
Effects and Mechanisms of Curcumin for the Prevention and Management of Cancers: An Updated Review

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
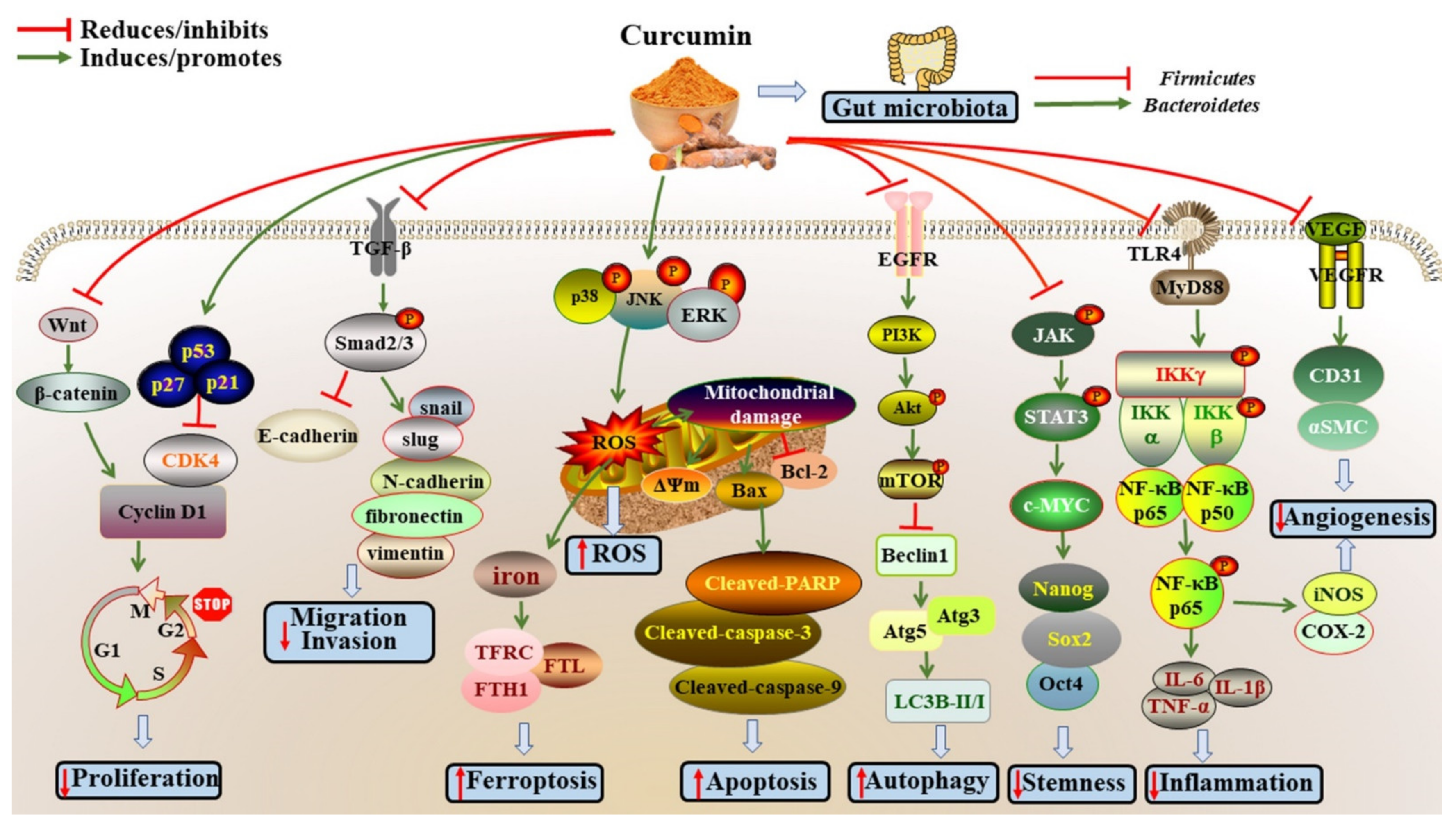
2. Effects and Mechanisms of Curcumin on Cancers
2.1. Inhibiting Cancer Cell Proliferation
2.2. Inhibiting Invasion and Migration
2.3. Inducing Cell Apoptosis
2.4. Inducing Autophagy
2.5. Suppressing Cancer Cell Stemness
2.6. Increasing ROS Production
2.7. Effects on Gut Microbiota
2.8. Adjuvant Therapy for Cancers
2.9. Other Mechanisms
3. Results from Clinical Trials
4. Enhancing Curcumin Bioavailability
5. Safety of Curcumin
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Fatiregun, O.A.; Lasebikan, N. High burden of postoperative cancer mortality in LMICs. Lancet 2021, 397, 349–351. [Google Scholar] [CrossRef]
- Hsu, H.C.; Tsai, S.Y.; Wu, S.L.; Jeang, S.R.; Ho, M.Y.; Liou, W.S.; Chiang, A.J.; Chang, T.H. Longitudinal perceptions of the side effects of chemotherapy in patients with gynecological cancer. Support. Care Cancer 2017, 25, 3457–3464. [Google Scholar] [CrossRef] [PubMed]
- Mayor, S. Side-effects of cancer drugs are under-reported in trials. Lancet Oncol. 2015, 16, e107. [Google Scholar] [CrossRef]
- Williams, P.A.; Cao, S.; Yang, D.; Jennelle, R.L. Patient-reported outcomes of the relative severity of side effects from cancer radiotherapy. Support. Care Cancer 2020, 28, 309–316. [Google Scholar] [CrossRef]
- Citrin, D.E. Recent developments in radiotherapy. N. Engl. J. Med. 2017, 377, 1065–1075. [Google Scholar] [CrossRef]
- Li, Y.; Li, S.; Meng, X.; Gan, R.Y.; Zhang, J.J.; Li, H.B. Dietary natural products for prevention and treatment of breast cancer. Nutrients 2017, 9, 728. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Li, Y.; Zhou, T.; Zheng, J.; Li, S.; Li, H.B. Dietary natural products for prevention and treatment of liver cancer. Nutrients 2016, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural polyphenols for prevention and treatment of cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Shang, A.; Cao, S.Y.; Xu, X.Y.; Gan, R.Y.; Tang, G.Y.; Corke, H.; Mavumengwana, V.; Li, H.B. Bioactive compounds and biological functions of garlic (Allium sativum L.). Foods 2019, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.Q.; Xu, X.Y.; Cao, S.Y.; Gan, R.Y.; Corke, H.; Beta, T.; Li, H.B. Bioactive compounds and bioactivities of ginger (Zingiber officinale Roscoe). Foods 2019, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Zhou, Y.; Li, Y.; Xu, D.P.; Li, S.; Li, H.B. Spices for prevention and treatment of cancers. Nutrients 2016, 8, 495. [Google Scholar] [CrossRef]
- Zhou, D.D.; Luo, M.; Huang, S.Y.; Saimaiti, A.; Shang, A.; Gan, R.Y.; Li, H.B. Effects and mechanisms of resveratrol on aging and age-related diseases. Oxid. Med. Cell. Longev. 2021, 2021, 9932218. [Google Scholar] [CrossRef]
- Xu, X.Y.; Zhao, C.N.; Cao, S.Y.; Tang, G.Y.; Gan, R.Y.; Li, H.B. Effects and mechanisms of tea for the prevention and management of cancers: An updated review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1693–1705. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Xu, X.Y.; Shang, A.; Gan, R.Y.; Wu, D.T.; Atanasov, A.G.; Li, H.B. Phytochemicals for the prevention and treatment of gastric cancer: Effects and mechanisms. Int. J. Mol. Sci. 2020, 21, 570. [Google Scholar] [CrossRef] [Green Version]
- Gescher, A.J.; Sharma, R.A.; Steward, W.P. Cancer chemoprevention by dietary constituents: A tale of failure and promise. Lancet. Oncol. 2001, 2, 371–379. [Google Scholar] [CrossRef]
- Jiao, Y.; Wilkinson, J.t.; Di, X.; Wang, W.; Hatcher, H.; Kock, N.D.; D’Agostino, R., Jr.; Knovich, M.A.; Torti, F.M.; Torti, S.V. Curcumin, a cancer chemopreventive and chemotherapeutic agent, is a biologically active iron chelator. Blood 2009, 113, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Schraufstatter, E.; Bernt, H. Antibacterial action of curcumin and related compounds. Nature 1949, 164, 456. [Google Scholar] [CrossRef]
- Marquardt, J.U.; Gomez-Quiroz, L.; Arreguin Camacho, L.O.; Pinna, F.; Lee, Y.H.; Kitade, M.; Dominguez, M.P.; Castven, D.; Breuhahn, K.; Conner, E.A.; et al. Curcumin effectively inhibits oncogenic NF-kappaB signaling and restrains stemness features in liver cancer. J. Hepatol. 2015, 63, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Esatbeyoglu, T.; Huebbe, P.; Ernst, I.M.; Chin, D.; Wagner, A.E.; Rimbach, G. Curcumin--from molecule to biological function. Angew. Chem. Int. Ed. Engl. 2012, 51, 5308–5332. [Google Scholar] [CrossRef]
- Egan, M.E.; Pearson, M.; Weiner, S.A.; Rajendran, V.; Rubin, D.; Glockner-Pagel, J.; Canny, S.; Du, K.; Lukacs, G.L.; Caplan, M.J. Curcumin, a major constituent of turmeric, corrects cystic fibrosis defects. Science 2004, 304, 600–602. [Google Scholar] [CrossRef] [Green Version]
- Anggakusuma; Colpitts, C.C.; Schang, L.M.; Rachmawati, H.; Frentzen, A.; Pfaender, S.; Behrendt, P.; Brown, R.J.; Bankwitz, D.; Steinmann, J.; et al. Turmeric curcumin inhibits entry of all hepatitis C virus genotypes into human liver cells. Gut 2014, 63, 1137–1149. [Google Scholar] [CrossRef]
- D’Incalci, M.; Steward, W.P.; Gescher, A.J. Use of cancer chemopreventive phytochemicals as antineoplastic agents. Lancet. Oncol. 2005, 6, 899–904. [Google Scholar] [CrossRef]
- Wang, G.; Duan, P.; Wei, Z.; Liu, F. Curcumin sensitizes carboplatin treatment in triple negative breast cancer through reactive oxygen species induced DNA repair pathway. Mol. Biol. Rep. 2022, 12, 3259–3270. [Google Scholar] [CrossRef]
- Ren, B.; Luo, S.; Tian, X.; Jiang, Z.; Zou, G.; Xu, F.; Yin, T.; Huang, Y.; Liu, J. Curcumin inhibits liver cancer by inhibiting DAMP molecule HSP70 and TLR4 signaling. Oncol. Rep. 2018, 40, 895–901. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Swelum, A.A.; Arif, M.; Abo Ghanima, M.M.; Shukry, M.; Noreldin, A.; Taha, A.E.; El-Tarabily, K.A. Curcumin, the active substance of turmeric: Its effects on health and ways to improve its bioavailability. J. Sci. Food Agric. 2021, 101, 5747–5762. [Google Scholar] [CrossRef]
- Li, M.; Guo, T.; Lin, J.; Huang, X.; Ke, Q.; Wu, Y.; Fang, C.; Hu, C. Curcumin inhibits the invasion and metastasis of triple negative breast cancer via Hedgehog/Gli1 signaling pathway. J. Ethnopharmacol. 2022, 283, 114689. [Google Scholar] [CrossRef]
- Kostrzewa, T.; Wolosewicz, K.; Jamrozik, M.; Drzezdzon, J.; Sieminska, J.; Jacewicz, D.; Gorska-Ponikowska, M.; Kolaczkowski, M.; Lazny, R.; Kuban-Jankowska, A. Curcumin and its new derivatives: Correlation between cytotoxicity against breast cancer cell lines, degradation of PTP1B phosphatase and ROS generation. Int. J. Mol. Sci. 2021, 22, 10368. [Google Scholar] [CrossRef]
- Cai, J.; Sun, H.; Zheng, B.; Xie, M.; Xu, C.; Zhang, G.; Huang, X.; Zhuang, J. Curcumin attenuates lncRNA H19 induced epithelialmesenchymal transition in tamoxifen-resistant breast cancer cells. Mol. Med. Rep. 2021, 23, 660712. [Google Scholar] [CrossRef]
