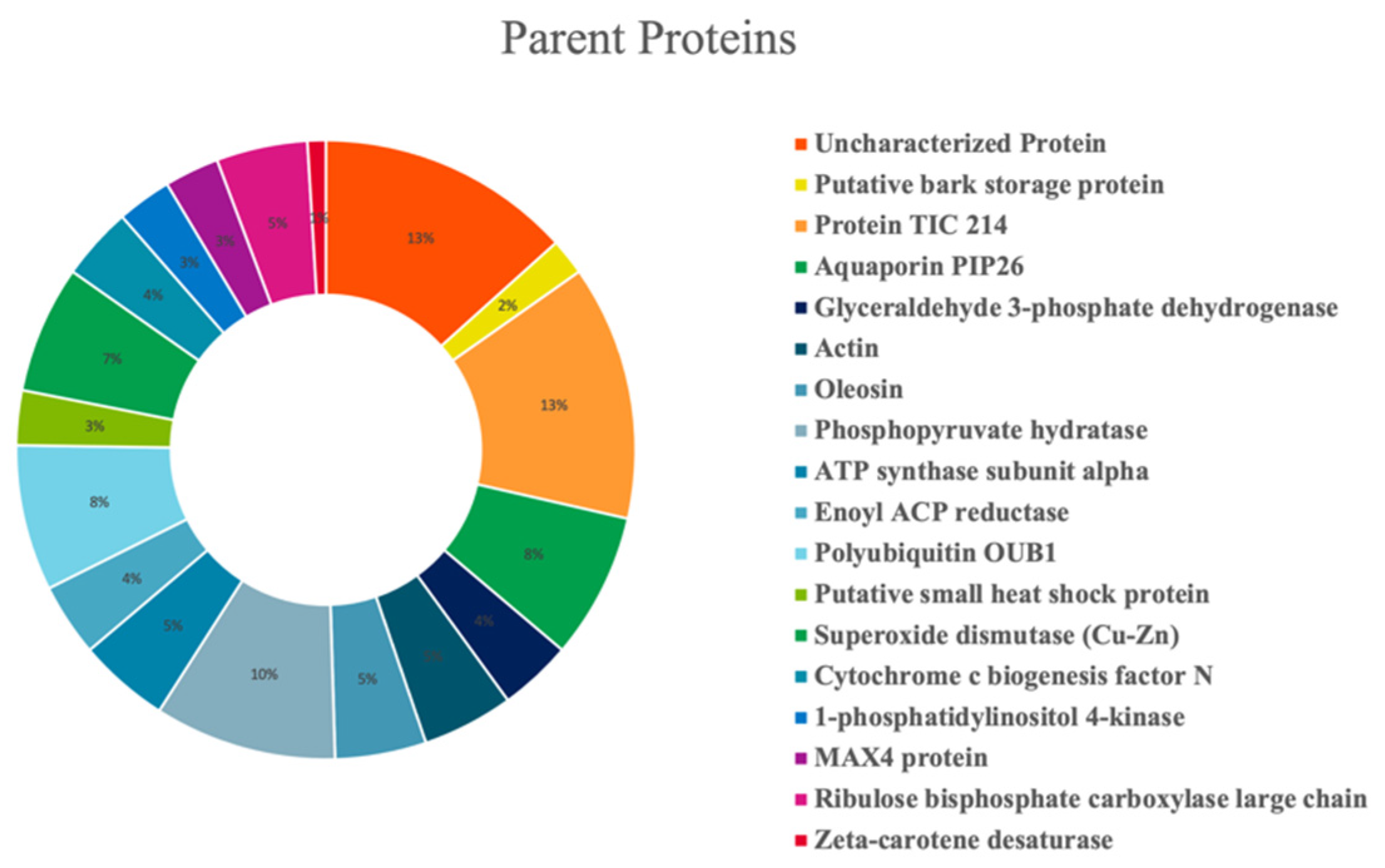
3.5. Peptidomics Characterization of Olive Seed Hydrolysates: Analysis of Medium- and Short-Sized Peptides
3.6. Antioxidant Activity of Olive Seed Hydrolysates
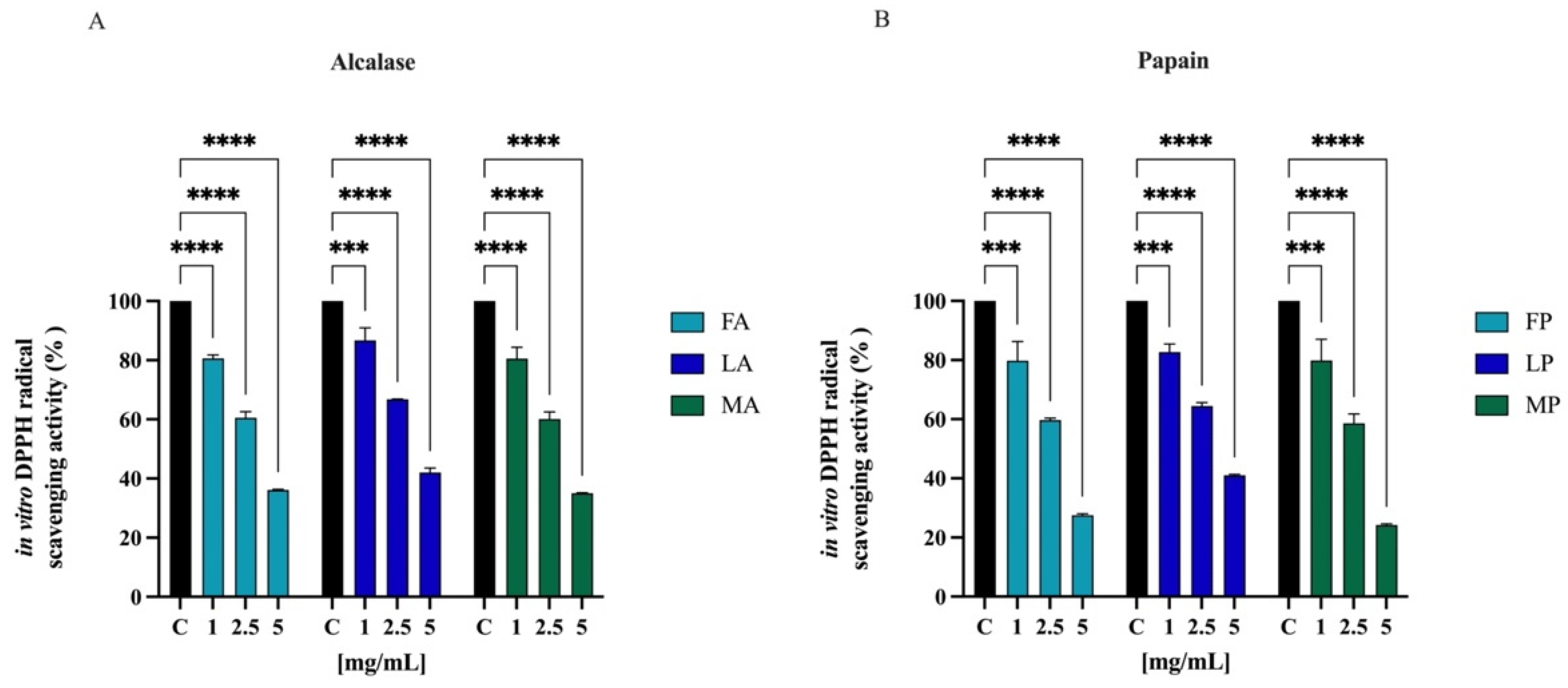
3.6.1. Direct Radical Scavenging Activity of Olive Seed Hydrolysates by DPPH Assay
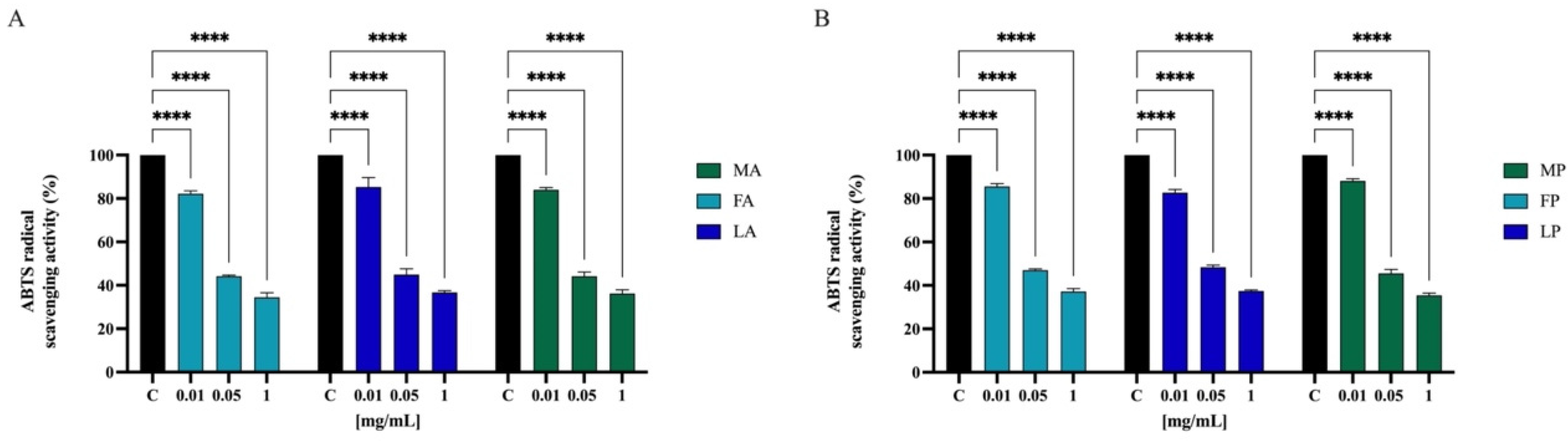
3.6.2. Direct Radical Scavenging Activity of Olive Seed Hydrolysates by ABTS Assay
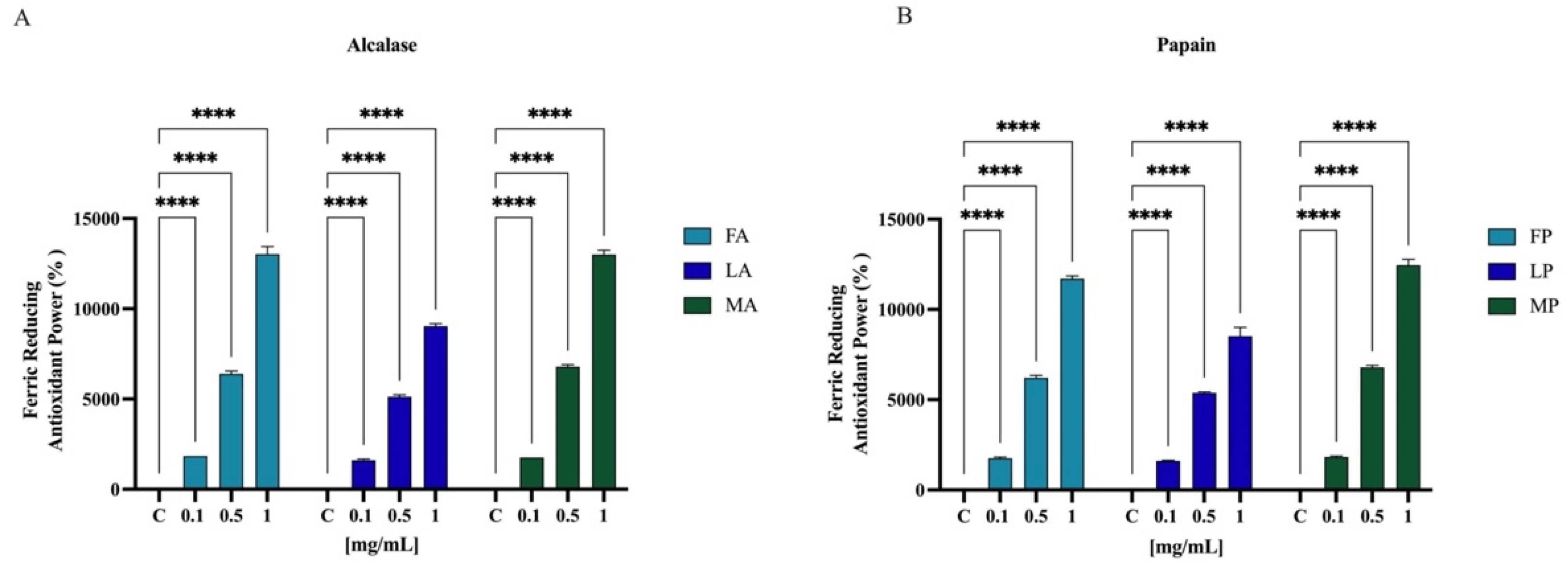
3.6.3. Ferric-Reducing Antioxidant Power (FRAP) Activity
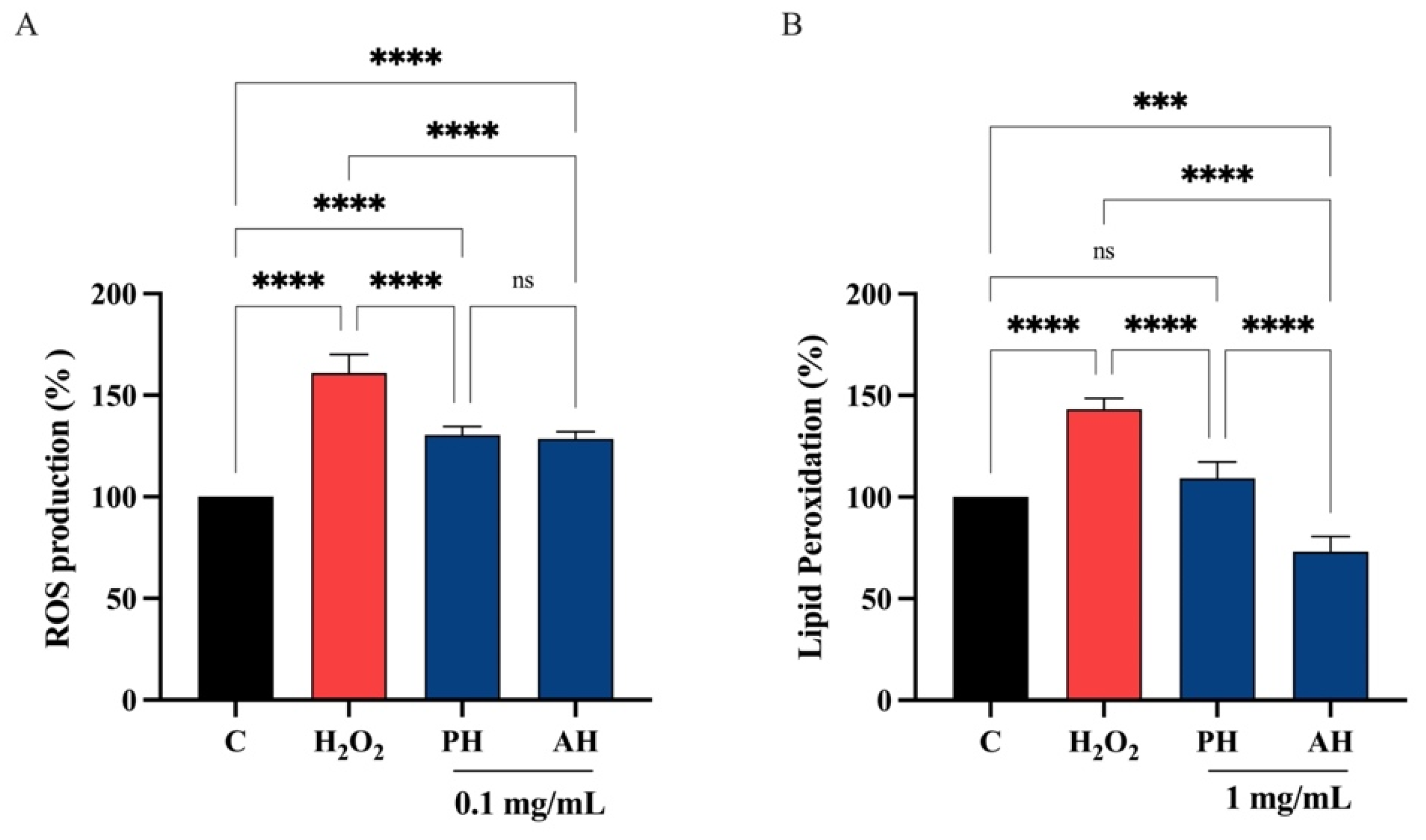
3.6.4. PH and AH Hydrolysates Decrease the H2O2-Induced ROS and Lipid Peroxidation Levels in Intestinal Caco-2 Cells
3.7. Antidiabetic Activity of Olive Seed Hydrolysates
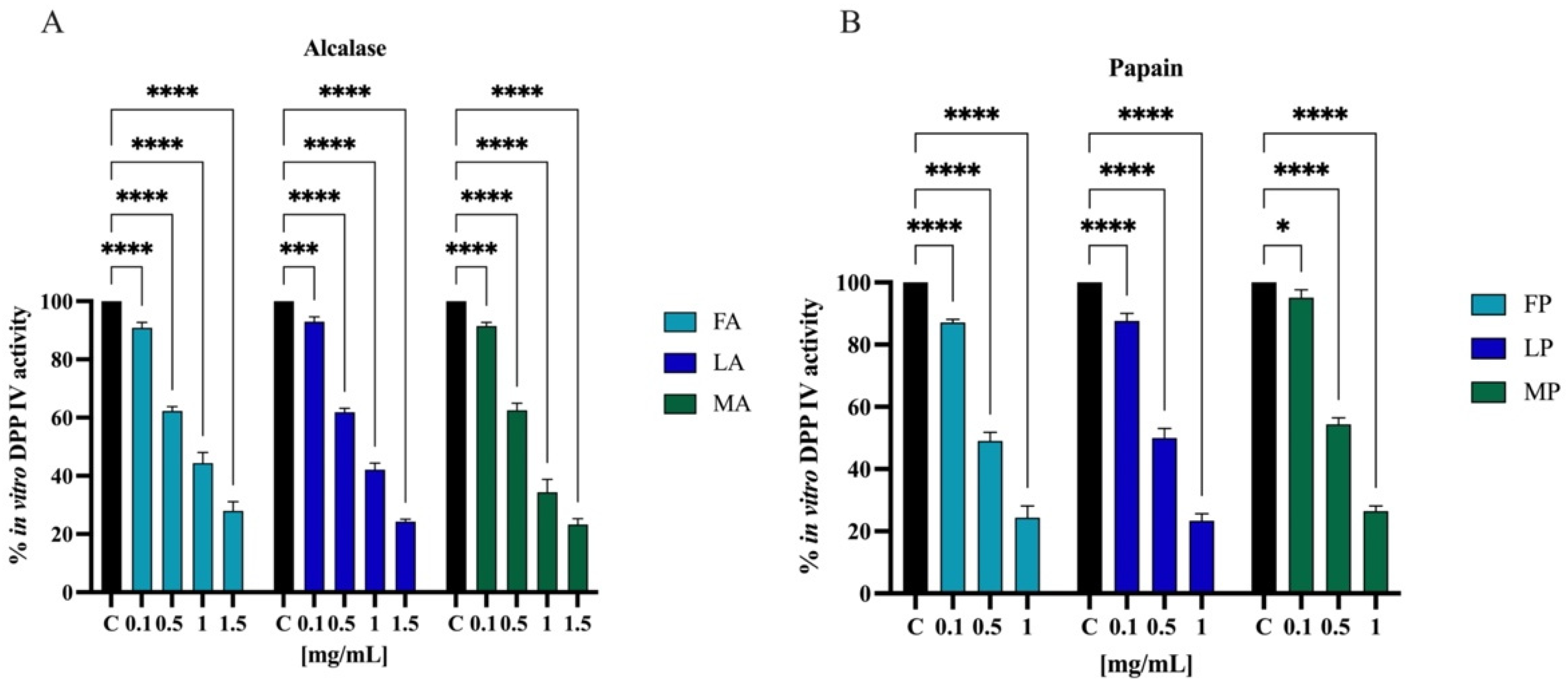
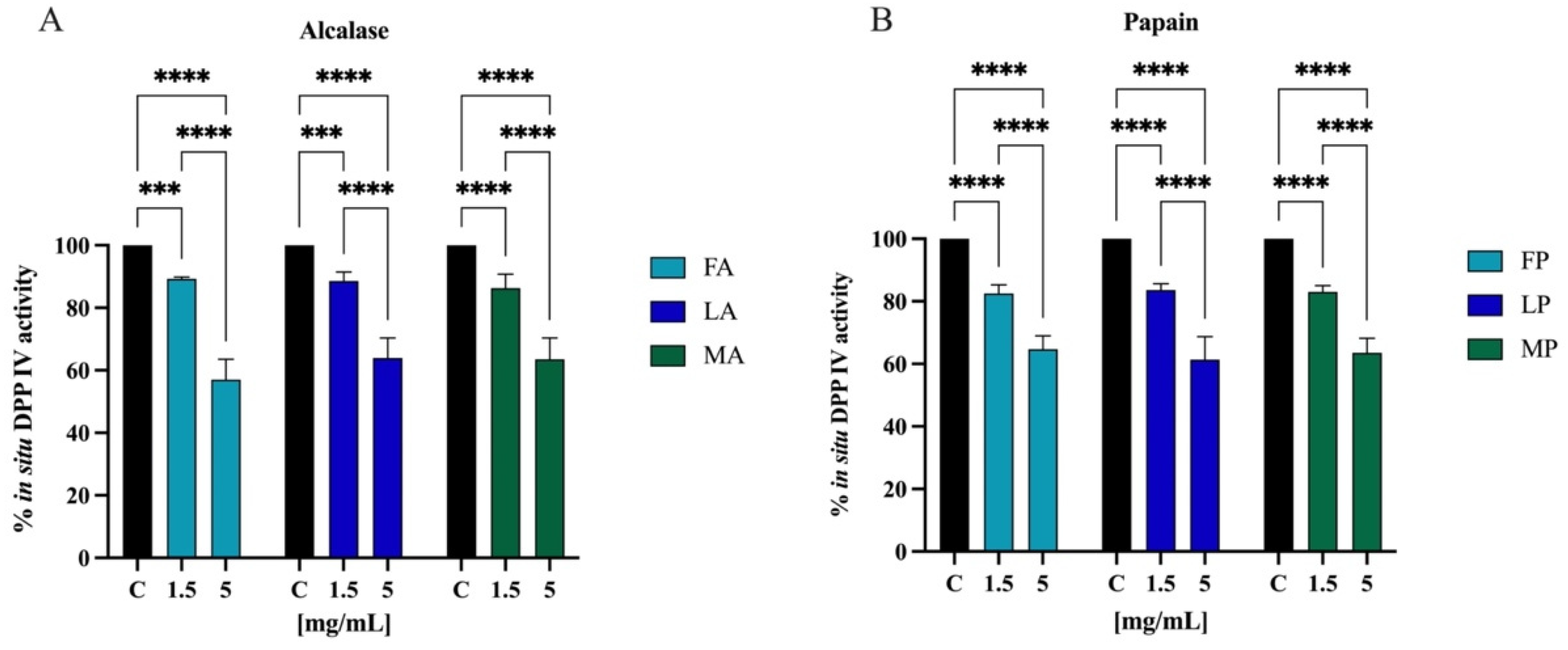
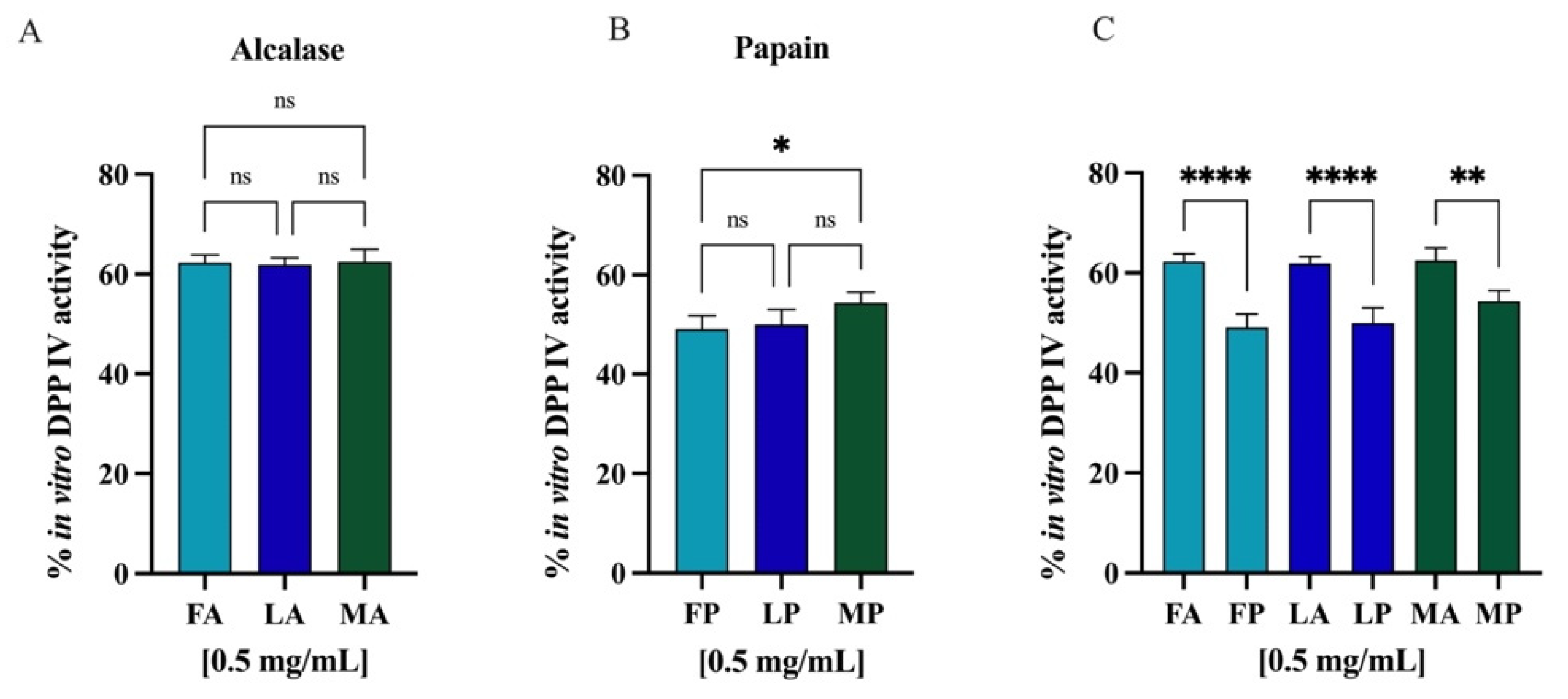
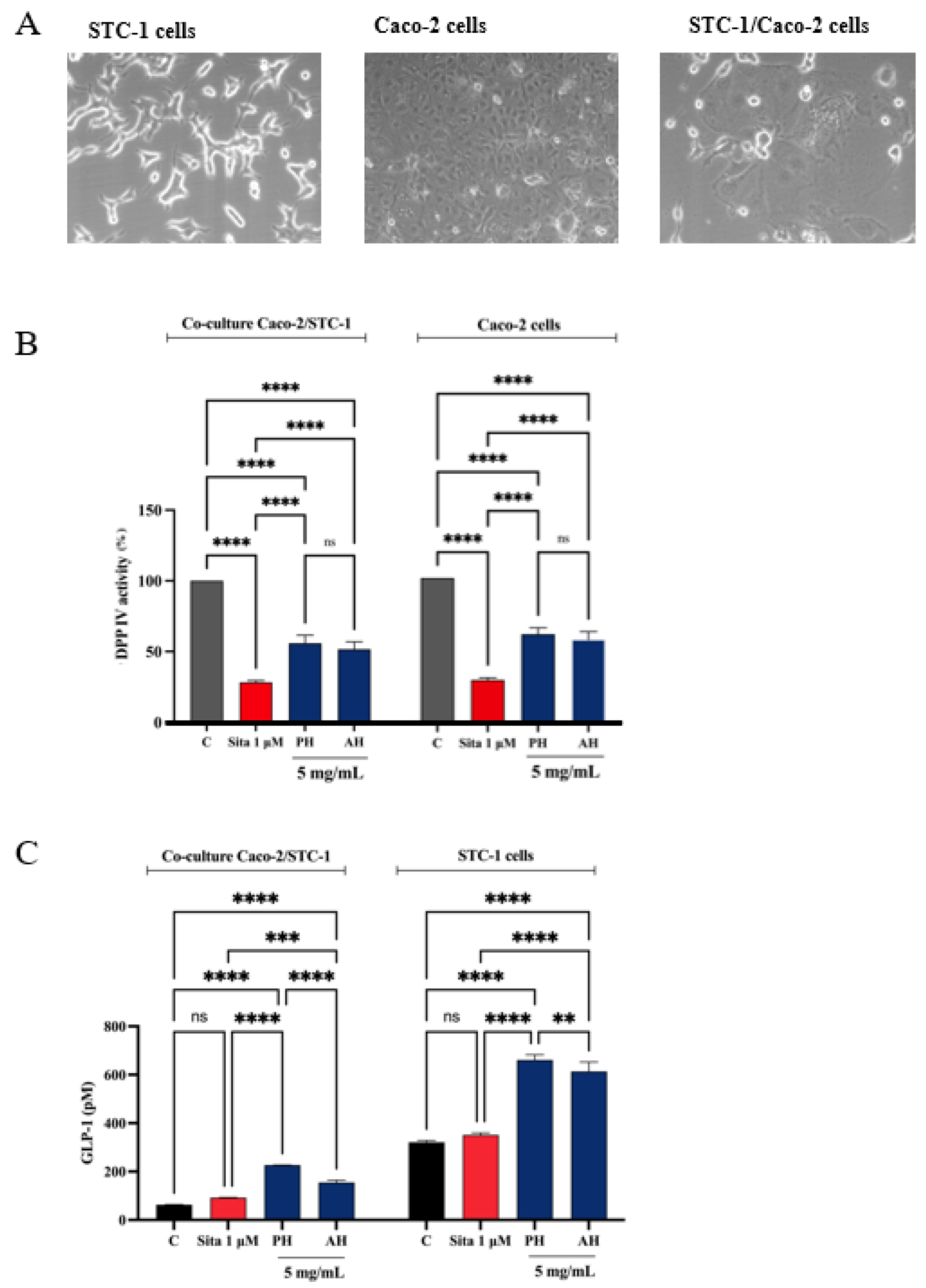
3.7.1. In Vitro and Cellular DPP-IV Inhibitory Activity of Olive Seed Hydrolysates
3.7.2. STC-1/Caco-2 Co-Culture System and Assessment of Olive Seed Hydrolysates Capacity to Modulate Stability and Secretion of GLP-1 Hormone
4. Discussion
4.1. Olive Seed Hydrolysates Exert Direct and Cellular Antioxidant Activity
4.2. Olive Seed Hydrolysates Inhibit DPP-IV Activity and Improve the Stability and Secretion of GLP-1
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguilar-Toalá, J.E.; Santiago-López, L.; Peres, C.M.; Peres, C.; Garcia, H.S.; Vallejo-Cordoba, B.; González-Córdova, A.F.; Hernández-Mendoza, A. Assessment of Multifunctional Activity of Bioactive Peptides Derived from Fermented Milk by Specific Lactobacillus Plantarum Strains. J. Dairy Sci. 2017, 100, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Cabizza, R.; Fancello, F.; Petretto, G.L.; Addis, R.; Pisanu, S.; Pagnozzi, D.; Piga, A.; Urgeghe, P.P. Exploring the DPP-IV Inhibitory, Antioxidant and Antibacterial Potential of Ovine “Scotta” Hydrolysates. Foods 2021, 10, 3137. [Google Scholar] [CrossRef] [PubMed]
- Eckert, E.; Zambrowicz, A.; Pokora, M.; Setner, B.; Dabrowska, A.; Szołtysik, M.; Szewczuk, Z.; Polanowski, A.; Trziszka, T.; Chrzanowska, J. Egg-Yolk Protein by-Product as a Source of ACE-Inhibitory Peptides Obtained with Using Unconventional Proteinase from Asian Pumpkin (Cucurbita Ficifolia). J. Proteom. 2014, 110, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Ferri, M.; Graen-Heedfeld, J.; Bretz, K.; Guillon, F.; Michelini, E.; Calabretta, M.M.; Lamborghini, M.; Gruarin, N.; Roda, A.; Kraft, A.; et al. Peptide Fractions Obtained from Rice By-Products by Means of an Environment-Friendly Process Show in Vitro Health-Related Bioactivities. PLoS ONE 2017, 12, 1–14. [Google Scholar] [CrossRef]
- Jin, R.; Teng, X.; Shang, J.; Wang, D.; Liu, N. Identification of Novel DPP–IV Inhibitory Peptides from Atlantic Salmon (Salmo Salar) Skin. Food Res. Int. 2020, 133, 109161. [Google Scholar] [CrossRef]
- Lammi, C.; Bollati, C.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A. Soybean-and Lupin-Derived Peptides Inhibit DPP-IV Activity on in Situ Human Intestinal Caco-2 Cells and Ex Vivo Human Serum. Nutrients 2018, 10, 1082. [Google Scholar] [CrossRef]
