
The Application of the Neuroprotective and Potential Antioxidant Effect of Ergotamine Mediated by Targeting N-Methyl-D-Aspartate Receptors

 ,
,
Abstract
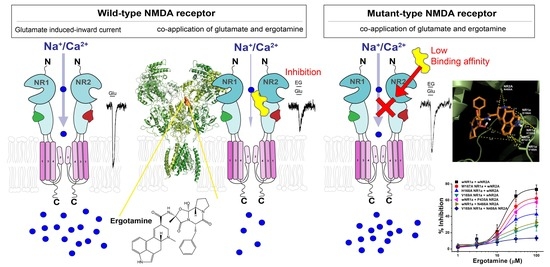
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Xenopus Oocytes and Microinjection
2.3. NR1a/NR2A Receptor Mutation and In Vitro Transcription of cDNAs
2.4. Molecular Docking Study with Three-Dimensional (3D) Modeling
2.5. Data Recording
2.6. Data Analysis
3. Results
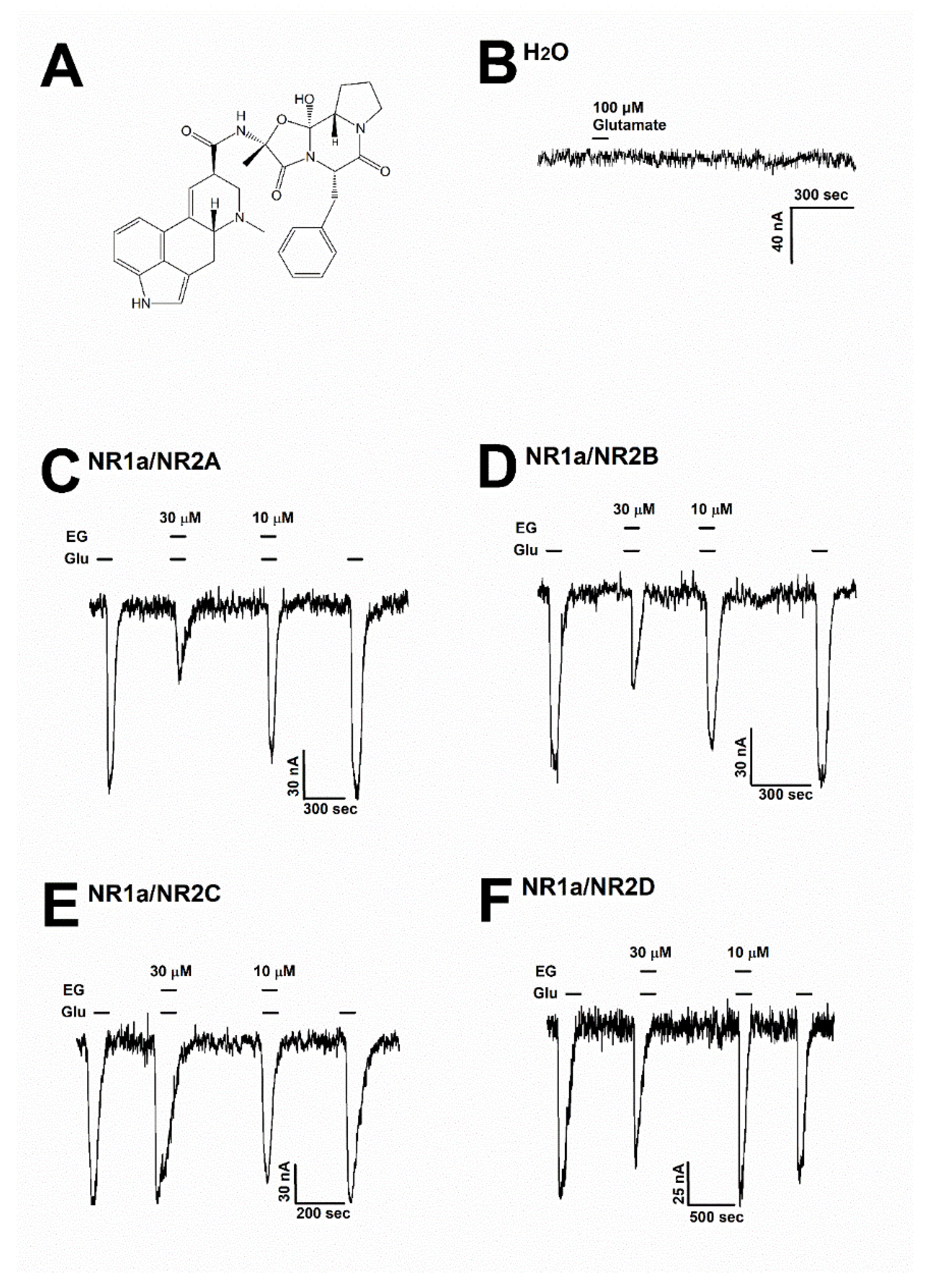
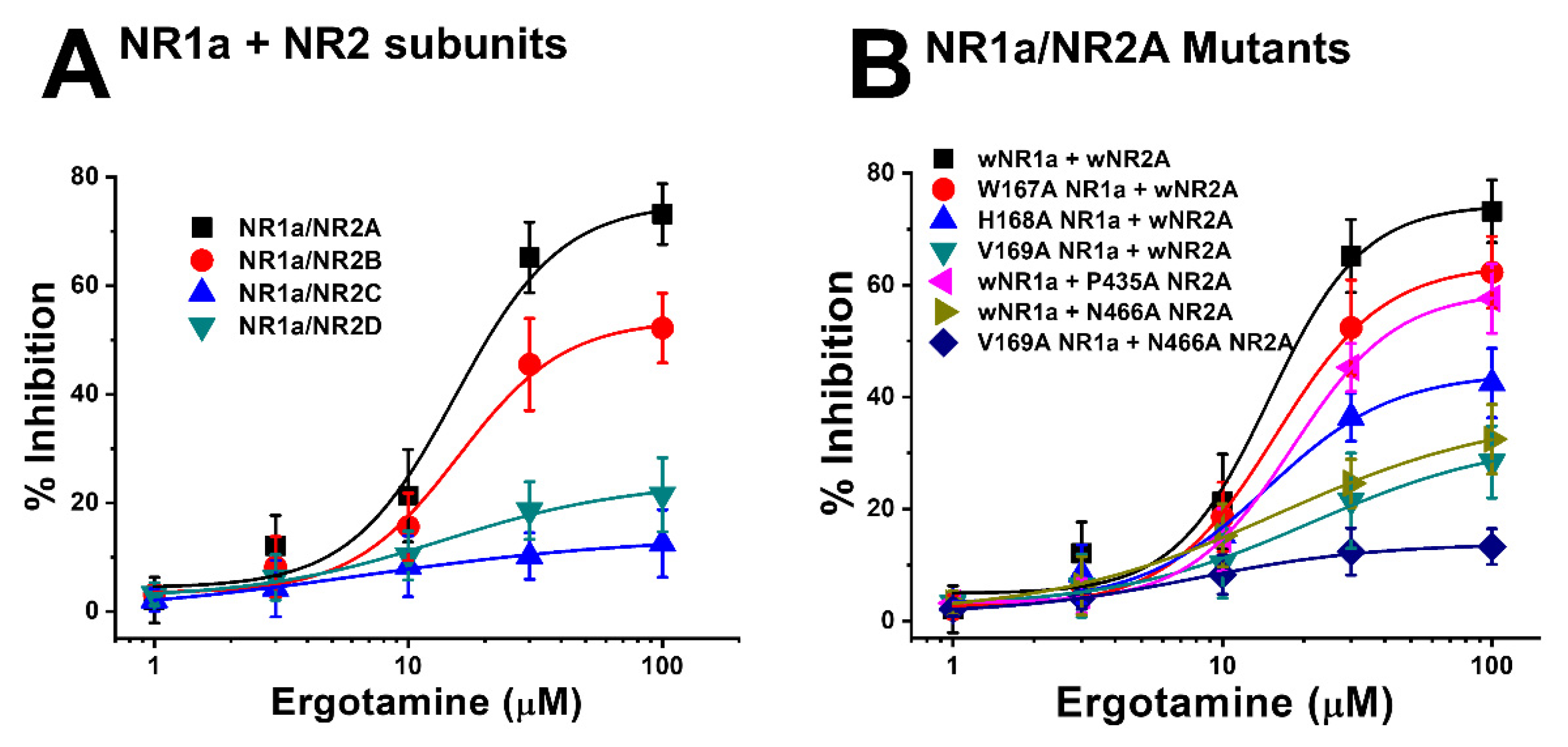
3.1. Inhibitory Effect of Ergotamine on Various N-Methyl-D-Aspartate Receptors
3.2. Ergotamine Concentration-Dependent Inhibition of IGlu
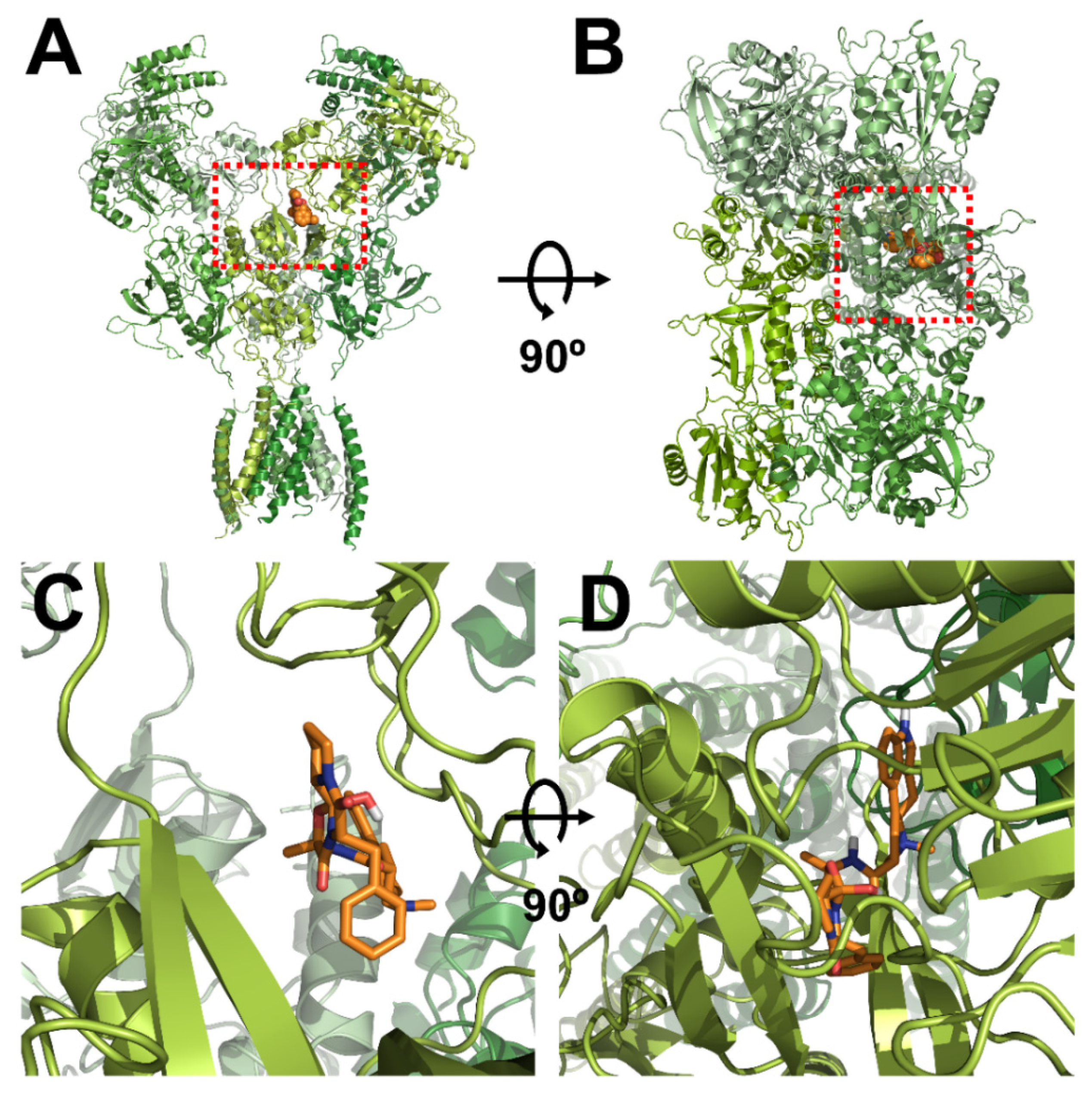
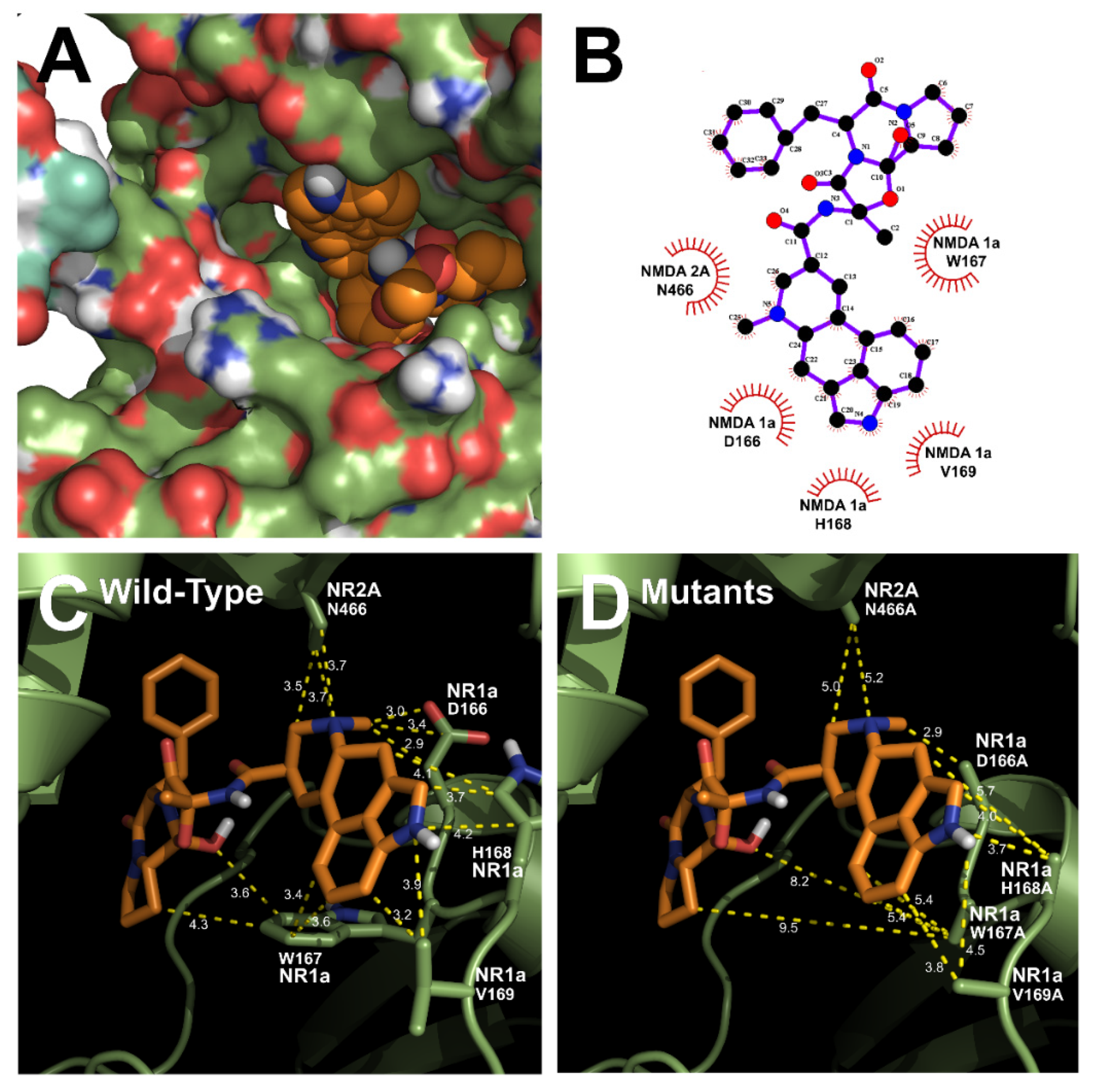
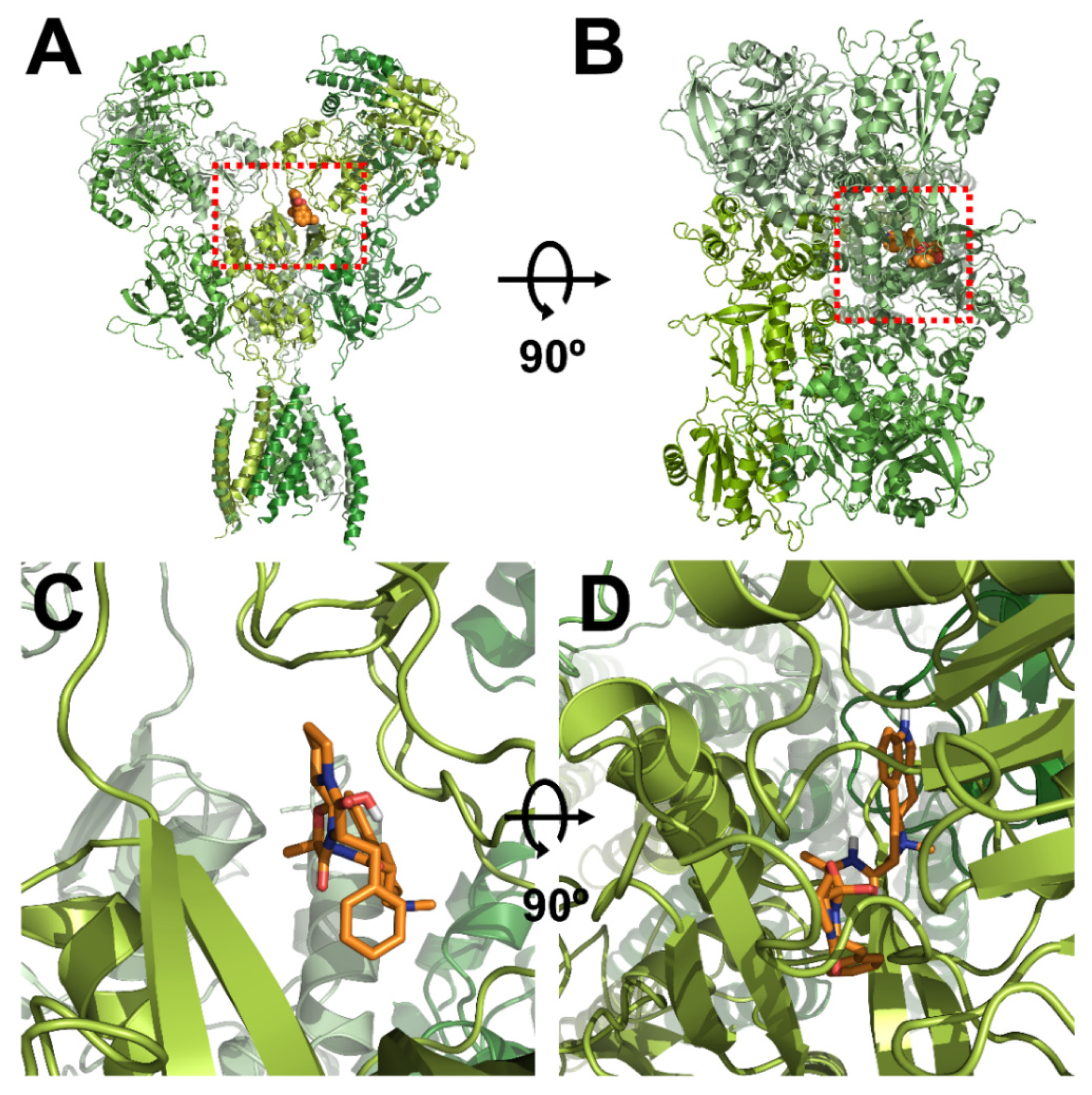
3.3. In Silico Docking of Ergotamine to Wild-Type and Mutant NR1a/NR2A Subunits
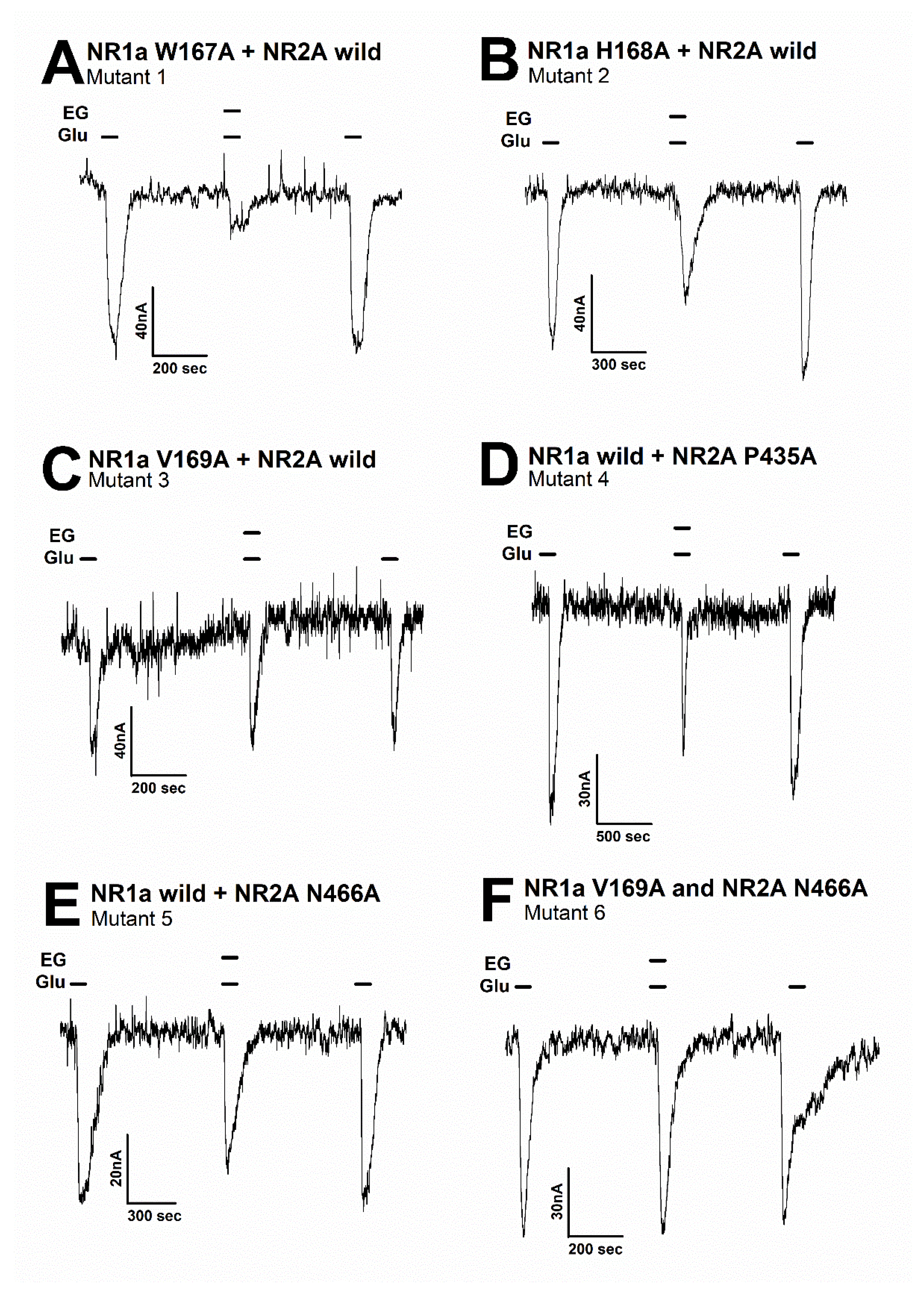
3.4. Effect of Ergotamine in Various NR1a/NR2A Subunit Mutations
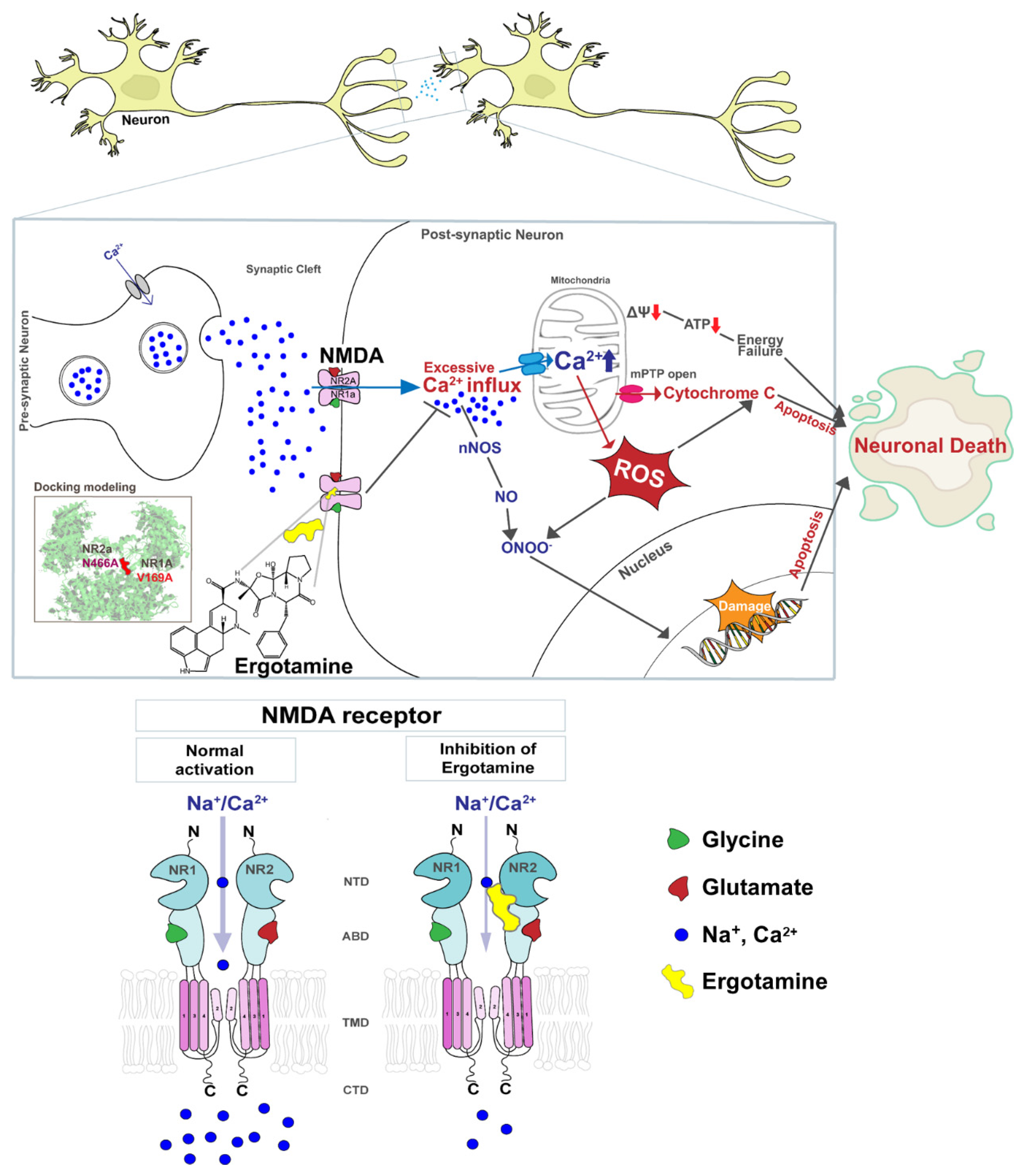
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nicoll, R.A.; Roche, K.W. Long-term potentiation: Peeling the onion. Neuropharmacology 2013, 74, 18–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure, regulation, and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA receptor subunit diversity: Impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Smothers, C.T.; Woodward, J.J. Pharmacological characterization of glycine-activated currents in HEK 293 cells expressing N-methyl-D-aspartate NR1 and NR3 subunits. J. Pharmacol. Exp. Ther. 2007, 322, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Masuko, T.; Kashiwagi, K.; Kuno, T.; Nguyen, N.D.; Pahk, A.J.; Fukuchi, J.I.; Igarashi, K.; Williams, K. A regulatory domain (R1–R2) in the amino terminus of theN-methyl-D-aspartate receptor: Effects of spermine, protons, and ifenprodil, and structural similarity to bacterial leucine/isoleucine/valine binding protein. Mol. Pharmacol. 1999, 55, 957–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakas, E.; Simorowski, N.; Furukawa, H. Structure of the zinc-bound amino-terminal domain of the NMDA receptor NR2B subunit. EMBO J. 2009, 28, 3910–3920. [Google Scholar] [CrossRef] [Green Version]
- Karakas, E.; Simorowski, N.; Furukawa, H. Subunit arrangement and phenylethanolamine binding in GluN1/GluN2B NMDA receptors. Nature 2011, 475, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Romero-Hernandez, A.; Furukawa, H. Novel mode of antagonist binding in NMDA receptors revealed by the crystal structure of the GluN1-GluN2A ligand-binding domain complexed to NVP-AAM077. Mol. Pharmacol. 2017, 92, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Slikker, W., Jr.; Paule, M.G.; Wang, C. Handbook of Developmental Neurotoxicology; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Banerjee, A.; Larsen, R.; Philpot, B.D.; Paulsen, O. Roles of presynaptic NMDA receptors in neurotransmission and plasticity. Trends Neurosci. 2016, 39, 26–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deutschenbaur, L.; Beck, J.; Kiyhankhadiv, A.; Mühlhauser, M.; Borgwardt, S.; Walter, M.; Hasler, G.; Sollberger, D.; Lang, U.E. Role of calcium, glutamate and NMDA in major depression and therapeutic application. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 64, 325–333. [Google Scholar] [CrossRef]
