
Lipid Peroxidation in Obesity: Can Bariatric Surgery Help?

 , , ,
, , ,  and
and
Abstract
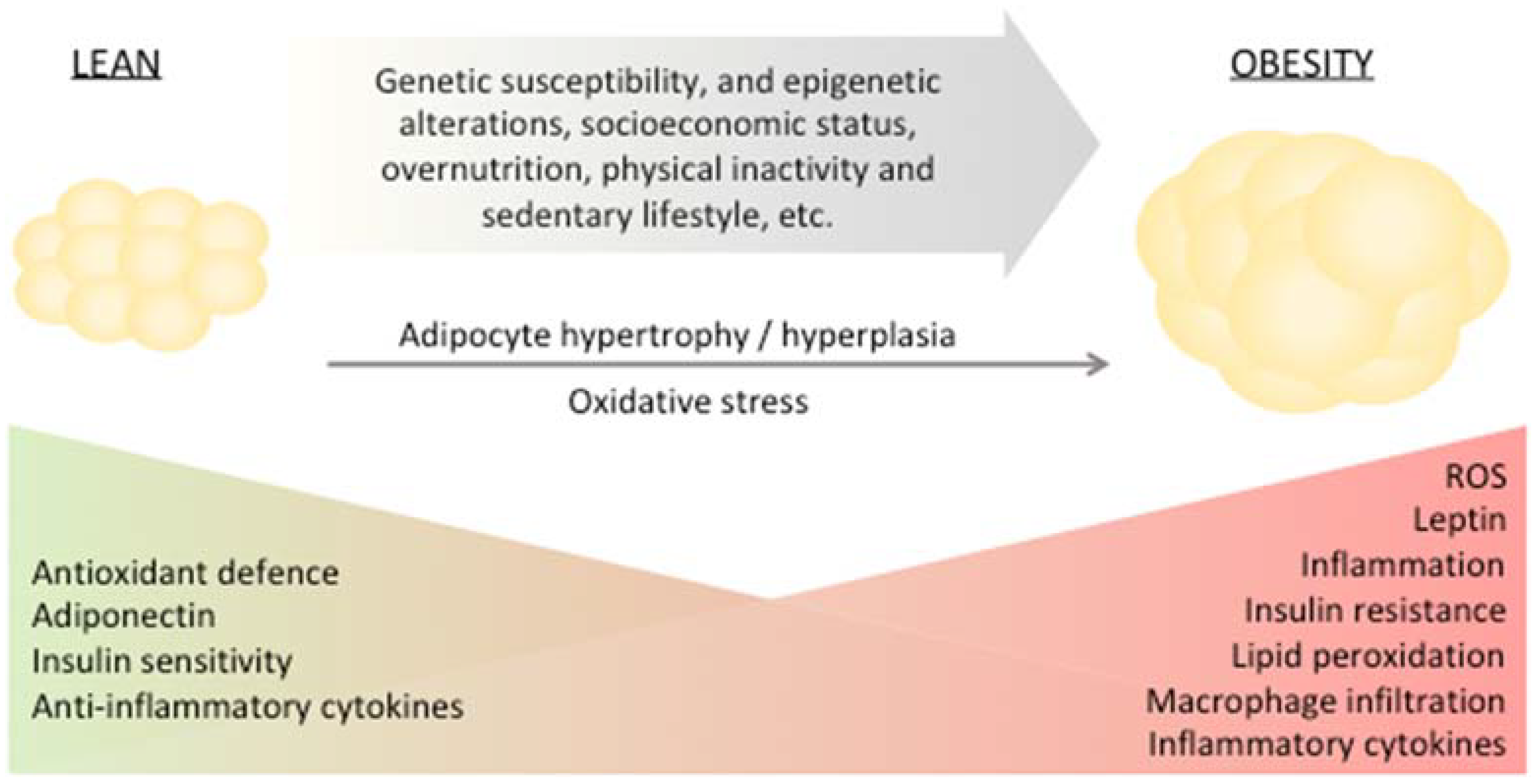
:1. Obesity
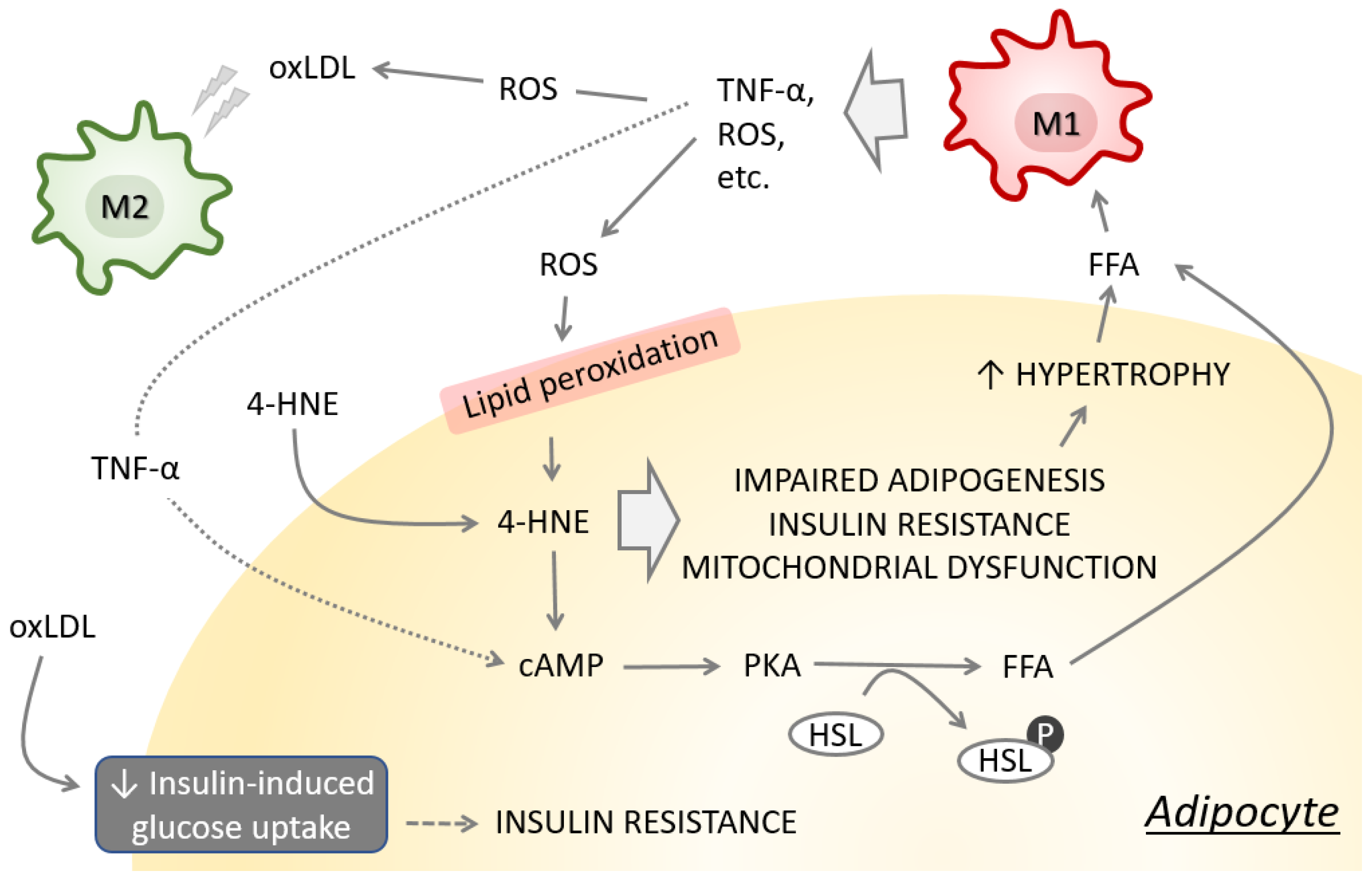
2. Adipose Tissue Remodeling, Impairment of Redox Homeostasis and Dysfunction
Obesity—A Major Risk Factor for T2DM
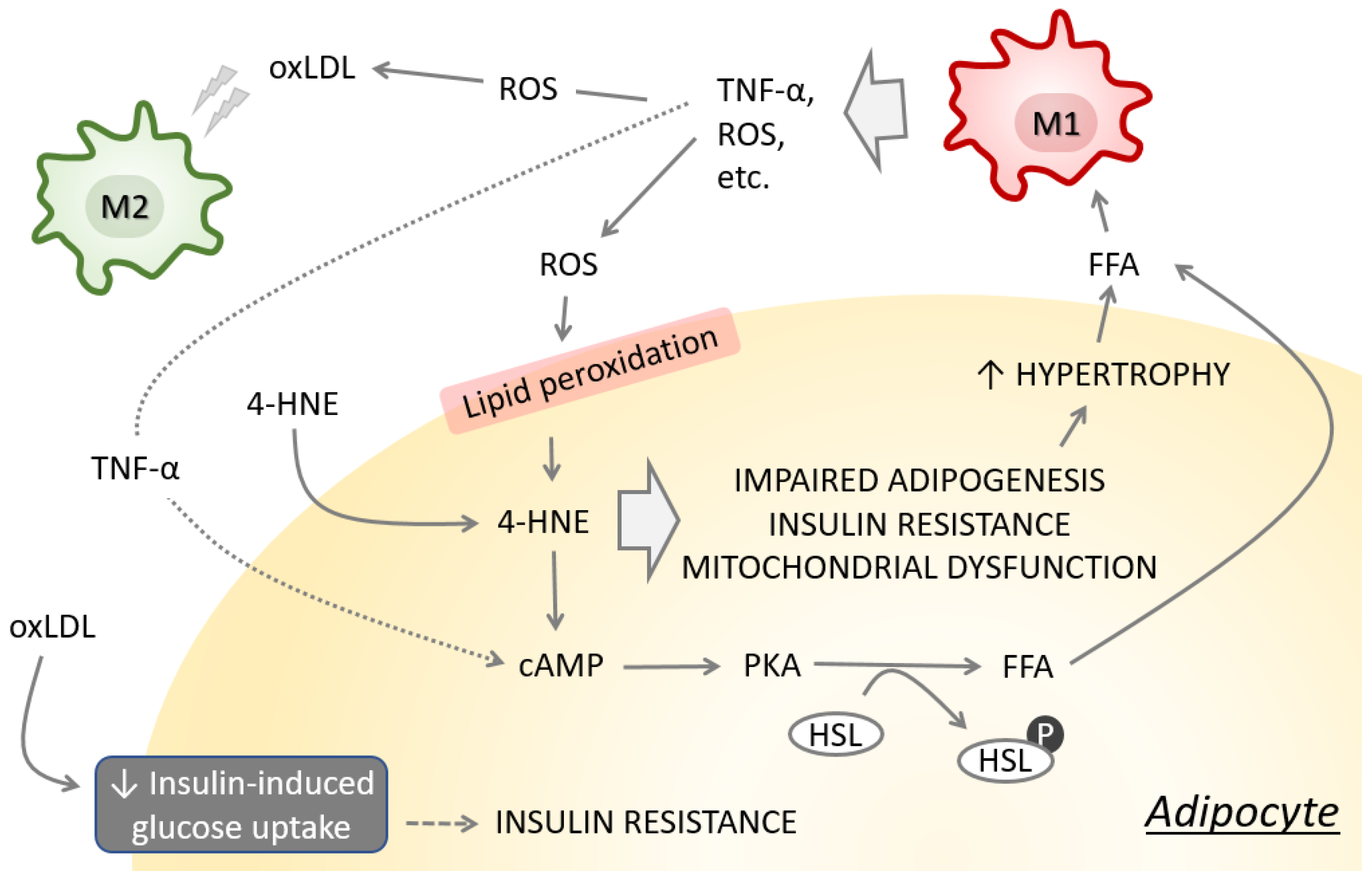
3. Consequences of Obesity-Induced Lipid Peroxidation

{kind=link}
{kind=link}
{kind=link}
| Process Affected | The Involvement of Obesity-Induced 4-HNE | Ref |
|---|---|---|
| Ferroptosis | HFD-induced obesity upregulates prostaglandin endoperoxide synthase 2 expression and promotes lipid peroxidation. Exosomes derived from obese adipose tissue macrophages upregulate prostaglandin endoperoxide synthase 2, promote formation of 4-HNE and induce mitochondrial injury. Obese adipose tissue macrophages exosomes contain a high level of miR-140-5p that affects GSH synthesis and promotes ferroptosis and cardiac injury in obesity. | [59] |
| Autophagy and proteostasis | HFD induces accumulation of lipid droplets in the liver and affects autophagy efficiency promoting accumulation of proteins modified with 4-HNE and 3-nitrotyrosine. | [60] |
| Carbonylation of histones | Obesity increases the level of 4-HNE-modified histones. | [64] |