- Lima, R.D.C.L.; Berg, R.S.; Rønning, S.B.; Afseth, N.K.; Knutsen, S.H.; Staerk, D.; Wubshet, S.G. Peptides from Chicken Processing By-Product Inhibit DPP-IV and Promote Cellular Glucose Uptake: Potential Ingredients for T2D Management. Food Funct. 2019, 10, 1619–1628. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Espejo-Carpio, F.J.; Guadix, E.M. Production and Identification of Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Peptides from Discarded Sardine Pilchardus Protein. Food Chem. 2020, 328, 127096. [Google Scholar] [CrossRef]
- Zamora-sillero, J.; Gharsallaoui, A.; Prentice, C.; Prentice, C. Peptides From Fish By-Product Protein Hydrolysates and Its Functional Properties: An Overview. Mar. Biotechnol. 2018, 20, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Socas-Rodríguez, B.; Álvarez-Rivera, G.; Valdés, A.; Ibáñez, E.; Cifuentes, A. Food By-Products and Food Wastes: Are They Safe Enough for Their Valorization? Trends Food Sci. Technol. 2021, 114, 133–147. [Google Scholar] [CrossRef]
- Power, O.; Jakeman, P.; Fitzgerald, R.J. Antioxidative Peptides: Enzymatic Production, in Vitro and in Vivo Antioxidant Activity and Potential Applications of Milk-Derived Antioxidative Peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jin, Y.; Lin, S.; Jones, G.S.; Chen, F. Purification and Identification of Novel Antioxidant Peptides from Egg White Protein and Their Antioxidant Activities. Food Chem. 2015, 175, 258–266. [Google Scholar] [CrossRef]
- De Campos Zani, S.C.; Wu, J.; Chan, C.B. Egg and Soy-Derived Peptides and Hydrolysates: A Review of Their Physiological Actions against Diabetes and Obesity. Nutrients 2018, 10, 549. [Google Scholar] [CrossRef] [PubMed]
- Rudich, A.; Tlrosh, A.; Potashnik, R.; Hemi, R.; Kanety, H.; Bashan, N. Prolonged Oxidative Stress Impairs Insulin-Induced GLUT4 Translocation in 3T3-L1 Adipocytes. Diabetes 1998, 47, 1562–1569. [Google Scholar] [CrossRef]
- Maddux, B.A.; See, W.; Lawrence, J.C.; Goldfine, A.L.; Goldfine, I.D.; Evans, J.L. Protection against Oxidative Stress-Induced Insulin Resistance in Rat L6 Muscle Cells by Micromolar Concentrations of α-Lipoic Acid. Diabetes 2001, 50, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A.; Testa, R.; Genovese, S. Clinical Implications of Oxidative Stress and Potential Role of Natural Antioxidants in Diabetic Vascular Complications. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 285–292. [Google Scholar] [CrossRef]
- Harding, J.L.; Pavkov, M.E.; Magliano, D.J.; Shaw, J.E.; Gregg, E.W. Global Trends in Diabetes Complications: A Review of Current Evidence. Diabetologia 2019, 62, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Dicker, D. DPP-4 Inhibitors: Impact on Glycemic Control and Cardiovascular Risk Factors. Diabetes Care 2011, 34, S276–S278. [Google Scholar] [CrossRef] [PubMed]
- Barnett, A. DPP-4 Inhibitors and Their Potential Role in the Management of Type 2 Diabetes. Int. J. Clin. Pract. 2006, 60, 1454–1470. [Google Scholar] [CrossRef] [PubMed]
