
Deferoxamine Supplementation Abolished Iron-Related Toxicity of Ilex paraguariensis Extract: Behavioral and Biochemical Evaluation in Adult Zebrafish (Danio rerio)

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Strategy
2.2. Ilex paraguariensis Extract Preparation, Microencapsulation, and Efficiency of Encapsulation Rate
2.3. Fish
2.4. Standards
2.5. Behavioral Analyses
2.6. Biochemical Analyzes
2.7. Statistics
3. Results
3.1. Microencapsulation Evaluation by Scanning Electron Microscopy and Efficiency Determination
3.2. Behavior
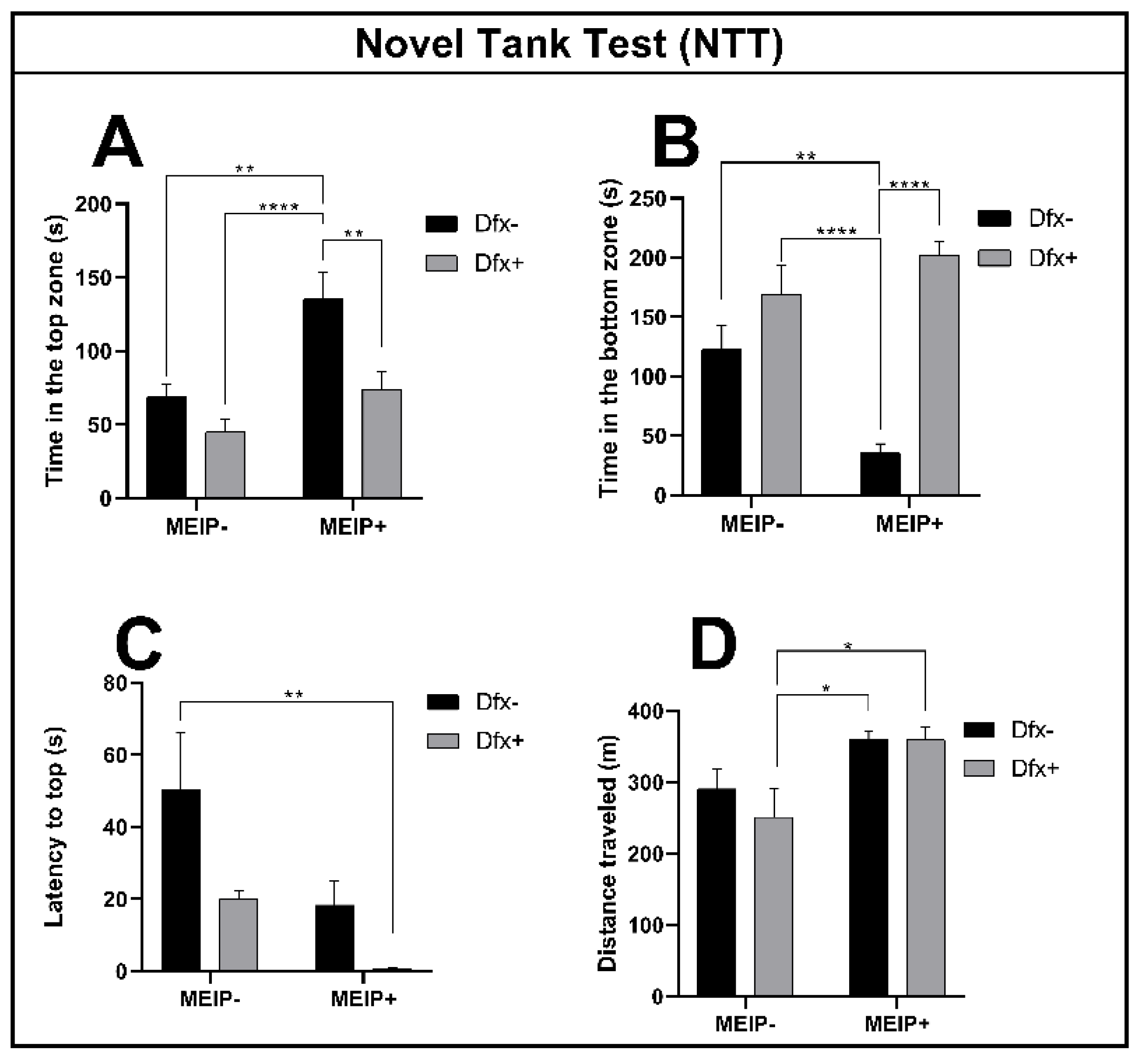
3.2.1. Novel Tank Test (NTT)
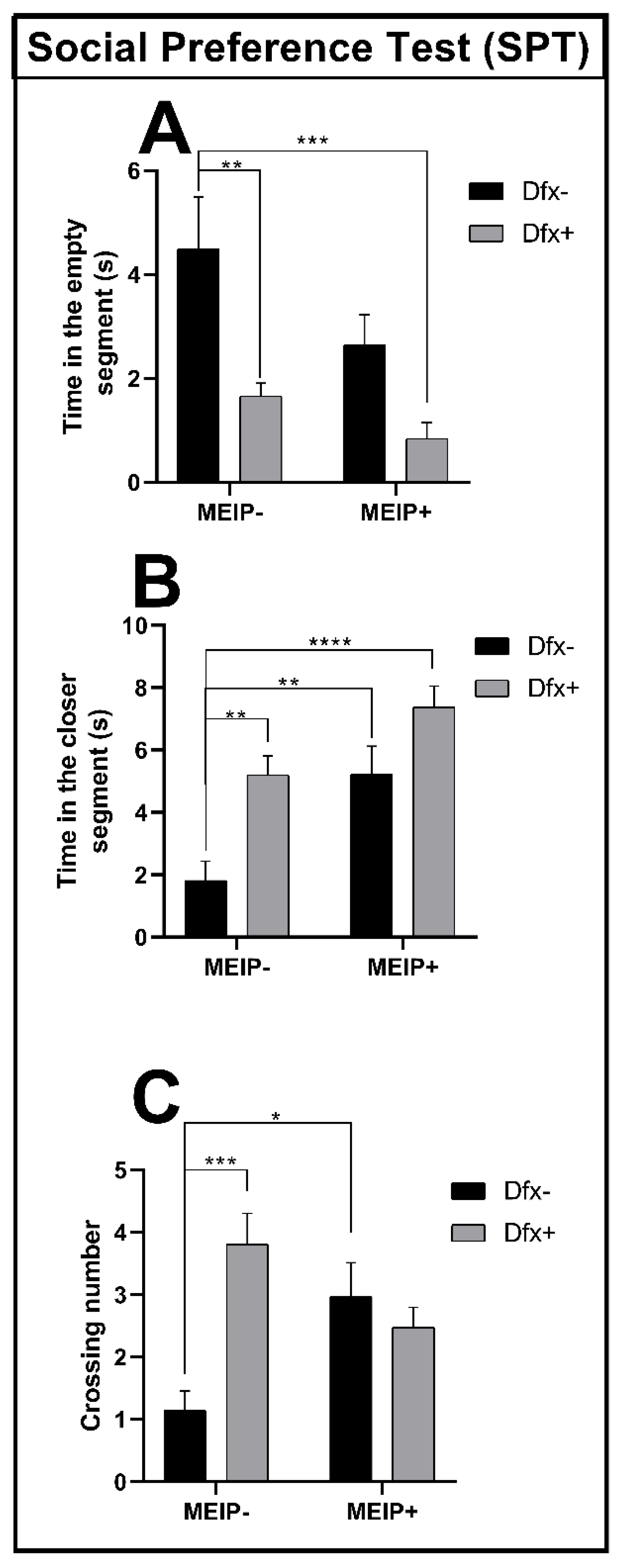
3.2.2. Social Preference Test (SPT)
3.3. Biochemical
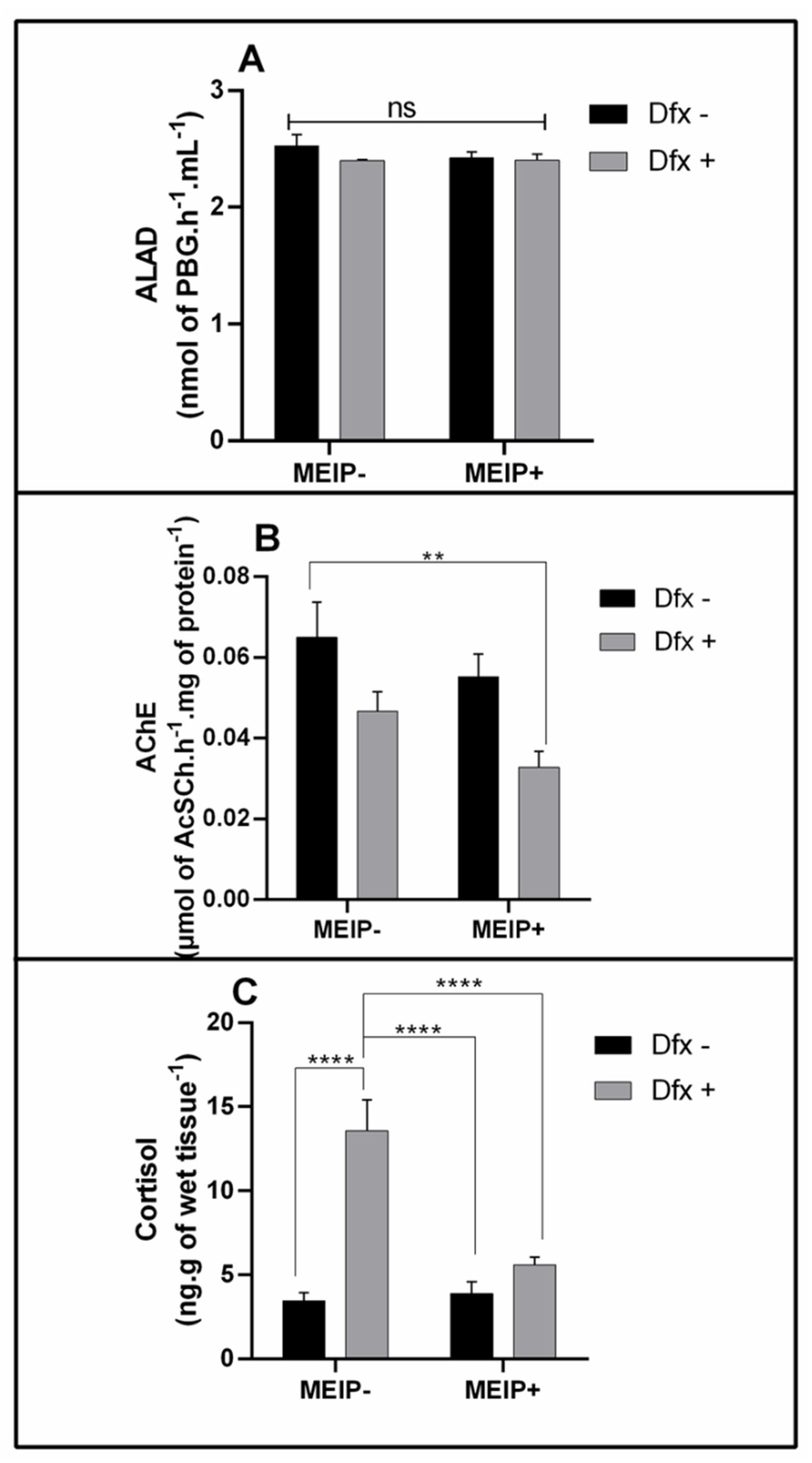
3.3.1. ALAD Activity
3.3.2. AChE Activity
3.3.3. Cortisol Levels
3.3.4. Antioxidant Status
Antioxidant Enzymes
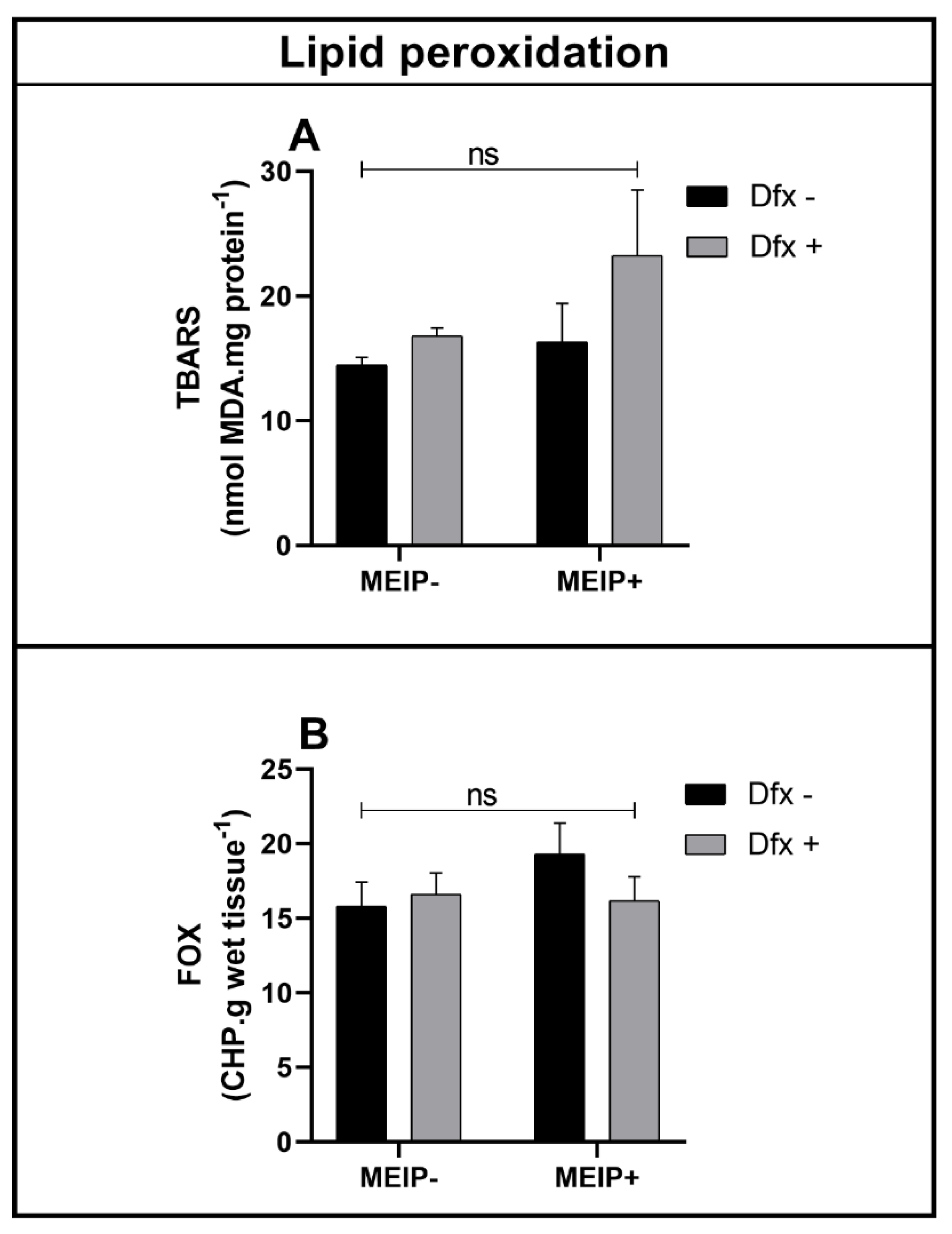
Lipid Peroxidation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vieira, M.A.; Maraschin, M.; Pagliosa, C.M.; Podestá, R.; De Simas, K.N.; Rockenbach, I.I.; Amboni, R.D.; Amboni, D.M.C.; Amante, E.R. Phenolic Acids and Methylxanthines Composition and Antioxidant Properties of Mate (Ilex Paraguariensis) Residue. J. Food Sci. 2010, 75, C280–C285. [Google Scholar] [CrossRef] [PubMed]
- Piovezan-Borges, A.C.; Valério-Júnior, C.; Gonçalves, I.L.; Mielniczki-Pereira, A.A.; Valduga, A.T. Antioxidant Potential of Yerba Mate (Ilex Paraguariensis St. Hil.) Extracts in Saccharomyces Cerevisae Deficient in Oxidant Defense Genes. Braz. J. Biol. 2016, 76, 539–544. [Google Scholar] [CrossRef] [Green Version]
- de Mejía, E.G.; Song, Y.S.; Heck, C.I.; Ramírez-Mares, M. Yerba Mate Tea (Ilex Paraguariensis): Phenolics, Antioxidant Capacity and in Vitro Inhibition of Colon Cancer Cell Proliferation. J. Funct. Foods 2010, 2, 23–34. [Google Scholar] [CrossRef]
- Heck, C.I.; Schmalko, M.; Gonzalez de Mejia, E. Effect of Growing and Drying Conditions on the Phenolic Composition of Mate Teas (Ilex Paraguariensis). J. Agric. Food Chem. 2008, 56, 8394–8403. [Google Scholar] [CrossRef] [PubMed]
- Tamagno, W.A.; Santini, W.; Alves, C.; Vanin, A.P.; Pompermaier, A.; Bilibio, D.; Sutorillo, N.T.; Kaizer, R.R.; Barcellos, L.J.G. Neuroprotective and Antioxidant Effects of Pitaya Fruit on Cu-Induced Stress in Adult Zebrafish. J. Food Biochem. 2022, e14147. [Google Scholar] [CrossRef] [PubMed]
- De Campos, R.M.L.; Hierro, E.; Ordóñez, J.A.; Bertol, T.M.; Terra, N.N.; De la Hoz, L. Fatty Acid and Volatile Compounds from Salami Manufactured with Yerba Mate (Ilex Paraguariensis) Extract and Pork Back Fat and Meat from Pigs Fed on Diets with Partial Replacement of Maize with Rice Bran. Food Chem. 2007, 103, 1159–1167. [Google Scholar] [CrossRef]