| Omental adipogenesis | Omental adipose tissue of morbidly obese individuals revealed that smaller size of adipocytes, increased adipocytes’ accumulation of 4-HNE-modified proteins and increased adipose tissue macrophage infiltration is associated with impaired adipogenesis. | [33] |
| Subcutaneous adipogenesis and insulin resistance | The 4-HNE-modified proteins accumulate in subcutaneous adipose tissue of obese individuals, with the highest presence in adipocytes. The 4-HNE affects redox homeostasis and inhibits growth subcutaneous preadipocytes. In addition, 4-HNE affects adipogenic capacity and insulin signaling promoting insulin resistance phenotype. | [34] |
| Inflammation | Obese individuals have higher amount of circulating 4-HNE. The 4-HNE inhibits miR-29b while it promotes adipokine EST1, resulting in TNF-α upregulation. In obesity, adipokines TNF-α, ETS1, and SP1 are upregulated while miR-29b is downregulated in the subcutaneous white adipose tissue. | [63] |
| Protein carbonylation | High fat, high sucrose diet induces carbonyl stress and accumulation of 4-HNE adducts and is accompanied with increased GPx4 enzyme in heart and liver. | [66] |
| Lipolysis | The 4-HNE stimulates lipolysis in adipocytes via upregulation of intracellular cyclic AMP level and increased phosphorylation of protein kinase A, resulting in elevated hormone sensitive lipase. In addition, 4-HNE downregulates AMP-activated protein kinase further supporting lipolysis. | [67] |
| Adiponectin secretion | HFD-induced obesity is associated with 4-HNE accumulation in adipose tissue and plasma adiponectin reduction. In adipocytes, 4-HNE promotes adiponectin ubiquitination enhancing its degradation via ubiquitin-proteasome system and ultimately resulting in its decreased secretion. | [68] |
| Protein carbonylation | High fat, high carbohydrate diet downregulates glutathione S-transferase A4 in adipose tissue, allowing 4-HNE carbonylation of proteins including fatty acid binding protein. | [65] |
| Protein carbonylation | Obesity leads to 4-HNE and 4-hydroxyhexenal carbonylation of nuclear zinc finger proteins. | [69] |
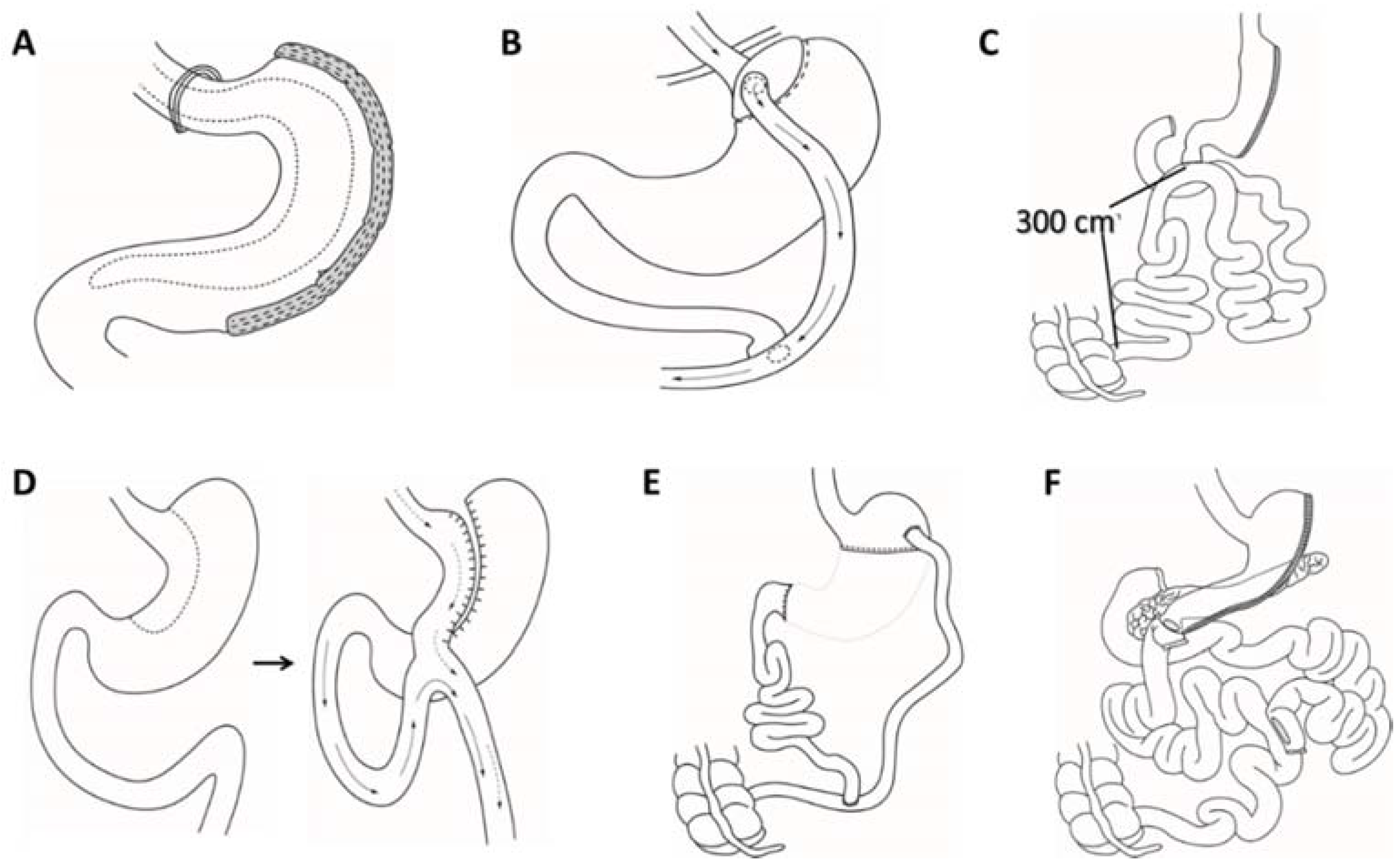
4. Surgical Interventions for Obesity and the Impact on Redox Homeostasis
4.1. Bariatric Surgery
4.2. Mental and Redox Balance Consequences of Bariatric Surgery
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Obesity and overweight, 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 4 May 2022).