- Zambrowicz, A.; Eckert, E.; Pokora, M.; Bobak, Ł.; Da¸browska, A.; Szołtysik, M.; Trziszka, T.; Chrzanowska, J. Antioxidant and Antidiabetic Activities of Peptides Isolated from a Hydrolysate of an Egg-Yolk Protein by-Product Prepared with a Proteinase from Asian Pumpkin (Cucurbita Ficifolia). RSC Adv. 2015, 5, 10460–10467. [Google Scholar] [CrossRef]
- Saidi, S.; Saoudi, M.; Ben Amar, R. Valorisation of Tuna Processing Waste Biomass: Isolation, Purification and Characterisation of Four Novel Antioxidant Peptides from Tuna by-Product Hydrolysate. Environ. Sci. Pollut. Res. 2018, 25, 17383–17392. [Google Scholar] [CrossRef] [PubMed]
- Choksawangkarn, W.; Phiphattananukoon, S.; Jaresitthikunchai, J.; Roytrakul, S. Antioxidative Peptides from Fish Sauce By-Product: Isolation and Characterization. Agric. Nat. Resour. 2018, 52, 460–466. [Google Scholar] [CrossRef]
- Ibáñez, E.; Palacios, J.; Señoráns, F.J.; Santa-María, G.; Tabera, J.; Reglero, G. Isolation and Separation of Tocopherols from Olive By-Products with Supercritical Fluids. JAOCS J. Am. Oil Chem. Soc. 2000, 77, 187–190. [Google Scholar] [CrossRef]
- Wang, B.; Qu, J.; Luo, S.; Feng, S.; Li, T.; Yuan, M.; Huang, Y.; Liao, J.; Yang, R.; Ding, C. Optimization of Ultrasound-Assisted Extraction of Flavonoids from Olive (Olea Europaea) Leaves, and Evaluation of Their Antioxidant and Anticancer Activities. Molecules 2018, 23, 2513. [Google Scholar] [CrossRef] [PubMed]
- Stathopoulos, P.; Koutra, C.; Bata, E.; Skaltsounis, A.-L. Optimization of Oleuropein Extraction from Olive Leaves. Alternative Approaches for the Recovery of Olive Bioactive Secoiridoids. Planta Med. 2021, 87, PC4–PC35. [Google Scholar] [CrossRef]
- Vilaplana-Pérez, C.; Auñón, D.; García-Flores, L.A.; Gil-Izquierdo, A. Hydroxytyrosol and Potential Uses in Cardiovascular Diseases, Cancer, and AIDS. Front. Nutr. 2014, 1, 18. [Google Scholar] [CrossRef]
- Bertelli, M.; Kiani, A.K.; Paolacci, S.; Manara, E.; Kurti, D.; Dhuli, K.; Bushati, V.; Miertus, J.; Pangallo, D.; Baglivo, M.; et al. Hydroxytyrosol: A Natural Compound with Promising Pharmacological Activities. J. Biotechnol. 2020, 309, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Pedroza, M.A.; Amendola, D.; Maggi, L.; Zalacain, A.; De Faveri, D.M.; Spigno, G. Microwave-Assisted Extraction of Phenolic Compounds from Dried Waste Grape Skins. Int. J. Food Eng. 2015, 11, 359–370. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Matsui, T.; Maeda, S.; Higashimoto, Y.; Yamagishi, S. ichi Advanced Glycation End Products Evoke Endothelial Cell Damage by Stimulating Soluble Dipeptidyl Peptidase-4 Production and Its Interaction with Mannose 6-Phosphate/Insulin-like Growth Factor II Receptor. Cardiovasc. Diabetol. 2013, 12, 125. [Google Scholar] [CrossRef] [PubMed]
- Malomo, S.A.; He, R.; Aluko, R.E. Structural and Functional Properties of Hemp Seed Protein Products. J. Food Sci. 2014, 79, 1512–1521. [Google Scholar] [CrossRef] [PubMed]