- Bortoli, P.M.; Alves, C.; Costa, E.; Vanin, A.P.; Sofiatti, J.R.; Siqueira, D.P.; Dallago, R.M.; Treichel, H.; Vargas, G.D.L.P.; Kaizer, R.R. Ilex Paraguariensis: Potential Antioxidant on Aluminium Toxicity, in an Experimental Model of Alzheimer’s Disease. J. Inorg. Biochem. 2018, 181, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Berté, K.A.; Beux, M.R.; Spada, P.K.; Salvador, M.; Hoffmann-Ribani, R. Chemical Composition and Antioxidant Activity of Yerba-Mate (Ilex Paraguariensis A. St.-Hil., Aquifoliaceae) Extract as Obtained by Spray Drying. J. Agric. Food Chem. 2011, 59, 5523–5527. [Google Scholar] [CrossRef]
- Ersus, S.; Yurdagel, U. Microencapsulation of Anthocyanin Pigments of Black Carrot (Daucus Carota L.) by Spray Drier. J. Food Eng. 2007, 80, 805–812. [Google Scholar] [CrossRef]
- Tamagno, W.A.; Vanin, A.P.; Sutorillo, N.T.; Bilibio, D.; Dada, R.A.; Colla, L.M.; Zamberlan, D.C.; Kaizer, R.R.; Barcellos, L.J.G. Fruit Extract of Red Pitaya (Hylocereus Undatus) Prevents and Reverses Stress-Induced Impairments in the Cholinergic and Antioxidant Systems of Caenorhabditis Elegans. J. Food Biochem. 2021, 46, e13981. [Google Scholar] [CrossRef]
- Seneviratne, M.; Rajakaruna, N.; Rizwan, M.; Madawala, H.; Ok, Y.S.; Vithanage, M. Heavy Metal-Induced Oxidative Stress on Seed Germination and Seedling Development: A Critical Review. Environ. Geochem. Health 2019, 41, 1813–1831. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.S.; Kadier, A.; Malyan, S.K.; Ahmad, A.; Bishnoi, N.R. Phytoremediation and Rhizoremediation: Uptake, Mobilization and Sequestration of Heavy Metals by Plants. In Plant-Microbe Interactions in Agro-Ecological Perspectives; Springer: Singapore, 2017; pp. 367–394. [Google Scholar]
- Amari, T.; Ghnaya, T.; Abdelly, C. Nickel, Cadmium and Lead Phytotoxicity and Potential of Halophytic Plants in Heavy Metal Extraction. S. Afr. J. Bot. 2017, 111, 99–110. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy Metal Hyperaccumulating Plants: How and Why Do They Do It? And What Makes Them so Interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Gaiad, S.; Rakocevic, M.; Reissmann, C.B. N Sources Affect Growth, Nutrient Content, and Net Photosynthesis in Mate (Ilex Paraguariensis St. Hil.). Braz. Arch. Biol. Technol. 2006, 49, 689–697. [Google Scholar] [CrossRef]
- Umemura, M.; Kim, J.-H.; Aoyama, H.; Hoshino, Y.; Fukumura, H.; Nakakaji, R.; Sato, I.; Ohtake, M.; Akimoto, T.; Narikawa, M. The Iron Chelating Agent, Deferoxamine Detoxifies Fe (Salen)-Induced Cytotoxicity. J. Pharmacol. Sci. 2017, 134, 203–210. [Google Scholar] [CrossRef]