- Bray, G.A. Medical consequences of obesity. J. Clin. Endocrinol. Metab. 2004, 89, 2583–2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lorenzo, A.; Gratteri, S.; Gualtieri, P.; Cammarano, A.; Bertucci, P.; Di Renzo, L. Why primary obesity is a disease? J. Transl. Med. 2019, 17, 169. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, J.; Farr, O.; Perakakis, N.; Ghaly, W.; Mantzoros, C. Obesity as a Disease. Med. Clin. North Am. 2018, 102, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Pi-Sunyer, X. The medical risks of obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef]
- Chooi, Y.C.; Ding, C.; Magkos, F. The epidemiology of obesity. Metabolism 2019, 92, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Keith, S.W.; Redden, D.T.; Katzmarzyk, P.T.; Boggiano, M.M.; Hanlon, E.C.; Benca, R.M.; Ruden, D.; Pietrobelli, A.; Barger, J.L.; Fontaine, K.R.; et al. Putative contributors to the secular increase in obesity: Exploring the roads less traveled. Int. J. Obes. 2006, 30, 1585–1594. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.; Jannah, N.; Kahan, S.; Gallagher, C.; Dietz, W. Current Knowledge of Obesity Treatment Guidelines by Health Care Professionals. Obesity 2018, 26, 665–671. [Google Scholar] [CrossRef]
- Tak, Y.J.; Lee, S.Y. Long-Term Efficacy and Safety of Anti-Obesity Treatment: Where Do We Stand? Curr. Obes. Rep. 2021, 10, 14–30. [Google Scholar] [CrossRef]
- White, U.; Ravussin, E. Dynamics of adipose tissue turnover in human metabolic health and disease. Diabetologia 2019, 62, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-K.; Lee, B.; Kim, C.Y. Natural Extracts That Stimulate Adipocyte Browning and Their Underlying Mechanisms. Antioxidants 2021, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Townsend, K.L.; Tseng, Y.-H. Brown fat fuel utilization and thermogenesis. Trends Endocrinol. Metab. 2014, 25, 168–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Okamatsu-Ogura, Y.; Matsushita, M.; Watanabe, K.; Yoneshiro, T.; Nio-Kobayashi, J.; Iwanaga, T.; Miyagawa, M.; Kameya, T.; Nakada, K.; et al. High Incidence of Metabolically Active Brown Adipose Tissue in Healthy Adult Humans: Effects of Cold Exposure and Adiposity. Diabetes 2009, 58, 1526–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijgen, G.H.E.J.; Bouvy, N.D.; Teule, G.J.J.; Brans, B.; Schrauwen, P.; van Marken Lichtenbelt, W.D. Brown adipose tissue in morbidly obese subjects. PLoS ONE 2011, 6, e17247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulterer, O.C.; Herz, C.T.; Prager, M.; Schmöltzer, C.; Langer, F.B.; Prager, G.; Marculescu, R.; Kautzky-Willer, A.; Hacker, M.; Haug, A.R.; et al. Brown Adipose Tissue Prevalence Is Lower in Obesity but Its Metabolic Activity Is Intact. Front. Endocrinol. 2022, 13, 858417. [Google Scholar] [CrossRef]
- Herz, C.T.; Kulterer, O.C.; Prager, M.; Schmöltzer, C.; Langer, F.B.; Prager, G.; Marculescu, R.; Kautzky-Willer, A.; Hacker, M.; Haug, A.R.; et al. Active Brown Adipose Tissue is Associated With a Healthier Metabolic Phenotype in Obesity. Diabetes 2021, 71, 93–103. [Google Scholar] [CrossRef]
- Zinngrebe, J.; Debatin, K.-M.; Fischer-Posovszky, P. Adipocytes in hematopoiesis and acute leukemia: Friends, enemies, or innocent bystanders? Leukemia 2020, 34, 2305–2316. [Google Scholar] [CrossRef]
- Orliaguet, L.; Ejlalmanesh, T.; Alzaid, F. Metabolic and Molecular Mechanisms of Macrophage Polarisation and Adipose Tissue Insulin Resistance. Int. J. Mol. Sci. 2020, 21, 5731. [Google Scholar] [CrossRef]
- Lumeng, C.N.; Deyoung, S.M.; Bodzin, J.L.; Saltiel, A.R. Increased inflammatory properties of adipose tissue macrophages recruited during diet-induced obesity. Diabetes 2007, 56, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W.J. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; DelProposto, J.B.; Westcott, D.J.; Saltiel, A.R. Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes 2008, 57, 3239–3246. [Google Scholar] [CrossRef] [Green Version]
- Blüher, M. Adipose tissue dysfunction in obesity. Exp. Clin. Endocrinol. Diabetes 2009, 117, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Cipak, A.; Schaur, R.J.; Zarkovic, N. Pathophysiology of neutrophil-mediated extracellular redox reactions. Front. Biosci. Landmark 2016, 21, 839–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulker, O.C.; Panieri, E.; Suzen, S.; Jaganjac, M.; Zarkovic, N.; Saso, L. Short overview on the relevance of microRNA-reactive oxygen species (ROS) interactions and lipid peroxidation for modulation of oxidative stress-mediated signalling pathways in cancer treatment. J. Pharm. Pharmacol. 2021, 74, 503–515. [Google Scholar] [CrossRef]
- Žarković, N.; Jaganjac, M.; Žarković, K.; Gęgotek, A.; Skrzydlewska, E. Spontaneous Regression of Cancer: Revealing Granulocytes and Oxidative Stress as the Crucial Double-edge Sword. Front. Biosci. 2022, 27, 119. [Google Scholar] [CrossRef]
- Jaganjac, M.; Milkovic, L.; Zarkovic, N.; Zarkovic, K. Oxidative stress and regeneration. Free Radic. Biol. Med. 2022, 181, 154–165. [Google Scholar] [CrossRef]
- Sredoja Tisma, V.; Bulimbasic, S.; Galesic Ljubanovic, D.; Galesic, K.; Morovic-Vergles, J.; Mitrovic, J.; Uchida, K.; Tatzber, F.; Zarkovic, N.; Jaganjac, M. The Onset of Systemic Oxidative Stress Associated with the Accumulation of Lipid Peroxidation Product Acrolein in the Skin of Patients with Small-Vessel Vasculitis. Molecules 2021, 26, 2344. [Google Scholar] [CrossRef]