- Aiello, G.; Pugliese, R.; Rueller, L.; Bollati, C.; Bartolomei, M.; Li, Y.; Robert, J.; Arnoldi, A.; Lammi, C. Assessment of the Physicochemical and Conformational Changes of Ultrasound-Driven Proteins Extracted from Soybean Okara Byproduct. Foods 2021, 10, 562. [Google Scholar] [CrossRef] [PubMed]
- Aiello, G.; Lammi, C.; Boschin, G.; Zanoni, C.; Arnoldi, A. Exploration of Potentially Bioactive Peptides Generated from the Enzymatic Hydrolysis of Hempseed Proteins. J. Agric. Food Chem. 2017, 65, 10174–10184. [Google Scholar] [CrossRef] [PubMed]
- Piovesana, R.; Faroni, A.; Magnaghi, V.; Reid, A.J.; Tata, A.M. M2 Receptors Activation Modulates Cell Growth, Migration and Differentiation of Rat Schwann-like Adipose-Derived Stem Cells. Cell Death Discov. 2019, 5, 92. [Google Scholar] [CrossRef] [PubMed]
- Aiello, G.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A.; Vistoli, G.; Lammi, C. Behavior of Three Hypocholesterolemic Peptides from Soy Protein in an Intestinal Model Based on Differentiated Caco-2 Cell. J. Funct. Foods 2018, 45, 363–370. [Google Scholar] [CrossRef]
- Lammi, C.; Zanoni, C.; Scigliuolo, G.M.; D’Amato, A.; Arnoldi, A. Lupin Peptides Lower Low-Density Lipoprotein (LDL) Cholesterol through an up-Regulation of the LDL Receptor/Sterol Regulatory Element Binding Protein 2 (SREBP2) Pathway at HepG2 Cell Line. J. Agric. Food Chem. 2014, 62, 7151–7159. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Peptides Derived from Soy and Lupin Protein as Dipeptidyl-Peptidase IV Inhibitors: In Vitro Biochemical Screening and in Silico Molecular Modeling Study. J. Agric. Food Chem. 2016, 64, 9601–9606. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Bollati, C.; Gelain, F.; Arnoldi, A.; Pugliese, R. Enhancement of the Stability and Anti-DPPIV Activity of Hempseed Hydrolysates through Self-Assembling Peptide-Based Hydrogels. Front. Chem. 2019, 6, 670. [Google Scholar] [CrossRef] [PubMed]
- Suda, C.N.K.; Giorgini, J.F. Seed Reserve Composition and Mobilization during Germination and Initial Seedling Development of Euphorbia Heterophylla. Rev. Bras. Fisiol. Veg. 2000, 12, 226–244. [Google Scholar] [CrossRef]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed Storage Proteins: Structures and Biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar] [CrossRef] [PubMed]
- De Dios Alché, J.; Jiménez-López, J.C.; Wang, W.; Castro-López, A.J.; Rodríguez-García, M.I. Biochemical Characterization and Cellular Localization of 11S Type Storage Proteins in Olive (Olea Europaea L.) Seeds. J. Agric. Food Chem. 2006, 54, 5562–5570. [Google Scholar] [CrossRef]
- Wu, M.; Li, Z.; Wei, R.; Luan, Y.; Hu, J.; Wang, Q.; Liu, R.; Ge, Q.; Yu, H. Role of Disulfide Bonds and Sulfhydryl Blocked by N-Ethylmaleimide on the Properties of Different Protein-Stabilized Emulsions. Foods 2021, 10, 3079. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; De Dios Alché, J.; Rodríguez-García, M.I. Characterization of Olive Seed Storage Proteins. Acta Physiol. Plant. 2007, 29, 439–444. [Google Scholar] [CrossRef]