- Li, R.; Zhang, J.; Zhou, Y.; Gao, Q.; Wang, R.; Fu, Y.; Zheng, L.; Yu, H. Transcriptome investigation and in vitro verification of curcumin-induced HO-1 as a feature of ferroptosis in breast cancer cells. Oxid. Med. Cell. Longev. 2020, 2020, 3469840. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Fu, Y.; Zheng, Y.; Ma, M.; Wang, C. Curcumin inhibits proteasome activity in triple-negative breast cancer cells through regulating p300/miR-142-3p/PSMB5 axis. Phytomedicine 2020, 78, 153312. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jiao, D.; Dou, M.; Zhang, W.; Lv, L.; Chen, J.; Li, L.; Wang, L.; Han, X. Curcumin inhibits the growth of triple-negative breast cancer cells by silencing EZH2 and restoring DLC1 expression. Int. J. Mol. Cell. Med. 2020, 24, 10648–10662. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Li, M.; Guo, T.; Wang, S.; Huang, W.; Yang, K.; Liao, Z.; Wang, J.; Zhang, F.; Wang, H. Anti-metastasis activity of curcumin against breast cancer via the inhibition of stem cell-like properties and EMT. Phytomedicine 2019, 58, 152740. [Google Scholar] [CrossRef] [PubMed]
- Al-Yousef, N.; Shinwari, Z.; Al-Shahrani, B.; Al-Showimi, M.; Al-Moghrabi, N. Curcumin induces reexpression of BRCA1 and suppression of gamma synuclein by modulating DNA promoter methylation in breast cancer cell lines. Oncol. Rep. 2020, 43, 827–838. [Google Scholar] [CrossRef] [Green Version]
- Guney Eskiler, G.; Deveci Ozkan, A.; Kaleli, S.; Bilir, C. Inhibition of TLR4/TRIF/IRF3 signaling pathway by curcumin in breast cancer cells. J. Pharm. Pharm. Sci. 2019, 22, 281–291. [Google Scholar] [CrossRef]
- Coker-Gurkan, A.; Celik, M.; Ugur, M.; Arisan, E.D.; Obakan-Yerlikaya, P.; Durdu, Z.B.; Palavan-Unsal, N. Curcumin inhibits autocrine growth hormone-mediated invasion and metastasis by targeting NF-kappaB signaling and polyamine metabolism in breast cancer cells. Amino Acids 2018, 50, 1045–1069. [Google Scholar] [CrossRef]
- Hu, S.; Xu, Y.; Meng, L.; Huang, L.; Sun, H. Curcumin inhibits proliferation and promotes apoptosis of breast cancer cells. Exp. Ther. Med. 2018, 16, 1266–1272. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Zhang, Y.; Wang, Z. Curcumin suppresses the malignancy of non-small cell lung cancer by modulating the circ-PRKCA/miR-384/ITGB1 pathway. Biomed. Pharmacother. 2021, 138, 111439. [Google Scholar] [CrossRef]
- Zhu, X.; Yu, Z.; Feng, L.; Deng, L.; Fang, Z.; Liu, Z.; Li, Y.; Wu, X.; Qin, L.; Guo, R.; et al. Chitosan-based nanoparticle co-delivery of docetaxel and curcumin ameliorates anti-tumor chemoimmunotherapy in lung cancer. Carbohydr. Polym. 2021, 268, 118237. [Google Scholar] [CrossRef]
- Xie, C.; Zhu, J.; Yang, X.; Huang, C.; Zhou, L.; Meng, Z.; Li, X.; Zhong, C. TAp63alpha is involved in tobacco smoke-induced lung cancer EMT and the anti-cancer activity of curcumin via miR-19 transcriptional suppression. Front. Cell Dev. Biol. 2021, 9, 645402. [Google Scholar] [CrossRef]
- Tang, X.; Ding, H.; Liang, M.; Chen, X.; Yan, Y.; Wan, N.; Chen, Q.; Zhang, J.; Cao, J. Curcumin induces ferroptosis in non-small-cell lung cancer via activating autophagy. Thorac. Cancer 2021, 12, 1219–1230. [Google Scholar] [CrossRef]
- Gao, L.; Shao, T.; Zheng, W.; Ding, J. Curcumin suppresses tumor growth of gemcitabine-resistant non-small cell lung cancer by regulating lncRNA-MEG3 and PTEN signaling. Clin. Transl. Oncol. 2021, 23, 1386–1393. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, X.; Tan, J.; Tian, R.; Shen, P.; Cai, W.; Liao, H. Curcumin suppresses the stemness of non-small cell lung cancer cells via promoting the nuclear-cytoplasm translocation of TAZ. Environ. Toxicol. 2021, 36, 1135–1142. [Google Scholar] [CrossRef]
- Mo, F.; Xiao, Y.; Zeng, H.; Fan, D.; Song, J.; Liu, X.; Luo, M.; Ma, X. Curcumin-induced global profiling of transcriptomes in small cell lung cancer cells. Front. Cell Dev. Biol. 2020, 8, 588299. [Google Scholar] [CrossRef]
- Deng, X.; Chen, C.; Wu, F.; Qiu, L.; Ke, Q.; Sun, R.; Duan, Q.; Luo, M.; Luo, Z. Curcumin inhibits the migration and invasion of non-small-cell lung cancer cells through radiation-induced suppression of epithelial-mesenchymal transition and soluble E-cadherin expression. Technol. Cancer Res. Treat. 2020, 19, 1533033820947485. [Google Scholar] [CrossRef]
- Endo, H.; Inoue, I.; Masunaka, K.; Tanaka, M.; Yano, M. Curcumin induces apoptosis in lung cancer cells by 14-3-3 protein-mediated activation of Bad. Biosci. Biotechnol. Biochem. 2020, 84, 2440–2447. [Google Scholar] [CrossRef]
- Liu, F.; Gao, S.; Yang, Y.; Zhao, X.; Fan, Y.; Ma, W.; Yang, D.; Yang, A.; Yu, Y. Antitumor activity of curcumin by modulation of apoptosis and autophagy in human lung cancer A549 cells through inhibiting PI3K/Akt/mTOR pathway. Oncol. Rep. 2018, 39, 1523–1531. [Google Scholar] [CrossRef]
- Zhan, J.W.; Jiao, D.M.; Wang, Y.; Song, J.; Wu, J.H.; Wu, L.J.; Chen, Q.Y.; Ma, S.L. Integrated microRNA and gene expression profiling reveals the crucial miRNAs in curcumin anti-lung cancer cell invasion. Thorac. Cancer 2017, 8, 461–470. [Google Scholar] [CrossRef]
- Elbadawy, M.; Hayashi, K.; Ayame, H.; Ishihara, Y.; Abugomaa, A.; Shibutani, M.; Hayashi, S.M.; Hazama, S.; Takenouchi, H.; Nakajima, M.; et al. Anti-cancer activity of amorphous curcumin preparation in patient-derived colorectal cancer organoids. Biomed. Pharmacother. 2021, 142, 112043. [Google Scholar] [CrossRef]
- Herrero de la Parte, B.; Rodeno-Casado, M.; Iturrizaga Correcher, S.; Mar Medina, C.; Garcia-Alonso, I. Curcumin reduces colorectal cancer cell proliferation and migration and slows in vivo growth of liver metastases in rats. Biomedicines 2021, 9, 1183. [Google Scholar] [CrossRef]
- Mao, X.; Zhang, X.; Zheng, X.; Chen, Y.; Xuan, Z.; Huang, P. Curcumin suppresses LGR5(+) colorectal cancer stem cells by inducing autophagy and via repressing TFAP2A-mediated ECM pathway. J. Nat. Med. 2021, 75, 590–601. [Google Scholar] [CrossRef]
- Li, G.; Fang, S.; Shao, X.; Li, Y.; Tong, Q.; Kong, B.; Chen, L.; Wang, Y.; Yang, J.; Yu, H.; et al. Curcumin reverses NNMT-induced 5-fluorouracil resistance via increasing ROS and cell cycle arrest in colorectal cancer cells. Biomolecules 2021, 11, 1295. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Yin, H.; Ma, H.; Wang, Y.; Kong, D.; Fan, Z. Curcumin regulates ERCC1 expression and enhances oxaliplatin sensitivity in resistant colorectal cancer cells through its effects on miR-409-3p. Evid. Based Complement. Altern. Med. 2020, 2020, 8394574. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, R.; Zhang, X.; Zhang, B.; Yao, Q. Curcumin may reverse 5-fluorouracil resistance on colonic cancer cells by regulating TET1-NKD-Wnt signal pathway to inhibit the EMT progress. Biomed. Pharmacother. 2020, 129, 110381. [Google Scholar] [CrossRef]
- Chen, T.; Yang, C.; Xi, Z.; Chen, F.; Li, H. Reduced caudal type homeobox 2 (CDX2) promoter methylation is associated with curcumin’s suppressive effects on epithelial-mesenchymal transition in colorectal cancer cells. Med. Sci. Monit. 2020, 26, e926443. [Google Scholar] [CrossRef]
- Zheng, Z.H.; You, H.Y.; Feng, Y.J.; Zhang, Z.T. LncRNA KCNQ1OT1 is a key factor in the reversal effect of curcumin on cisplatin resistance in the colorectal cancer cells. Mol. Cell. Biochem. 2021, 476, 2575–2585. [Google Scholar] [CrossRef]
