
Simulated Gastrointestinal Digestion of Bioprocessed Spelt Seeds: Bioaccessibility and Bioactivity of Phenolics




Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Preparation
2.3. Simulated In Vitro Gastrointestinal Digestion of Spelt Seeds
2.4. Extraction of Extractable and Bound Phenolics
2.5. Determination of Total Phenolics Content
2.6. Phenolics Profile
2.7. Determination of Antioxidant Activity
2.7.1. DPPH• Scavenging Activity
2.7.2. ABTS•+ Scavenging Activity
2.8. Bioaccessibility and Biostability of Phenolics after Digestion
2.9. Statistical Analysis
3. Results and Discussion
3.1. Total Phenolic Content
3.2. Phenolic Profiles
3.3. Antioxidant Activity
3.4. Bioaccessibility and Biostability of Digested Phenolics
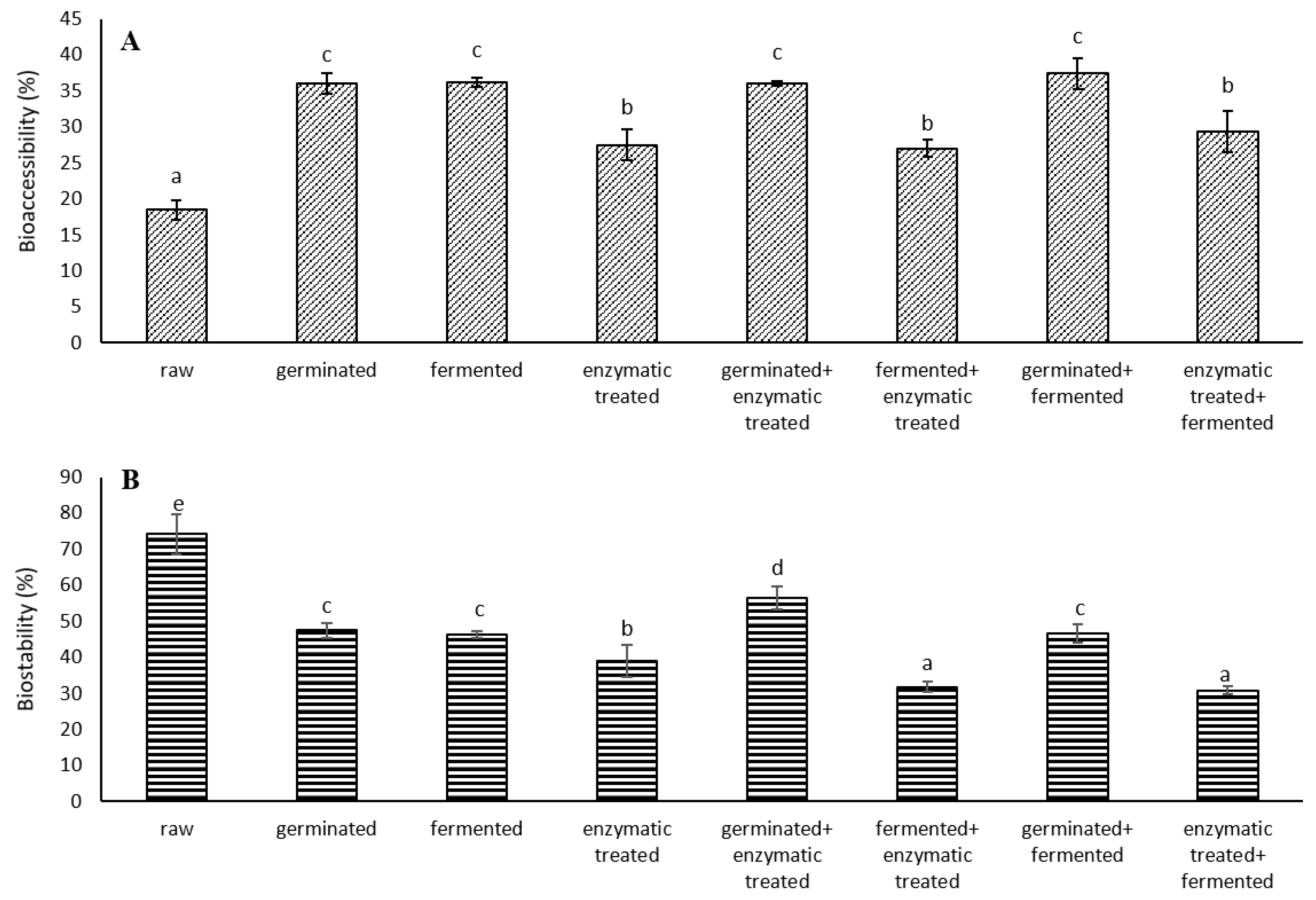
3.4.1. Bioaccessibility and Biostability of Total Phenolics Content in Analysed Spelt Seeds
3.4.2. Bioaccesebility and Biostability of Individual Phenolics in Analysed Spelt Seeds
3.5. Pearson Correlation Analysis
3.5.1. Correlations between the Proportion of Extractable Phenolics and Bioaccessibility
3.5.2. Correlations between Extractable Phenolic Acids of Digested and Undigested Spelt Seeds
3.6. Principal Component Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahidi, F.; Pan, Y. Influence of food matrix and food processing on the chemical interaction and bioaccessibility of dietary phytochemicals: A review. Crit. Rev. Food Sci. Nutr. 2021, 62, 6421–6445. [Google Scholar] [CrossRef] [PubMed]
- Anson, N.M.; Selinheimo, E.; Havenaar, R.; Aura, A.-M.; Mattila, I.; Lehtinen, P.; Bast, A.; Poutanen, K.; Haenen, G.R.M.M. Bioprocessing of wheat bran improves in vitro bioaccessibility and colonic metabolism of phenolic compounds. J. Agric. Food Chem. 2009, 57, 6148–6155. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Grijalva, E.; Ambriz-Pérez, D.; Leyva-López, N.; Castillo, R.; Heredia, J.B. Review: Dietary phenolic compounds, health benefits and bioaccessibility. Arch. Latinoam. Nutr. 2016, 66, 87–100. [Google Scholar]
- Mencin, M.; Abramovič, H.; Jamnik, P.; Mikulič Petkovšek, M.; Veberič, R.; Terpinc, P. Abiotic stress combinations improve the phenolics profiles and activities of extractable and bound antioxidants from germinated spelt (Triticum spelta L.) seeds. Food Chem. 2021, 344, 128704. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Wang, W.-K.; Wu, Q.-C.; Yang, H.-J. The release and catabolism of ferulic acid in plant cell wall by rumen microbes: A review. Anim. Nutr. 2022, 9, 335–344. [Google Scholar] [CrossRef]