- Haque, M.A.; Timilsena, Y.P.; Adhikari, B. Food Proteins, Structure, and Function. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Ulloa, J.A.; Villalobos Barbosa, M.C.; Resendiz Vazquez, J.A.; Rosas Ulloa, P.; Ramírez Ramírez, J.C.; Silva Carrillo, Y.; González Torres, L. Producción y Caracterización Fisico-Química y Functional de Un Aislado Proteínico de Semillas de Jaca (Artocarpus Heterophyllus). CYTA - J. Food 2017, 15, 497–507. [Google Scholar] [CrossRef]
- Kanu, P.J.; Kanu, J.B.; Sandy, E.H.; Kandeh, J.B.A.; Mornya, P.M.P.; Huiming, Z. Optimization of Enzymatic Hydrolysis of Defatted Sesame Flour by Different Proteases and Their Effect on the Functional Properties of the Resulting Protein Hydrolysate. Am. J. Food Technol. 2009, 4, 226–240. [Google Scholar] [CrossRef]
- Cerrato, A.; Aita, S.E.; Capriotti, A.L.; Cavaliere, C.; Montone, C.M.; Laganà, A.; Piovesana, S. A New Opening for the Tricky Untargeted Investigation of Natural and Modified Short Peptides. Talanta 2020, 219, 121262. [Google Scholar] [CrossRef] [PubMed]
- Mishra, K.; Ojha, H.; Chaudhury, N.K. Estimation of Antiradical Properties of Antioxidants Using DPPH- Assay: A Critical Review and Results. Food Chem. 2012, 130, 1036–1043. [Google Scholar] [CrossRef]
- Cruz-Casas, D.E.; Aguilar, C.N.; Ascacio-Valdés, J.A.; Rodríguez-Herrera, R.; Chávez-González, M.L.; Flores-Gallegos, A.C. Enzymatic Hydrolysis and Microbial Fermentation: The Most Favorable Biotechnological Methods for the Release of Bioactive Peptides. Food Chem. Mol. Sci. 2021, 3, 1000047. [Google Scholar] [CrossRef] [PubMed]
- Ahmadifard, N.; Murueta, J.H.C.; Abedian-Kenari, A.; Motamedzadegan, A.; Jamali, H. Comparison the Effect of Three Commercial Enzymes for Enzymatic Hydrolysis of Two Substrates (Rice Bran Protein Concentrate and Soy-Been Protein) with SDS-PAGE. J. Food Sci. Technol. 2016, 53, 1279–1284. [Google Scholar] [CrossRef]
- Piovesana, S.; Montone, C.M.; Cavaliere, C.; Crescenzi, C.; La Barbera, G.; Laganà, A.; Capriotti, A.L. Sensitive Untargeted Identification of Short Hydrophilic Peptides by High Performance Liquid Chromatography on Porous Graphitic Carbon Coupled to High Resolution Mass Spectrometry. J. Chromatogr. A 2019, 1590, 73–79. [Google Scholar] [CrossRef]
- Koskinen, V.R.; Emery, P.A.; Creasy, D.M.; Cottrell, J.S. Hierarchical Clustering of Shotgun Proteomics Data. Mol. Cell. Proteomics 2011, 10, 1–12. [Google Scholar] [CrossRef]
- Kim, D.O.; Lee, K.W.; Lee, H.J.; Lee, C.Y. Vitamin C Equivalent Antioxidant Capacity (VCEAC) of Phenolic Phytochemicals. J. Agric. Food Chem. 2002, 50, 3713–3717. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Fractionation and Identification of Antioxidant Peptides from an Enzymatically Hydrolysed Palmaria Palmata Protein Isolate. Food Res. Int. 2017, 100, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, I.D.; Aluko, R.E. Structural and Functional Properties of Food Protein-derived Antioxidant Peptides. J. Food Biochem. 2019, 43, e12761. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, I.D.; Aluko, R.E. Antioxidant Properties of Flaxseed Protein Hydrolysates: Influence of Hydrolytic Enzyme Concentration and Peptide Size. JAOCS, J. Am. Oil Chem. Soc. 2018, 95, 1105–1118. [Google Scholar] [CrossRef]
