
Limited Heme Oxygenase Contribution to Modulating the Severity of Salmonella enterica serovar Typhimurium Infection

 ,
,  , , , , ,
, , , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. In Vivo Infection and Monitoring
2.3. Bacterial Strain and Infective Dose Preparation
2.4. CoPP and SnPP Preparation
2.5. Enrofloxacin Administration
2.6. Bacterial Loads in Organs and Tissues
2.7. RNA Extraction
2.8. Quantitative Real-Time PCR
2.9. Western Blot
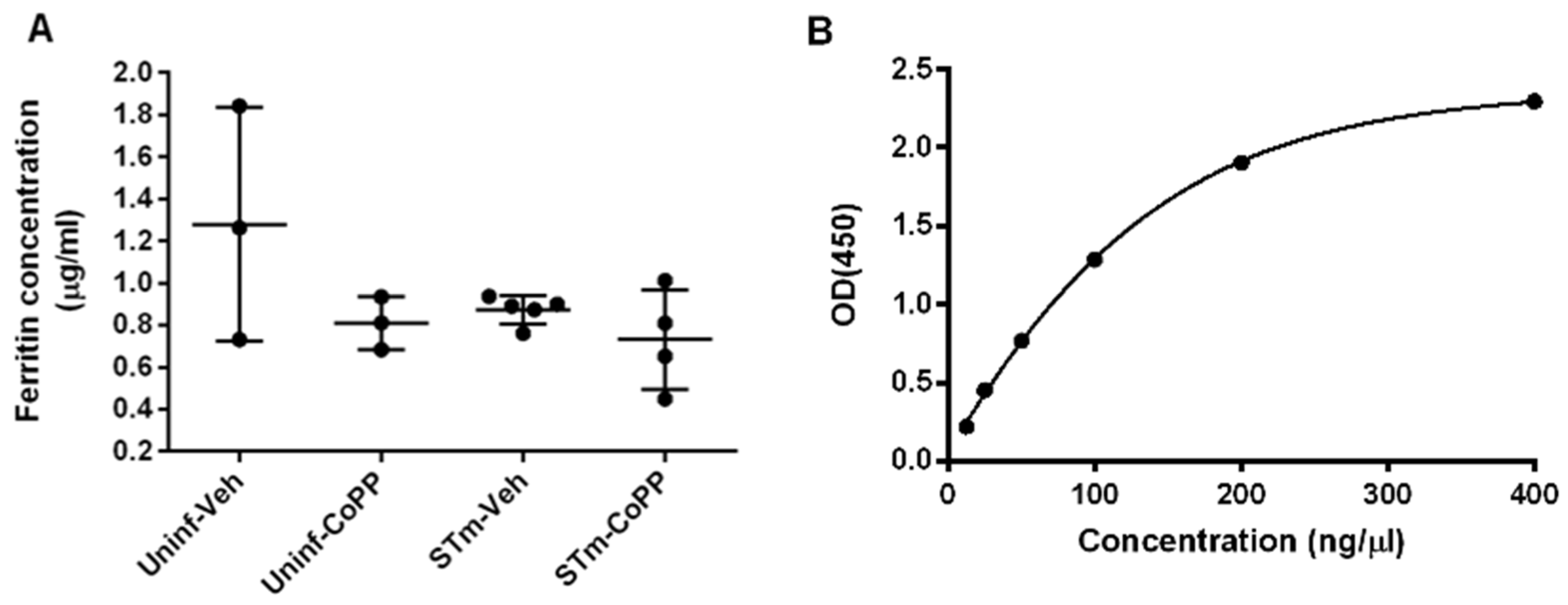
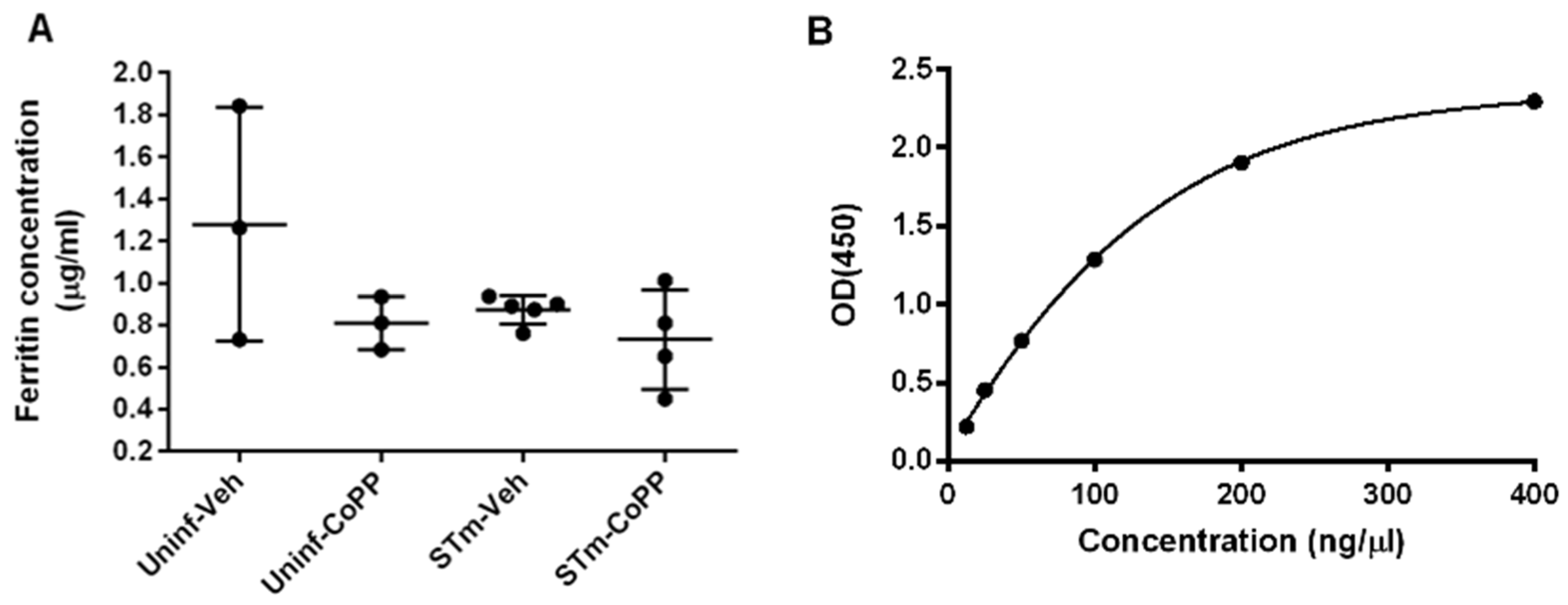
2.10. Ferritin ELISA
2.11. Cell Culture, Treatment and Infection
2.12. Flow Cytometry
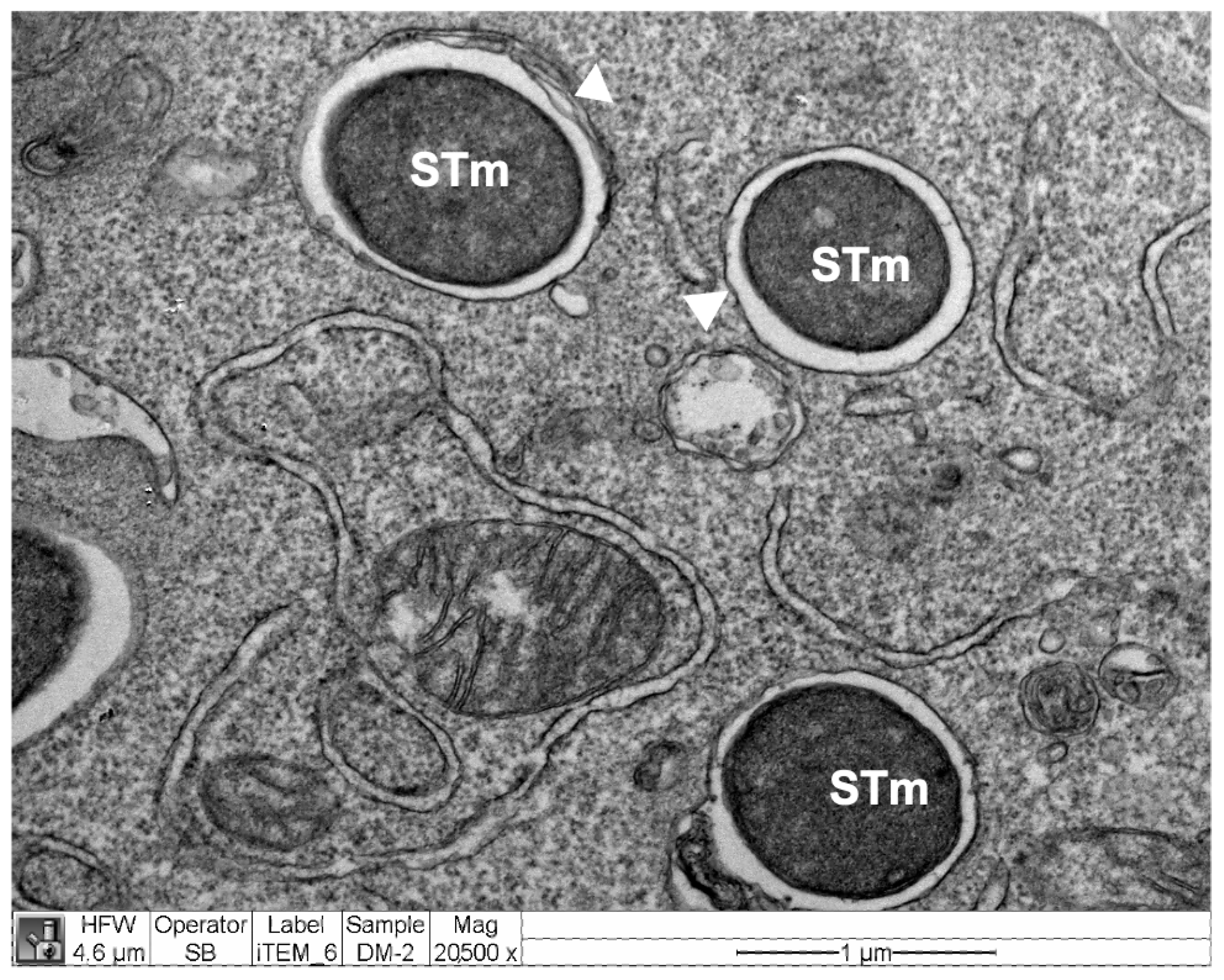