- Vitaglione, P.; Napolitano, A.; Fogliano, V. Cereal Dietary Fibre: A natural functional ingredient to deliver phenolic compounds into the gut. Trends Food Sci. Technol. 2008, 19, 451–463. [Google Scholar] [CrossRef]
- Koistinen, V.M.; Nordlund, E.; Katina, K.; Mattila, I.; Poutanen, K.; Hanhineva, K.; Aura, A.-M. Effect of bioprocessing on the in vitro colonic microbial metabolism of phenolic acids from rye bran fortified breads. J. Agric. Food Chem. 2017, 65, 1854–1864. [Google Scholar] [CrossRef]
- Rocchetti, G.; Giuberti, G.; Lucini, L. Gluten-free cereal-based food products: The potential of metabolomics to investigate changes in phenolics profile and their in vitro bioaccessibility. Curr. Opin. Food Sci. 2018, 22, 1–8. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Dziki, D.; Baraniak, B.; Lin, R. The Effect of simulated digestion in vitro on bioactivity of wheat bread with tartary buckwheat flavones addition. LWT-Food Sci. Technol. 2009, 42, 137–143. [Google Scholar] [CrossRef]
- Saura-Calixto, F.; Serrano, J.; Goñi, I. Intake and bioaccessibility of total polyphenols in a whole diet. Food Chem. 2007, 101, 492–501. [Google Scholar] [CrossRef]
- Mencin, M.; Jamnik, P.; Mikulič Petkovšek, M.; Veberič, R.; Terpinc, P. Improving accessibility and bioactivity of raw, germinated and enzymatic-treated spelt (Triticum Spelta L.) seed antioxidants by fermentation. Food Chem. 2022, 394, 133483. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Mencin, M.; Mikulic-Petkovsek, M.; Veberič, R.; Terpinc, P. Development and optimisation of solid-phase extraction of extractable and bound phenolic acids in spelt (Triticum spelta L.) seeds. Antioxidants 2021, 10, 1085. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D.; Refaey, M.M.; Xu, W. High spatial and temporal variations of microbial community along the southern catfish gastrointestinal tract: Insights into dynamic food digestion. Front. Microbiol. 2017, 8, 1531. [Google Scholar] [CrossRef]
- Tomé-Sánchez, I.; Martín-Diana, A.B.; Peñas, E.; Frias, J.; Rico, D.; Jiménez-Pulido, I.; Martínez-Villaluenga, C. Bioprocessed wheat ingredients: Characterization, bioaccessibility of phenolic compounds, and bioactivity during in vitro digestion. Front. Plant Sci. 2021, 12, 790898. [Google Scholar] [CrossRef]
- Xu, K.; She, Z.; Wang, H.; Liu, X.; Zhang, Y.; Chi, C.; Zhang, H. Textural quality of sweet dumpling: Effect of germination of waxy rice. Int. J. Food Sci. Technol. 2021, 56, 4081–4090. [Google Scholar] [CrossRef]
- Ortega, N.; Macià, A.; Romero, M.-P.; Reguant, J.; Motilva, M.-J. Matrix composition effect on the digestibility of carob flour phenols by an in-vitro digestion model. Food Chem. 2011, 124, 65–71. [Google Scholar] [CrossRef]