- Bollati, C.; Cruz-Chamorro, I.; Aiello, G.; Li, J.; Bartolomei, M.; Santos-Sánchez, G.; Ranaldi, G.; Ferruzza, S.; Sambuy, Y.; Arnoldi, A.; et al. Investigation of the Intestinal Trans-Epithelial Transport and Antioxidant Activity of Two Hempseed Peptides WVSPLAGRT (H2) and IGFLIIWV (H3). Food Res. Int. 2022, 152, 110720. [Google Scholar] [CrossRef] [PubMed]
- Esteve, C.; Marina, M.L.; García, M.C. Novel Strategy for the Revalorization of Olive (Olea Europaea) Residues Based on the Extraction of Bioactive Peptides. Food Chem. 2015, 167, 272–280. [Google Scholar] [CrossRef]
- Lammi, C.; Bollati, C.; Arnoldi, A. Antioxidant Activity of Soybean Peptides on Human Hepatic HepG2 Cells. J. Food Bioact. 2019. [Google Scholar] [CrossRef]
- Girgih, A.T.; Udenigwe, C.C.; Aluko, R.E. In Vitro Antioxidant Properties of Hemp Seed (Cannabis Sativa L.) Protein Hydrolysate Fractions. JAOCS, J. Am. Oil Chem. Soc. 2011, 88, 381–389. [Google Scholar] [CrossRef]
- Wattanasiritham, L.; Kubglomsong, S.; Theerakulkait, C. Antioxidant Activity of Rice Bran Protein Extract, Its Enzymatic Hydrolysates and Its Combination with Commercial Antioxidants. Pakistan J. Nutr. 2015, 14, 647–652. [Google Scholar] [CrossRef]
- Centenaro, G.S.; Mellado, M.S.; Prentice-Hernández, C. Antioxidant Activity of Protein Hydrolysates of Fish and Chicken Bones. Adv. J. Food Sci. Technol. 2011, 3, 282–288. [Google Scholar]
- Han, R.; Hernández Álvarez, A.J.; Maycock, J.; Murray, B.S.; Boesch, C. Comparison of Alcalase- and Pepsin-Treated Oilseed Protein Hydrolysates—Experimental Validation of Predicted Antioxidant, Antihypertensive and Antidiabetic Properties. Curr. Res. Food Sci. 2021, 4, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zheng, L.; Huang, M.; Zhao, M. Exploring Structural Features of Potent Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Peptides Derived from Tilapia (Oreochromis Niloticus) Skin Gelatin by an Integrated Approach of Multivariate Analysis and Gly-Pro-Based Peptide Library. Food Chem. 2022, 397, 133821. [Google Scholar] [CrossRef] [PubMed]
- Mentlein, R. Cell-Surface Peptidases. Int. Rev. Cytol. 2004, 235, 165–213. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Aiello, G.; Bollati, C.; Bartolomei, M.; Arnoldi, A.; Lammi, C. Phycobiliproteins from Arthrospira Platensis (Spirulina): A New Source of Peptides with Dipeptidyl Peptidase-IV Inhibitory Activity. Nutrients 2020, 13, 1624. [Google Scholar] [CrossRef] [PubMed]
- Aiello, G.; Li, Y.; Boschin, G.; Bollati, C.; Arnoldi, A.; Lammi, C. Chemical and Biological Characterization of Spirulina Protein Hydrolysates: Focus on ACE and DPP-IV Activities Modulation. J. Funct. Foods 2019, 63, 103592. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Temperature (°C) | Hydrolysis Time | Enzyme: Substrate Ratio | pH |
|---|---|---|---|---|
| Alcalase | 50 | 4 h | 0.15 UA/g | 8.5 |
| Papain | 65 | 8 h | 100 UA/g | 7 |
| ID | Extraction Buffer | (mg/mL) |
|---|---|---|
| U | UREA 8 M, 1% CHAPS, 0.1% DTT, NH4HCO3 | 9.2 |
| T | 0.1 M Tris (pH 7.5), 0.5 M NaCl, 0.5% SDS, 0.1% DTT | 1.9 |
