
Phytochemical and Safety Evaluations of Finger Lime, Mountain Pepper, and Tamarind in Zebrafish Embryos

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Plant Extracts
2.3. Determination of Total Phenolic Content (TPC) and Antioxidant Activity
2.4. Screening for Bioactive Compounds in Plant Extracts
2.5. Quantification of Individual Bioactive Antioxidant Compounds in Plant Extracts
2.6. In Vivo Acute Toxicity Test of Australian Plants
2.7. Statistical Analysis
3. Results and Discussion
3.1. Characterization of Australian Native Plant Extracts
3.1.1. Total Phenolic Content and Antioxidant Activity
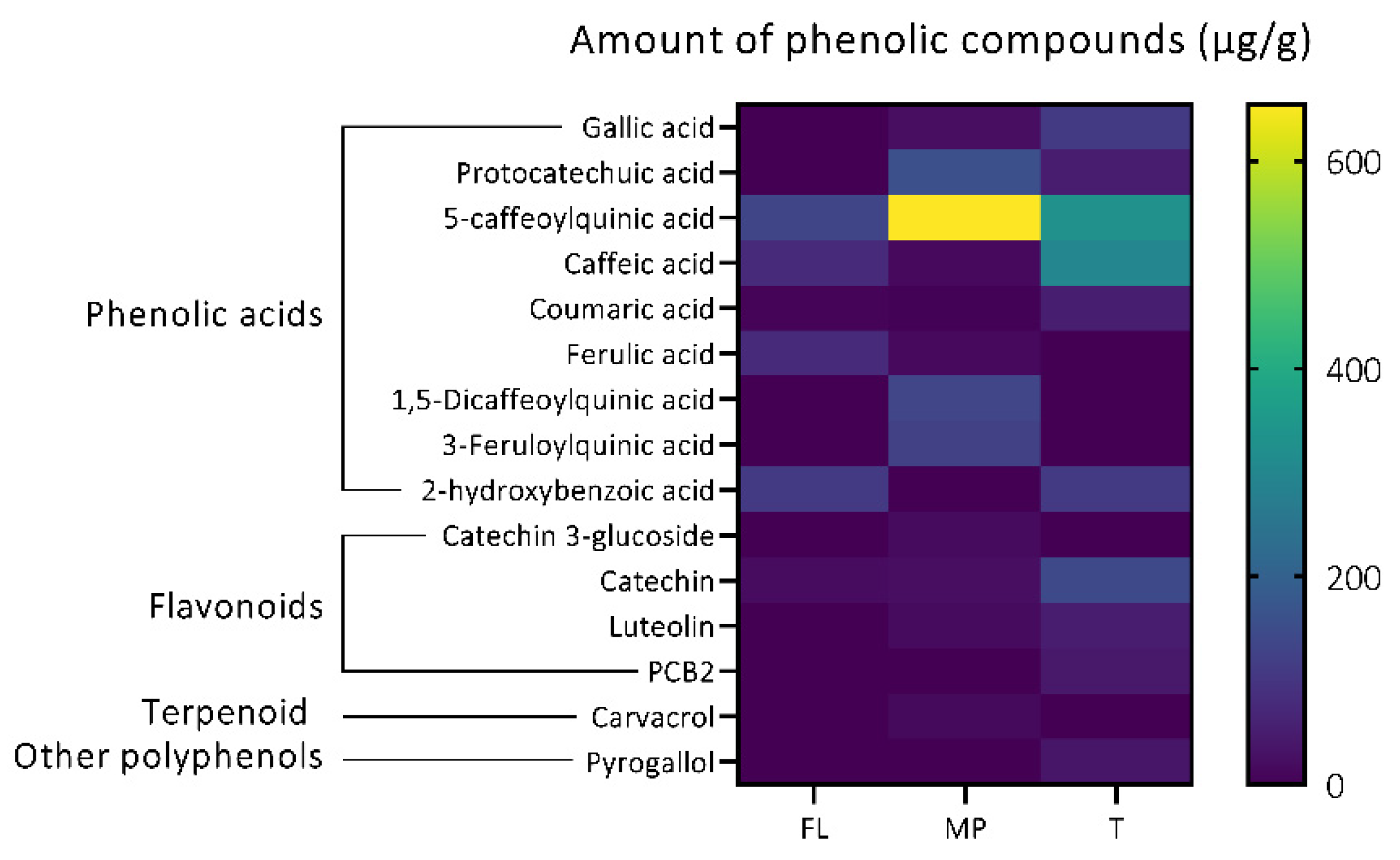
3.1.2. Identification of Individual Bioactive Antioxidant Compounds in Plant Extracts
Phenolic Acids
Flavonoids
Other Biologically Relevant Compounds Identified
3.1.3. Targeted Quantification of Bioactive Metabolites in Plant Extracts
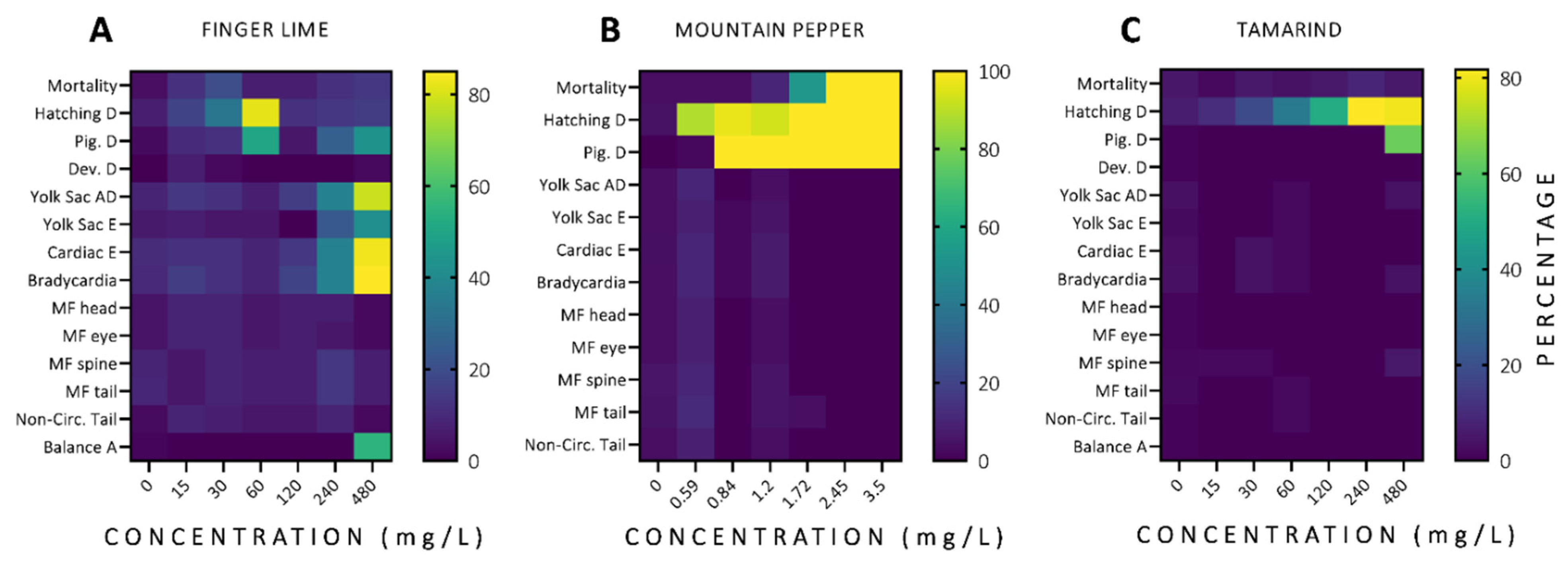
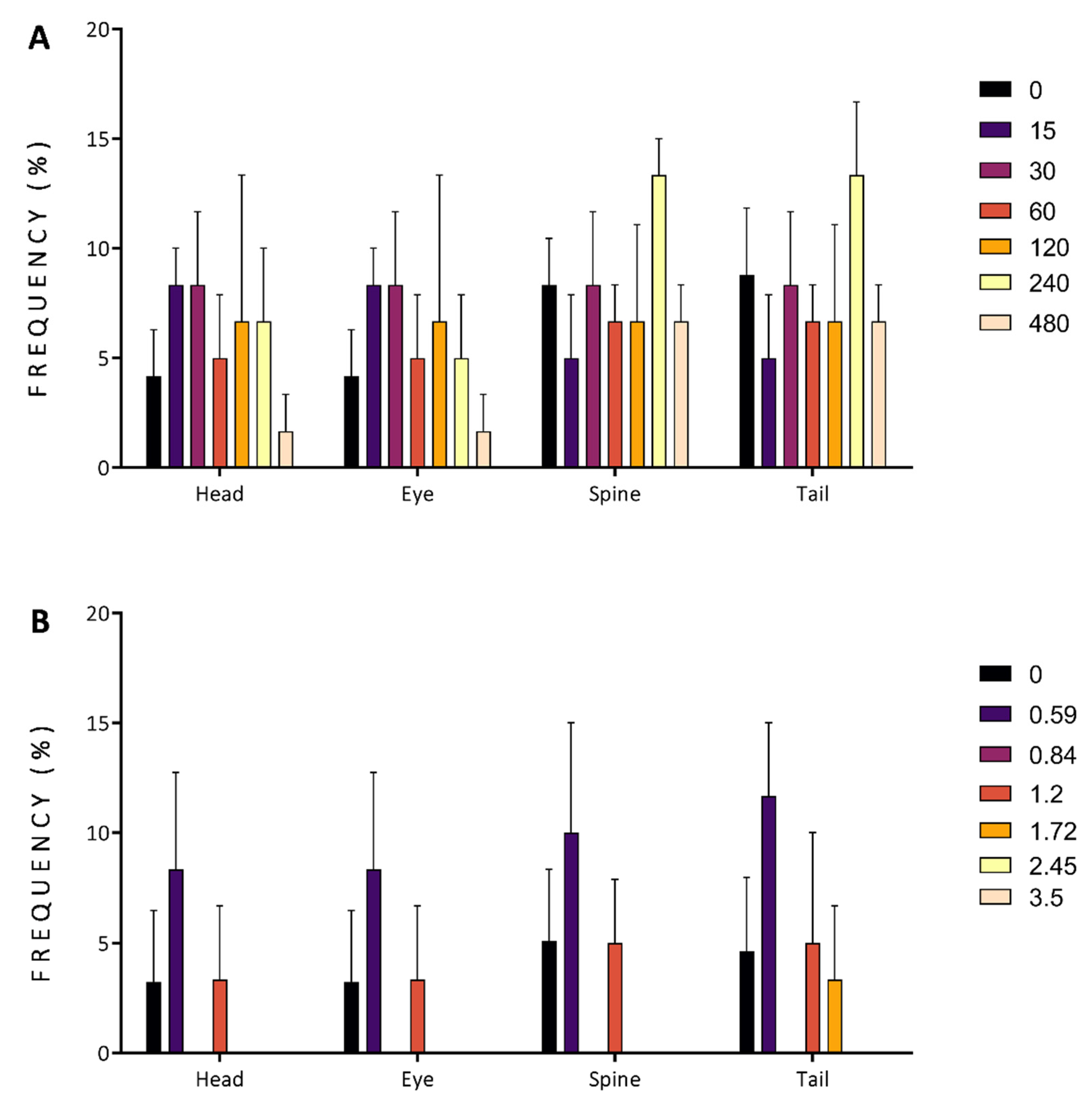
3.2. In Vivo Acute Toxicity of Australian Plants
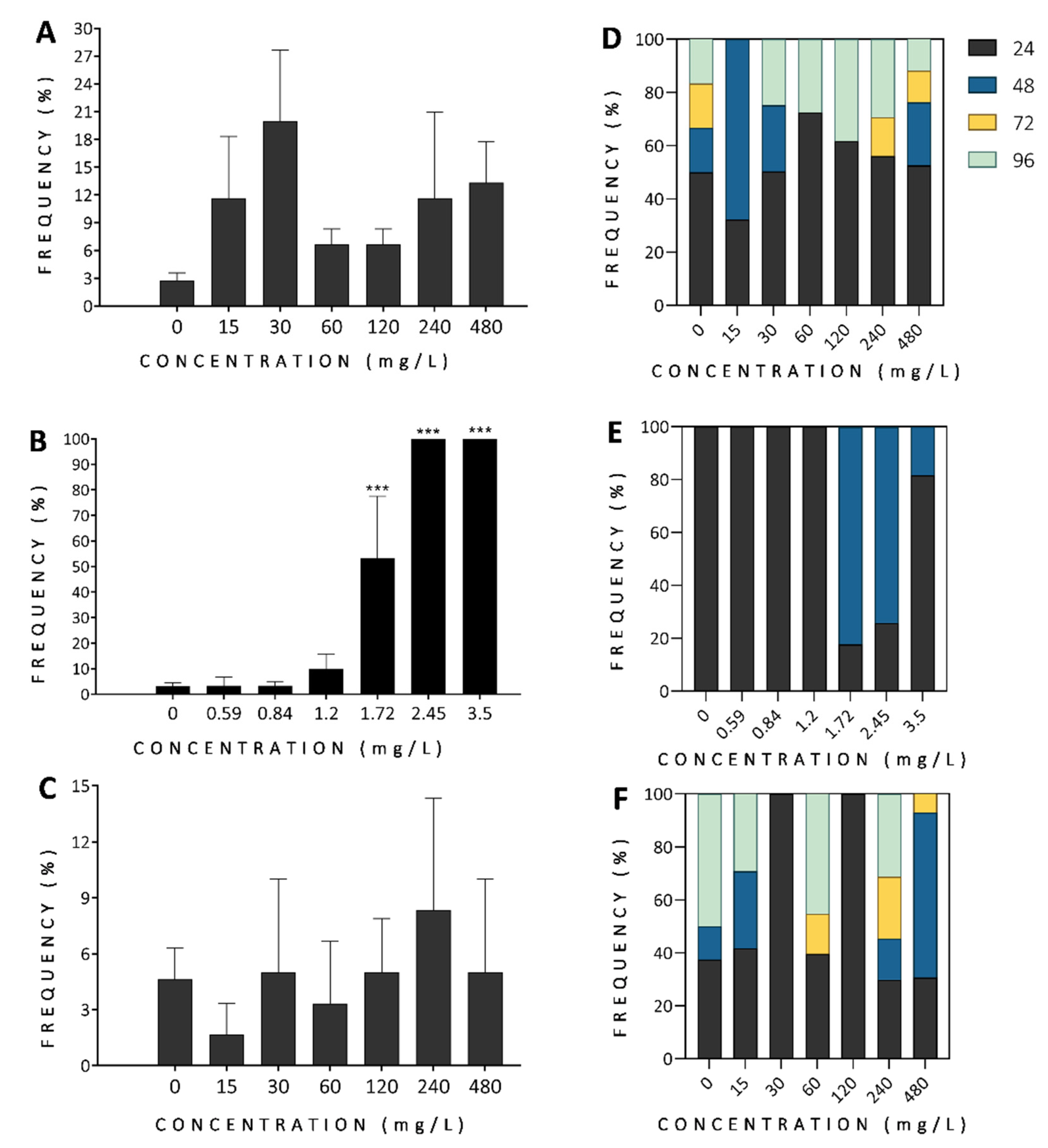
3.2.1. Mortality and Morbidity
3.2.2. Developmental Alterations
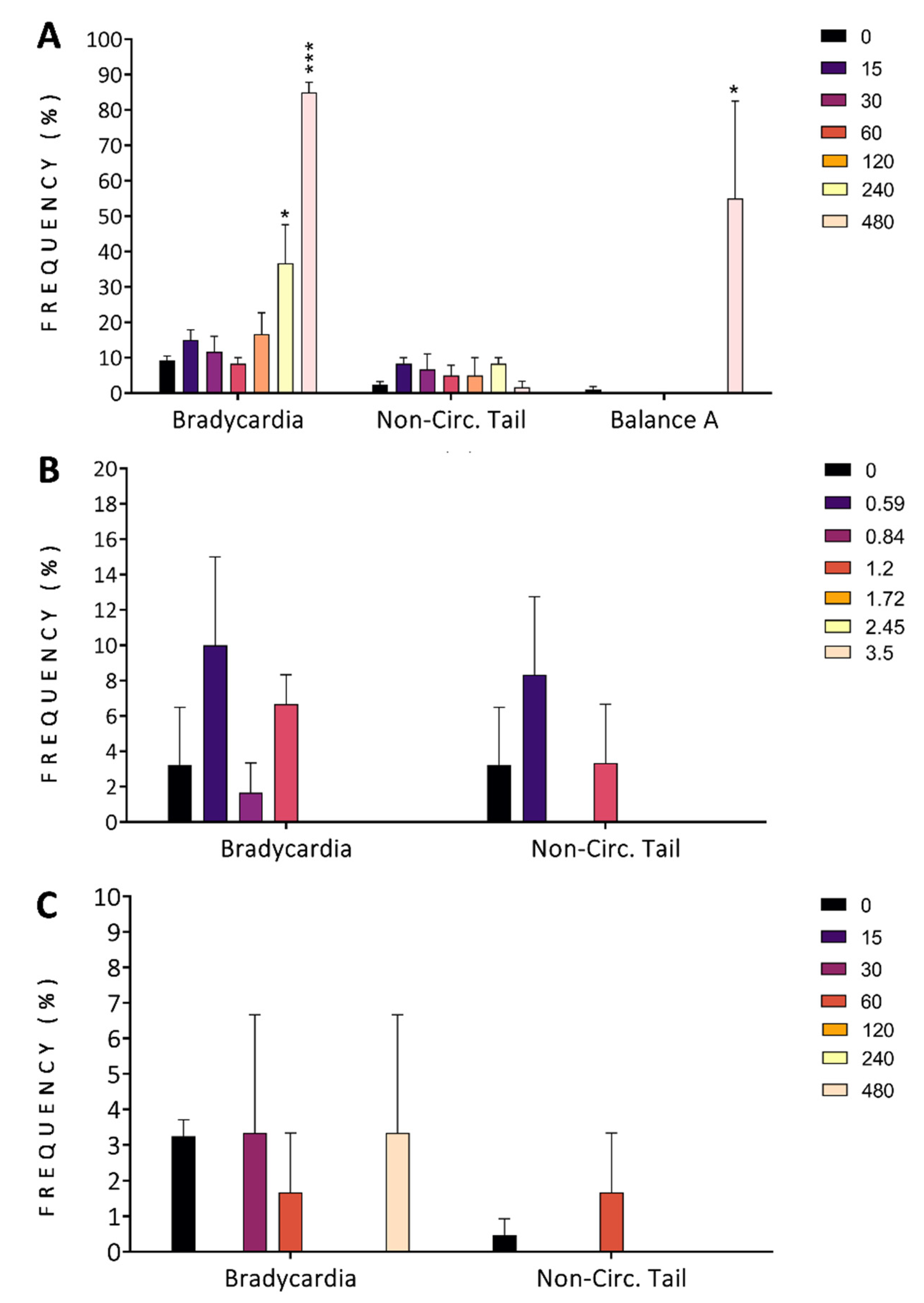
3.2.3. Malformations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Velu, G.; Palanichamy, V.; Rajan, A.P. Phytochemical and Pharmacological Importance of Plant Secondary Metabolites in Modern Medicine. In Bioorganic Phase in Natural Food: An Overview; Springer International Publishing: Cham, Switzerland, 2018; pp. 135–156. [Google Scholar]
- Hussein, R.A.; El-Anssary, A.A. Plants Secondary Metabolites: The Key Drivers of the Pharmacological Actions of Medicinal Plants. In Herbal Medicine; IntechOpen: London, UK, 2019. [Google Scholar]
- Anand, U.; Jacobo-Herrera, N.; Altemimi, A.; Lakhssassi, N. A Comprehensive Review on Medicinal Plants as Antimicrobial Therapeutics: Potential Avenues of Biocompatible Drug Discovery. Metabolites 2019, 9, 258. [Google Scholar] [CrossRef] [Green Version]
- Michael, D. New Pharmaceutical, Nutraceutical & Industrial Products: The Potential for Australian Agriculture; Wondu Holdings Pty Limited: Barton, Australia, 2000. [Google Scholar]