- Nikaido, Y.; Midorikawa, Y.; Furukawa, T.; Shimoyama, S.; Takekawa, D.; Kitayama, M.; Ueno, S.; Kushikata, T.; Hirota, K. The Role of Neutrophil Gelatinase-Associated Lipocalin and Iron Homeostasis in Object Recognition Impairment in Aged Sepsis-Survivor Rats. Sci. Rep. 2022, 12, 1–10. [Google Scholar] [CrossRef]
- Avdesh, A.; Chen, M.; Martin-Iverson, M.T.; Mondal, A.; Ong, D.; Rainey-Smith, S.; Taddei, K.; Lardelli, M.; Groth, D.M.; Verdile, G. Regular Care and Maintenance of a Zebrafish (Danio Rerio) Laboratory: An Introduction. JoVE (J. Vis. Exp.) 2012, 69, e4196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Mascaraque, L.G.; Sipoli, C.C.; de La Torre, L.G.; López-Rubio, A. Microencapsulation Structures Based on Protein-Coated Liposomes Obtained through Electrospraying for the Stabilization and Improved Bioaccessibility of Curcumin. Food Chem. 2017, 233, 343–350. [Google Scholar] [CrossRef]
- Ghorbanzade, T.; Jafari, S.M.; Akhavan, S.; Hadavi, R. Nano-Encapsulation of Fish Oil in Nano-Liposomes and Its Application in Fortification of Yogurt. Food Chem. 2017, 216, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Dametto, F.S.; Fior, D.; Idalencio, R.; Rosa, J.G.S.; Fagundes, M.; Marqueze, A.; Barreto, R.E.; Piato, A.; Barcellos, L.J. Feeding Regimen Modulates Zebrafish Behavior. PeerJ 2018, 6, e5343. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R. Zebra Fish: An Uncharted Behavior Genetic Model. Behav. Genet. 2003, 33, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Goth, L. A Simple Method for Determination of Serum Catalase Activity and Revision of Reference Range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Johansson, L.H.; Borg, L.H. A Spectrophotometric Method for Determination of Catalase Activity in Small Tissue Samples. Anal. Biochem. 1988, 174, 331–336. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-Transferases: The First Enzymatic Step in Mercapturic Acid Formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Habig, W.H.; Jakoby, W.B. [51] Assays for Differentiation of Glutathione S-Transferases. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1981; Volume 77, pp. 398–405. [Google Scholar]