| U/T | UREA 6 M, 0.1 M Tris-HCl (pH 8), 0.5 M NaCl, 0.5% SDS, 0.1% DTT | 5.3 |
| Cultivar | DH % Alcalase Digestion | DH % Papain Digestion |
|---|---|---|
| Frantoio | 26.9% | 12.5% |
| Leccino | 27.5% | 15.7% |
| Moraiolo | 15.3% | 14.7% |
| Score a | Sequence | Potential Bioactive Peptides b | Biological Functions c |
|---|---|---|---|
| 0.998974 | FF | FF | DPP IV inhibitor |
| 0.996643 | MF | MF | DPP IV inhibitor |
| 0.994712 | GF | GF | DPP IV inhibitor, |
| 0.994084 | GGFF | GF, GG, FF | DPP IV inhibitor |
| GF | DPP III inhibitor | ||
| 0.993120 | FFJ | FF | DPP IV inhibitor |
| 0.991656 | WPM | WP, PM | DPP IV inhibitor |
| 0.989681 | VY | VY | DPP IV inhibitor, antioxidant |
| 0.987345 | GGF | GF, GG | DPP IV inhibitor |
| GF | DPP III inhibitor | ||
| 0.985719 | FR | FR | DPP IV inhibitor |
| 0.977983 | WJAF | AF | DPP IV inhibitor |
| 0.977525 | FGQW | QW | DPP IV inhibitor |
| 0.974885 | WY | WY | DPP IV inhibitor, antioxidant |
| 0.968518 | SMF | MF | DPP IV inhibitor |
| 0.962442 | FFJA | FF | DPP IV inhibitor |
| 0.954571 | FNFJ | NF, FN | DPP IV inhibitor |
| 0.952926 | HW | HW | DPP IV inhibitor |
| 0.951176 | FN | FN | DPP IV inhibitor |
| 0.948462 | FFDR | FF, DR | DPP IV inhibitor |
| 0.947101 | WSM | WS | DPP IV inhibitor |
| 0.946135 | QF | QF | DPP IV inhibitor |
| 0.941387 | AGRF | AG | DPP IV inhibitor |
| RF | DPP III inhibitor | ||
| 0.940972 | QGF | GF, QG | DPP IV inhibitor |
| 0.937189 | FPAG | PA, FP, AG | DPP IV inhibitor |
| 0.915352 | AYF | AY | DPP IV inhibitor |
| 0.913344 | DGJF | DG | DPP IV inhibitor, DPP III inhibitor |
| 0.912957 | WVAF | AF, VA, WV | DPP IV inhibitor |
| 0.909891 | WQ | WQ | DPP IV inhibitor |
| 0.906764 | KF | KF | DPP IV inhibitor |
| 0.905338 | NGJF | NG | DPP IV inhibitor |
| 0.894996 | FJPH | PH | DPP IV inhibitor |
| 0.888582 | MPJ | MP | DPP IV inhibitor |
| 0.887708 | KGF | GF, KG | DPP IV inhibitor |
| 0.887194 | JAF | AF | DPP IV inhibitor |
| 0.881235 | AFPA | PA, FP, AF | DPP IV inhibitor |
| 0.879704 | AYFG | AY | DPP IV inhibitor, antioxidant |
| YF | DPP IV inhibitor | ||
| 0.874411 | SFY | FR | DPP IV inhibitor |
| QR | DPP IV inhibitor | ||
| 0.873986 | WSMH | MH, WS | DPP IV inhibitor |
| 0.867049 | SFJ | SF | DPP IV inhibitor |
| 0.867026 | FNR | FN NR | DPP IV inhibitor |
| 0.866328 | PFGD | FG, GD | DPP IV inhibitor |
| DPP III inhibitor | |||
| 0.865974 | GPR | GP | DPP IV inhibitor |
| 0.863356 | RFN | FN | DPP IV inhibitor |
| 0.85592 | HWJY | HW | DPP IV inhibitor |
| 0.849148 | MR | MR | DPP IV inhibitor |
| 0.846587 | RPFD | RP | DPP IV inhibitor |
| PF | DPP IV inhibitor | ||
| 0.843478 | MY | MY | DPP IV inhibitor, antioxidant |
| 0.835091 | JSF | SF | DPP IV inhibitor |
| 0.833322 | AAF | AF, AA | DPP IV inhibitor |
| 0.825853 | FQR | FQ | DPP IV inhibitor |
| 0.820516 | SAF | AF | DPP IV inhibitor |
| 0.798127 | JWQ | WQ | DPP IV inhibitor |
| 0.796676 | WJYN | YN | DPP IV inhibitor |
| 0.785013 | YMDM | YM | DPP IV inhibitor |
| 0.768611 | GJYP | YP | DPP IV inhibitor |
| 0.761687 | RFT | TF | DPP IV inhibitor |
| DPP III inhibitor | |||
| 0.759378 | JFQ | FQ | DPP IV inhibitor |