- Yu, H.; Xie, Y.; Zhou, Z.; Wu, Z.; Dai, X.; Xu, B. Curcumin regulates the progression of colorectal cancer via lncRNA NBR2/AMPK pathway. Technol. Cancer Res. Treat. 2019, 18, 1533033819870781. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Zand, H.; Cheraghpour, M. The Influence of curcumin on the downregulation of MYC, insulin and IGF-1 receptors: A possible mechanism underlying the anti-growth and anti-migration in chemoresistant colorectal cancer cells. Medicina 2019, 55, 90. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Wang, L.; Wang, Y.; Shen, H.; Wang, X.; Wu, L. Curcumin reverses oxaliplatin resistance in human colorectal cancer via regulation of TGF-beta/Smad2/3 signaling pathway. Onco Targets Ther. 2019, 12, 3893–3903. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.H.; Huang, C.Y.; Chou, C.W.; Makondi, P.T.; Huang, M.T.; Wei, P.L.; Chang, Y.J. Heat shock protein 27 influences the anti-cancer effect of curcumin in colon cancer cells through ROS production and autophagy activation. Life Sci. 2018, 209, 43–51. [Google Scholar] [CrossRef]
- Dou, H.; Shen, R.; Tao, J.; Huang, L.; Shi, H.; Chen, H.; Wang, Y.; Wang, T. Curcumin suppresses the colon cancer proliferation by inhibiting Wnt/beta-catenin pathways via miR-130a. Front. Pharmacol. 2017, 8, 877. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Zhu, M.; Qiu, F.; Li, W.; Wang, M.; Guo, Y.; Xi, X.; Du, B. Curcumin may be a potential adjuvant treatment drug for colon cancer by targeting CD44. Int. Immunopharmacol. 2020, 88, 106991. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Lim, M.A.; Jung, S.N.; Oh, C.; Won, H.R.; Jin, Y.L.; Piao, Y.; Kim, H.J.; Chang, J.W.; Koo, B.S. The effect of curcumin on multi-level immune checkpoint blockade and T cell dysfunction in head and neck cancer. Phytomedicine 2021, 92, 153758. [Google Scholar] [CrossRef]
- Borges, G.A.; Elias, S.T.; Amorim, B.; de Lima, C.L.; Coletta, R.D.; Castilho, R.M.; Squarize, C.H.; Guerra, E.N.S. Curcumin downregulates the PI3K-AKT-mTOR pathway and inhibits growth and progression in head and neck cancer cells. Phytother. Res. 2020, 34, 3311–3324. [Google Scholar] [CrossRef]
- Emami, A.; Babaei, E.; Nagishbandi, A.; Azeez, H.J.; Feizi, M.A.H.; Golizadeh, A. Cellular uptake and apoptotic properties of gemini curcumin in gastric cancer cells. Mol. Biol. Rep. 2021, 48, 7215–7222. [Google Scholar] [CrossRef]
- Tong, R.; Wu, X.; Liu, Y.; Liu, Y.; Zhou, J.; Jiang, X.; Zhang, L.; He, X.; Ma, L. Curcumin-induced DNA demethylation in human gastric cancer cells is mediated by the DNA-damage response pathway. Oxid. Med. Cell. Longev. 2020, 2020, 2543504. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, C.; Ren, Z.; Zhang, F.; Xu, J.; Zhang, X.; Zheng, H. Curcumin affects gastric cancer cell migration, invasion and cytoskeletal remodeling through Gli1-beta-Catenin. Cancer Manag. Res. 2020, 12, 3795–3806. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, S.; Liu, C.; Liu, X. Curcumin promoted miR-34a expression and suppressed proliferation of gastric cancer cells. Cancer Biother. Radiopharm. 2019, 34, 634–641. [Google Scholar] [CrossRef]
- Fu, H.; Wang, C.; Yang, D.; Wei, Z.; Xu, J.; Hu, Z.; Zhang, Y.; Wang, W.; Yan, R.; Cai, Q. Curcumin regulates proliferation, autophagy, and apoptosis in gastric cancer cells by affecting PI3K and P53 signaling. J. Cell. Physiol. 2018, 233, 4634–4642. [Google Scholar] [CrossRef]
- Zhou, S.; Yao, D.; Guo, L.; Teng, L. Curcumin suppresses gastric cancer by inhibiting gastrin-mediated acid secretion. FEBS Open Bio 2017, 7, 1078–1084. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yang, G.; Zhang, R.; Dong, L.; Chen, H.; Bo, J.; Xue, W.; Huang, Y. Curcumin inhibits cell proliferation and motility via suppression of TROP2 in bladder cancer cells. Int. J. Oncol. 2018, 53, 515–526. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Tan, S.L.; Lu, Q.; Xu, R.; Cao, J.; Wu, S.Q.; Wang, Y.H.; Zhao, X.K.; Zhong, Z.H. Curcumin suppresses microRNA-7641-mediated regulation of p16 expression in bladder cancer. Am. J. Chin. Med. 2018, 46, 1357–1368. [Google Scholar] [CrossRef]
- Pan, L.; Sha, J.; Lin, W.; Wang, Y.; Bian, T.; Guo, J. Curcumin inhibits prostate cancer progression by regulating the miR-30a-5p/PCLAF axis. Exp. Ther. Med. 2021, 22, 969. [Google Scholar] [CrossRef]
- Zeng, Y.; Du, Q.; Zhang, Z.; Ma, J.; Han, L.; Wang, Y.; Yang, L.; Tao, N.; Qin, Z. Curcumin promotes cancer-associated fibroblasts apoptosis via ROS-mediated endoplasmic reticulum stress. Arch. Biochem. Biophys. 2020, 694, 108613. [Google Scholar] [CrossRef]
- Ide, H.; Lu, Y.; Noguchi, T.; Muto, S.; Okada, H.; Kawato, S.; Horie, S. Modulation of AKR1C2 by curcumin decreases testosterone production in prostate cancer. Cancer Sci. 2018, 109, 1230–1238. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Zheng, Z.; Huang, J.; Ma, X.; Huang, C.; Wu, R.; Li, X.; Liang, Z.; Deng, F.; Wu, J.; et al. Modulation of miR-34a in curcumin-induced antiproliferation of prostate cancer cells. J. Cell. Biochem. 2019, 120, 15616–15624. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, S.; Cheng, X.; Wu, J.; Wu, L.; Wang, Y.; Wang, X.; Bao, J.; Yu, H. Curcumin induces autophagic cell death in human thyroid cancer cells. Toxicol. In Vitro 2022, 78, 105254. [Google Scholar] [CrossRef]
- Liang, Y.; Kong, D.; Zhang, Y.; Li, S.; Li, Y.; Dong, L.; Zhang, N.; Ma, J. Curcumin inhibits the viability, migration and invasion of papillary thyroid cancer cells by regulating the miR-301a-3p/STAT3 axis. Exp. Ther. Med. 2021, 22, 875. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Liao, L.; Zhou, Q.; Huang, X.; Zheng, P.; Guo, Y.; Deng, T.; Tian, X. Curcumin inhibits the growth of liver cancer by impairing myeloid-derived suppressor cells in murine tumor tissues. Oncol. Lett. 2021, 21, 286. [Google Scholar] [CrossRef] [PubMed]
- Soni, V.K.; Shukla, D.; Kumar, A.; Vishvakarma, N.K. Curcumin circumvent lactate-induced chemoresistance in hepatic cancer cells through modulation of hydroxycarboxylic acid receptor-1. Int. J. Biochem. Cell Biol. 2020, 123, 105752. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Kong, L.; Han, Y.; Zhang, S. Gut microbiota enhances the chemosensitivity of hepatocellular carcinoma to 5-fluorouracil in vivo by increasing curcumin bioavailability. Phytother. Res. 2021, 35, 5823–5837. [Google Scholar] [CrossRef]
- Wu, R.; Mei, X.; Ye, Y.; Xue, T.; Wang, J.; Sun, W.; Lin, C.; Xue, R.; Zhang, J.; Xu, D. Zn(II)-curcumin solid dispersion impairs hepatocellular carcinoma growth and enhances chemotherapy by modulating gut microbiota-mediated zinc homeostasis. Pharmacol. Res. 2019, 150, 104454. [Google Scholar] [CrossRef]
- Kim, M.J.; Park, K.S.; Kim, K.T.; Gil, E.Y. The inhibitory effect of curcumin via fascin suppression through JAK/STAT3 pathway on metastasis and recurrence of ovary cancer cells. BMC Womens Health 2020, 20, 256. [Google Scholar] [CrossRef]
- Yen, H.Y.; Tsao, C.W.; Lin, Y.W.; Kuo, C.C.; Tsao, C.H.; Liu, C.Y. Regulation of carcinogenesis and modulation through Wnt/beta-catenin signaling by curcumin in an ovarian cancer cell line. Sci. Rep. 2019, 9, 17267. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.D.; Pang, Y.X.; Zhao, X.R.; Li, R.; Jin, C.J.; Xue, J.; Dong, R.Y.; Liu, P.S. Curcumin induces apoptotic cell death and protective autophagy by inhibiting AKT/mTOR/p70S6K pathway in human ovarian cancer cells. Arch. Gynecol. Obstet. 2019, 299, 1627–1639. [Google Scholar] [CrossRef]