- Choromańska, B.; Myśliwiec, P.; Dadan, J.; Maleckas, A.; Zalewska, A.; Maciejczyk, M. Effects of age and gender on the redox homeostasis of morbidly obese people. Free Radic. Biol. Med. 2021, 175, 108–120. [Google Scholar] [CrossRef]
- Jaganjac, M.; Milkovic, L.; Gegotek, A.; Cindric, M.; Zarkovic, K.; Skrzydlewska, E.; Zarkovic, N. The relevance of pathophysiological alterations in redox signaling of 4-hydroxynonenal for pharmacological therapies of major stress-associated diseases. Free Radic. Biol. Med. 2020, 157, 128–153. [Google Scholar] [CrossRef]
- Jaganjac, M.; Almuraikhy, S.; Al-Khelaifi, F.; Al-Jaber, M.; Bashah, M.; Mazloum, N.A.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Kafienah, W.; et al. Combined metformin and insulin treatment reverses metabolically impaired omental adipogenesis and accumulation of 4-hydroxynonenal in obese diabetic patients. Redox Biol. 2017, 12, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elrayess, M.A.; Almuraikhy, S.; Kafienah, W.; Al-Menhali, A.; Al-Khelaifi, F.; Bashah, M.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Alsayrafi, M.; et al. 4-hydroxynonenal causes impairment of human subcutaneous adipogenesis and induction of adipocyte insulin resistance. Free Radic. Biol. Med. 2017, 104, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozata, M.; Mergen, M.; Oktenli, C.; Aydin, A.; Yavuz Sanisoglu, S.; Bolu, E.; Yilmaz, M.I.; Sayal, A.; Isimer, A.; Ozdemir, I.C. Increased oxidative stress and hypozincemia in male obesity. Clin. Biochem. 2002, 35, 627–631. [Google Scholar] [CrossRef]
- Tinahones, F.J.; Murri-Pierri, M.; Garrido-Sánchez, L.; García-Almeida, J.M.; García-Serrano, S.; García-Arnés, J.; García-Fuentes, E. Oxidative stress in severely obese persons is greater in those with insulin resistance. Obesity 2009, 17, 240–246. [Google Scholar] [CrossRef]
- Samjoo, I.A.; Safdar, A.; Hamadeh, M.J.; Raha, S.; Tarnopolsky, M.A. The effect of endurance exercise on both skeletal muscle and systemic oxidative stress in previously sedentary obese men. Nutr. Diabetes 2013, 3, e88. [Google Scholar] [CrossRef] [Green Version]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The Impact of Oxidative Stress on Adipose Tissue Energy Balance. Front. Physiol. 2019, 10, 1638. [Google Scholar] [CrossRef]
- Hajer, G.R.; van Haeften, T.W.; Visseren, F.L.J. Adipose tissue dysfunction in obesity, diabetes, and vascular diseases. Eur. Heart J. 2008, 29, 2959–2971. [Google Scholar] [CrossRef] [Green Version]
- Chan, P.-C.; Hsieh, P.-S. The Role of Adipocyte Hypertrophy and Hypoxia in the Development of Obesity-Associated Adipose Tissue Inflammation and Insulin Resistance. In Adiposity; Gordeladze, J.O., Ed.; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Suganami, T.; Nishida, J.; Ogawa, Y. A paracrine loop between adipocytes and macrophages aggravates inflammatory changes: Role of free fatty acids and tumor necrosis factor alpha. Arter. Thromb. Vasc. Biol. 2005, 25, 2062–2068. [Google Scholar] [CrossRef] [Green Version]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S.; et al. Role of the Toll-like receptor 4/NF-kappaB pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Arter. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Permana, P.A.; Menge, C.; Reaven, P.D. Macrophage-secreted factors induce adipocyte inflammation and insulin resistance. Biochem. Biophys. Res. Commun. 2006, 341, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Hacohen, N.; Golub, T.R.; Van Parijs, L.; Lodish, H.F. Tumor necrosis factor-alpha suppresses adipocyte-specific genes and activates expression of preadipocyte genes in 3T3-L1 adipocytes: Nuclear factor-kappaB activation by TNF-alpha is obligatory. Diabetes 2002, 51, 1319–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Njajou, O.T.; Kanaya, A.M.; Holvoet, P.; Connelly, S.; Strotmeyer, E.S.; Harris, T.B.; Cummings, S.R.; Hsueh, W.-C. Association between oxidized LDL, obesity and type 2 diabetes in a population-based cohort, the Health, Aging and Body Composition Study. Diabetes Metab. Res. Rev. 2009, 25, 733–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Fernández, C.; Martin-Reyes, F.; Tome, M.; Ocaña-Wilhelmi, L.; Rivas-Becerra, J.; Tatzber, F.; Pursch, E.; Tinahones, F.J.; García-Fuentes, E.; Garrido-Sánchez, L. Oxidized LDL Modify the Human Adipocyte Phenotype to an Insulin Resistant, Proinflamatory and Proapoptotic Profile. Biomolecules 2020, 10, 534. [Google Scholar] [CrossRef] [Green Version]
- Isa, S.A.; Ruffino, J.S.; Ahluwalia, M.; Thomas, A.W.; Morris, K.; Webb, R. M2 macrophages exhibit higher sensitivity to oxLDL-induced lipotoxicity than other monocyte/macrophage subtypes. Lipids Health Dis. 2011, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Petan, T.; Jarc, E.; Jusović, M. Lipid Droplets in Cancer: Guardians of Fat in a Stressful World. Molecules 2018, 23, 1941. [Google Scholar] [CrossRef] [Green Version]
- Jarc, E.; Petan, T. Lipid Droplets and the Management of Cellular Stress. Yale J. Biol. Med. 2019, 92, 435–452. [Google Scholar]
- Korac, B.; Kalezic, A.; Pekovic-Vaughan, V.; Korac, A.; Jankovic, A. Redox changes in obesity, metabolic syndrome, and diabetes. Redox Biol. 2021, 42, 101887. [Google Scholar] [CrossRef]