| 0.754757 | JJSF | SF | DPP IV inhibitor |
| 0.739381 | YP | YP | DPP IV inhibitor |
| 0.728473 | FYP | YT | DPP IV inhibitor |
| 0.72244 | WVEF | VE | DPP IV inhibitor |
| WV | DPP IV inhibitor | ||
| 0.720745 | SRSF | SF | DPP IV inhibitor |
| 0.720069 | GFE | GF | DPP IV inhibitor, DPP III inhibitor |
| 0.717504 | QQF | QF, QQ | DPP IV inhibitor |
| 0.717158 | JSAF | AF | DPP IV inhibitor |
| 0.714532 | AQFJ | QF | DPP IV inhibitor |
| 0.713582 | AAFJ | AF, AA | DPP IV inhibitor |
| 0.693293 | MA | MA | DPP IV inhibitor |
| 0.689222 | GQGP | GP, QG | DPP IV inhibitor |
| 0.679365 | JVF | VF | DPP IV inhibitor |
| 0.659026 | KGFA | GF | DPP IV inhibitor |
| KG | DPP IV inhibitor | ||
| FA | DPP IV inhibitor | ||
| 0.65522 | SJVF | VF | DPP IV inhibitor |
| 0.642689 | GQP | QP | DPP IV inhibitor |
| 0.64074 | JPVM | VM | DPP IV inhibitor |
| PV | DPP IV inhibitor | ||
| 0.636164 | HFQ | FQ | DPP IV inhibitor |
| HF | DPP IV inhibitor | ||
| 0.623398 | MPJQ | MP | DPP IV inhibitor |
| 0.612254 | SFJQ | SF | DPP IV inhibitor |
| 0.601894 | KFN | KF | DPP IV inhibitor |
| FN | DPP IV inhibitor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartolomei, M.; Capriotti, A.L.; Li, Y.; Bollati, C.; Li, J.; Cerrato, A.; Cecchi, L.; Pugliese, R.; Bellumori, M.; Mulinacci, N.; et al. Exploitation of Olive (Olea europaea L.) Seed Proteins as Upgraded Source of Bioactive Peptides with Multifunctional Properties: Focus on Antioxidant and Dipeptidyl-Dipeptidase—IV Inhibitory Activities, and Glucagon-like Peptide 1 Improved Modulation. Antioxidants 2022, 11, 1730. https://doi.org/10.3390/antiox11091730
Bartolomei M, Capriotti AL, Li Y, Bollati C, Li J, Cerrato A, Cecchi L, Pugliese R, Bellumori M, Mulinacci N, et al. Exploitation of Olive (Olea europaea L.) Seed Proteins as Upgraded Source of Bioactive Peptides with Multifunctional Properties: Focus on Antioxidant and Dipeptidyl-Dipeptidase—IV Inhibitory Activities, and Glucagon-like Peptide 1 Improved Modulation. Antioxidants. 2022; 11(9):1730. https://doi.org/10.3390/antiox11091730
Chicago/Turabian StyleBartolomei, Martina, Anna Laura Capriotti, Yuchen Li, Carlotta Bollati, Jianqiang Li, Andrea Cerrato, Lorenzo Cecchi, Raffaele Pugliese, Maria Bellumori, Nadia Mulinacci, and et al. 2022. "Exploitation of Olive (Olea europaea L.) Seed Proteins as Upgraded Source of Bioactive Peptides with Multifunctional Properties: Focus on Antioxidant and Dipeptidyl-Dipeptidase—IV Inhibitory Activities, and Glucagon-like Peptide 1 Improved Modulation" Antioxidants 11, no. 9: 1730. https://doi.org/10.3390/antiox11091730
APA StyleBartolomei, M., Capriotti, A. L., Li, Y., Bollati, C., Li, J., Cerrato, A., Cecchi, L., Pugliese, R., Bellumori, M., Mulinacci, N., Laganà, A., Arnoldi, A., & Lammi, C. (2022). Exploitation of Olive (Olea europaea L.) Seed Proteins as Upgraded Source of Bioactive Peptides with Multifunctional Properties: Focus on Antioxidant and Dipeptidyl-Dipeptidase—IV Inhibitory Activities, and Glucagon-like Peptide 1 Improved Modulation. Antioxidants, 11(9), 1730. https://doi.org/10.3390/antiox11091730