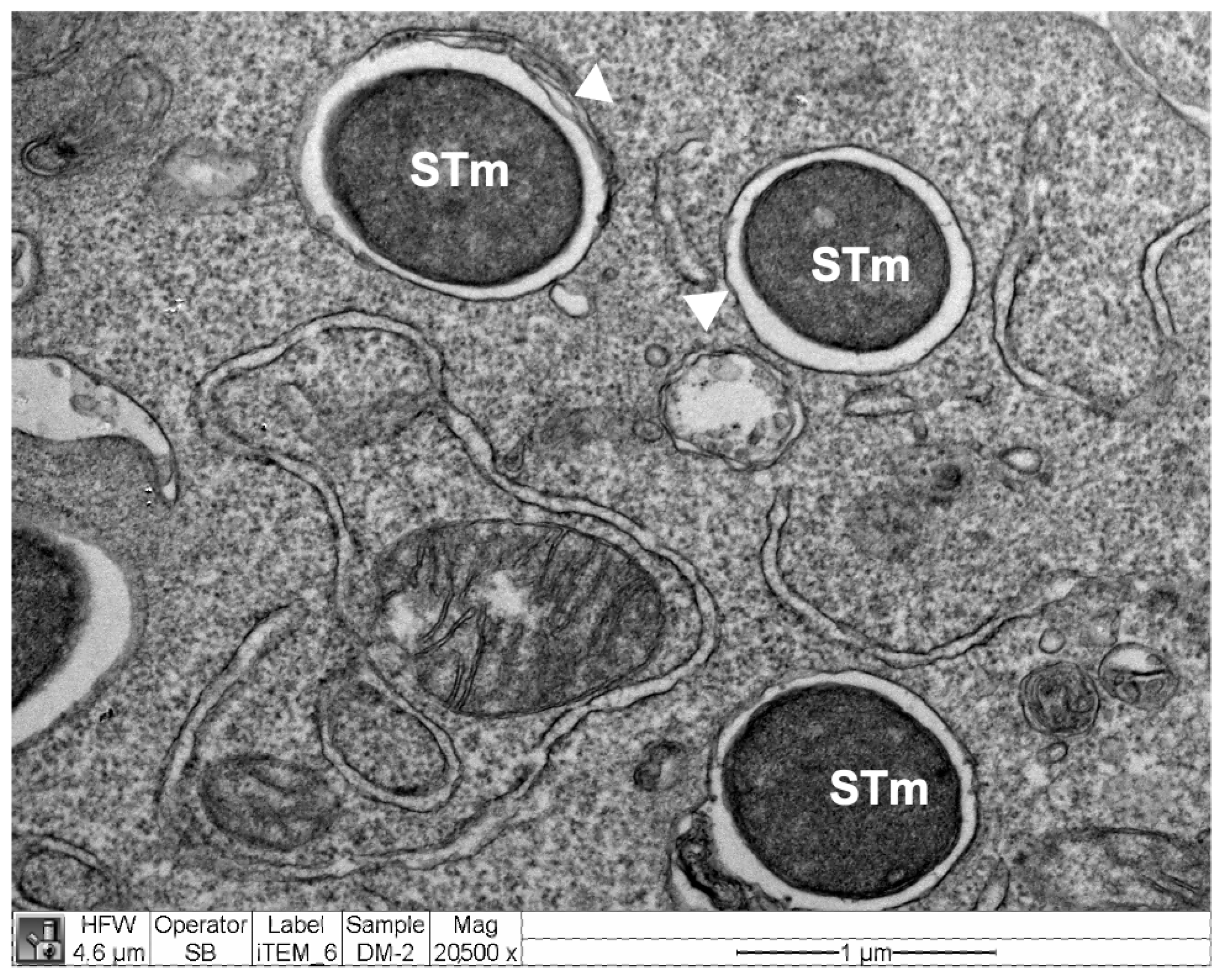
2.13. Electron Microscopy
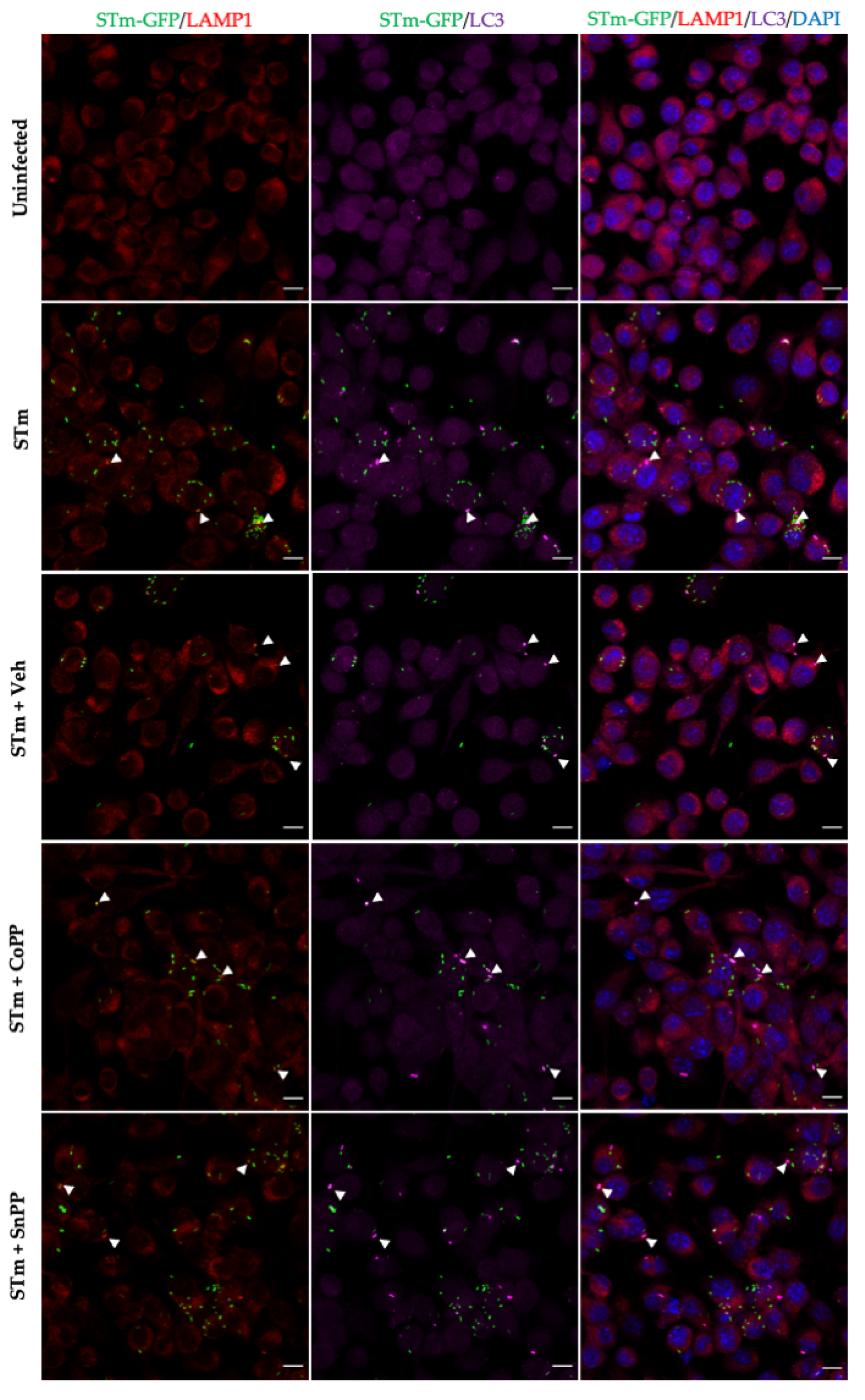
2.14. Immunofluorescence and Fluorescence Microscopy
2.15. Statistical Analysis
3. Results
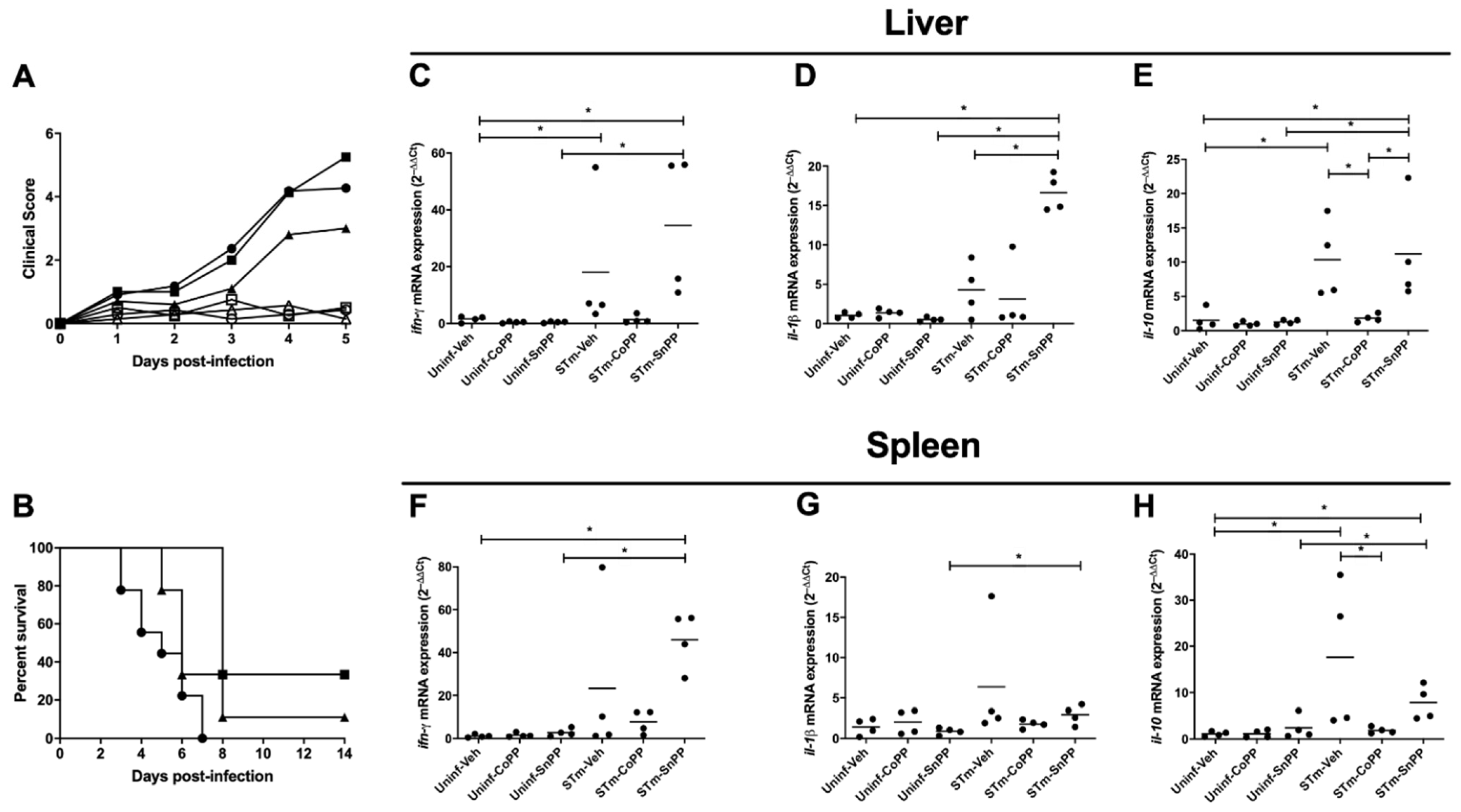