- Hedegaard, M.; Hansen, K.B.; Andersen, K.T.; Bräuner-Osborne, H.; Traynelis, S.F. Molecular pharmacology of human NMDA receptors. Neurochem. Int. 2012, 61, 601–609. [Google Scholar] [CrossRef] [Green Version]
- Duchen, M.R. Mitochondria, calcium-dependent neuronal death and neurodegenerative disease. Pflügers Arch.-Eur. J. Physiol. 2012, 464, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delierneux, C.; Kouba, S.; Shanmughapriya, S.; Potier-Cartereau, M.; Trebak, M.; Hempel, N. Mitochondrial calcium regulation of redox signaling in cancer. Cells 2020, 9, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfraind, T. Antioxidant effects and the therapeutic mode of action of calcium channel blockers in hypertension and atherosclerosis. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 2259–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernorudskiy, A.L.; Zito, E. Regulation of calcium homeostasis by ER redox: A close-up of the ER/mitochondria connection. J. Mol. Biol. 2017, 429, 620–632. [Google Scholar] [CrossRef]
- Sibarov, D.; Antonov, S. Calcium-dependent desensitization of NMDA receptors. Biochemistry 2018, 83, 1173–1183. [Google Scholar] [CrossRef]
- Mammucari, C.; Raffaello, A.; Reane, D.V.; Gherardi, G.; De Mario, A.; Rizzuto, R. Mitochondrial calcium uptake in organ physiology: From molecular mechanism to animal models. Pflügers Arch.-Eur. J. Physiol. 2018, 470, 1165–1179. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Rodriguez, M.; Bacskai, B.J. High mitochondrial calcium levels precede neuronal death in vivo in Alzheimer’s disease. Cell Stress 2020, 4, 187. [Google Scholar] [CrossRef]