- Ydjedd, S.; Bouriche, S.; López-Nicolás, R.; Sánchez-Moya, T.; Frontela-Saseta, C.; Ros-Berruezo, G.; Rezgui, F.; Louaileche, H.; Kati, D.-E. Effect of in vitro gastrointestinal digestion on encapsulated and nonencapsulated phenolic compounds of carob (Ceratonia Siliqua L.) pulp extracts and their antioxidant capacity. J. Agric. Food Chem. 2017, 65, 827–835. [Google Scholar] [CrossRef]
- Chait, Y.A.; Gunenc, A.; Bendali, F.; Hosseinian, F. Simulated gastrointestinal digestion and in vitro colonic fermentation of carob polyphenols: Bioaccessibility and bioactivity. LWT 2020, 117, 108623. [Google Scholar] [CrossRef]
- Konishi, Y.; Zhao, Z.; Shimizu, M. Phenolic acids are absorbed from the rat stomach with different absorption rates. J. Agric. Food Chem. 2006, 54, 7539–7543. [Google Scholar] [CrossRef]
- Zhao, Z.; Egashira, Y.; Sanada, H. Ferulic acid is quickly absorbed from rat stomach as the free form and then conjugated mainly in liver. J. Nutr. 2004, 134, 3083–3088. [Google Scholar] [CrossRef]
- Liu, M.; Liu, X.; Luo, J.; Bai, T.; Chen, H. Effect of digestion on bound phenolic content, antioxidant activity and hypoglycemic ability of insoluble dietary fibre from four Triticeae crops. J. Food Biochem. 2021, 45, e13746. [Google Scholar] [CrossRef]
- Mosele, J.I.; Motilva, M.-J.; Ludwig, I.A. Beta-glucan and phenolic compounds: Their concentration and behavior during in vitro gastrointestinal digestion and colonic fermentation of different barley-based food products. J. Agric. Food Chem. 2018, 66, 8966–8975. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free. Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Shahidi, F. Bioaccessibility and antioxidant potential of millet grain phenolics as affected by simulated in vitro digestion and microbial fermentation. J. Funct. Foods 2012, 4, 226–237. [Google Scholar] [CrossRef]
- Terpinc, P.; Abramovič, H. A kinetic approach for evaluation of the antioxidant activity of selected phenolic acids. Food Chem. 2010, 121, 366–371. [Google Scholar] [CrossRef]
- Correa-Betanzo, J.; Allen-Vercoe, E.; McDonald, J.; Schroeter, K.; Corredig, M.; Paliyath, G. Stability and biological activity of wild blueberry (Vaccinium Angustifolium) polyphenols during simulated in vitro gastrointestinal digestion. Food Chem. 2014, 165, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lin, H.; Lin, M.; Zheng, Y.; Chen, J. Effect of roasting and in vitro digestion on phenolic profiles and antioxidant activity of water-soluble extracts from sesame. Food Chem. Toxicol. 2020, 139, 111239. [Google Scholar] [CrossRef] [PubMed]