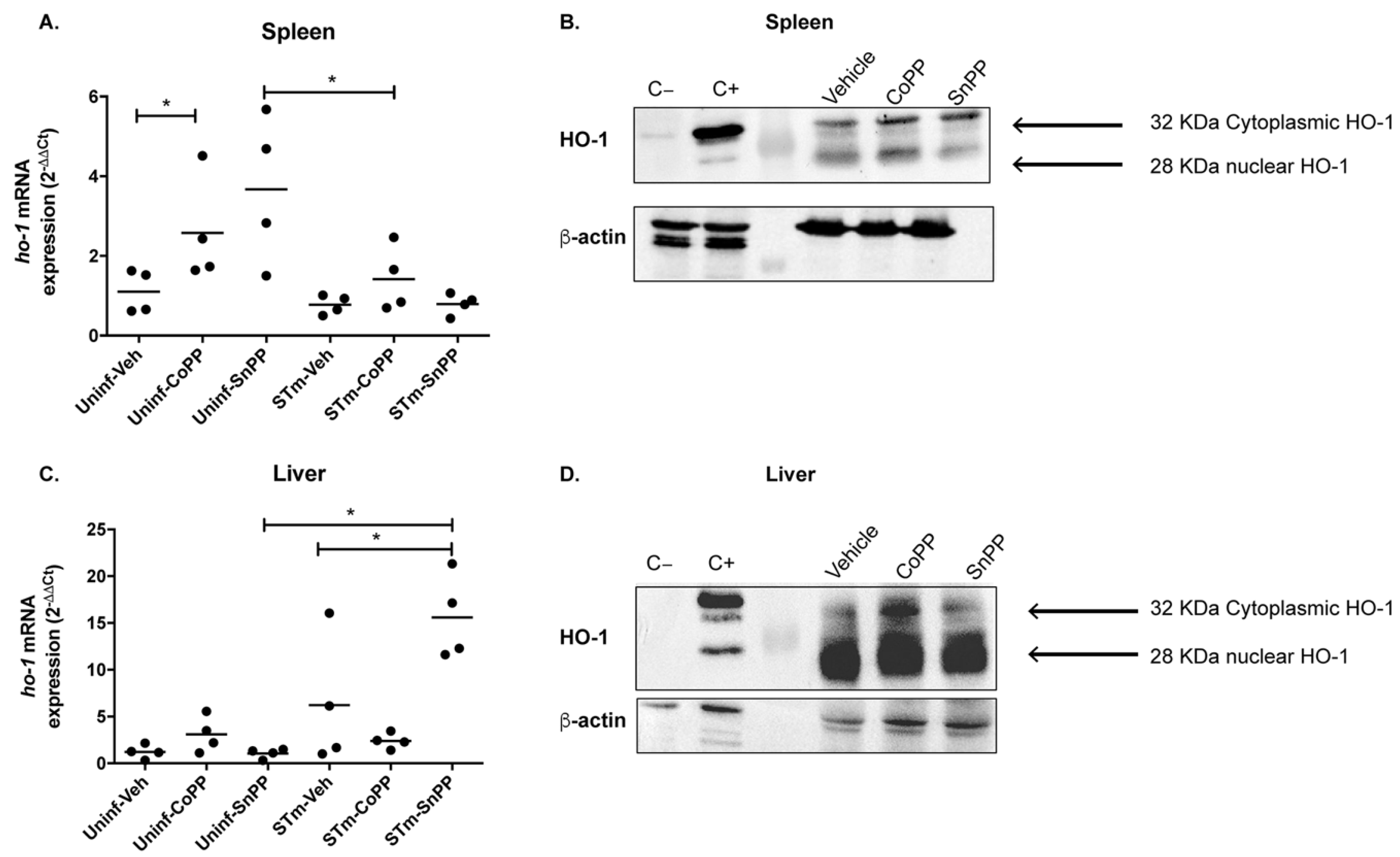
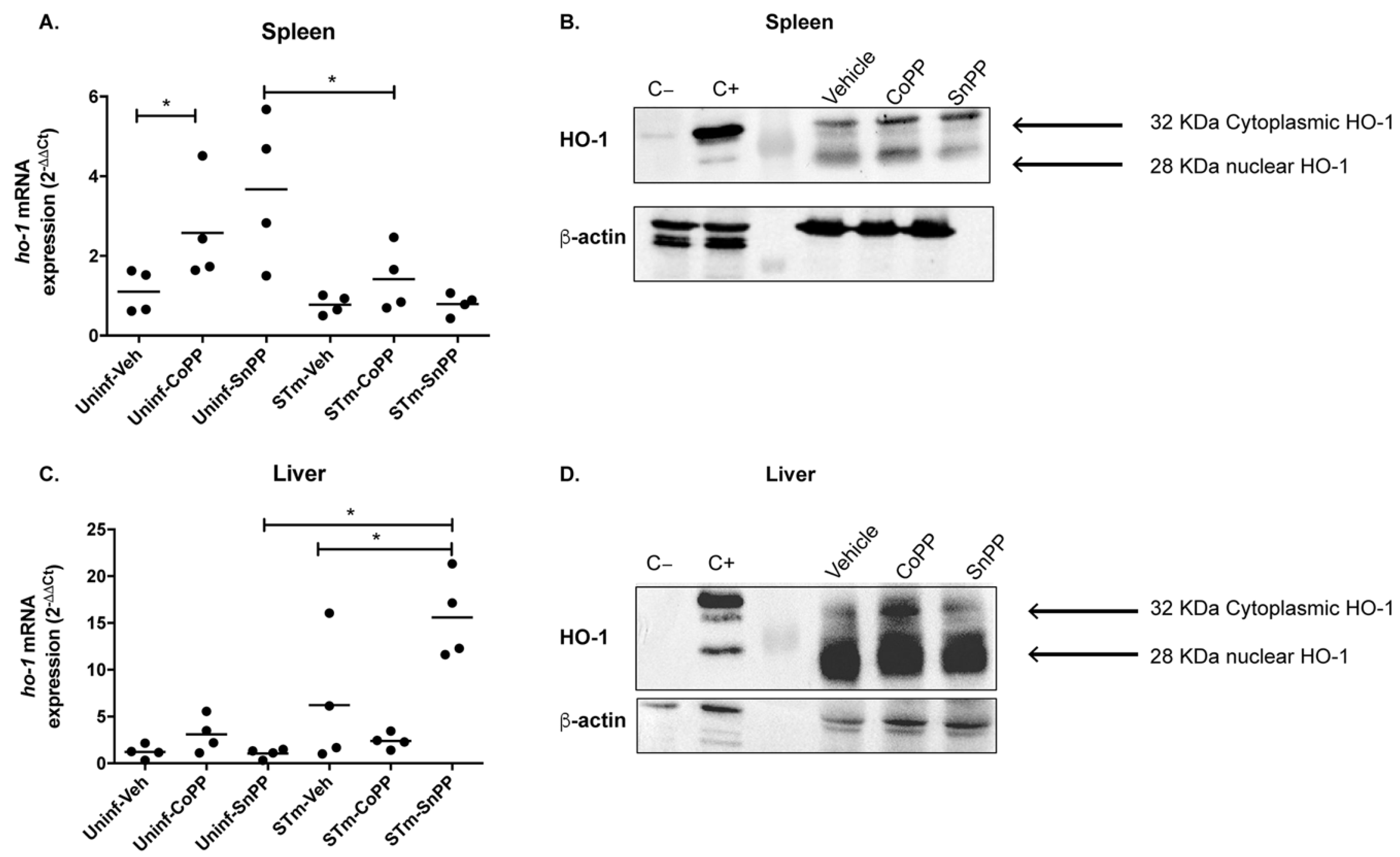
3.1. CoPP-Mediated Hmox1 Induction Does Not Influence an Acute S. Typhimurium Infection
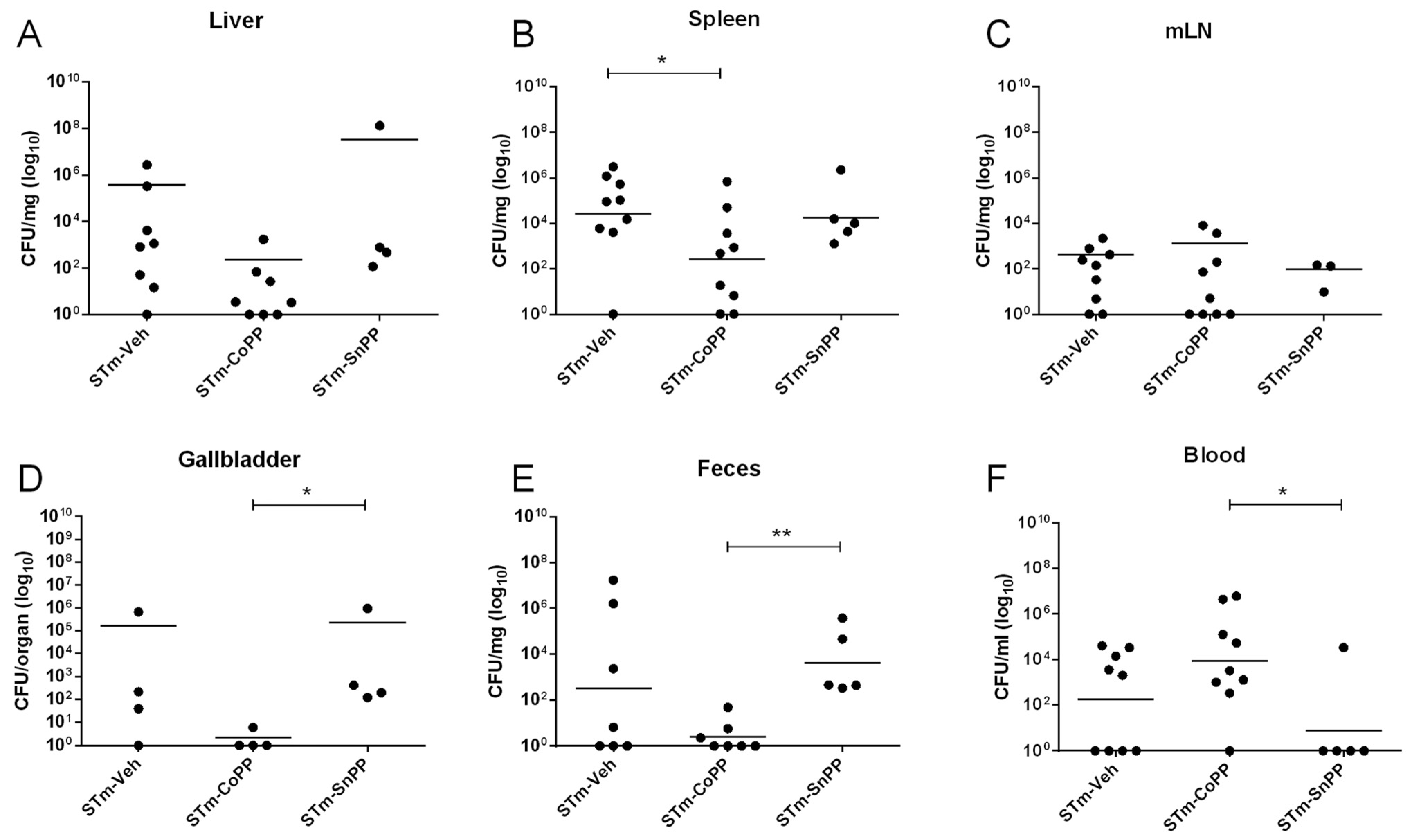