- Zhivotovsky, B.; Orrenius, S. Calcium and cell death mechanisms: A perspective from the cell death community. Cell Calcium 2011, 50, 211–221. [Google Scholar] [CrossRef]
- Silberstein, S.D.; McCrory, D.C. Ergotamine and dihydroergotamine: History, pharmacology, and efficacy. Headache J. Head Face Pain 2003, 43, 144–166. [Google Scholar] [CrossRef]
- Tfelt-Hansen, P.; Saxena, P.R.; Dahlöf, C.; Pascual, J.; Láinez, M.; Henry, P.; Diener, H.-C.; Schoenen, J.; Ferrari, M.D.; Goadsby, P.J. Ergotamine in the acute treatment of migraine: A review and European consensus. Brain 2000, 123, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Seol, H.-S.; Eom, S.; Lee, J.; Kim, C.; Park, J.-H.; Kim, T.-H.; Lee, J.H. Hydroxy Pentacyclic Triterpene Acid, Kaempferol, Inhibits the Human 5-Hydroxytryptamine Type 3A Receptor Activity. Int. J. Mol. Sci. 2022, 23, 544. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, S.; Guillemin, G.J.; Abiramasundari, R.S.; Essa, M.M.; Akbar, M.; Akbar, M.D. The role of reactive oxygen species in the pathogenesis of Alzheimer’s disease, Parkinson’s disease, and Huntington’s disease: A mini review. Oxidative Med. Cell. Longev. 2016, 2016, 8590578. [Google Scholar] [CrossRef] [PubMed]
- Behl, C.; Moosmann, B. Antioxidant neuroprotection in Alzheimer’s disease as preventive and therapeutic approach. Free. Radic. Biol. Med. 2002, 33, 182–191. [Google Scholar] [CrossRef]
- Teleanu, R.I.; Chircov, C.; Grumezescu, A.M.; Volceanov, A.; Teleanu, D.M. Antioxidant therapies for neuroprotection—A review. J. Clin. Med. 2019, 8, 1659. [Google Scholar] [CrossRef] [Green Version]