- Sun, S.; Fang, H. Curcumin inhibits ovarian cancer progression by regulating circ-PLEKHM3/miR-320a/SMG1 axis. J. Ovarian Res. 2021, 14, 158. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Sakamoto, T.; Zhengguang, L.; Yasui, H.; Hamada, H.; Kubo, H.; Nakajima, M. Curcumin inhibits epithelial-mesenchymal transition in oral cancer cells via c-Met blockade. Oncol. Lett. 2020, 19, 4177–4182. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, K.; Dobiasch, S.; Nguyen, L.; Schilling, D.; Combs, S.E. Modification of radiosensitivity by curcumin in human pancreatic cancer cell lines. Sci. Rep. 2020, 10, 3815. [Google Scholar] [CrossRef]
- Yang, D.; Li, Y.; Zhao, D. Curcumin induces apoptotic cell death in human pancreatic cancer cells via the miR-340/XIAP signaling pathway. Oncol. Lett. 2017, 14, 1811–1816. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Zhou, X.; Yin, X.; Wang, L.; Zhao, Z.; Hou, Y.; Zheng, N.; Xia, J.; Wang, Z. The effects of curcumin on proliferation, apoptosis, invasion, and NEDD4 expression in pancreatic cancer. Biochem. Pharmacol. 2017, 140, 28–40. [Google Scholar] [CrossRef]
- Wang, T.; Wu, X.; Al Rudaisat, M.; Song, Y.; Cheng, H. Curcumin induces G2/M arrest and triggers autophagy, ROS generation and cell senescence in cervical cancer cells. J. Cancer 2020, 11, 6704–6715. [Google Scholar] [CrossRef] [PubMed]
- Aedo-Aguilera, V.; Carrillo-Beltran, D.; Calaf, G.M.; Munoz, J.P.; Guerrero, N.; Osorio, J.C.; Tapia, J.C.; Leon, O.; Contreras, H.R.; Aguayo, F. Curcumin decreases epithelialmesenchymal transition by a Pirindependent mechanism in cervical cancer cells. Oncol. Rep. 2019, 42, 2139–2148. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Zhuang, Z.; Su, Q.; He, J.; Li, H. Curcumin has anti-proliferative and pro-apoptotic effects on tongue cancer in vitro: A study with bioinformatics analysis and in vitro experiments. Drug Des. Devel. Ther. 2020, 14, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Deng, J.; Yuan, J.; Tang, X.; Wang, Y.; Chen, H.; Liu, Y.; Zhou, L. Curcumin exerts its tumor suppressive function via inhibition of NEDD4 oncoprotein in glioma cancer cells. Int. J. Oncol. 2017, 51, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Golias, C.H.; Charalabopoulos, A.; Charalabopoulos, K. Cell proliferation and cell cycle control: A mini review. Int. J. Clin. Pract. 2004, 58, 1134–1141. [Google Scholar] [CrossRef]
- Roy, D.; Sheng, G.Y.; Herve, S.; Carvalho, E.; Mahanty, A.; Yuan, S.; Sun, L. Interplay between cancer cell cycle and metabolism: Challenges, targets and therapeutic opportunities. Biomed. Pharmacother. 2017, 89, 288–296. [Google Scholar] [CrossRef]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef]
- Suski, J.M.; Braun, M.; Strmiska, V.; Sicinski, P. Targeting cell-cycle machinery in cancer. Cancer Cell 2021, 39, 759–778. [Google Scholar] [CrossRef]
- Otto, T.; Sicinski, P. Cell cycle proteins as promising targets in cancer therapy. Nat. Rev. Cancer 2017, 17, 93–115. [Google Scholar] [CrossRef] [Green Version]
- Ingham, M.; Schwartz, G.K. Cell-cycle therapeutics come of age. J. Clin. Oncol. 2017, 35, 2949–2959. [Google Scholar] [CrossRef]
- Leal-Esteban, L.C.; Fajas, L. Cell cycle regulators in cancer cell metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165715. [Google Scholar] [CrossRef]
- Friedl, P.; Alexander, S. Cancer invasion and the microenvironment: Plasticity and reciprocity. Cell 2011, 147, 992–1009. [Google Scholar] [CrossRef] [Green Version]
- Friedl, P.; Wolf, K. Tumour-cell invasion and migration: Diversity and escape mechanisms. Nat. Rev. Cancer 2003, 3, 362–374. [Google Scholar] [CrossRef]
- Thiery, J.P. Epithelial-mesenchymal transitions in tumour progression. Nat. Rev. Cancer 2002, 2, 442–454. [Google Scholar] [CrossRef]
- Mittal, V. Epithelial mesenchymal transition in tumor metastasis. Annu. Rev. Pathol. 2018, 13, 395–412. [Google Scholar] [CrossRef]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhou, D.D.; Huang, S.Y.; Fang, A.P.; Li, H.B.; Zhu, H.L. Effects and mechanisms of natural products on Alzheimer’s disease. Crit. Rev. Food Sci. Nutr. 2021, 1–21. [Google Scholar] [CrossRef]
- Mohammad, R.M.; Muqbil, I.; Lowe, L.; Yedjou, C.; Hsu, H.Y.; Lin, L.T.; Siegelin, M.D.; Fimognari, C.; Kumar, N.B.; Dou, Q.P.; et al. Broad targeting of resistance to apoptosis in cancer. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2015; Volume 35, pp. S78–S103. [Google Scholar] [CrossRef]
- Mortezaee, K.; Salehi, E.; Mirtavoos-Mahyari, H.; Motevaseli, E.; Najafi, M.; Farhood, B.; Rosengren, R.J.; Sahebkar, A. Mechanisms of apoptosis modulation by curcumin: Implications for cancer therapy. J. Cell. Physiol. 2019, 234, 12537–12550. [Google Scholar] [CrossRef]
- Qiao, L.; Wong, B.C. Targeting apoptosis as an approach for gastrointestinal cancer therapy. Drug Resist. Updates 2009, 12, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Onorati, A.V.; Dyczynski, M.; Ojha, R.; Amaravadi, R.K. Targeting autophagy in cancer. Cancer 2018, 124, 3307–3318. [Google Scholar] [CrossRef] [Green Version]
- Ishaq, M.; Ojha, R.; Sharma, A.P.; Singh, S.K. Autophagy in cancer: Recent advances and future directions. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2020; Volume 66, pp. 171–181. [Google Scholar] [CrossRef]
- Xia, H.; Green, D.R.; Zou, W. Autophagy in tumour immunity and therapy. Nat. Rev. Cancer 2021, 21, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Srivastava, S.K. Cancer cells stemness: A doorstep to targeted therapy. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165424. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Hsu, W.H.; Han, J.; Xia, Y.; DePinho, R.A. Cancer stemness meets immunity: From mechanism to therapy. Cell Rep. 2021, 34, 108597. [Google Scholar] [CrossRef]
- Tsui, Y.M.; Chan, L.K.; Ng, I.O. Cancer stemness in hepatocellular carcinoma: Mechanisms and translational potential. Br. J. Cancer 2020, 122, 1428–1440. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative stress in cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef]
- Pelicano, H.; Carney, D.; Huang, P. ROS stress in cancer cells and therapeutic implications. Drug Resist. Updates 2004, 7, 97–110. [Google Scholar] [CrossRef]
- Cheung, E.C.; Vousden, K.H. The role of ROS in tumour development and progression. Nat. Rev. Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Li, S.; Gan, R.Y.; Zhou, T.; Xu, D.P.; Li, H.B. Impacts of gut bacteria on human health and diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519. [Google Scholar] [CrossRef]
- Helmink, B.A.; Khan, M.A.W.; Hermann, A.; Gopalakrishnan, V.; Wargo, J.A. The microbiome, cancer, and cancer therapy. Nat. Med. 2019, 25, 377–388. [Google Scholar] [CrossRef]
- Sepich-Poore, G.D.; Zitvogel, L.; Straussman, R.; Hasty, J.; Wargo, J.A.; Knight, R. The microbiome and human cancer. Science 2021, 371, eabc4552. [Google Scholar] [CrossRef]
- Park, E.M.; Chelvanambi, M.; Bhutiani, N.; Kroemer, G.; Zitvogel, L.; Wargo, J.A. Targeting the gut and tumor microbiota in cancer. Nat. Med. 2022, 28, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Li, S.; Gan, R.Y.; Zhao, C.N.; Meng, X.; Li, H.B. Targeting gut microbiota with dietary components on cancer: Effects and potential mechanisms of action. Crit. Rev. Food Sci. Nutr. 2020, 60, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- McFadden, R.-M.T.; Larmonier, C.B.; Shehab, K.W.; Midura-Kiela, M.; Ramalingam, R.; Harrison, C.A.; Besselsen, D.G.; Chase, J.H.; Caporaso, J.G.; Jobin, C.; et al. The Role of curcumin in modulating colonic microbiota during colitis and colon cancer prevention. Inflamm. Bowel Dis. 2015, 21, 2483–2494. [Google Scholar] [CrossRef] [PubMed]