- Yan, S.D.; Schmidt, A.M.; Anderson, G.M.; Zhang, J.; Brett, J.; Zou, Y.S.; Pinsky, D.; Stern, D. Enhanced cellular oxidant stress by the interaction of advanced glycation end products with their receptors/binding proteins. J. Biol. Chem. 1994, 269, 9889–9897. [Google Scholar] [CrossRef]
- Chang, P.-C.; Chen, T.-H.; Chang, C.-J.; Hou, C.-C.; Chan, P.; Lee, H.-M. Advanced glycosylation end products induce inducible nitric oxide synthase (iNOS) expression via a p38 MAPK-dependent pathway. Kidney Int. 2004, 65, 1664–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. The Nitration of Proteins, Lipids and DNA by Peroxynitrite Derivatives-Chemistry Involved and Biological Relevance. Stresses 2022, 2, 53–64. [Google Scholar] [CrossRef]
- Villa-Martínez, E.; López-Vaquera, S.R.; Alvarado-Coutiño, L.K.; Gámez-Méndez, A.M.; Ríos, A.; Escalante, B. Thromboxane-dependent coronary vasoconstriction in obese mice: Role of peroxynitrite. Prostaglandins Other Lipid Mediat. 2022, 160, 106631. [Google Scholar] [CrossRef]
- Jaganjac, M.; Tirosh, O.; Cohen, G.; Sasson, S.; Zarkovic, N. Reactive aldehydes-second messengers of free radicals in diabetes mellitus. Free Radic. Res. 2013, 47, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Jankovic, A.; Korac, A.; Srdic-Galic, B.; Buzadzic, B.; Otasevic, V.; Stancic, A.; Vucetic, M.; Markelic, M.; Velickovic, K.; Golic, I.; et al. Differences in the redox status of human visceral and subcutaneous adipose tissues-Relationships to obesity and metabolic risk. Metabolism. 2014, 63, 661–671. [Google Scholar] [CrossRef]
- Zhao, X.; Si, L.; Bian, J.; Pan, C.; Guo, W.; Qin, P.; Zhu, W.; Xia, Y.; Zhang, Q.; Wei, K. Adipose tissue macrophage-derived exosomes induce ferroptosis via glutathione synthesis inhibition by targeting SLC7A11 in obesity-induced cardiac injury. Free Radic. Biol. Med. 2022, 182, 232–245. [Google Scholar] [CrossRef]
- Korovila, I.; Höhn, A.; Jung, T.; Grune, T.; Ott, C. Reduced liver autophagy in high-fat diet induced liver steatosis in new zealand obese mice. Antioxidants 2021, 10, 501. [Google Scholar] [CrossRef]
- Al-Menhali, A.S.; Anderson, C.; Gourine, A.V.; Abramov, A.Y.; D’Souza, A.; Jaganjac, M. Proteomic Analysis of Cardiac Adaptation to Exercise by High Resolution Mass Spectrometry. Front. Mol. Biosci. 2021, 8, 723858. [Google Scholar] [CrossRef]
- Zarkovic, N.; Cipak, A.; Jaganjac, M.; Borovic, S.; Zarkovic, K. Pathophysiological relevance of aldehydic protein modifications. J. Proteomics 2013, 92, 239–247. [Google Scholar] [CrossRef]
- Zhang, X.-M.; Guo, L.; Huang, X.; Li, Q.-M.; Chi, M.-H. 4-Hydroxynonenal Regulates TNF-α Gene Transcription Indirectly via ETS1 and microRNA-29b in Human Adipocytes Induced From Adipose Tissue-Derived Stromal Cells. Anat. Rec. 2016, 299, 1145–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauck, A.K.; Zhou, T.; Upadhyay, A.; Sun, Y.; O’connor, M.B.; Chen, Y.; Bernlohr, D.A. Histone carbonylation is a redox-regulated epigenomic mark that accumulates with obesity and aging. Antioxidants 2020, 9, 1210. [Google Scholar] [CrossRef] [PubMed]
- Grimsrud, P.A.; Picklo, M.J.S.; Griffin, T.J.; Bernlohr, D.A. Carbonylation of adipose proteins in obesity and insulin resistance: Identification of adipocyte fatty acid-binding protein as a cellular target of 4-hydroxynonenal. Mol. Cell. Proteomics 2007, 6, 624–637. [Google Scholar] [CrossRef] [Green Version]
- Katunga, L.A.; Gudimella, P.; Efird, J.T.; Abernathy, S.; Mattox, T.A.; Beatty, C.; Darden, T.M.; Thayne, K.A.; Alwair, H.; Kypson, A.P.; et al. Obesity in a model of gpx4 haploinsufficiency uncovers a causal role for lipid-derived aldehydes in human metabolic disease and cardiomyopathy. Mol. Metab. 2015, 4, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Z.; Li, J.; Gu, D.; Li, S.; Shen, C.; Song, Z. Increased 4-Hydroxynonenal Formation Contributes to Obesity-Related Lipolytic Activation in Adipocytes. PLoS ONE 2013, 8, e70663. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Dou, X.; Gu, D.; Shen, C.; Yao, T.; Nguyen, V.; Braunschweig, C.; Song, Z. 4-Hydroxynonenal differentially regulates adiponectin gene expression and secretion via activating PPARγ and accelerating ubiquitin-proteasome degradation. Mol. Cell. Endocrinol. 2012, 349, 222–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauck, A.K.; Zhou, T.; Hahn, W.; Petegrosso, R.; Kuang, R.; Chen, Y.; Bernlohr, D.A. Obesity-induced protein carbonylation in murine adipose tissue regulates the DNA-binding domain of nuclear zinc finger proteins. J. Biol. Chem. 2018, 293, 13464–13476. [Google Scholar] [CrossRef] [Green Version]
- Albaugh, V.L.; Abumrad, N.N. Surgical treatment of obesity. F1000Research 2018, 7, 617. [Google Scholar] [CrossRef] [Green Version]
- Choromańska, B.; Myśliwiec, P.; Łuba, M.; Wojskowicz, P.; Dadan, J.; Myśliwiec, H.; Choromańska, K.; Zalewska, A.; Maciejczyk, M. A Longitudinal Study of the Antioxidant Barrier and Oxidative Stress in Morbidly Obese Patients after Bariatric Surgery. Does the Metabolic Syndrome Affect the Redox Homeostasis of Obese People? J. Clin. Med. 2020, 9, 976. [Google Scholar] [CrossRef] [Green Version]
- Soldo, I.; Kolak, T. Pretilost I Liječenje; Medicinska Naklada: Zagreb, Croatia, 2020; ISBN 978-953-176-976-1. [Google Scholar]
- Angrisani, L.; Santonicola, A.; Iovino, P.; Vitiello, A.; Zundel, N.; Buchwald, H.; Scopinaro, N. Bariatric Surgery and Endoluminal Procedures: IFSO Worldwide Survey 2014. Obes. Surg. 2017, 27, 2279–2289. [Google Scholar] [CrossRef]