- Lucas-Gonzalez, R.; Navarro-Coves, S.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Assessment of polyphenolic profile stability and changes in the antioxidant potential of maqui berry (Aristotelia chilensis (Molina) Stuntz) during in vitro gastrointestinal digestion. Ind. Crops Prod. 2016, 94, 774–782. [Google Scholar] [CrossRef]
- Doniec, J.; Florkiewicz, A.; Dziadek, K.; Filipiak-Florkiewicz, A. Hydrothermal treatment effect on antioxidant activity and polyphenols concentration and profile of brussels sprouts (Brassica Oleracea Var. Gemmifera) in an in vitro simulated gastrointestinal digestion model. Antioxidants 2022, 11, 446. [Google Scholar] [CrossRef]
- Angelino, D.; Cossu, M.; Marti, A.; Zanoletti, M.; Chiavaroli, L.; Brighenti, F.; Rio, D.D.; Martini, D. Bioaccessibility and bioavailability of phenolic compounds in bread: A review. Food Funct. 2017, 8, 2368–2393. [Google Scholar] [CrossRef]
- Ren, Y.; Liu, H.; Wang, D.; Liu, T.; Zhang, R.; Wu, Y.; Zhang, Y.; Han, J.; Liu, M. Antioxidant activity, stability, in vitro digestion and cytotoxicity of two dietary polyphenols co-loaded by β-lactoglobulin. Food Chem. 2022, 371, 131385. [Google Scholar] [CrossRef]
- Goulas, V.; Hadjisolomou, A. Dynamic changes in targeted phenolic compounds and antioxidant potency of carob fruit (Ceratonia siliqua L.) products during in vitro digestion. LWT 2019, 101, 269–275. [Google Scholar] [CrossRef]
- Lima, K.; Silva, O.; Figueira, M.E.; Pires, C.; Cruz, D.; Gomes, S.; Maurício, E.M.; Duarte, M.P. Influence of the in vitro gastrointestinal digestion on the antioxidant activity of Artemisia gorgonum Webb and Hyptis pectinata (L.) Poit. infusions from Cape Verde. Food Res. Int. 2019, 115, 150–159. [Google Scholar] [CrossRef]
- Sęczyk, Ł.; Sugier, D.; Świeca, M.; Gawlik-Dziki, U. The effect of in vitro digestion, food matrix, and hydrothermal treatment on the potential bioaccessibility of selected phenolic compounds. Food Chem. 2021, 344, 128581. [Google Scholar] [CrossRef]
- Xu, M.; Rao, J.; Chen, B. Phenolic compounds in germinated cereal and pulse seeds: Classification, transformation, and metabolic process. Crit. Rev. Food Sci. Nutr. 2020, 60, 740–759. [Google Scholar] [CrossRef]
- Gullon, B.; Pintado, M.E.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. In vitro gastrointestinal digestion of pomegranate peel (Punica granatum) flour obtained from co-products: Changes in the antioxidant potential and bioactive compounds stability. J. Funct. Foods 2015, 19, 617–628. [Google Scholar] [CrossRef]
- Kroon, P.A.; Faulds, C.B.; Ryden, P.; Robertson, J.A.; Williamson, G. Release of covalently bound ferulic acid from fiber in the human colon. J. Agric. Food Chem. 1997, 45, 661–667. [Google Scholar] [CrossRef]