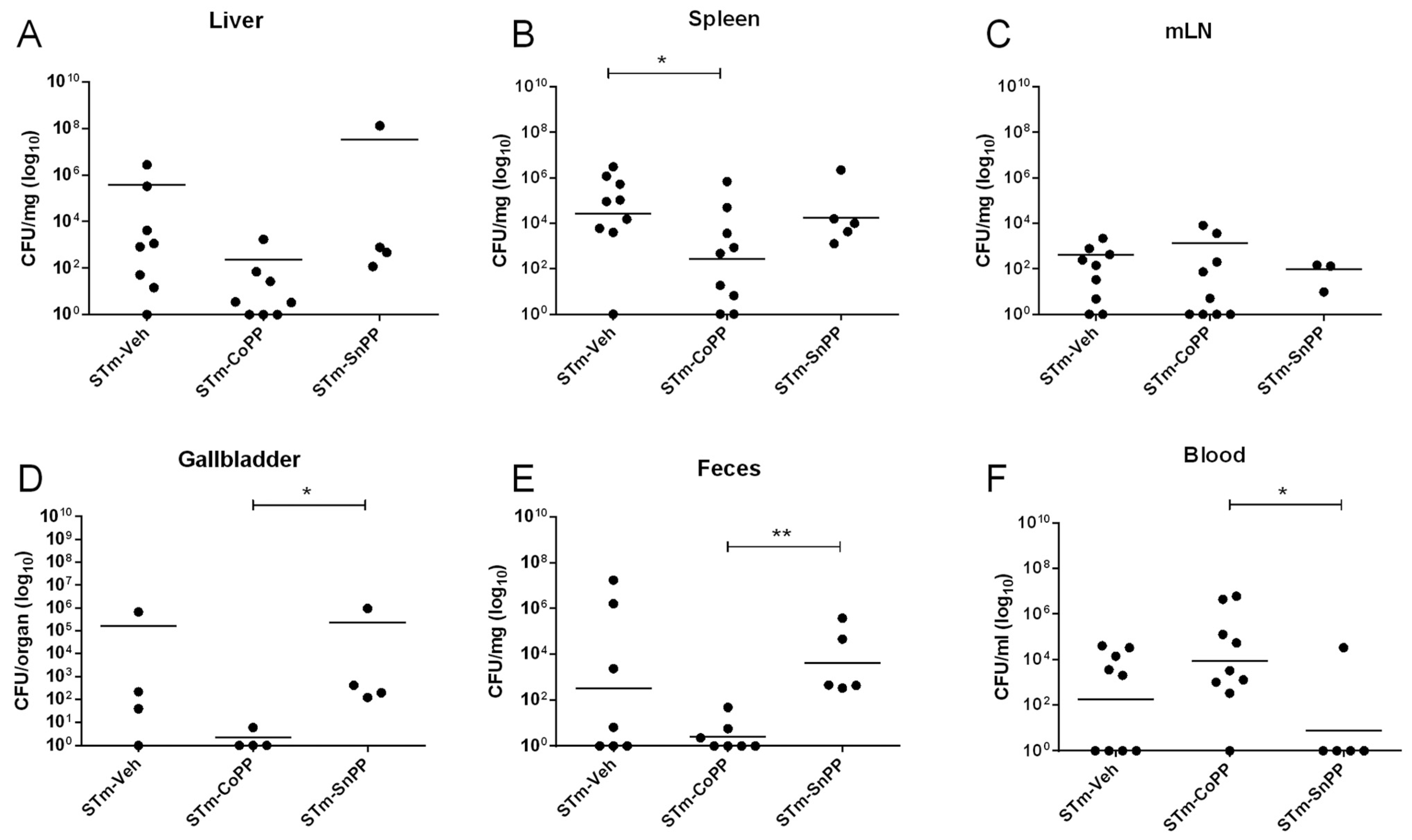
3.2. Hmox1 Induction by CoPP Treatment Reduces Bacteria Load in Tissues but Not in the Blood of S. Typhimurium-Infected Mice
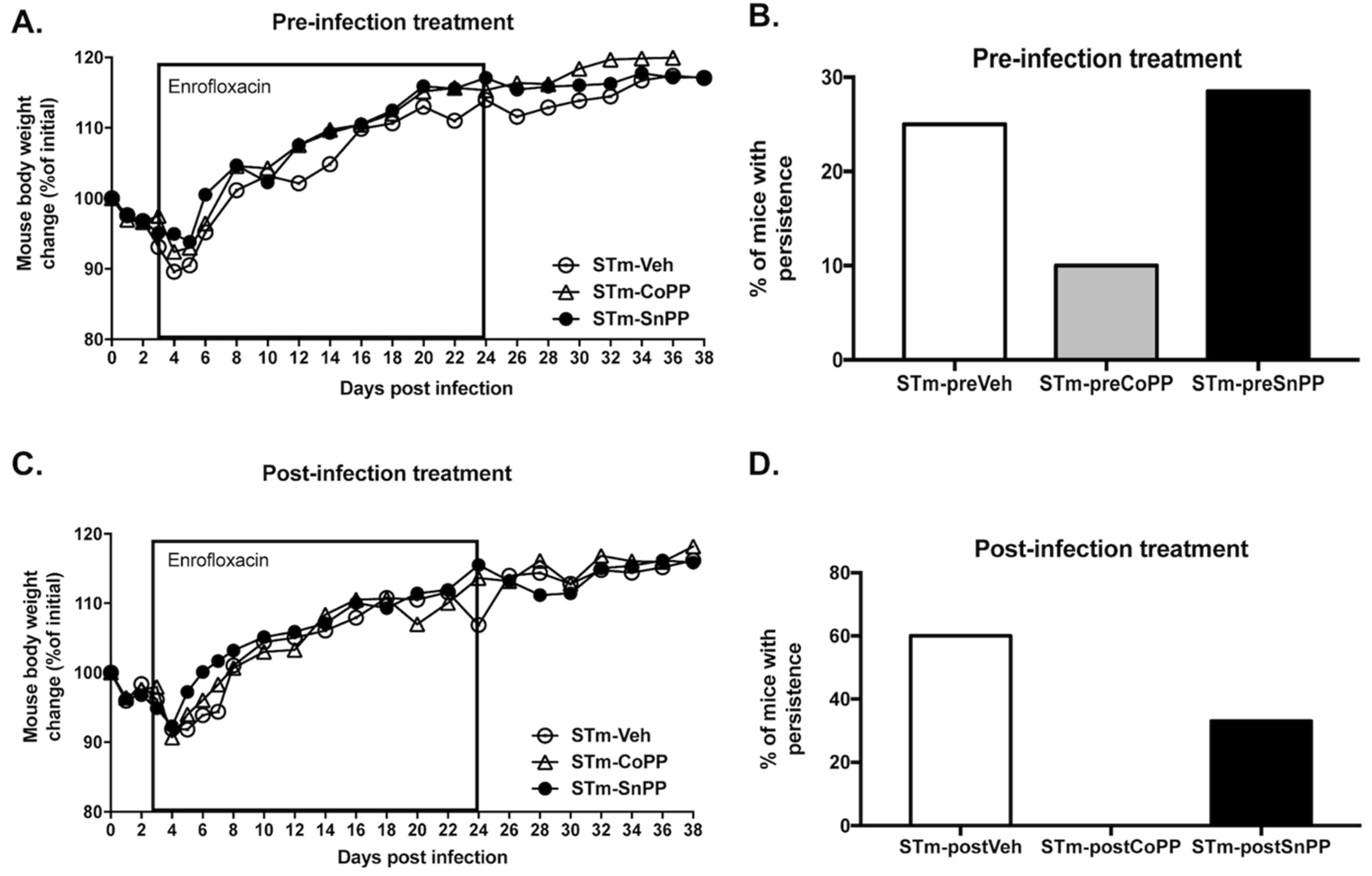