- Weiner, S.; Sauerland, S.; Weiner, R.A.; Pomhoff, I. Quality of Life after Bariatric Surgery-Is There a Difference? Visc. Med. 2005, 21 (Suppl. 1), 34–36. [Google Scholar] [CrossRef]
- Lomanto, D.; Lee, W.-J.; Goel, R.; Lee, J.J.-M.; Shabbir, A.; So, J.B.Y.; Huang, C.-K.; Chowbey, P.; Lakdawala, M.; Sutedja, B.; et al. Bariatric surgery in Asia in the last 5 years (2005–2009). Obes. Surg. 2012, 22, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Spivak, H.; Abdelmelek, M.F.; Beltran, O.R.; Ng, A.W.; Kitahama, S. Long-term outcomes of laparoscopic adjustable gastric banding and laparoscopic Roux-en-Y gastric bypass in the United States. Surg. Endosc. 2012, 26, 1909–1919. [Google Scholar] [CrossRef]
- Tieu, K.; Allison, N.; Snyder, B.; Wilson, T.; Toder, M.; Wilson, E. Robotic-assisted Roux-en-Y gastric bypass: Update from 2 high-volume centers. Surg. Obes. Relat. Dis. Off. J. Am. Soc. Bariatr. Surg. 2013, 9, 284–288. [Google Scholar] [CrossRef]
- Buchwald, H.; Avidor, Y.; Braunwald, E.; Jensen, M.D.; Pories, W.; Fahrbach, K.; Schoelles, K. Bariatric surgery: A systematic review and meta-analysis. JAMA 2004, 292, 1724–1737. [Google Scholar] [CrossRef]
- Bekavac-Beslin, M. New surgical procedure for treatment of morbid obesity. Acta Chir. Croat. 2013, 10, 9–10. [Google Scholar] [CrossRef]
- Crea, N.; Pata, G.; Di Betta, E.; Greco, F.; Casella, C.; Vilardi, A.; Mittempergher, F. Long-term results of biliopancreatic diversion with or without gastric preservation for morbid obesity. Obes. Surg. 2011, 21, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Risstad, H.; Søvik, T.T.; Engström, M.; Aasheim, E.T.; Fagerland, M.W.; Olsén, M.F.; Kristinsson, J.A.; le Roux, C.W.; Bøhmer, T.; Birkeland, K.I.; et al. Five-year outcomes after laparoscopic gastric bypass and laparoscopic duodenal switch in patients with body mass index of 50 to 60: A randomized clinical trial. JAMA Surg. 2015, 150, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Hedberg, J.; Sundström, J.; Sundbom, M. Duodenal switch versus Roux-en-Y gastric bypass for morbid obesity: Systematic review and meta-analysis of weight results, diabetes resolution and early complications in single-centre comparisons. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2014, 15, 555–563. [Google Scholar] [CrossRef]
- Villarreal-Calderon, J.R.; Cuellar-Tamez, R.; Castillo, E.C.; Luna-Ceron, E.; García-Rivas, G.; Elizondo-Montemayor, L. Metabolic shift precedes the resolution of inflammation in a cohort of patients undergoing bariatric and metabolic surgery. Sci. Rep. 2021, 11, 12127. [Google Scholar] [CrossRef]
- Cornejo-Pareja, I.; Clemente-Postigo, M.; Tinahones, F.J. Metabolic and Endocrine Consequences of Bariatric Surgery. Front. Endocrinol. 2019, 10, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.; Lee, H.; Kaura, S.; Yip, J.; Sun, H.; Guan, L.; Han, W.; Ding, Y. Effect of Bariatric Surgery on Metabolic Diseases and Underlying Mechanisms. Biomolecules 2021, 11, 1582. [Google Scholar] [CrossRef] [PubMed]
- Mulla, C.M.; Middelbeek, R.J.W.; Patti, M.-E. Mechanisms of weight loss and improved metabolism following bariatric surgery. Ann. N. Y. Acad. Sci. 2018, 1411, 53–64. [Google Scholar] [CrossRef]
- Syn, N.L.; Cummings, D.E.; Wang, L.Z.; Lin, D.J.; Zhao, J.J.; Loh, M.; Koh, Z.J.; Chew, C.A.; Loo, Y.E.; Tai, B.C.; et al. Association of metabolic-bariatric surgery with long-term survival in adults with and without diabetes: A one-stage meta-analysis of matched cohort and prospective controlled studies with 174 772 participants. Lancet 2021, 397, 1830–1841. [Google Scholar] [CrossRef]
- Sudlow, A.; le Roux, C.W.; Pournaras, D.J. The metabolic benefits of different bariatric operations: What procedure to choose? Endocr. Connect. 2020, 9, R28–R35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, D.A.; Trac, J.Z.; Glazer, S.A.; Terenzi, D.C.; Quan, A.; Teoh, H.; Al-Omran, M.; Bhatt, D.L.; Mazer, C.D.; Rotstein, O.D.; et al. Vascular Risk Reduction in Obesity through Reduced Granulocyte Burden and Improved Angiogenic Monocyte Content following Bariatric Surgery. Cell Rep. Med. 2020, 1, 100018. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jin, J.; Chen, Y.; Chen, C.; Chen, Z.; Xu, L. Integrative analyses of biomarkers and pathways for adipose tissue after bariatric surgery. Adipocyte 2020, 9, 384–400. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-W.; Zhou, Y.; Zhou, P.-Y.; Zhang, T.-Y.; Hu, J.-Y.; Bai, X.-T. Protective Effects of Bariatric Surgery on Kidney Functions by Inhibiting Oxidative Stress Responses Through Activating PPARα in Rats With Diabetes. Front. Physiol. 2021, 12, 662666. [Google Scholar] [CrossRef]
- Uzun, H.; Zengin, K.; Taskin, M.; Aydin, S.; Simsek, G.; Dariyerli, N. Changes in leptin, plasminogen activator factor and oxidative stress in morbidly obese patients following open and laparoscopic Swedish adjustable gastric banding. Obes. Surg. 2004, 14, 659–665. [Google Scholar] [CrossRef]
- Jamialahmadi, T.; Reiner, Ž.; Alidadi, M.; Kroh, M.; Cardenia, V.; Xu, S.; Al-Rasadi, K.; Santos, R.D.; Sahebkar, A. The Effect of Bariatric Surgery on Circulating Levels of Oxidized Low-Density Lipoproteins Is Apparently Independent of Changes in Body Mass Index: A Systematic Review and Meta-Analysis. Oxid. Med. Cell. Longev. 2021, 2021, 4136071. [Google Scholar] [CrossRef]