- Szakacs, G.; Paterson, J.K.; Ludwig, J.A.; Booth-Genthe, C.; Gottesman, M.M. Targeting multidrug resistance in cancer. Nat. Rev. Drug Discov. 2006, 5, 219–234. [Google Scholar] [CrossRef]
- Lim, Z.F.; Ma, P.C. Emerging insights of tumor heterogeneity and drug resistance mechanisms in lung cancer targeted therapy. J. Hematol. Oncol. 2019, 12, 134. [Google Scholar] [CrossRef] [Green Version]
- Zarei, E.; Sefidi-Heris, Y.; Saadat, I. Synergistic effects of metformin and curcumin on cytotoxicity of chemotherapy drugs using a gastric cancer cell line model. EXCLI J. 2021, 20, 1488–1498. [Google Scholar] [CrossRef]
- DiMarco-Crook, C.; Rakariyatham, K.; Li, Z.; Du, Z.; Zheng, J.; Wu, X.; Xiao, H. Synergistic anticancer effects of curcumin and 3′,4′-didemethylnobiletin in combination on colon cancer cells. J. Food Sci. 2020, 85, 1292–1301. [Google Scholar] [CrossRef]
- Xie, L.; Ji, X.; Zhang, Q.; Wei, Y. Curcumin combined with photodynamic therapy, promising therapies for the treatment of cancer. Biomed. Pharmacother. 2022, 146, 112567. [Google Scholar] [CrossRef]
- Hidayat, Y.M.; Wagey, F.; Suardi, D.; Susanto, H.; Laihad, B.J.; Tobing, M.D.L. Analysis of curcumin as a radiosensitizer in cancer therapy with serum survivin examination: Randomised control trial. Asian Pac. J. Cancer Prev. 2021, 22, 139–143. [Google Scholar] [CrossRef]
- Saghatelyan, T.; Tananyan, A.; Janoyan, N.; Tadevosyan, A.; Petrosyan, H.; Hovhannisyan, A.; Hayrapetyan, L.; Arustamyan, M.; Arnhold, J.; Rotmann, A.R.; et al. Efficacy and safety of curcumin in combination with paclitaxel in patients with advanced, metastatic breast cancer: A comparative, randomized, double-blind, placebo-controlled clinical trial. Phytomedicine 2020, 70, 153218. [Google Scholar] [CrossRef]
- Choi, Y.H.; Han, D.H.; Kim, S.W.; Kim, M.J.; Sung, H.H.; Jeon, H.G.; Jeong, B.C.; Seo, S.I.; Jeon, S.S.; Lee, H.M.; et al. A randomized, double-blind, placebo-controlled trial to evaluate the role of curcumin in prostate cancer patients with intermittent androgen deprivation. Prostate 2019, 79, 614–621. [Google Scholar] [CrossRef]
- Saadipoor, A.; Razzaghdoust, A.; Simforoosh, N.; Mahdavi, A.; Bakhshandeh, M.; Moghadam, M.; Abdollahi, H.; Mofid, B. Randomized, double-blind, placebo-controlled phase II trial of nanocurcumin in prostate cancer patients undergoing radiotherapy. Phytother. Res. 2019, 33, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Passildas-Jahanmohan, J.; Eymard, J.C.; Pouget, M.; Kwiatkowski, F.; Van Praagh, I.; Savareux, L.; Atger, M.; Durando, X.; Abrial, C.; Richard, D.; et al. Multicenter randomized phase II study comparing docetaxel plus curcumin versus docetaxel plus placebo in first-line treatment of metastatic castration-resistant prostate cancer. Cancer Med. 2021, 10, 2332–2340. [Google Scholar] [CrossRef]
- Howells, L.M.; Iwuji, C.O.O.; Irving, G.R.B.; Barber, S.; Walter, H.; Sidat, Z.; Griffin-Teall, N.; Singh, R.; Foreman, N.; Patel, S.R.; et al. Curcumin combined with FOLFOX chemotherapy is safe and tolerable in patients with metastatic colorectal cancer in a randomized phase IIa trial. J. Nutr. 2019, 149, 1133–1139. [Google Scholar] [CrossRef] [Green Version]
- Panahi, Y.; Saberi-Karimian, M.; Valizadeh, O.; Behnam, B.; Saadat, A.; Jamialahmadi, T.; Majeed, M.; Sahebkar, A. Effects of curcuminoids on systemic inflammation and quality of life in patients with colorectal cancer undergoing chemotherapy: A randomized controlled trial. In Natural Products and Human Diseases; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2021; Volume 1328, pp. 1–9. [Google Scholar] [CrossRef]
- Pastorelli, D.; Fabricio, A.S.C.; Giovanis, P.; D’Ippolito, S.; Fiduccia, P.; Solda, C.; Buda, A.; Sperti, C.; Bardini, R.; Da Dalt, G.; et al. Phytosome complex of curcumin as complementary therapy of advanced pancreatic cancer improves safety and efficacy of gemcitabine: Results of a prospective phase II trial. Pharmacol. Res. 2018, 132, 72–79. [Google Scholar] [CrossRef]
- Tuyaerts, S.; Rombauts, K.; Everaert, T.; Van Nuffel, A.M.T.; Amant, F. A Phase 2 study to assess the immunomodulatory capacity of a lecithin-based delivery system of curcumin in endometrial cancer. Front. Nutr. 2018, 5, 138. [Google Scholar] [CrossRef] [Green Version]
- Basak, S.K.; Bera, A.; Yoon, A.J.; Morselli, M.; Jeong, C.; Tosevska, A.; Dong, T.S.; Eklund, M.; Russ, E.; Nasser, H.; et al. A randomized, phase 1, placebo-controlled trial of APG-157 in oral cancer demonstrates systemic absorption and an inhibitory effect on cytokines and tumor-associated microbes. Cancer 2020, 126, 1668–1682. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, N.; He, H.; Tang, X. Pharmaceutical strategies of improving oral systemic bioavailability of curcumin for clinical application. J. Control. Release 2019, 316, 359–380. [Google Scholar] [CrossRef]
- Sanidad, K.Z.; Sukamtoh, E.; Xiao, H.; McClements, D.J.; Zhang, G. Curcumin: Recent advances in the development of strategies to improve oral bioavailability. Annu. Rev. Food Sci. Technol. 2019, 10, 597–617. [Google Scholar] [CrossRef] [Green Version]
- Elbialy, N.S.; Aboushoushah, S.F.; Mohamed, N. Bioinspired synthesis of protein/polysaccharide-decorated folate as a nanocarrier of curcumin to potentiate cancer therapy. Int. J. Pharm. 2022, 613, 121420. [Google Scholar] [CrossRef]
- Freidus, L.G.; Kumar, P.; Marimuthu, T.; Pradeep, P.; Choonara, Y.E. Theranostic mesoporous silica nanoparticles loaded with a curcumin-naphthoquinone conjugate for potential cancer intervention. Front. Mol. Biosci. 2021, 8, 670792. [Google Scholar] [CrossRef]
- Karimpour, M.; Feizi, M.A.H.; Mahdavi, M.; Krammer, B.; Verwanger, T.; Najafi, F.; Babaei, E. Development of curcumin-loaded gemini surfactant nanoparticles: Synthesis, characterization and evaluation of anticancer activity against human breast cancer cell lines. Phytomedicine 2019, 57, 183–190. [Google Scholar] [CrossRef]
- Lai, H.; Ding, X.; Ye, J.; Deng, J.; Cui, S. pH-responsive hyaluronic acid-based nanoparticles for targeted curcumin delivery and enhanced cancer therapy. Colloids Surf. B Biointerfaces 2021, 198, 111455. [Google Scholar] [CrossRef]
- Ghaffari, S.B.; Sarrafzadeh, M.H.; Fakhroueian, Z.; Khorramizadeh, M.R. Flower-like curcumin-loaded folic acid-conjugated ZnO-MPA- betacyclodextrin nanostructures enhanced anticancer activity and cellular uptake of curcumin in breast cancer cells. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 103, 109827. [Google Scholar] [CrossRef]
- Ndong Ntoutoume, G.M.A.; Granet, R.; Mbakidi, J.P.; Brégier, F.; Léger, D.Y.; Fidanzi-Dugas, C.; Lequart, V.; Joly, N.; Liagre, B.; Chaleix, V.; et al. Development of curcumin-cyclodextrin/cellulose nanocrystals complexes: New anticancer drug delivery systems. Bioorg. Med. Chem. Lett. 2016, 26, 941–945. [Google Scholar] [CrossRef]
- Fernandez-Lazaro, D.; Mielgo-Ayuso, J.; Seco Calvo, J.; Cordova Martinez, A.; Caballero Garcia, A.; Fernandez-Lazaro, C.I. Modulation of exercise-induced muscle damage, inflammation, and oxidative markers by curcumin supplementation in a physically active population: A systematic review. Nutrients 2020, 12, 501. [Google Scholar] [CrossRef] [Green Version]