- Hemery, Y.M.; Anson, N.M.; Havenaar, R.; Haenen, G.R.M.M.; Noort, M.W.J.; Rouau, X. Dry-fractionation of wheat bran increases the bioaccessibility of phenolic acids in breads made from processed bran fractions. Food Res. Int. 2010, 43, 1429–1438. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant activity of grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Liu, C.; Luo, S.; Chen, J.; Gong, E. The profile and bioaccessibility of phenolic compounds in cereals influenced by improved extrusion cooking treatment. PLoS ONE 2016, 11, e0161086. [Google Scholar] [CrossRef] [PubMed]
- Mateo Anson, N.; van den Berg, R.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Bioavailability of ferulic acid is determined by its bioaccessibility. J. Cereal Sci. 2009, 49, 296–300. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| SSF (pH 7) | SGF (pH 3) | SIF (pH 7) | |||||
|---|---|---|---|---|---|---|---|
| Salt Solution | Stock Concentration (mol/L) | mL of Stock Added to Prepare 0.4 L (mL) | Final Salt Concentration in SSF (mmol/L) | mL of Stock Added to Prepare 0.5 L (mL) | Final Salt Concentration in SGF (mmol/L) | mL of Stock Added to Prepare 1 L (mL) | Final Salt Concentration in SIF (mmol/L) |
| KCl | 0.5 | 15.1 | 15.1 | 8.6 | 6.9 | 17 | 6.8 |
| KH2PO4 | 0.5 | 3.7 | 3.7 | 1.1 | 0.9 | 2 | 0.8 |
| NaHCO3 | 1 | 6.8 | 13.6 | 15.6 | 25 | 106.3 | 85 |
| NaCl | 2 | / | / | 14.8 | 47.2 | 24 | 38.4 |
| MgCl2(H2O)6 | 0.15 | 0.5 | 0.15 | 0.5 | 0.12 | 2.8 | 0.33 |
| (NH4)2CO3 | 0.5 | 0.06 | 0.06 | 0.6 | 0.5 | / | / |
| CaCl2(H2O)2 | 0.3 | / | 1.50 | / | 0.15 | / | 0.6 |
| TPC | DPPH | ABTS | ||||
|---|---|---|---|---|---|---|
| mg TE/g DW | mg TE/g DW | mg TE/g DW | ||||
| Sample | Extractable | Bound | Extractable | Bound | Extractable | Bound |
| Undigested | ||||||
| raw | 1.28 a | 6.73 D | 0.04 a | 0.85 EF | 1.30 a | 6.98 F |
| germinated | 4.85 e | 10.76 G | 0.43 d | 1.50 H | 4.21 d | 8.90 HI |
| fermented | 3.89 c | 7.77 E | 0.26 b | 0.87 F | 4.96 e | 7.13 F |
| enzymatic treated | 4.52 de | 6.13 C | 0.89 g | 0.80 E | 6.31 f | 6.01 E |
| germinated + enzymatic treated | 7.29 h | 9.90 F | 0.91 g | 1.69 J | 6.97 g | 8.14 G |
| fermented + enzymatic treated | 8.57 i | 5.72 BC | 1.21 i | 0.79 E | 7.45 h | 5.61 CD |
| germinated + fermented | 14.19 k | 13.08 H | 1.57 k | 1.84 K | 9.61 i | 9.05 I |
| enzymatic treated +fermented | 8.02 i | 6.10 C | 1.15 h | 0.85 EF | 7.16 g | 5.83 DE |
| Digested | ||||||
| raw | 1.48 a | 5.98 BC | 0.06 a | 0.70 D | 1.35 a | 6.10 E |
| germinated | 5.62 f | 7.39 E | 0.70 e | 0.40 B | 4.28 d | 7.21 F |