- Geiszt, M.; Leto, T.L. The Nox family of NAD (P) H oxidases: Host defense and beyond. J. Biol. Chem. 2004, 279, 51715–51718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, L.B.; Nelson, K.J. Discovering mechanisms of signaling-mediated cysteine oxidation. Curr. Opin. Chem. Biol. 2008, 12, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Beal, M.F. Aging, energy, and oxidative stress in neurodegenerative diseases. Ann. Neurol. 1995, 38, 357–366. [Google Scholar] [CrossRef]
- Tramutola, A.; Lanzillotta, C.; Perluigi, M.; Butterfield, D.A. Oxidative stress, protein modification and Alzheimer disease. Brain Res. Bull. 2017, 133, 88–96. [Google Scholar] [CrossRef]
- Rauner, C.; Köhr, G. Triheteromeric NR1/NR2A/NR2B receptors constitute the major N-methyl-D-aspartate receptor population in adult hippocampal synapses. J. Biol. Chem. 2011, 286, 7558–7566. [Google Scholar] [CrossRef] [Green Version]
- Tovar, K.R.; McGinley, M.J.; Westbrook, G.L. Triheteromeric NMDA receptors at hippocampal synapses. J. Neurosci. 2013, 33, 9150–9160. [Google Scholar] [CrossRef]
- Sheng, M.; Cummings, J.; Roldan, L.A.; Jan, Y.N.; Jan, L.Y. Changing subunit composition of heteromeric NMDA receptors during development of rat cortex. Nature 1994, 368, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Cull-Candy, S.; Brickley, S.; Farrant, M. NMDA receptor subunits: Diversity, development and disease. Curr. Opin. Neurobiol. 2001, 11, 327–335. [Google Scholar] [CrossRef]
- Kumar, A. NMDA receptor function during senescence: Implication on cognitive performance. Front. Neurosci. 2015, 9, 473. [Google Scholar] [CrossRef] [PubMed]