- Mehta, H.J.; Patel, V.; Sadikot, R.T. Curcumin and lung cancer-A review. Target. Oncol. 2014, 9, 295–310. [Google Scholar] [CrossRef]
- Soleimani, V.; Sahebkar, A.; Hosseinzadeh, H. Turmeric (Curcuma longa) and its major constituent (curcumin) as nontoxic and safe substances: Review. Phytother. Res. PTR 2018, 32, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.A.; Euden, S.A.; Platton, S.L.; Cooke, D.N.; Shafayat, A.; Hewitt, H.R.; Marczylo, T.H.; Morgan, B.; Hemingway, D.; Plummer, S.M.; et al. Phase I clinical trial of oral curcumin: Biomarkers of systemic activity and compliance. Clin. Cancer Res. 2004, 10, 6847–6854. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Study Type | Models | Dose & Duration | Effects | Mechanisms | Ref. |
|---|---|---|---|---|---|
| Breast cancer | |||||
| In vitro In vivo | MDA-MB-231 and MDA-MB-468 cells; female BALB/c-nu/nu mice with MDA-MB-231 adherent cells | 10, 15, 20, 25, 30 and 35 µM, 24, 48 and 72 h | Inhibiting proliferation, invasion and migration, EMT and stemness | ↓PTCH1, SMO, Gli1, Gli2, N-cadherin, vimentin, Oct4, Sox2 | [27] |
| In vitro | MCF-7 and MDA-MB-231 cells | 6.25, 25 and 100 µM, 24 h | Cytotoxicity and photosensitizing effect | ↓PTP1B; ↑ROS | [28] |
| In vitro | MCF-7/TAMR cells | 5, 10, 20, 30 and 40 µM, 48 h | Preventing cell migration and invasion, and EMT | ↓N-cadherin, H19; ↑E-cadherin | [29] |
| In vitro | MCF-7 and MDA-MB-231 cells | 5, 10, 20, 40, 60, 80, 100, 120 and140 μM, 24 and 48 h | Inhibiting cell viability; Promoting oxidative stress, ER stress, and ferroptosis | ↑HO-1, Nrf2, ROS, HSPA5, ATF4, DDIT3, MDA, FTL, TFRC, FTH1, BACH1, RELA, USF1, NFE2L2; ↓GPX4, GSH | [30] |
| In vitro In vivo | MDA-MB-231 cell; BALB/ c nude mice with MDA-MB-231 cells | 5, 10, 20 and 50 μM, 24 h; 25 g/kg, 4 weeks | Inhibiting cell proliferation and cancer growth | ↑GFPu, miR-142-3p; ↓PSMB5, PSMB1, P300, CT-1 | [31] |
| In vitro, In vivo | MCF-7, MDA-MB-231 and MDA-MB-468 cells; female BALB/c nude mice with MDA-MB-231 cells | 20 and 40 µM, 48 h; 100 mg/kg/2 days, 21 days | Inhibiting proliferation, migration and invasion; Promoting apoptosis; Blocking the cell cycle | ↓cyclin A1, CDK1, Bcl-2, EZH2; ↑Caspase-9, DLC1 | [32] |
| In vitro | MCF-7 and MDA-MB-231 cells | 10, 15, 20, 25, 30, 35 and 40 µM, 24 and 48 h | Inhibiting cell viability, invasion and migration, mammosphere formation and differentiation abilities, stem cell properties | ↓CD44+CD24− subpopulation, vimentin, fibronectin, β-catenin, Oct4, Nanog, Sox2; ↑E-cadherin | [33] |
| In vitro | HCC-38, UACC-3199, and T47D cells | 5 and 10 µM, 3 days | Suppressing proliferation and methylation | ↓DNMT1, miR-29b, SNCG; ↑BRCA1, TET1, DNMT3 | [34] |
| In vitro | MCF-7 and MDA-MB-231 cells | 5, 10 and 25 µM, 48 h | Inhibiting cell vitality; Inducing apoptosis | ↓TLR4, TRIF, IRF3, IFN-α/β | [35] |
| In vitro | MCF-7, MDA-MB-453 and MDA-MB-231 cells | 5, 10, 15, 20, 25 and 30 µM, 24, 48 and 72 h | Inhibiting proliferation, invasion and metastasis; Inducing apoptotic cell death and cell cycle arrest | ↓Src, pSTAT-1, p-Akt, p-p44/42, Ras, c-raf, vimentin, β-catenin, p53, Rb, p-Rb, Bax, Bcl-2, Bcl-xL, Mcl-1; ↑PIAS-3, SOCS-1, SOCS-3, ROS, NF-κB, PAO, SSAT, p21, Bak | [36] |
| In vitro | T47D, MCF7, MDA-MB-415, SK-BR-3, MDA-MB-231, MDA-MB-468 and BT-20 cells | 10 and 30 µM, 24 and 48 h | Inhibiting proliferation; Inducing G2/M arrest and apoptosis | ↓CDC25, CDC2, p-Akt, p-mTOR, p-S6, Bcl-2; ↑p21, Bax, Cleaved-caspase-3 | [37] |
| In vitro | MDA-MB-231 and CAL-51 cells | 5 µM, 48 h | Inhibiting proliferation; Inducing apoptosis | ↓Bcl-2, RAD51; ↑ROS, Bax, γH2AX | [24] |
| Lung cancer | |||||
| In vitro In vivo | H1650, H1299, H460 and A549 cells; BALB/c nude mice with A549 cells | 10, 20 and 40 μM, 24 h; 50 mg/kg, 22 days | Accelerating apoptosis; Inhibiting migration, invasion and xenograft tumor growth | ↓circ-PRKCA, ITGB1; ↑miR-384 | [38] |
| In vitro In vivo | H460, H1299, H1975, A549, SCC-827, PC-9 and CMT-64 cells; female C57bl/6j mice with CMT-64 cells | 4, 8, 12, 16, 20, 24 and 28 μg/mL, 24 h; 5 mg/kg, 24 h | Inhibiting of tumor growth and volume; Ameliorating the immunosuppressive micro-environment | ↓MDSCs cells, Treg cells, IL-10; ↑NK cells | [39] |
| In vitro | H1299 and A549 cells | 2.5, 5 and 7.5 μM, 48 h | Decreasing migration, invasion and EMT Process | ↑TAp63α, E-cadherin, ZO-1; ↓Vimentin, N-cadherin, miR-19a, miR-19b | [40] |
| In vitro In vivo | A549 and H1299 cells; female C57BL/6 mice with Lewis lung carcinomas cells | 5, 10, 20, 30 and 40 μM, 24 h; 100 mg/kg/day, 15days | Inhibiting tumor growth; Inducing ferroptosis and autophagy | ↓SOD, GSH, SLC7A11, GPX4, p62; ↑MDA, iron, ACSL4, Beclin1, LC3-II, autolysosome, mitochondrial damage | [41] |
| In vitro In vivo | A549/GR and H520/GR cells; BALB/c nude mice with A549/GR cells | 50, 100 and 150 μM, 48 h; 100 mg/kg, 3 weeks | Suppressing proliferation; Promoting apoptosis | ↑lncRNA-MEG3, PTEN | [42] |
| In vitro | A549, NCI-H1299 | 5, 25, 125 and 250 nM, 24, 48 and 72 h | Suppressing sphere size and number, and stemness | ↓ALDH, CD133, Epcam, Oct4, TAZ; ↑Hippo pathway, p-TAZ | [43] |
| In vitro | H446 cells | 5, 10, 15 and 20 μM, 24 and 48 h | Inducing cell apoptosis; Regulating cell cycle | ↓Bcl-2, CCNF, LOX1, MRGPRF, and VEGFB; ↑Bax, cytochrome-C, miR-548ah-5p | [44] |
| In vitro | A549 cells | 1, 2, 5, 10 and 20 μM, 24 and 48 h | Inhibiting migration and invasion | ↓E-cadherin, sE-cad, vimentin, slug; ↑N-cadherin, snail, MMP-9 | [45] |
| In vitro | A549 cells | 25, 50 and 100 μM, 48 h | Inhibiting proliferation; Inducing apoptosis | ↓14-3-3 proteins, p-Bad, p-AKT/AKT, Caspase-9, PARP; ↑Cleaved-caspase-9, Cleaved-PARP | [46] |
| In vitro | A549 cells | 5, 10, 20 and 40 μM, 24, 48, 72 and 96 h | Inhibiting proliferation; Inducing apoptosis and autophagy | ↓p-Akt, p-mTOR, p62, LC3-I; ↑Beclin1, LC3-II | [47] |
| In vitro | A549 cells | 10, 20 and 40 μM, 12, 24 and 48 h | Inhibiting migration and invasion | ↓miR-25-5p; ↑miR-330-5p | [48] |
| In vitro | A549 and H1299 cells | 0.5, 1, 5, 10 and 20 µM, 24, 48 and 72 h | Inhibiting colony formation; Promoting apoptosis and autophagy | ↓p-mTOR, p-S6, p-PI3K, p-Akt ↑LC3-II/ LC3-I, Beclin-1 | |
| Colorectal cancer | |||||
| In vitro In vivo | TCO1 and TCO2 cells; SCID mice with organoid cells | 0.6, 2, 6 and 20 µg/mL, 72 h; 20 mg/day, 21 days | Inducing necrotic lesions and apoptosis; Inhibiting stemness and proliferation | ↓cyclin D1, c-MYC, p-ERK, CD44, CD133, LGR5 | [49] |
| In vitro In vivo | CC531 cells; tumor-bearing rats with CC531 cells | 15, 20, 25 and 30 µM, 24, 48 and 72 h; 200 mg/kg/day, 28 days | Reducing proliferation and migration; Diminishing global tumor progression | ↑AST, ALP, albumin; ↓cholinesterase, cholesterol, and total protein | [50] |
| In vitro | SW620 cells | 1, 5 and 25 μM, 48 h | Inhibiting tumor sphere formation; Inducing apoptosis and autophagy | ↓GP1BB, COL9A3, COMP, AGRN, ITGB4, LAMA5, COL2A1, ITGB6, LGR5, TFAP2A, ECM; ↑Autolysosomes, autophagosomes | [51] |