| fermented | 4.22 cd | 5.39 B | 0.23 b | 0.35 AB | 2.33 c | 6.09 E |
| enzymatic treated | 2.92 b | 4.13 A | 0.24 b | 0.56 C | 1.94 b | 5.36 BC |
| germinated + enzymatic treated | 6.19 g | 9.71 F | 0.75 f | 1.32 G | 4.28 d | 7.26 F |
| fermented + enzymatic treated | 3.85 c | 4.53 A | 0.36 c | 0.36 AB | 2.28 c | 5.15 B |
| germinated + fermented | 10.19 j | 12.48 H | 1.28 j | 1.60 I | 6.24 f | 8.57 H |
| enzymatic treated + fermented | 4.14 c | 4.34 A | 0.44 d | 0.31 A | 2.37 c | 4.51 A |
| Phenolic Acids (µg/g DW) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sample | p-Coumaric Acid | trans-Ferulic Acid | cis-Ferulic Acid | Caffeic Acid | p-Hydroxybenzoic Acid | |||||
| Extractable | Bound | Extractable | Bound | Extractable | Bound | Extractable | Bound | Extractable | Bound | |
| Undigested | ||||||||||
| raw | 0.27 a | 28.86 A | 0.59 a | 834.99 E | 0.30 a | 37.14 E | 0.29 a | 7.87 E | 1.20 a | 12.57 B |
| germinated | 3.54 d | 221.60 E | 9.19 bc | 1466.45 J | 4.75 c | 80.77 K | 2.92 e | 13.98 I | 14.24 f | 18.13 D |
| fermented | 1.09 b | 32.11 BC | 3.58 a | 965.96 G | 1.56 b | 54.02 G | 0.90 b | 5.17 BC | 9.21 c | 15.30 C |
| enzymatic treated | 6.37 h | 36.07 C | 140.68 j | 578.91 E | 8.14 f | 37.86 E | 5.06 h | 8.46 E | 24.81 i | 30.12 EF |
| germinated + enzymatic treated | 7.90 j | 265.04 G | 111.56 i | 1351.42 I | 7.62 ef | 61.57 H | 3.93 g | 14.48 I | 12.19 de | 49.63 G |
| fermented + enzymatic treated | 7.95 j | 38.21 C | 345.74 l | 532.01 CD | 22.87 h | 26.87 B | 7.63 j | 6.39 D | 36.29 j | 29.62 E |
| germinated + fermented | 15.31 l | 298.17 H | 68.32 g | 1699.14 K | 30.84 i | 77.15 J | 22.56 l | 14.40 I | 97.27 m | 31.50 EF |
| enzymatic treated + fermented | 6.84 i | 35.67 C | 320.92 k | 549.15 DE | 19.17 g | 30.38 C | 6.42 i | 6.30 D | 70.08 l | 29.34 E |
| Digested | ||||||||||
| raw | 0.44 a | 28.07 A | 0.61 a | 820.05 F | 0.49 a | 33.27 D | 0.53 a | 7.59 E | 2.27 b | 11.18 B |
| germinated | 4.06 e | 185.19 D | 14.39 c | 1210.44 H | 6.36 d | 44.52 F | 3.27 f | 9.19 F | 18.39 g | 17.60 D |
| fermented | 2.25 c | 27.80 A | 4.26 ab | 805.45 F | 2.05 b | 33.76 D | 1.35 c | 2.61 A | 11.57 d | 10.48 B |
| enzymatic treated | 3.69 d | 33.56 BC | 12.48 c | 493.10 B | 2.10 b | 28.01 BC | 2.40 d | 8.65 EF | 11.26 d | 16.62 CD |
| germinated + enzymatic treated | 5.70 g | 238.38 F | 34.95 d | 1209.11 H | 7.26 e | 55.35 G | 5.24 h | 11.05 G | 21.03 h | 29.64 E |
| fermented + enzymatic treated | 3.90 de | 39.17 C | 41.55 e | 508.46 BC | 2.04 b | 21.90 A | 3.73 g | 5.73 C | 26.82 i | 9.73 AB |
| germinated + fermented | 11.05 k | 293.85 H | 85.61 h | 1668.17 K | 31.45 i | 65.84 I | 15.93 k | 12.84 H | 52.57 k | 31.75 F |