- João Cabrera, E.; Valezi, A.C.; Delfino, V.D.A.; Lavado, E.L.; Barbosa, D.S. Reduction in plasma levels of inflammatory and oxidative stress indicators after Roux-en-Y gastric bypass. Obes. Surg. 2010, 20, 42–49. [Google Scholar] [CrossRef]
- Monzo-Beltran, L.; Vazquez-Tarragón, A.; Cerdà, C.; Garcia-Perez, P.; Iradi, A.; Sánchez, C.; Climent, B.; Tormos, C.; Vázquez-Prado, A.; Girbés, J.; et al. One-year follow-up of clinical, metabolic and oxidative stress profile of morbid obese patients after laparoscopic sleeve gastrectomy. 8-oxo-dG as a clinical marker. Redox Biol. 2017, 12, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Horn, R.C.; Gelatti, G.T.; Mori, N.C.; Tissiani, A.C.; Mayer, M.S.; Pereira, E.A.; Ross, M.; Moreira, P.R.; Bortolotto, J.W.; Felippin, T. Obesity, bariatric surgery and oxidative stress. Rev. Assoc. Med. Bras. 2017, 63, 229–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venâncio, F.A.; Almeida, L.A.; Zovico, P.V.; Barauna, V.G.; Miguel, G.P.S.; Pedrosa, R.G.; Haraguchi, F.K. Roux-en-Y Gastric Bypass and Sleeve Gastrectomy Differently Affect Oxidative Damage Markers and their Correlations with Body Parameters. Obes. Surg. 2021, 31, 1680–1687. [Google Scholar] [CrossRef] [PubMed]
- Choromańska, B.; Myśliwiec, P.; Łuba, M.; Wojskowicz, P.; Myśliwiec, H.; Choromańska, K.; Żendzian-Piotrowska, M.; Dadan, J.; Zalewska, A.; Maciejczyk, M. Impact of weight loss on the total antioxidant/oxidant potential in patients with morbid obesity—A longitudinal study. Antioxidants 2020, 9, 376. [Google Scholar] [CrossRef] [PubMed]
- Fejfer, K.; Buczko, P.; Niczyporuk, M.; Ładny, J.R.; Hady, H.R.; Knaś, M.; Waszkiel, D.; Klimiuk, A.; Zalewska, A.; Maciejczyk, M. Oxidative Modification of Biomolecules in the Nonstimulated and Stimulated Saliva of Patients with Morbid Obesity Treated with Bariatric Surgery. Biomed Res. Int. 2017, 2017, 4923769. [Google Scholar] [CrossRef] [Green Version]
- Skrzep-Poloczek, B.; Poloczek, J.; Chełmecka, E.; Dulska, A.; Romuk, E.; Idzik, M.; Kazura, W.; Nabrdalik, K.; Gumprecht, J.; Jochem, J.; et al. The Oxidative Stress Markers in the Erythrocytes and Heart Muscle of Obese Rats: Relate to a High-Fat Diet but Not to DJOS Bariatric Surgery. Antioxidants 2020, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Lopes Pinto, S.; da Silva, D.C.G.; Bressan, J. Absolute and Relative Changes in Ultra-processed Food Consumption and Dietary Antioxidants in Severely Obese Adults 3 Months After Roux-en-Y Gastric Bypass. Obes. Surg. 2019, 29, 1810–1815. [Google Scholar] [CrossRef]
- Onyango, A.N. Lipid Peroxidation as a Link between Unhealthy Diets and the Metabolic Syndrome. In Accenting Lipid Peroxidation; Atukeren, P., Ed.; IntechOpen: Rijeka, Croatia, 2021. [Google Scholar]
- Macho-González, A.; Garcimartín, A.; López-Oliva, M.E.; Bastida, S.; Benedí, J.; Ros, G.; Nieto, G.; Sánchez-Muniz, F.J. Can Meat and Meat-Products Induce Oxidative Stress? Antioxidants 2020, 9, 638. [Google Scholar] [CrossRef]
- Taherkhani, S.; Suzuki, K.; Ruhee, R.T. A brief overview of oxidative stress in adipose tissue with a therapeutic approach to taking antioxidant supplements. Antioxidants 2021, 10, 594. [Google Scholar] [CrossRef]
- Tun, S.; Spainhower, C.J.; Cottrill, C.L.; Lakhani, H.V.; Pillai, S.S.; Dilip, A.; Chaudhry, H.; Shapiro, J.I.; Sodhi, K. Therapeutic Efficacy of Antioxidants in Ameliorating Obesity Phenotype and Associated Comorbidities. Front. Pharmacol. 2020, 11, 1234. [Google Scholar] [CrossRef] [PubMed]
- Jumbe, S.; Hamlet, C.; Meyrick, J. Psychological Aspects of Bariatric Surgery as a Treatment for Obesity. Curr. Obes. Rep. 2017, 6, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, Y.C.; Huang, C.K.; Tai, C.M. Psychiatric aspects of bariatric surgery. Curr. Opin. Psychiatry 2014, 27, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.; Kang, S.; Lee, Y.; Rosenblat, J.D.; Brietzke, E.; Zuckerman, H.; McIntyre, R.S. The long-term effect of bariatric surgery on depression and anxiety. J. Affect. Disord. 2019, 246, 886–894. [Google Scholar] [CrossRef]
- Kubik, J.F.; Gill, R.S.; Laffin, M.; Karmali, S. The impact of bariatric surgery on psychological health. J. Obes. 2013, 2013, 837989. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soldo, A.M.; Soldo, I.; Karačić, A.; Konjevod, M.; Perkovic, M.N.; Glavan, T.M.; Luksic, M.; Žarković, N.; Jaganjac, M. Lipid Peroxidation in Obesity: Can Bariatric Surgery Help? Antioxidants 2022, 11, 1537. https://doi.org/10.3390/antiox11081537
Soldo AM, Soldo I, Karačić A, Konjevod M, Perkovic MN, Glavan TM, Luksic M, Žarković N, Jaganjac M. Lipid Peroxidation in Obesity: Can Bariatric Surgery Help? Antioxidants. 2022; 11(8):1537. https://doi.org/10.3390/antiox11081537
Chicago/Turabian StyleSoldo, Ana Maria, Ivo Soldo, Andrija Karačić, Marcela Konjevod, Matea Nikolac Perkovic, Tanja Matijevic Glavan, Martina Luksic, Neven Žarković, and Morana Jaganjac. 2022. "Lipid Peroxidation in Obesity: Can Bariatric Surgery Help?" Antioxidants 11, no. 8: 1537. https://doi.org/10.3390/antiox11081537
APA StyleSoldo, A. M., Soldo, I., Karačić, A., Konjevod, M., Perkovic, M. N., Glavan, T. M., Luksic, M., Žarković, N., & Jaganjac, M. (2022). Lipid Peroxidation in Obesity: Can Bariatric Surgery Help? Antioxidants, 11(8), 1537. https://doi.org/10.3390/antiox11081537