- Misra, H.P.; Fridovich, I. The Role of Superoxide Anion in the Autoxidation of Epinephrine and a Simple Assay for Superoxide Dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- de Quadros Camargo, C.; Mocellin, M.C.; Brunetta, H.S.; Chagas, T.R.; de Souza Fabre, M.E.; de Moraes Trindade, E.B.S.; da Silva, E.L.; Nunes, E.A. Fish Oil Decreases the Severity of Treatment-Related Adverse Events in Gastrointestinal Cancer Patients Undergoing Chemotherapy: A Randomized, Placebo-Controlled, Triple-Blind Clinical Trial. Clin. Nutr. ESPEN 2019, 31, 61–70. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres Jr, V.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Kaizer, R.R.; Correa, M.C.; Gris, L.R.S.; Da Rosa, C.S.; Bohrer, D.; Morsch, V.M.; Schetinger, M.R.C. Effect of Long-Term Exposure to Aluminum on the Acetylcholinesterase Activity in the Central Nervous System and Erythrocytes. Neurochem. Res. 2008, 33, 2294–2301. [Google Scholar] [CrossRef]
- Sassa, S. Delta-Aminolevulinic Acid Dehydratase Assay. Enzyme 1982, 28, 133–145. [Google Scholar] [CrossRef]
- Moore, K.; Roberts, L.J. Measurement of Lipid Peroxidation. Free. Radic. Res. 1998, 28, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.C.; Aparício, R.; Blanco-Vives, B.; Chereguini, O.; Martín, I.; Javier Sánchez-Vazquez, F. Endocrine (Plasma Cortisol and Glucose) and Behavioral (Locomotor and Self-Feeding Activity) Circadian Rhythms in Senegalese Sole (Solea Senegalensis Kaup 1858) Exposed to Light/Dark Cycles or Constant Light. Fish Physiol. Biochem. 2013, 39, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Sink, T.D.; Lochmann, R.T.; Fecteau, K.A. Validation, Use, and Disadvantages of Enzyme-Linked Immunosorbent Assay Kits for Detection of Cortisol in Channel Catfish, Largemouth Bass, Red Pacu, and Golden Shiners. Fish Physiol. Biochem. 2008, 34, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Bastos, D.H.; Saldanha, L.A.; Catharino, R.R.; Sawaya, A.; Cunha, I.B.; Carvalho, P.O.; Eberlin, M.N. Phenolic Antioxidants Identified by ESI-MS from Yerba Mate (Ilex Paraguariensis) and Green Tea (Camelia Sinensis) Extracts. Molecules 2007, 12, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Mateos, R.; Baeza, G.; Sarriá, B.; Bravo, L. Improved LC-MSn Characterization of Hydroxycinnamic Acid Derivatives and Flavonols in Different Commercial Mate (Ilex Paraguariensis) Brands. Quantification of Polyphenols, Methylxanthines, and Antioxidant Activity. Food Chem. 2018, 241, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, J.S.; Deolindo, C.T.; Hoffmann, J.F.; Chaves, F.C.; do Prado-Silva, L.; Sant’Ana, A.S.; Azevedo, L.; do Carmo, M.A.V.; Granato, D. Optimized Camellia Sinensis Var. Sinensis, Ilex Paraguariensis, and Aspalathus Linearis Blend Presents High Antioxidant and Antiproliferative Activities in a Beverage Model. Food Chem. 2018, 254, 348–358. [Google Scholar] [CrossRef]