| In vitro In vivo | SW480 and HT-29 cells; BALB/c nude mice with SW480 cells | 10, 20, 30, 40, 50 and 60 µM, 24 h; 100 mg/kg/day, 3 weeks | Inhibiting proliferation and tumor volume and weight; Inducing apoptosis | ↓NNMT, p-STAT3, G2/M phase cell cycle arrest; ↑ROS | [52] |
| In vitro | HCT-116/L-OHP cells | 10, 20, 30 and 40 µM, 48 h | Inhibiting proliferation, migration and invasion; Arresting cell cycle distribution | ↓ERCC1, Bcl-2, GST-π, MRP, P-gp; ↑miR-409-3p | [53] |
| In vitro | 5-FU resistant HCT-116 cells | 5, 10, 20 and 40 μM, 48 h | Inhibiting proliferation; Inducing apoptosis; Blocking G0/G1 phase | ↓E-cadherin, β-catenin, TCF4, Axin; ↑TET1, NKD2, vimentin | [54] |
| In vitro | SW480 cells | 0.1, 0.2 and 0.4 µM, 24 h | Inhibiting EMT and the expression of DNMTs | ↑E-cadherin; ↓N-cadherin, twist, snail, vimentin, CDX2, DNMT1, DNMT3a, Wnt3a, β-catenin | [55] |
| In vitro In vivo | HCT8 and HCT8/DDP cells; Nude mice with HCT8/DDP cells | 10 μM, 48 h; 1 g/kg/week, 42 days | Reducing tumor volume and weight; Promoting apoptosis | ↓Bcl-2, KCNQ1OT1; ↑cytochrome C, Bax, Cleaved-caspase-3, Cleaved-PARP1, miR-497 | [56] |
| In vitro | HCT116, HCT8, SW480 and SW620 cells | 10 μM, 24 h | Reducing clone formation | ↑NBR2, p-AMPK, p-ACC; ↓p-S6K/p-S6, Mtor, S-phase | [57] |
| In vitro | SW480 and 5FU-SW480 cells | 5, 10, 15, 20, 25, 30, and 50 μM, 48 and 72 h | Inducing apoptosis; Decreasing colony formation and migration | ↓insulin, IGF-1 receptors | [58] |
| In vitro, In vivo | HCT116/OXA and HCT116 cells; BALB/c nude mice with HCT116/OXA cells | 1, 2, 4, 8, 16, 32 and 64 μM, 48 h; 60 mg/kg, 3 weeks | Inhibiting tumor volumes and weights; Decreasing the migratory ability | ↓p-p65, Bcl-2, p-Smad2, p-Smad3, N-cadherin, TGF-β; ↑Cleaved-caspase3, E-cadherin | [59] |
| In vitro | HT-29 and DLD-1 cells | 15, 20 and 25 μM, 48 h | Inducing apoptosis and G2/M cell cycle arrest | ↓p-Akt, p-Bad, Bcl-2, GPX1, GPX4; ↑ROS, HSP27, Bad, cPARP, Beclin 1, p62 | [60] |
| In vitro In vivo | SW480 cells; female nude mice with SW480 cells | 40 μM, 24 h; 200 mg/kg, 5 days | Suppressing proliferation | ↓β-catenin, TCF4, miR-21, miR-130a; ↑Nkd2 | [61] |
| In vitro | HCT-116 and HCT-8 cells | 2.5, 5, 10, 20 and 40 µM, 24 h | Inhibiting proliferation, migration and stem-cell like characteristics | ↑CD44 | [62] |
| Head and Neck Cancer | |||||
| In vitro In vivo | HNSCC cell lines SNU1076, SNU1041, FaDu and SCC15; C57BL/6 mice with SCC15 cells | 1, 2, 5, 10, 20, 40 and 80 µM, 1, 3, 6, 12 and 24 h; 50 mg/kg, 6 weeks | Inhibiting cell viability, invasion, EMT, and tumor formation and growth; Enhancing ability of effector T cells to kill cancer cells and immune response to tumors | ↓p-STAT3, TIM-3+CD4+ T cells, PD-1+CD8+ T cells, TIM-3+CD8+ T cells, CD4+CD25+FoxP3+ Treg cells, PD-1, TIM-3; ↑E-cadherin, CD8+ T cells, IFN-γ | [63] |
| In vitro | SCC-9, FaDu and HaCaT cells | 50, 25, 10, 5, 2.5, 1.25 and 0.75 μM, 24 and 48 h | Reducing cell viability; Inducing cell cycle arrest; Modifying cytoskeleton organization | ↓procaspase-3, EGFR, PLD1, RPS6KA1, p-mTOR, p-AKT, PI3K; ↑Caspase-3, PRKCG, EGF | [64] |
| Gastric cancer | |||||
| In vitro | AGS cells | 10, 20, 30, 40, 50, 60, 70, 80, 90 and 100 µM, 24, 48 and 72 h; 50 mg/kg, 6weeks | Inducing apoptosis; Suppressing proliferation | ↓Bcl-2, survivin; ↑Bax, the proportion of Sub-G1 cells | [65] |
| In vitro | MGC-803 cells | 5, 10, 15, 20, 40 and 60 μM, 24, 48 and 72 h | Inhibiting proliferation and migration; Promoting mitochondrial and DNA damage, and apoptosis | ↓Δ ψm, cyclin E1, DNMT1, p-Rb, methylated CpG sites; ↑ROS, ATM, ATR, GADD45A, p21, p-p53, p-γH2AX | [66] |
| In vitro | SGC-7901 cells | 10, 20, 40 and 80 µM, 48 h | Suppressing proliferation, invasion, and cytoskeletal remodeling ability; Inducing apoptosis | ↓Gli1, Foxm1, β-catenin, pseudopods, skeleton fibers, vimentin; ↑S stage, E-cadherin | [67] |
| In vitro In vivo | SGC-7901 cells; BALB/c male nude mice with SGC-7901 cells | 50 μM, 24, 48 and 96 h | Decreasing migration, invasion and growth of transplanted tumors; Promoting cell apoptosis | ↓Bcl-2, cyclin D1, CDK4; ↑miR-34a | [68] |
| In vitro | SGC-7901 and BGC-823 cells | 10, 20 and 40 μM, 24 h | Inhibiting proliferation; Promoting apoptosis and autophagy | ↓Bcl-2, Bcl-xL, LC3I, PI3K, p-Akt, p-mTOR; ↑Bax, Beclin1, ATG3, Cleaved-caspase-3, Cleaved-PARP, ATG5, LC3II, p53, p21 | [69] |
| In vitro In vivo | SGC-7901 cells; Balc/c nude mice with SGC7901 cells | 25 μM, 3, 5 and 7 days; 100 mg/kg, 2 weeks | Inhibiting proliferation, gastrin and gastric acid secretion; Promoting apoptosis | ↑Caspase-3 | [70] |
| Bladder cancer | |||||
| In vitro | T24 and RT4 cells | 10, 15, 20 and 25 µM, 48 and 72 h | Inhibiting cell growth, migration and invasion; Inducing cell cycle arrest | ↓Trop2, cyclin E1; ↑G2/M cell populations, p27 | [71] |
| In vitro | J82, TCCSUP and T24 cells | 1, 5, 10 and 20 µM, 24, 48 and 72 h | Decreasing invasion and tumorigenicity; Increasing apoptosis | ↓miR-7641; ↑p16 | [72] |
| Prostate Cancer | |||||
| In vitro | PC-3 and DU145 cells | 10, 20, 30, 40 and 50 µM, 12, 24 and 48 h | Reducing cell viability, migration and invasion; Promoting apoptosis | ↓PCLAF, Bcl-2, Caspase-3; ↑miR-30a-5p, Bax, Cleaved-caspase-3 | [73] |
| In vitro | Prostate-CAFs, PC-3 and NAFs cells | 10, 20 and 30 μM, 8, 12 and 24 h | Inducing apoptosis and ER stress; Regulating cell cycle | ↓Bcl-2, ΔΨm; ↑Cleaved-caspase-3, Bax, Bims, Cleaved-PARP, Puma, p-p53, ROS, p-ERK, p-eIF2α, CHOP, ATF4 | [74] |
| In vitro In vivo | LNCaP and 22Rv1 cells; male TRAMP mice | 5, 25 and 50 μM, 24, 48 and 72 h; 200 mg/kg/day, 30days | Inhibiting growth; Inducing apoptosis | ↓CYP11A1, HSD3B2, StAR, testosterone, dihydrotestosterone; ↑AKR1C2, SRD5A1, CYP17A1 | [75] |
| In vitro | 22RV1, PC-3 and DU145 cells | 1, 5, 10 and 20 μM, 4 days | Suppressing proliferation | ↓cyclin D1, PCNA, β-catenin, c-MYC; ↑p21, miR-34a | [76] |
| Thyroid cancer | |||||
| In vitro | K1, FTC-133, BCPAP and 8505C cells | 10, 12.5, 20, 25, 30, 40 and 50 µM, 24 and 72 h | Inhibiting cell growth; Inducing autophagy | ↑LC3-II, Beclin-1, p-p38, p-JNK, p-ERK1/2; ↓p62, p-PDK1, p-Akt, p-p70S6, p-p85S6, p-S6, p-4E-BP1 | [77] |
| In vitro | TPC-1 and BCPAP-R cells | 2.5, 5, 10, 20 and 40 µM, 24 h | Inhibiting cell viability, invasion, migration and EMT | ↓MMP-9, MMP-2, N-cadherin, vimentin, fibronectin, p-JAK, p-JAK2, p-JAK3, p-STAT1, p-STAT2; ↑E-cadherin, miR-301a-3p | [78] |
| Liver cancer | |||||
| In vitro In vivo | HepG2, Huh-7 and MHCC-97H cells; BALB/c-nu nude mice with HepG2 cells | 1.2, 2.4, 4.8 and 9.6 µg/mL, 24 and 48 h; 120 and 240 mg/kg/day, 15 days | Reducing tumor volume and weight, and angiogenesis | ↓MDSCs, GM-CSF, G-CSF, TLR4, MyD88, p-IKKα, p-IKKβ, NF-κB, TNF-α, IL-6, IL-1β, PGE2, COX-2, VEGF, CD31, α-smooth | [79] |
| In vitro | HepG2 and HuT78 cells | 5 and 10 μM, 24 h | Inducing cell death | ↓lactate, ldh-a, mct-1, mdr-1, stat-3, HIF-1α, HCAR-1; ↑NO | [80] |
| In vitro | HepG2 cells | 20, 50, 80 and 100 μM, 24, 48 and 72 h | Inhibiting proliferation, migration and invasion; Promoting apoptosis | ↓HSP70, eHSP70, TLR4 | [25] |
| In vitro In vivo | Bel-7,402 and HepG2 cells; male BALB/c mice with H22 cells | 15 and 30 μM, 24, 48 and 72 h; 100 mg/kg/day, 14 days | Inducing apoptosis, G2/M cell cycle arrest; Modulating gut microbiota | ↓p-PI3K, p-Akt, p-mTOR, tumors weights and sizes; ↑Cleaved-caspase-3, Lactobacillus, Epsilonbacteraeota, Helicobacterac-eae, Campylobacterales, Helicobacter, Escherichia-shigella, Bifidobacterium, Campylobacteria | [81] |