| enzymatic treated + fermented | 5.32 f | 36.29 C | 55.21 f | 382.21 A | 6.31 d | 22.23 A | 5.06 h | 4.95 B | 53.46 k | 8.30 A |
| Flavonoids (µg/g DW) | |||||
|---|---|---|---|---|---|
| Gallocatechin | Apigenin | Unknown C-Glycosyl Derivative | |||
| I | II | III | |||
| Sample | Extractable | Extractable | Extractable | Extractable | Extractable |
| Undigested | |||||
| raw | 1.01 ± 0.13 a | 4.28 ± 0.06 a | 1.36 ± 0.04 a | 8.95 ± 0.29 d | 0.45 ± 0.03 a |
| germinated | 7.16 ± 0.28 e | 6.17 ± 0.13 d | 7.72 ± 0.30 i | 7.21 ± 0.29 b | 2.86 ± 0.16 e |
| fermented | 3.42 ± 0.13 b | 5.88 ± 0.15 d | 2.36 ± 0.14 c | 9.70 ± 0.29 e | 1.32 ± 0.03 b |
| enzymatic treated | 6.24 ± 0.26 d | 8.47 ± 0.23 g | 6.02 ± 0.19 g | 13.02 ± 0.40 g | 4.82 ± 0.11 j |
| germinated + enzymatic treated | 31.21 ± 1.48 l | 7.56 ± 0.22 f | 7.33 ± 0.25 h | 10.69 ± 0.27 f | 4.70 ± 0.12 j |
| fermented + enzymatic treated | 22.95 ± 0.59 i | 9.85 ± 0.28 i | 5.03 ± 0.22 f | 15.28 ± 0.31 h | 4.04 ± 0.10 i |
| germinated + fermented | 39.57 ± 1.00 m | 23.21 ± 0.57 k | 18.95 ± 0.68 j | 35.19 ± 0.95 j | 7.13 ± 0.37 k |
| enzymatic treated + fermented | 24.37 ± 0.69 j | 9.23 ± 0.38 h | 5.17 ± 0.15 f | 15.67 ± 0.25 h | 2.11 ± 0.13 d |
| Digested | |||||
| raw | 1.00 ± 0.17 a | 4.33 ± 0.06 a | 3.74 ± 0.20 e | 10.66 ± 0.15 f | 1.82 ± 0.04 c |
| germinated | 11.34 ± 0.22 f | 4.78 ± 0.16 b | 5.92 ± 0.52 g | 5.98 ± 0.30 a | 2.83 ± 0.14 e |
| fermented | 3.60 ± 0.07 bc | 4.26 ± 0.31 a | 1.90 ± 0.05 b | 7.91 ± 0.51 c | 3.37 ± 0.09 g |
| enzymatic treated | 3.93 ± 0.27 c | 4.27 ± 0.18 a | 2.11 ± 0.09 bc | 8.03 ± 0.41 c | 3.44 ± 0.11 g |
| germinated + enzymatic treated | 32.13 ± 0.84 k | 7.49 ± 0.71 f | 7.58 ± 0.34 hi | 7.75 ± 0.50 c | 8.20 ± 0.21 l |
| fermented + enzymatic treated | 20.93 ± 0.92 h | 6.93 ± 0.61 e | 2.82 ± 0.18 d | 7.78 ± 0.44 c | 3.72 ± 0.22 h |
| germinated + fermented | 25.89 ± 0.43 k | 18.19 ± 0.75 j | 19.27 ± 0.57 k | 16.83 ± 0.82 i | 10.62 ± 0.38 m |
| enzymatic treated + fermented | 17.51 ± 0.81 g | 5.44 ± 0.54 c | 2.84 ± 0.16 d | 9.79 ± 0.35 e | 3.14 ± 0.16 f |
| Bioaccessibility (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Digested Spelt Seeds | p-Coumaric Acid | trans-Ferulic Acid | cis-Ferulic Acid | Caffeic Acid | p-Hydroxybenzoic Acid | Gallocatechin | Apigenin | Unknown C-glycosyl Derivative | ||
| I | II | III | ||||||||
| raw | 1.5 a | 0.1 a | 1.3 a | 6.5 a | 16.5 a | 43.1 a | 45.6 e | 83.4 g | 61.0 e | 10.9 b |
| germinated | 1.8 b | 1.0 c | 7.4 e | 19.3 c | 56.8 g | 85.9 f | 32.5 b | 32.7 b | 33.5 a | 8.1 a |
| fermented | 6.8 e | 0.4 b | 3.7 b | 22.2 d | 47.2 e | 66.9 d | 39.1 d | 36.2 d | 44.2 d | 22.8 f |