- Teselkin, Y.O.; Babenkova, I.V.; Pavlova, L.A.; Lee, A.; Kochetova, A.A.; Osipov, A.N.; Vladimirov, Y.A. The Antioxidant Capacity of Aqueous Extracts from Yerba Mate (Ilex Paraguariensis). Biophysics 2021, 66, 125–132. [Google Scholar] [CrossRef]
- Fenoglio, D.; Soto Madrid, D.; Alarcón Moyano, J.; Ferrario, M.; Guerrero, S.; Matiacevich, S. Active Food Additive Based on Encapsulated Yerba Mate (Ilex Paraguariensis) Extract: Effect of Drying Methods on the Oxidative Stability of a Real Food Matrix (Mayonnaise). J. Food Sci. Technol. 2021, 58, 1574–1584. [Google Scholar] [CrossRef]
- Pilatti-Riccio, D.; Dos Santos, D.F.; Meinhart, A.D.; Knapp, M.A.; dos Santos Hackbart, H.C.; Pinto, V.Z. Impact of the Use of Saccharides in the Encapsulation of Ilex Paraguariensis Extract. Food Res. Int. 2019, 125, 108600. [Google Scholar] [CrossRef]
- Vargas, B.K.; Frota, E.G.; dos Santos, L.F.; Gutkoski, J.P.; Lopes, S.T.; Bertol, C.D.; Bertolin, T.E. Yerba Mate (Ilex Paraguariensis) Microparticles Modulate Antioxidant Markers in the Plasma and Brains of Rats. Food Biosci. 2021, 41, 100999. [Google Scholar] [CrossRef]
- Sablania, V.; Bosco, S.J.D. Optimization of Spray Drying Parameters for Murraya Koenigii (Linn) Leaves Extract Using Response Surface Methodology. Powder Technol. 2018, 335, 35–41. [Google Scholar] [CrossRef]
- Kim, J.-J.; Kim, Y.-S.; Kumar, V. Heavy Metal Toxicity: An Update of Chelating Therapeutic Strategies. J. Trace Elem. Med. Biol. 2019, 54, 226–231. [Google Scholar] [CrossRef]
- Nasrallah, G.K.; Salem, R.; Da’as, S.; Al-Jamal, O.L.A.; Scott, M.; Mustafa, I. Biocompatibility and Toxicity of Novel Iron Chelator Starch-Deferoxamine (S-DFO) Compared to Zinc Oxide Nanoparticles to Zebrafish Embryo: An Oxidative Stress Based Apoptosis, Physicochemical and Neurological Study Profile. Neurotoxicol. Teratol. 2019, 72, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Olivari, I.; Paz, S.; Gutiérrez, Á.J.; González-Weller, D.; Hardisson, A.; Sagratini, G.; Rubio, C. Macroelement, Trace Element, and Toxic Metal Levels in Leaves and Infusions of Yerba Mate (Ilex Paraguariensis). Environ. Sci. Pollut. Res. 2020, 27, 21341–21352. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, V.E.; Vanin, A.P.; Concato, A.C.; Tamagno, W.A.; Treichel, H.; Kaizer, R.R. The Toxicological Paradigms of Aluminum and the Etiology of Alzheimer’s Disease in the Nematode Caenorhabditis Elegans. Environ. Qual. Manag. 2019, 29, 147–154. [Google Scholar] [CrossRef]
- Alves, C.; Zancan, N.P.; Concato, A.C.; Vanin, A.P.; Tamagno, W.A.; Tessaro, D.; Kaizer, R.R. Deferoxamine Pretreatment Inhibits Metal Neurotoxicity in Caenorhabditis Elegans. Environ. Qual. Manag. 2019, 29, 121–127. [Google Scholar] [CrossRef]