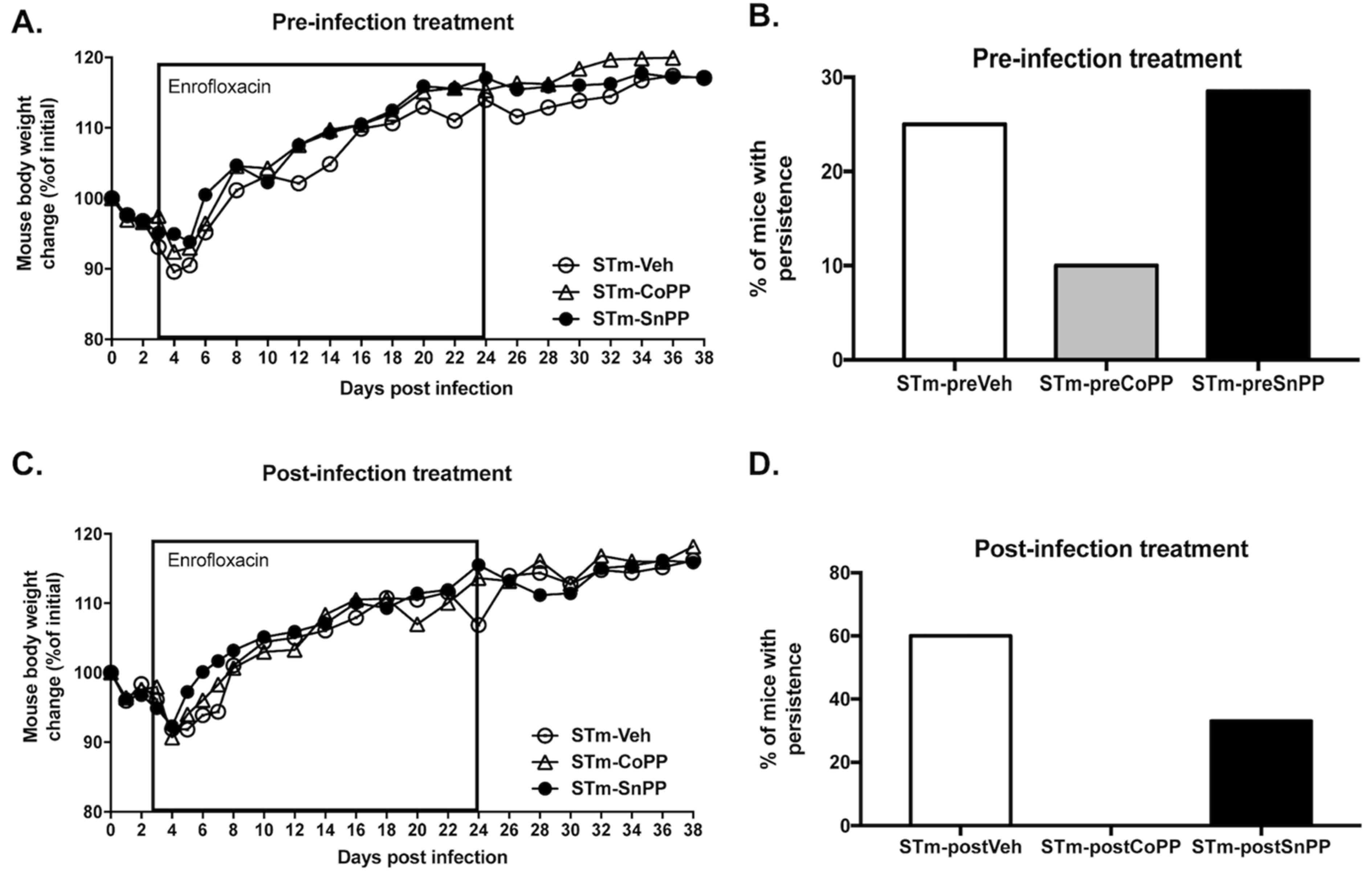
3.3. Prophylactic Hmox1 Induction by CoPP Reduces the Persistence of S. Typhimurium in Mice
3.4. Hmox1 Induction by CoPP Treatment Reduces the Persistence of S. Typhimurium When Administrated Post-Infection