- Shami, A.M.; Philip, K.; Muniandy, S. Synergy of Antibacterial and Antioxidant Activities from Crude Extracts and Peptides of Selected Plant Mixture. BMC Complement. Altern. Med. 2013, 13, 360. [Google Scholar] [CrossRef] [Green Version]
- Weatherhead, J. Australian Native Food Harvest: A Guide for the Passionate Cook and Gardener; Peppermint Ridge Farm: Melbourne, Australia, 2016. [Google Scholar]
- Winnett, V.; Boyer, H.; Sirdaarta, J.; Cock, I. The Potential of Tasmannia lanceolata as a Natural Preservative and Medicinal Agent: Antimicrobial Activity and Toxicity. Pharmacogn. Commun. 2014, 4, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Cock, I.E. Medicinal and Aromatic Plants—Australia, in Ethnopharmacology Section, Biological, Physiological and Health Sciences. In Encyclopedia of Life Support Systems (EOLSS); Developed under the Auspices of the UNESCO; EOLSS Publishers: Oxford, UK, 2011; Available online: http://www.eolss.net (accessed on 8 April 2022).
- Sirdaarta, J.; Maen, A.; Rayan, P.; Matthews, B.; Cock, I. High Performance Liquid Chromatography-Mass Spectrometry Analysis of High Antioxidant Australian Fruits with Antiproliferative Activity against Cancer Cells. Pharmacogn. Mag. 2016, 12, 181. [Google Scholar]
- Lee, C.J.; Wright, M.H.; Arnold, M.S.J.; Greene, A.C.; Cock, I.E. Inhibition of Streptococcus Pyogenes Growth by Native Australian Plants: New Approaches towards the Management of Impetigo, Pharyngitis and Rheumatic Heart Disease. Pharmacogn. Commun. 2016, 6, 164–173. [Google Scholar]
- Konczak, I.; Roulle, P. Nutritional Properties of Commercially Grown Native Australian Fruits: Lipophilic Antioxidants and Minerals. Food Res. Int. 2011, 44, 2339–2344. [Google Scholar] [CrossRef]
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S.; et al. Beneficial Role of Phytochemicals on Oxidative Stress and Age-Related Diseases. Biomed Res. Int. 2019, 2019, 8748253. [Google Scholar]
- Wu, M.; Luo, Q.; Nie, R.; Yang, X.; Tang, Z.; Chen, H. Potential Implications of Polyphenols on Aging Considering Oxidative Stress, Inflammation, Autophagy, and Gut Microbiota. Crit. Rev. Food Sci. Nutr. 2021, 61, 2175–2193. [Google Scholar] [CrossRef]