- Guidi, M.; Kumar, A.; Foster, T.C. Impaired attention and synaptic senescence of the prefrontal cortex involves redox regulation of NMDA receptors. J. Neurosci. 2015, 35, 3966–3977. [Google Scholar] [CrossRef] [Green Version]
- Guidi, M.; Rani, A.; Karic, S.; Severance, B.; Kumar, A.; Foster, T.C. Contribution of N-methyl-D-aspartate receptors to attention and episodic spatial memory during senescence. Neurobiol. Learn. Mem. 2015, 125, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chang, L.; Song, Y.; Li, H.; Wu, Y. The role of NMDA receptors in Alzheimer’s disease. Front. Neurosci. 2019, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Hallett, P.J.; Standaert, D.G. Rationale for and use of NMDA receptor antagonists in Parkinson’s disease. Pharmacol. Ther. 2004, 102, 155–174. [Google Scholar] [CrossRef]
- Collingridge, G. The role of NMDA receptors in learning and memory. Nature 1987, 330, 604–605. [Google Scholar] [CrossRef]
- Gray, J.A.; Shi, Y.; Usui, H.; During, M.J.; Sakimura, K.; Nicoll, R.A. Distinct modes of AMPA receptor suppression at developing synapses by GluN2A and GluN2B: Single-cell NMDA receptor subunit deletion in vivo. Neuron 2011, 71, 1085–1101. [Google Scholar] [CrossRef] [Green Version]
- Kochlamazashvili, G.; Bukalo, O.; Senkov, O.; Salmen, B.; Gerardy-Schahn, R.; Engel, A.K.; Schachner, M.; Dityatev, A. Restoration of synaptic plasticity and learning in young and aged NCAM-deficient mice by enhancing neurotransmission mediated by GluN2A-containing NMDA receptors. J. Neurosci. 2012, 32, 2263–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumas, T.C. Developmental regulation of cognitive abilities: Modified composition of a molecular switch turns on associative learning. Prog. Neurobiol. 