- Holden, P.; Nair, L.S. Deferoxamine: An Angiogenic and Antioxidant Molecule for Tissue Regeneration. Tissue Eng. Part B: Rev. 2019, 25, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kysil, E.V.; Meshalkina, D.A.; Frick, E.E.; Echevarria, D.J.; Rosemberg, D.B.; Maximino, C.; Lima, M.G.; Abreu, M.S.; Giacomini, A.C.; Barcellos, L.J. Comparative Analyses of Zebrafish Anxiety-like Behavior Using Conflict-Based Novelty Tests. Zebrafish 2017, 14, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.D. Heme Biosynthesis and the Porphyrias. Mol. Genet. Metab. 2019, 128, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Roh, Y.J.; Han, S.-J.; Park, I.; Lee, H.M.; Ok, Y.S.; Lee, B.C.; Lee, S.-R. Role of Selenoproteins in Redox Regulation of Signaling and the Antioxidant System: A Review. Antioxidants 2020, 9, 383. [Google Scholar] [CrossRef]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. The Copper-Zinc Superoxide Dismutase Activity in Selected Diseases. Eur. J. Clin. Investig. 2019, 49, e13036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendez, J.I.; Nicholson, W.J.; Taylor, W.R. SOD Isoforms and Signaling in Blood Vessels: Evidence for the Importance of ROS Compartmentalization. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 887–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signorella, S.; Palopoli, C.; Ledesma, G. Rationally Designed Mimics of Antioxidant Manganoenzymes: Role of Structural Features in the Quest for Catalysts with Catalase and Superoxide Dismutase Activity. Coord. Chem. Rev. 2018, 365, 75–102. [Google Scholar] [CrossRef]
- Vinagre, C.; Madeira, D.; Narciso, L.; Cabral, H.N.; Diniz, M. Effect of Temperature on Oxidative Stress in Fish: Lipid Peroxidation and Catalase Activity in the Muscle of Juvenile Seabass, Dicentrarchus Labrax. Ecol. Indic. 2012, 23, 274–279. [Google Scholar] [CrossRef]
- Spasojević, I.; Mojović, M.; Stević, Z.; Spasić, S.D.; Jones, D.R.; Morina, A.; Spasić, M.B. Bioavailability and Catalytic Properties of Copper and Iron for Fenton Chemistry in Human Cerebrospinal Fluid. Redox Rep. 2010, 15, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Savastano, M.; Boscaro, F.; Bianchi, A. Metal Coordination Properties of a Chromophoric Desferrioxamine (DFO) Derivative: Insight on the Coordination Stoichiometry and Thermodynamic Stability of Zr4+ Complexes. Molecules 2021, 27, 184. [Google Scholar] [CrossRef] [PubMed]
- Rudneva, I.I.; Kuzminova, N.S.; Skuratovskaya, E.N. Glutathione-S-Transferase Activity in Tissues of Black Sea Fish Species. Asian J. Exp. Biol. Sci. 2010, 1, 141–150. [Google Scholar]