- Ratnam, D.V.; Ankola, D.D.; Bhardwaj, V.; Sahana, D.K.; Kumar, M.N.V.R. Role of Antioxidants in Prophylaxis and Therapy: A Pharmaceutical Perspective. J. Control. Release 2006, 113, 189–207. [Google Scholar] [CrossRef]
- Ota, A.; Ulrih, N.P. An Overview of Herbal Products and Secondary Metabolites Used for Management of Type Two Diabetes. Front. Pharmacol. 2017, 8, 436. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative Stress, Aging, and Diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Cáceres-Vélez, P.R.; Hui, F.; Hercus, J.; Bui, B.; Jusuf, P.R. Restoring the Oxidative Balance in Age-Related Diseases—An Approach in Glaucoma. Ageing Res. Rev. 2022, 75, 101572. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [Green Version]
- Golpich, M.; Amini, E.; Mohamed, Z.; Ali, R.A.; Ibrahim, N.M. Mitochondrial Dysfunction and Biogenesis in Neurodegenerative Diseases: Pathogenesis and Treatment. Neurosci. Ther. 2017, 23, 5–22. [Google Scholar] [CrossRef]
- Wilkinson-Berka, J.L.; Pattamatta, U.; Ung, L.; White, A.J.R.; Liew, G.; Carnt, N. Oxidative Stress and Reactive Oxygen Species: A Review of Their Role in Ocular Disease. Clin. Sci. 2017, 131, 2865–2883. [Google Scholar]
- Madreiter-Sokolowski, C.T.; Thomas, C.; Ristow, M. Interrelation between ROS and Ca2+ in Aging and Age-Related Diseases. Redox Biol. 2020, 36, 101678. [Google Scholar] [CrossRef]
- Rupesinghe, E.J.R.; Jones, A.; Shalliker, R.A.; Pravadali-Cekic, S. A Rapid Screening Analysis of Antioxidant Compounds in Native Australian Food Plants Using Multiplexed Detection with Active Flow Technology Columns. Molecules 2016, 21, 118. [Google Scholar] [CrossRef] [Green Version]
- Aldosary, H.; Wright, M.H.; Lee, C.J.; Greene, A.C.; Cock, I.E. The Growth Inhibitory Activity of Tasmannia lanceolata (Poir.) A.C. Sm against the Food-Poisoning Pathogen Yersinia enterocolitica. Pharmacogn. Commun. 2019, 9, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Parasuraman, S. Toxicological Screening. J. Pharmacol. Pharmacother. 2011, 2, 74. [Google Scholar]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a Model Vertebrate for Investigating Chemical Toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [Green Version]