2005, 76, 189–211. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cheng, X.; Hu, J.; Gao, Z. The role of GluN2A in cerebral ischemia: Promoting neuron death and survival in the early stage and thereafter. Mol. Neurobiol. 2018, 55, 1208–1216. [Google Scholar] [CrossRef] [PubMed]
- Hafenbreidel, M.; Todd, C.R.; Mueller, D. Infralimbic GluN2A-containing NMDA receptors modulate reconsolidation of cocaine self-administration memory. Neuropsychopharmacology 2017, 42, 1113–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulac, D.; Hüwel, S.; Galla, H.-J.; Humpf, H.-U. Permeability of ergot alkaloids across the blood-brain barrier in vitro and influence on the barrier integrity. Mol. Nutr. Food Res. 2012, 56, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Imax | IC50 | nH | |
|---|---|---|---|
| NR1a/NR2A | 75.2 ± 9.7 | 15.3 ± 6.1 | 2.1 ± 1.5 |
| NR1a/NR2B | 53.7 ± 5.8 | 15.7 ± 4.8 | 2.1 ± 1.0 |
| NR1a/NR2C | 13.9 ± 3.0 | 6.5 ± 3.6 | 0.8 ± 0.5 |
| NR1a/NR2D | 24.0 ± 5.1 | 13.8 ± 6.4 | 1.2 ± 0.6 |
| Binding Energy | KI (mM) | Intermole-Cular Energy | Internal Energy | RMSD * (nm) | Binding Residues (Amino Acids) | |
|---|---|---|---|---|---|---|
| #1 | –7.54 | 0.15 | –9.54 | –1.21 | 0.21 | NR1a W167, NR1a H168, NR2A V169, NR2A P435, NR2A N466 |
| #2 | –6.54 | 1.01 | –6.54 | –1.01 | 0.19 | NR1a N210, NR1a G215, NR2A N216, NR2A C510, NR2A E551 |
| #3 | –5.44 | 1.24 | –5.25 | –1.20 | 0.19 | NR1a K155, NR1a H156, NR1a G773, NR2A N523, NR2A R767 |
| #4 | –4.57 | 1.98 | –4.25 | –1.32 | 0.18 | NR1a E528, NR2A T433, NR2A V434, NR2A S468 |
| No. | Mutants | Imax | IC50 | nH |
|---|---|---|---|---|
| NR1a wild + NR2A wild | 74.4 ± 8.4 | 14.9 ± 4.7 | 2.5 ± 1.4 | |