| enzymatic treated | 8.7 f | 1.7 d | 4.6 d | 17.8 b | 20.5 b | 41.2 a | 30.4 a | 21.4 a | 37.6 b | 16.8 c |
| germinated + enzymatic treated | 2.1 c | 2.4 e | 10.5 f | 28.5 f | 34.0 c | 85.3 f | 47.2 f | 45.5 e | 35.5 b | 20.1 e |
| fermented + enzymatic treated | 8.5 f | 4.7 f | 4.1 c | 26.6 e | 40.7 d | 78.6 e | 47.8 f | 34.9 c | 34.9 a | 21.7 f |
| germinated + fermented | 3.5 d | 4.8 f | 29.1 h | 43.1 h | 40.8 d | 58.1 b | 61.5 g | 69.8 f | 36.6 b | 26.0 g |
| enzymatic treated + fermented | 12.5 g | 6.3 g | 12.7 g | 39.8 g | 53.8 f | 61.5 c | 36.4 c | 30.6 b | 41.2 c | 17.4 d |
| Biostability (%) | ||||||||||
| raw | 96.4 f | 98.1 f | 88.9 e | 93.1 h | 81.2 h | 48.4 f | 48.3 e | 53.0 h | 42.7 e | 91.7 e |
| germinated | 82.3 b | 82.0 d | 52.1 b | 54.4 e | 54.4 g | 37.0 e | 48.5 e | 40.9 e | 59.3 g | 89.0 d |
| fermented | 83.7 b | 83.1 d | 60.8 c | 43.0 d | 42.8 e | 29.4 c | 13.8 a | 43.7 f | 29.9 b | 80.9 c |
| enzymatic treated | 79.1 a | 68.5 c | 60.9 c | 64.0 f | 30.2 d | 35.5 d | 39.2 d | 37.1 d | 38.4 d | 74.6 b |
| germinated + enzymatic treated | 87.3 d | 82.6 d | 80.0 d | 60.0 g | 47.9 f | 7.6 a | 53.7 f | 51.9 g | 47.9 f | 88.3 d |
| fermented + enzymatic treated | 84.8 c | 57.9 b | 44.0 a | 40.9 c | 14.8 b | 13.2 b | 20.0 c | 33.3 b | 32.4 c | 75.9 b |
| germinated + fermented | 93.7 e | 94.4 e | 61.0 c | 34.7 a | 24.7 c | 8.0 a | 20.4 c | 30.1 a | 22.8 a | 81.6 c |
| enzymatic treated + fermented | 85.4 c | 43.9 a | 44.9 a | 38.9 b | 8.3 a | 13.8 b | 18.6 b | 35.7 c | 33.7 c | 65.4 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mencin, M.; Mikulič Petkovšek, M.; Veberič, R.; Terpinc, P. Simulated Gastrointestinal Digestion of Bioprocessed Spelt Seeds: Bioaccessibility and Bioactivity of Phenolics. Antioxidants 2022, 11, 1703. https://doi.org/10.3390/antiox11091703
Mencin M, Mikulič Petkovšek M, Veberič R, Terpinc P. Simulated Gastrointestinal Digestion of Bioprocessed Spelt Seeds: Bioaccessibility and Bioactivity of Phenolics. Antioxidants. 2022; 11(9):1703. https://doi.org/10.3390/antiox11091703
Chicago/Turabian StyleMencin, Marjeta, Maja Mikulič Petkovšek, Robert Veberič, and Petra Terpinc. 2022. "Simulated Gastrointestinal Digestion of Bioprocessed Spelt Seeds: Bioaccessibility and Bioactivity of Phenolics" Antioxidants 11, no. 9: 1703. https://doi.org/10.3390/antiox11091703
APA StyleMencin, M., Mikulič Petkovšek, M., Veberič, R., & Terpinc, P. (2022). Simulated Gastrointestinal Digestion of Bioprocessed Spelt Seeds: Bioaccessibility and Bioactivity of Phenolics. Antioxidants, 11(9), 1703. https://doi.org/10.3390/antiox11091703