- OECD. Test No. 236: Fish Embryo Acute Toxicity (FET) Test; OECD: Paris, France, 2013. [Google Scholar]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of Zebrafish in Drug Discovery Toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Pyati, U.J.; Look, A.T.; Hammerschmidt, M. Zebrafish as a Powerful Vertebrate Model System for in Vivo Studies of Cell Death. Semin. Cancer Biol. 2007, 17, 154–165. [Google Scholar] [CrossRef]
- Ali, A.; Wu, H.; Ponnampalam, E.N.; Cottrell, J.J.; Dunshea, F.R.; Suleria, H.A.R. Comprehensive Profiling of Most Widely Used Spices for Their Phenolic Compounds through LC-ESI-QTOF-MS 2 and Their Antioxidant Potential. Antioxidants 2021, 10, 721. [Google Scholar]
- Chou, O.; Ali, A.; Subbiah, V.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF-MS/MS Characterisation of Phenolics in Herbal Tea Infusion and Their Antioxidant Potential. Fermentation 2021, 7, 73. [Google Scholar]
- Bashmil, Y.M.; Ali, A.; Bk, A.; Dunshea, F.R. Screening and Characterization of Phenolic Compounds from Australian Grown Bananas and Their Antioxidant Capacity. Antioxidants 2021, 10, 1521. [Google Scholar] [CrossRef]
- Ali, A.; Bashmil, Y.M.; Cottrell, J.J.; Suleria, H.A.R.; Dunshea, F.R. LC-MS/MS-QTOF Screening and Identification of Phenolic Compounds from Australian Grown Herbs and Their Antioxidant Potential. Antioxidants 2021, 10, 1770. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Song, S.; Ali, A.; Subbiah, V.; Taheri, Y.; Suleria, H.A.R. LC-ESI-QTOF-MS/MS Characterization of Phenolic Compounds from Pyracantha coccinea M.Roem. and Their Antioxidant Capacity. Cell. Mol. Biol. 2021, 67, 201–211. [Google Scholar]
- Cáceres-Vélez, P.R.; Fascineli, M.L.; Sousa, M.H.; Grisolia, C.K.; Yate, L.; de Souza, P.E.N.; Estrela-Lopis, I.; Moya, S.; Azevedo, R.B. Humic Acid Attenuation of Silver Nanoparticle Toxicity by Ion Complexation and the Formation of a Ag3+ Coating. J. Hazard. Mater. 2018, 353, 173–181. [Google Scholar] [CrossRef]