- Marcon, M.; Mocelin, R.; Sachett, A.; Siebel, A.M.; Herrmann, A.P.; Piato, A. Enriched Environment Prevents Oxidative Stress in Zebrafish Submitted to Unpredictable Chronic Stress. PeerJ 2018, 6, e5136. [Google Scholar] [CrossRef]
- Pérez, J.M.; Maldonado, M.E.; Rojano, B.A.; Alzate, F.; Sáez, J.; Cardona, W. Comparative Antioxidant, Antiproliferative and Apoptotic Effects of Ilex Laurina and Ilex Paraguariensis on Colon Cancer Cells. Trop. J. Pharm. Res. 2014, 13, 1279–1286. [Google Scholar] [CrossRef] [Green Version]
- Santetti, G.S.; Dacoreggio, M.V.; Silva, A.C.M.; Biduski, B.; Bressiani, J.; Oro, T.; de Francisco, A.; Gutkoski, L.C.; Amboni, R.D. de M.C. Effect of Yerba Mate (Ilex Paraguariensis) Leaves on Dough Properties, Antioxidant Activity, and Bread Quality Using Whole Wheat Flour. J. Food Sci. 2021, 86, 4354–4364. [Google Scholar] [CrossRef]
- Tamagno, W.A.; Santini, W.; Dos Santos, A.; Alves, C.; Bilibio, D.; Sutorillo, N.T.; Zamberlan, D.C.; Kaizer, R.R.; Barcellos, L.J.G. Pitaya Fruit Extract Ameliorates the Healthspan on Copper-Induced Toxicity of Caenorhabditis Elegans. J. Food Biochem. 2022, 43(3), e14050. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Figure | F Value | p Value | |
|---|---|---|---|---|
| Novel Tank Test | ||||
| Time spent in the top zone | 2A | Interaction | F1,44 = 2.325 | 0.1345 |
| Herb effect | F1,44 = 14.98 | 0.0004 | ||
| Dfx effect | F1,44 = 12.22 | 0.0011 | ||
| Time spent in the bottom zone | 2B | Interaction | F1,44 = 11.98 | 0.0012 |
| Herb effect | F1,44 = 2.528 | 0.1190 | ||
| Dfx effect | F1,44 = 38.38 | <0.0001 | ||
| Latency to top | 2C | Interaction | F1,44 = 0.505 | 0.4811 |
| Herb effect | F1,44 = 8.444 | 0.0057 | ||
| Dfx effect | F1,44 = 7.353 | 0.0095 | ||
| Distance traveled | 2D | Interaction | F1,44 = 0.531 | 0.4699 |
| Herb effect | F1,44 = 10.94 | 0.0019 | ||
| Dfx effect | F1,44 = 0.523 | 0.4654 | ||
| Social Preference Test | ||||
| Time spent at the conspecific segment | 3A | Interaction | F1,52 = 0.7465 | 0.3915 |
| Herb effect | F1,52 = 15.26 | 0.0003 | ||
| Dfx effect | F1,52 = 15.05 | 0.0003 | ||
| Time far from the conspecific segment | 3B | Interaction | F1,52 = 0.7059 | 0.4047 |
| Herb effect | F1,52 = 4.714 | 0.0345 | ||
| Dfx effect | F1,52 = 14.37 | 0.0004 | ||
| Crossing number | 3C | Interaction | F1,52 = 13.45 | 0.0006 |
| Herb effect | F1,52 = 0.3301 | 0.5681 | ||
| Dfx effect | F1,52 = 6.344 | 0.0149 | ||
| Biochemical parameters | ||||
| ALAD activity | 4A | Interaction | F1,32 = 0.8297 | 0.3692 |
| Herb effect | F1,32 = 0.6972 | 0.4099 | ||