| 1 | NR1a W167A + NR2A wild | 63.7 ± 3.5 | 15.7 ± 2.0 | 2.2 ± 0.5 |
| 2 | NR1a H168A + NR2A wild | 44.3 ± 5.9 | 14.6 ± 4.0 | 1.9 ± 0.8 |
| 3 | NR1a V169A + NR2A wild | 32.7 ± 6.8 | 21.4 ± 10.7 | 1.2 ± 0.5 |
| 4 | NR1a wild + NR2A P435A | 58.8 ± 0.9 | 18.2 ± 0.5 | 2.3 ± 0.1 |
| 5 | NR1a wild + NR2A N466A | 37.2 ± 2.2 | 16.1 ± 2.0 | 1.0 ± 0.1 |
| 6 | NR1a V169A + NR2A N466A | 13.9 ± 0.7 | 7.8 ± 1.1 | 1.3 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Eom, S.; Nguyen, K.V.A.; Lee, J.; Park, Y.; Yeom, H.D.; Lee, J.H. The Application of the Neuroprotective and Potential Antioxidant Effect of Ergotamine Mediated by Targeting N-Methyl-D-Aspartate Receptors. Antioxidants 2022, 11, 1471. https://doi.org/10.3390/antiox11081471
Lee S, Eom S, Nguyen KVA, Lee J, Park Y, Yeom HD, Lee JH. The Application of the Neuroprotective and Potential Antioxidant Effect of Ergotamine Mediated by Targeting N-Methyl-D-Aspartate Receptors. Antioxidants. 2022; 11(8):1471. https://doi.org/10.3390/antiox11081471
Chicago/Turabian StyleLee, Shinhui, Sanung Eom, Khoa V. A. Nguyen, Jiwon Lee, Youngseo Park, Hye Duck Yeom, and Junho H. Lee. 2022. "The Application of the Neuroprotective and Potential Antioxidant Effect of Ergotamine Mediated by Targeting N-Methyl-D-Aspartate Receptors" Antioxidants 11, no. 8: 1471. https://doi.org/10.3390/antiox11081471
APA StyleLee, S., Eom, S., Nguyen, K. V. A., Lee, J., Park, Y., Yeom, H. D., & Lee, J. H. (2022). The Application of the Neuroprotective and Potential Antioxidant Effect of Ergotamine Mediated by Targeting N-Methyl-D-Aspartate Receptors. Antioxidants, 11(8), 1471. https://doi.org/10.3390/antiox11081471