- Konczak, I.; Zabaras, D.; Dunstan, M.; Aguas, P. Antioxidant Capacity and Hydrophilic Phytochemicals in Commercially Grown Native Australian Fruits. Food Chem. 2010, 123, 1048–1054. [Google Scholar] [CrossRef]
- Sommano, S.; Caffin, N.; Kerven, G. Screening for Antioxidant Activity, Phenolic Content, and Flavonoids from Australian Native Food Plants. Int. J. Food Prop. 2013, 16, 1394–1406. [Google Scholar] [CrossRef] [Green Version]
- Shlini, P.; Siddalinga, M.K.R. Extraction of Phenolics, Proteins and Antioxidant Activity from Defatted Tamarind Kernel Powder. Asian J. Res. Chem. 2011, 4, 936–941. [Google Scholar]
- Subbiah, V.; Zhong, B.; Nawaz, M.A.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. Screening of Phenolic Compounds in Australian Grown Berries by LC-ESI-QTOF-MS/MS and Determination of Their Antioxidant Potential. Antioxidants 2020, 10, 26. [Google Scholar] [CrossRef]
- Sandeep Kumar, A.; Yadav, M.Y.; Yadav, J.P. Effect of Climate Change on Phytochemical Diversity, Total Phenolic Content and in Vitro Antioxidant Activity of Aloe vera (L.) Burm.F. BMC Res. Notes 2017, 10, 60. [Google Scholar]
- Selvamuthukumaran, M.; Shi, J. Recent Advances in Extraction of Antioxidants from Plant By-Products Processing Industries. Food Qual. Saf. 2017, 1, 61–81. [Google Scholar] [CrossRef]
- Robbins, R.J. Phenolic Acids in Foods: An Overview of Analytical Methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef]
- El-Haddad, A.E.-S.; Saadeldeen, A.M.; El-Emam, S.Z. Anti-Angiogenic Activity of Major Phenolics in Tamarind Assessed with Molecular Docking Study on VEGF Kinase Proteins. Pak. J. Biol. Sci. 2019, 22, 502–509. [Google Scholar] [CrossRef] [Green Version]
- Namazi, N.; Khodamoradi, K.; Khamechi, S.P.; Heshmati, J.; Ayati, M.H.; Larijani, B. The Impact of Cinnamon on Anthropometric Indices and Glycemic Status in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Clinical Trials. Complement. Ther. Med. 2019, 43, 92–101. [Google Scholar] [CrossRef]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant Properties of Ferulic Acid and Its Possible Application. Skin Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Sudjaroen, Y.; Haubner, R.; Würtele, G.; Hull, W.E.; Erben, G.; Spiegelhalder, B.; Changbumrung, S.; Bartsch, H.; Owen, R.W. Isolation and Structure Elucidation of Phenolic Antioxidants from Tamarind (Tamarindus indica L.) Seeds and Pericarp. Food Chem. Toxicol. 2005, 43, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Daussin, F.N.; Heyman, E.; Burelle, Y. Effects of (−)-Epicatechin on Mitochondria. Nutr. Rev. 2021, 79, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a Flavonoid, as an Anticancer Agent: A Review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, H.; Xie, K.; Yin, L.; Li, Y.; Kwik-Uribe, C.L.; Zhu, X. Procyanidin Dimer B2 [Epicatechin-(4β-8)-Epicatechin] Suppresses the Expression of Cyclooxygenase-2 in Endotoxin-Treated Monocytic Cells. Biochem. Biophys. Res. Commun. 2006, 345, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.-X.; Wang, S.; Wei, L.; Cui, Y.-Y.; Chen, Y.-H. Proanthocyanidins: Components, Pharmacokinetics and Biomedical Properties. Am. J. Chin. Med. 2020, 48, 813–869. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-Ovando, A.; Pacheco-Hernández, M.D.L.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical Studies of Anthocyanins: A Review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Chen, L.; Qiang, P.; Xu, Q.; Zhao, Y.; Dai, F.; Zhang, L. Trans-3,4,5,4′-Tetramethoxystilbene, a Resveratrol Analog, Potently Inhibits Angiogenesis in Vitro and in Vivo. Acta Pharmacol. Sin. 2013, 34, 1174–1182. [Google Scholar] [CrossRef] [Green Version]
- Lo, J.Y.; Kamarudin, M.N.A.; Hamdi, O.A.A.; Awang, K.; Kadir, H.A. Curcumenol Isolated from Curcuma Zedoaria Suppresses Akt-Mediated NF-ΚB Activation and P38 MAPK Signaling Pathway in LPS-Stimulated BV-2 Microglial Cells. Food Funct. 2015, 6, 3550–3559. [Google Scholar] [CrossRef]
- Detsi, A.; Kontogiorgis, C.; Hadjipavlou-Litina, D. Coumarin Derivatives: An Updated Patent Review (2015–2016). Expert Opin. Ther. Pat. 2017, 27, 1201–1226. [Google Scholar] [CrossRef]