| Dfx effect | F1,32 = 1.475 | 0.2334 | ||
| AChE activity | 4B | Interaction | F1,56 = 0.1121 | 0.7390 |
| Herb effect | F1,56 = 3.756 | 0.0577 | ||
| Dfx effect | F1,56 = 11.25 | 0.0014 | ||
| Cortisol levels | 4C | Interaction | F1,72 = 16.19 | 0.0001 |
| Herb effect | F1,72 = 12.98 | 0.0006 | ||
| Dfx effect | F1,72 = 31.83 | <0.0001 | ||
| SOD activity | 5A | Interaction | F1,44 = 17.08 | 0.0002 |
| Herb effect | F1,44 = 0.7868 | 0.3799 | ||
| Dfx effect | F1,44 = 5.858 | 0.0197 | ||
| CAT activity | 5B | Interaction | F1,40 = 41.80 | <0.0001 |
| Herb effect | F1,40 = 2.382 | 0.1306 | ||
| Dfx effect | F1,40 = 5.554 | 0.0234 | ||
| GST activity | 5C | Interaction | F1,60 = 0.07510 | 0.7850 |
| Herb effect | F1,60 = 2.094 | 0.1531 | ||
| Dfx effect | F1,60 = 0.5466 | 0.4626 | ||
| FOX brain | Interaction | F1,36 = 1.334 | 0.2557 | |
| Herb effect | F1,36 = 0.8083 | 0.3746 | ||
| Dfx effect | F1,36 = 0.4564 | 0.5036 | ||
| TBARS brain | Interaction | F1,40 = 0.5606 | 0.4584 | |
| Herb effect | F1,40 = 1.809 | 0.1862 | ||
| Dfx effect | F1,40 = 2.259 | 0.1407 | ||
| TBARS whole body | Interaction | F1,4 0 = 7.321 | 0.0100 | |
| Herb effect | F1,40 = 135.6 | <0.0001 | ||
| Dfx effect | F1,40 = 34.19 | <0.0001 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamagno, W.A.; Alves, C.; Tessaro, D.; Sutorillo, N.T.; Santin, W.; Barcellos, L.J.G. Deferoxamine Supplementation Abolished Iron-Related Toxicity of Ilex paraguariensis Extract: Behavioral and Biochemical Evaluation in Adult Zebrafish (Danio rerio). Antioxidants 2022, 11, 1507. https://doi.org/10.3390/antiox11081507
Tamagno WA, Alves C, Tessaro D, Sutorillo NT, Santin W, Barcellos LJG. Deferoxamine Supplementation Abolished Iron-Related Toxicity of Ilex paraguariensis Extract: Behavioral and Biochemical Evaluation in Adult Zebrafish (Danio rerio). Antioxidants. 2022; 11(8):1507. https://doi.org/10.3390/antiox11081507
Chicago/Turabian StyleTamagno, Wagner Antonio, Carla Alves, Diego Tessaro, Nathália Tafarel Sutorillo, Wallace Santin, and Leonardo José Gil Barcellos. 2022. "Deferoxamine Supplementation Abolished Iron-Related Toxicity of Ilex paraguariensis Extract: Behavioral and Biochemical Evaluation in Adult Zebrafish (Danio rerio)" Antioxidants 11, no. 8: 1507. https://doi.org/10.3390/antiox11081507
APA StyleTamagno, W. A., Alves, C., Tessaro, D., Sutorillo, N. T., Santin, W., & Barcellos, L. J. G. (2022). Deferoxamine Supplementation Abolished Iron-Related Toxicity of Ilex paraguariensis Extract: Behavioral and Biochemical Evaluation in Adult Zebrafish (Danio rerio). Antioxidants, 11(8), 1507. https://doi.org/10.3390/antiox11081507