| In vitro In vivo | HepG2 and SK-HEP1 cells; male BALB/c mice H22 and HepG2 cells | 20, 40, 60, 80, 100, 120 and 140 nM, 24 h; 100 mg/kg curcumin or Zn (II)-curcumin, 2 weeks | Inhibiting tumor growth; Regulating gut microbiota; Improving intestinal permeability | ↓Firmicutes, unclassified Lachnospiraceae, Clostridium cluster XIVa, Pseudoflavonifractor, Oscillibacter; ↑Bacteroidetes, Barnesiella, Unclassified_Porphyromonadaceae, Paraprevotella, Prevotella, zonula occludens-1, occludin | [82] |
| Ovarian cancer | |||||
| In vitro | SKOV3 cells | 10, 20, 30, 40 and 50 μM, 6, 12 and 24 h | Inhibiting migration and invasion | ↓STAT3, fascin | [83] |
| In vitro | SKOV3 cells | 20 μM, 96 h | Inhibiting cell migration and EMT | ↓DNMT3a, β-catenin, cyclin D1, c-Myc, fibronectin, vimentin; ↑SFRP5, E-cadherin | [84] |
| In vitro | SK-OV-3 and A2780 cells | 5, 10, 20, 40 and 80 μM, 24, 48 and 72 h | Inducing apoptosis and autophagy | ↓p62, p-AKT, p-mTOR, p-p70S6K; ↑Caspase-9, PARP, Atg3, Beclin-1, LC3B-I/II | [85] |
| In vitro In vivo | SKOV3 and A2780 cells; BALB/c athymic mice with A2780 cells | 10, 20 and 40 μM, 24, 48 and 72 h; 15 mg/kg/2days, 5 weeks | Inhibiting proliferation; Promoting apoptosis | ↓PCNA, miR-320a; ↑Bax, Cleaved-caspase-3, circ-PLEKHM3, SMG1 | [86] |
| Oral Cancer | |||||
| In vitro | HSC-4 and Ca9-22 cells | 15 μM, 48 h | Decreasing invasion, migration and EMT | ↓vimentin, p-c-Met, p- ERK, pro-MMP9; ↑E-cadherin | [87] |
| Pancreatic Cancer | |||||
| In vitro | Panc-1 and MiaPaCa-2 cells | 6, 10 and 12 µM, 24 h | Reducing cell survival; Inducing apoptosis and DNA damage | ↓G0/G1-fraction; ↑yH2AX-MFI, G2/M-fraction, S-phase cells | [88] |
| In vitro | PANC-1 cells | 2.5, 5, 10 and 20 µM, 72 h | Inducing apoptosis | ↑Cleaved-caspase-3, miR-340, Cleaved-PARP; ↓PARP, XIAP | [89] |
| In vitro | Patu8988 and Panc-1 cells | 5, 10, 15 and 20 μM, 48 and 72 h | Inhibiting migration and invasion; Inducing apoptosis | ↓NEDD4, p-Akt, p-mTOR; ↑PTEN, p73, β-TRCP | [90] |
| Cervical Cancer | |||||
| In vitro | Siha cells | 5, 15, 30 and 50 µM, 6, 12, 24 and 48 h | Inhibiting proliferation; Inducing G2/M cell cycle arrest, apoptosis, autophagy | ↓cyclins B1, cdc25; ↑ROS, p62, LC3I/II, Cleaved-caspase-3, Cleaved-PARP, p53, p21 | [91] |
| In vitro | Siha cells | 20 µM, 72 h | Decreasing EMT and migration | ↓N-cadherin, vimentin, slug, Zeb1, PIR, pirin; ↑E-cadherin | [92] |
| Tongue Cancer | |||||
| In vitro | CAL 27 cells | 10, 25, 50 and 100 µM, 16 and 24 h | Inhibiting proliferation and migration; Promoting apoptosis and S-phase cell cycle arrest | ↓Bcl-2; ↑Bax, Cleaved-caspase-3, S-phase cells | [93] |
| Brain Cancer | |||||
| In vitro | SNB19 and A1207 cells | 10, 15, 20 and 25 µM, 48 and 72 h | Suppressing proliferation, migration and invasion; Inducing apoptosis and cell cycle arrest | ↓NEDD4, Notch1, p-Akt; ↑G2/M phase | [94] |
| Therapy | Study Type | Subjects | Administration Methods | Dose & Duration | Outcomes | Ref. |
|---|---|---|---|---|---|---|
| Cervical Cancer | ||||||
| Curcumin + radiation | Quasi-experiment | 40 advanced cervical cancer patients | Oral administration | 4 g/day, 7 days | Lowering survivin levels | [133] |
| Breast Cancer | ||||||
| Curcumin + paclitaxel | RCT | 150 women with metastatic and advanced breast cancer | Intravenous administration | 300 mg/week (curcumin), 12 weeks; 80 mg/m2 body surface area/week (paclitaxel), once a week for 12 consecutive weeks | Improving objective response rate and patient self-assessed overall performance status | [134] |
| Colorectal Cancer | ||||||
| Curcumin C3 complex + standard-of-care chemotherapy (FOLFOX ± bevacizumab) | Open-labelled RCT | 27 patients with stage IV disease metastatic colorectal cancer, aged >18 y | Oral administration | 2 g curcumin C3 complex/d (80% curcumin and 20% dimethoxy-curcumin and bisdemethoxycurcumin), ≤ 12 cycles of chemotherapy | Curcumin was safe and tolerable, increasing overall survival and objective response rate | [138] |
| Curcuminoids capsules | RCT | 72 patients with stage 3 colorectal cancer, aged >20 y | Oral administration | 500 mg/day, 8 weeks | Lowering serum C-reactive protein levels, enhancing functional scales and the global quality of life | [139] |
| Prostate Cancer | ||||||
| Curcumin | RCT | 97 prostate cancer patients | Oral administration | 1440 mg/day, 6–36 months | Reducing prostate specific antigen | [135] |
| Nanocurcumin | RCT | 64 prostate cancer patients | Oral administration | 120 mg/day, 3 days before and during radiotherapy | Not efficacious | [136] |
| Curcumin + chemo-therapy with docetaxel | Phase II RCT | 50 metastatic castration-resistant prostate cancer patients, aged >18 y | Oral administration | 6 g/d (curcumin), 3 weeks; 75 mg/m2 body surface area (docetaxel), first day of every 3 weeks for 6 cycles | Not efficacious | [137] |
| Pancreatic Cancer | ||||||
| Gemcitabine + Meriva® (curcumin complexed with soy lecithin, 1:2) | Single center, single arm, prospective phase II trial | 52 pancreatic cancer patients, aged >18 y | Oral administration | 2000 mg (Meriva®), 28 day; 10 mg/m2/min (gemcitabine), on days 1, 8, 15 | Raising the efficiency of gemcitabine translating in a response rate | [140] |
| Endometrial Cancer | ||||||
| Curcumin phytosome | Open-label, non-randomized phase II study | 7 endometrial cancer patients | Oral administration | 2 g/day, 2 weeks | Reducing major histocompatibility complex expression levels on leukocytes, inducible T cell costimulatory expression by CD8+ T cells and the frequency of monocytes, increasing CD69 levels on CD16− NK cells | [141] |
| Oral Cancer | ||||||
| APG-157 (including curcumin) | Phase II RCT | 13 normal subjects and 12 patients with oral cancer | Oral administration | 100 and 200 mg, each hour for 3 consecutive hours | Reducing inflammation, Bacteroides and ratio of Firmicutes/Bacteroidetes | [142] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.-J.; Huang, S.-Y.; Zhou, D.-D.; Xiong, R.-G.; Zhao, C.-N.; Fang, A.-P.; Zhang, Y.-J.; Li, H.-B.; Zhu, H.-L. Effects and Mechanisms of Curcumin for the Prevention and Management of Cancers: An Updated Review. Antioxidants 2022, 11, 1481. https://doi.org/10.3390/antiox11081481
Yang Z-J, Huang S-Y, Zhou D-D, Xiong R-G, Zhao C-N, Fang A-P, Zhang Y-J, Li H-B, Zhu H-L. Effects and Mechanisms of Curcumin for the Prevention and Management of Cancers: An Updated Review. Antioxidants. 2022; 11(8):1481. https://doi.org/10.3390/antiox11081481
Chicago/Turabian StyleYang, Zhi-Jun, Si-Yu Huang, Dan-Dan Zhou, Ruo-Gu Xiong, Cai-Ning Zhao, Ai-Ping Fang, Yun-Jian Zhang, Hua-Bin Li, and Hui-Lian Zhu. 2022. "Effects and Mechanisms of Curcumin for the Prevention and Management of Cancers: An Updated Review" Antioxidants 11, no. 8: 1481. https://doi.org/10.3390/antiox11081481
APA StyleYang, Z.-J., Huang, S.-Y., Zhou, D.-D., Xiong, R.-G., Zhao, C.-N., Fang, A.-P., Zhang, Y.-J., Li, H.-B., & Zhu, H.-L. (2022). Effects and Mechanisms of Curcumin for the Prevention and Management of Cancers: An Updated Review. Antioxidants, 11(8), 1481. https://doi.org/10.3390/antiox11081481