- Bhattarai, N.; Kumbhar, A.A.; Pokharel, Y.R.; Yadav, P.N. Anticancer Potential of Coumarin and Its Derivatives. Mini-Rev. Med. Chem. 2021, 21, 2996–3029. [Google Scholar] [CrossRef]
- Sakulnarmrat, K.; Konczak, I. Composition of Native Australian Herbs Polyphenolic-Rich Fractions and in Vitro Inhibitory Activities against Key Enzymes Relevant to Metabolic Syndrome. Food Chem. 2012, 134, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.H.; Lee, C.J.; Sarah, M.; Arnold, J.; Shalom, J.; White, A.; Greene, A.C.; Cock, I.E.; Wright, M.H.; Lee, C.J.; et al. GC-MS Analysis of Tasmannia lanceolata Extracts Which Inhibit the Growth of the Pathogenic Bacterium Clostridium Perfringens. Pharmacogn. J. 2017, 9, 626–637. [Google Scholar] [CrossRef] [Green Version]
- Cock, I. The Phytochemistry and Chemotherapeutic Potential of Tasmannia lanceolata (Tasmanian Pepper): A Review. Pharmacogn. Commun. 2013, 3, 13–25. [Google Scholar]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, A.; Sudwarts, A.; Parker, M.O.; Brennan, C.H. Zebrafish Behavioral Models of Ageing. In The Rights and Wrongs of Zebrafish: Behavioral Phenotyping of Zebrafish; Springer: Cham, Switzerland, 2017; pp. 241–258. [Google Scholar]
- Rayan, P.; Matthews, B.; Donnell, P.M.; Cock, I.E. Phytochemical Analysis of Tasmannia lanceolata Extracts and Inhibition of Giardia Duodenalis Proliferation. Pharmacogn. J. 2016, 8, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, J.; Cock, I.E. Acai, Cacao and Maca Extracts: Anticancer Activity and Growth Inhibition of Microbial Triggers of Selected Autoimmune Inflammatory Diseases. Pharmacogn. Commun. 2016, 6, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Singh, C.; Upadhyay, R.; Nath, K. Comparative Analysis of the Seasonal Influence on Polyphenolic Content, Antioxidant Capacity, Identification of Bioactive Constituents and Hepatoprotective Biomarkers by in Silico Docking Analysis in Premna integrifolia L. Physiol. Mol. Biol. Plants 2022, 28, 223–249. [Google Scholar] [CrossRef]
- Schulte-Hubbert, R.; Küpper, J.-H.; Thomas, A.D.; Schrenk, D. Estragole: DNA Adduct Formation in Primary Rat Hepatocytes and Genotoxic Potential in HepG2-CYP1A2 Cells. Toxicology 2020, 444, 152566. [Google Scholar] [CrossRef]
- Kirsch, V.; Bakuradze, T.; Richling, E. Toxicological Testing of Syringaresinol and Enterolignans. Curr. Res. Toxicol. 2020, 1, 104–110. [Google Scholar] [CrossRef]
- Bezerra, A.H.; Bezerra, S.R.; Macêdo, N.S.; de Sousa Silveira, Z.; dos Santos Barbosa, C.R.; de Freitas, T.S.; Muniz, D.F.; de Sousa Júnior, D.L.; Júnior, J.P.S.; Donato, I.A.; et al. Effect of Estragole over the RN4220 Staphylococcus aureus Strain and Its Toxicity in Drosophila Melanogaster. Life Sci. 2021, 264, 118675. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | TPC (mg GAE/g) | DPPH (mg TE/g) | ABTS (mg TE/g) | RPA (mg TE/g) |
|---|---|---|---|---|
| Finger lime | 0.71 ± 0.00 c | 0.44 ± 0.06 b | 0.92 ± 0.05 b | 0.69 ± 0.04 c |
| Mountain pepper | 5.91 ± 0.32 a | 4.48 ± 0.03 a | 6.68 ± 0.54 a | 13.23 ± 0.17 a |
| Tamarind | 3.72 ± 0.12 b | 4.07 ± 0.00 a | 6.86 ± 0.35 a | 12.52 ± 0.46 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cáceres-Vélez, P.R.; Ali, A.; Fournier-Level, A.; Dunshea, F.R.; Jusuf, P.R. Phytochemical and Safety Evaluations of Finger Lime, Mountain Pepper, and Tamarind in Zebrafish Embryos. Antioxidants 2022, 11, 1280. https://doi.org/10.3390/antiox11071280
Cáceres-Vélez PR, Ali A, Fournier-Level A, Dunshea FR, Jusuf PR. Phytochemical and Safety Evaluations of Finger Lime, Mountain Pepper, and Tamarind in Zebrafish Embryos. Antioxidants. 2022; 11(7):1280. https://doi.org/10.3390/antiox11071280
Chicago/Turabian StyleCáceres-Vélez, Paolin Rocio, Akhtar Ali, Alexandre Fournier-Level, Frank R. Dunshea, and Patricia Regina Jusuf. 2022. "Phytochemical and Safety Evaluations of Finger Lime, Mountain Pepper, and Tamarind in Zebrafish Embryos" Antioxidants 11, no. 7: 1280. https://doi.org/10.3390/antiox11071280
APA StyleCáceres-Vélez, P. R., Ali, A., Fournier-Level, A., Dunshea, F. R., & Jusuf, P. R. (2022). Phytochemical and Safety Evaluations of Finger Lime, Mountain Pepper, and Tamarind in Zebrafish Embryos. Antioxidants, 11(7), 1280. https://doi.org/10.3390/antiox11071280