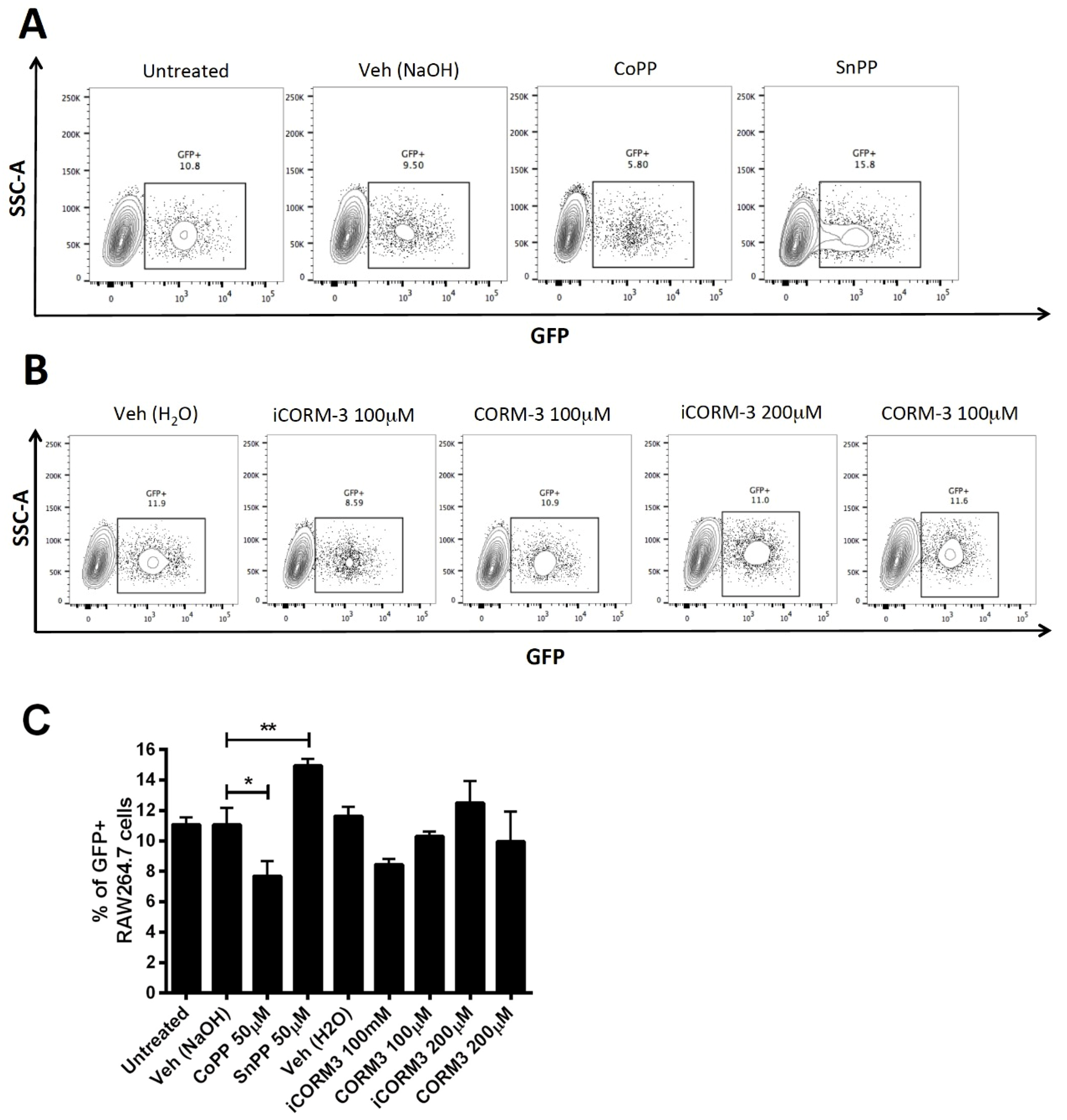
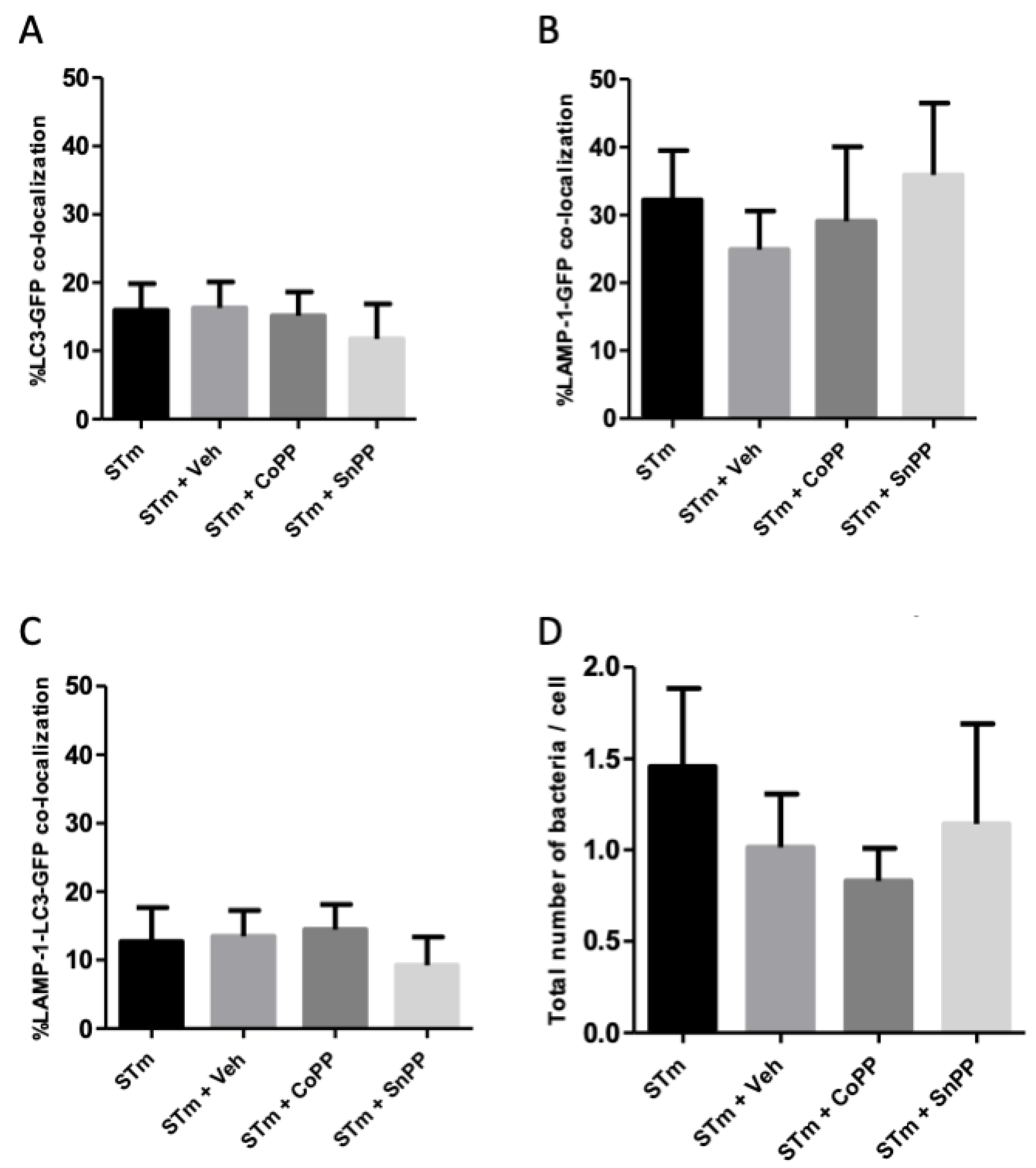
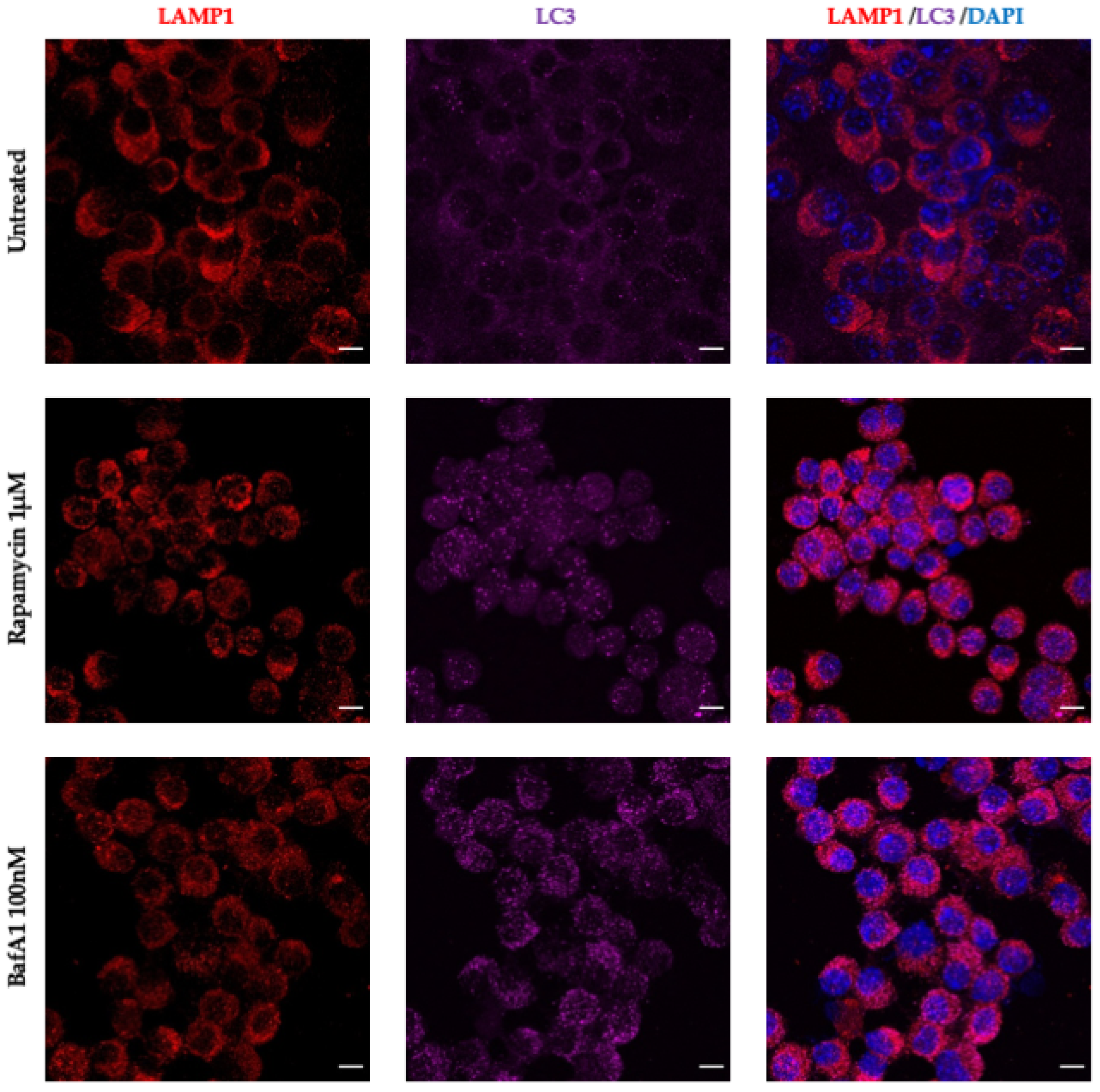
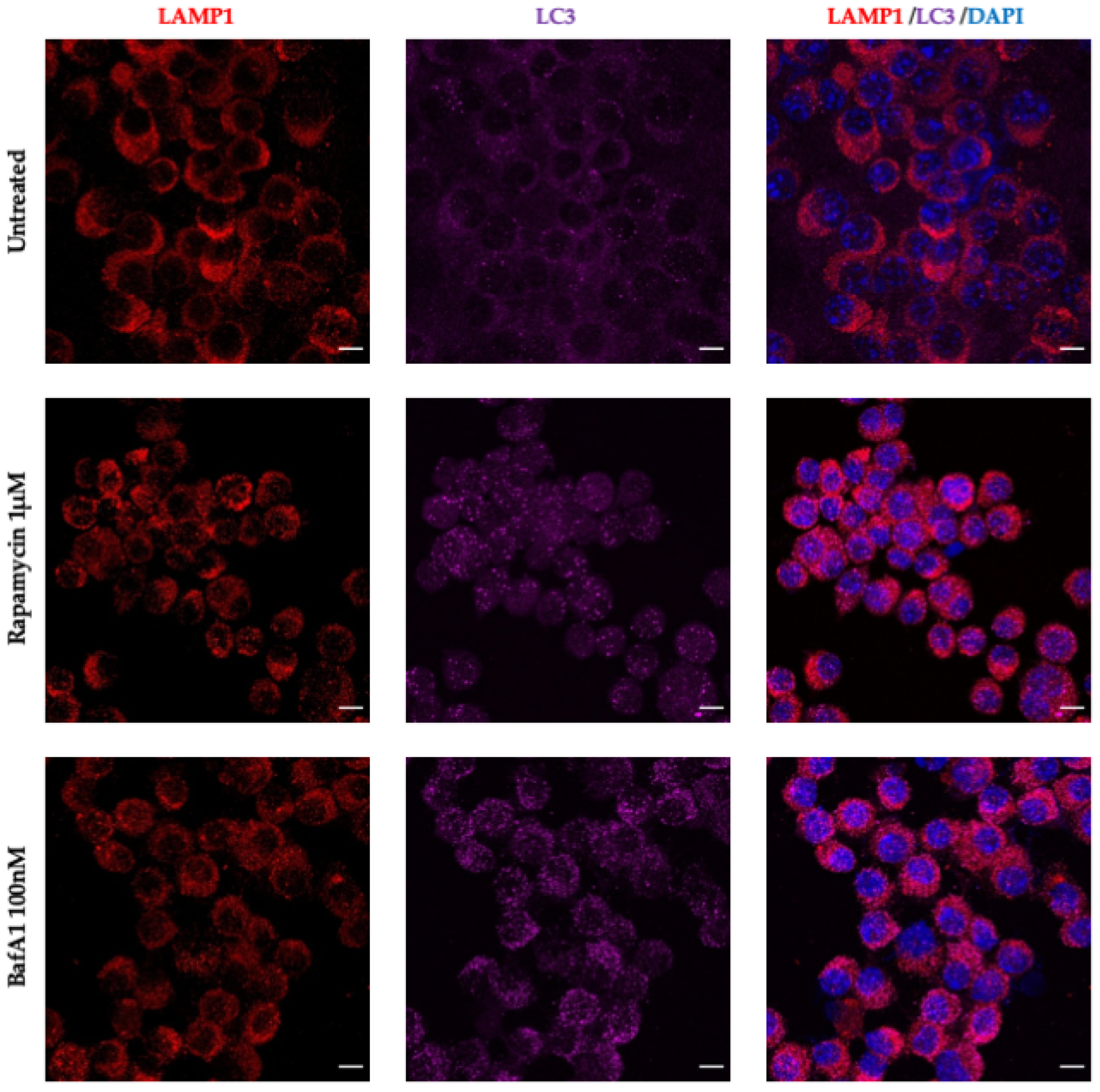
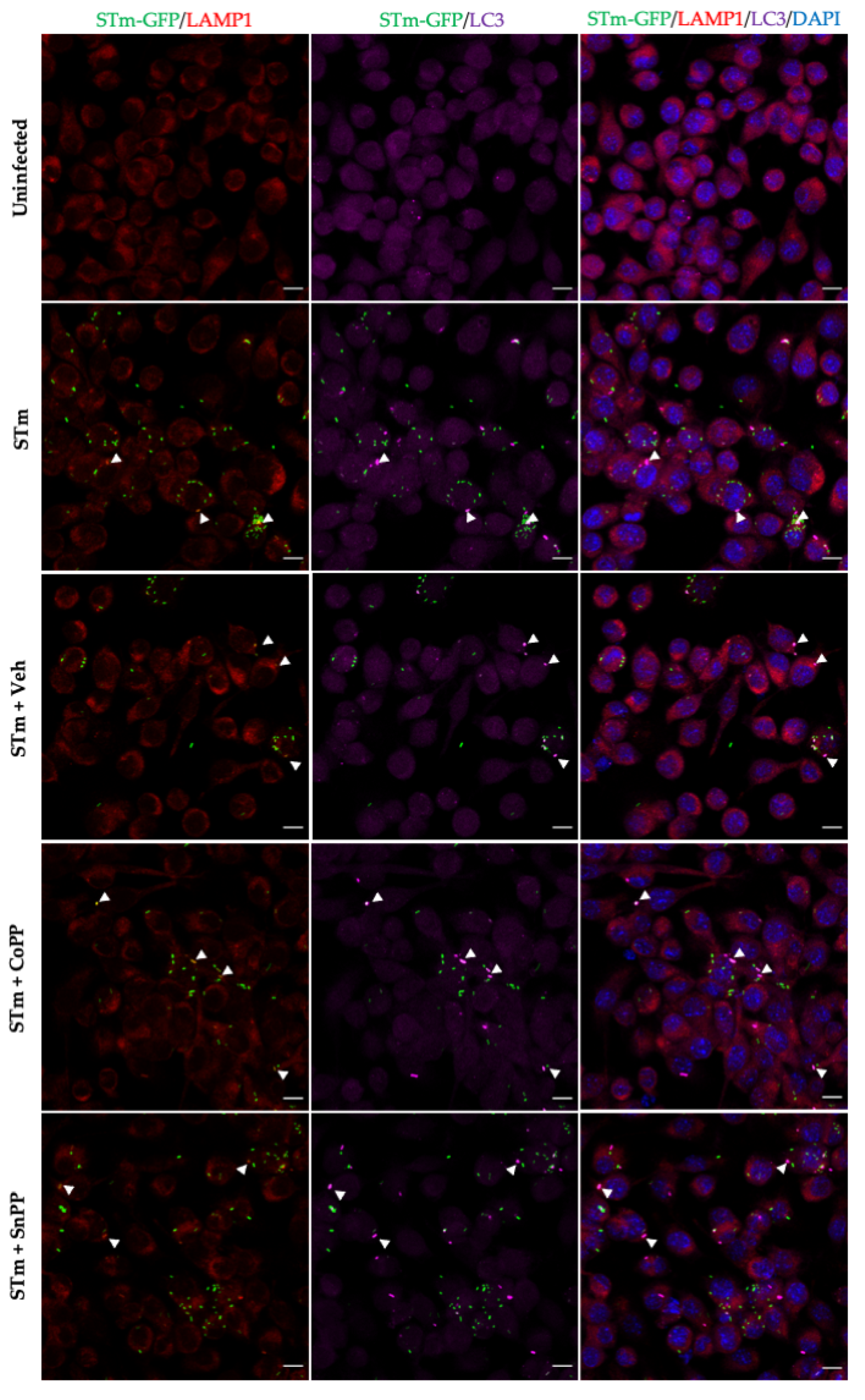
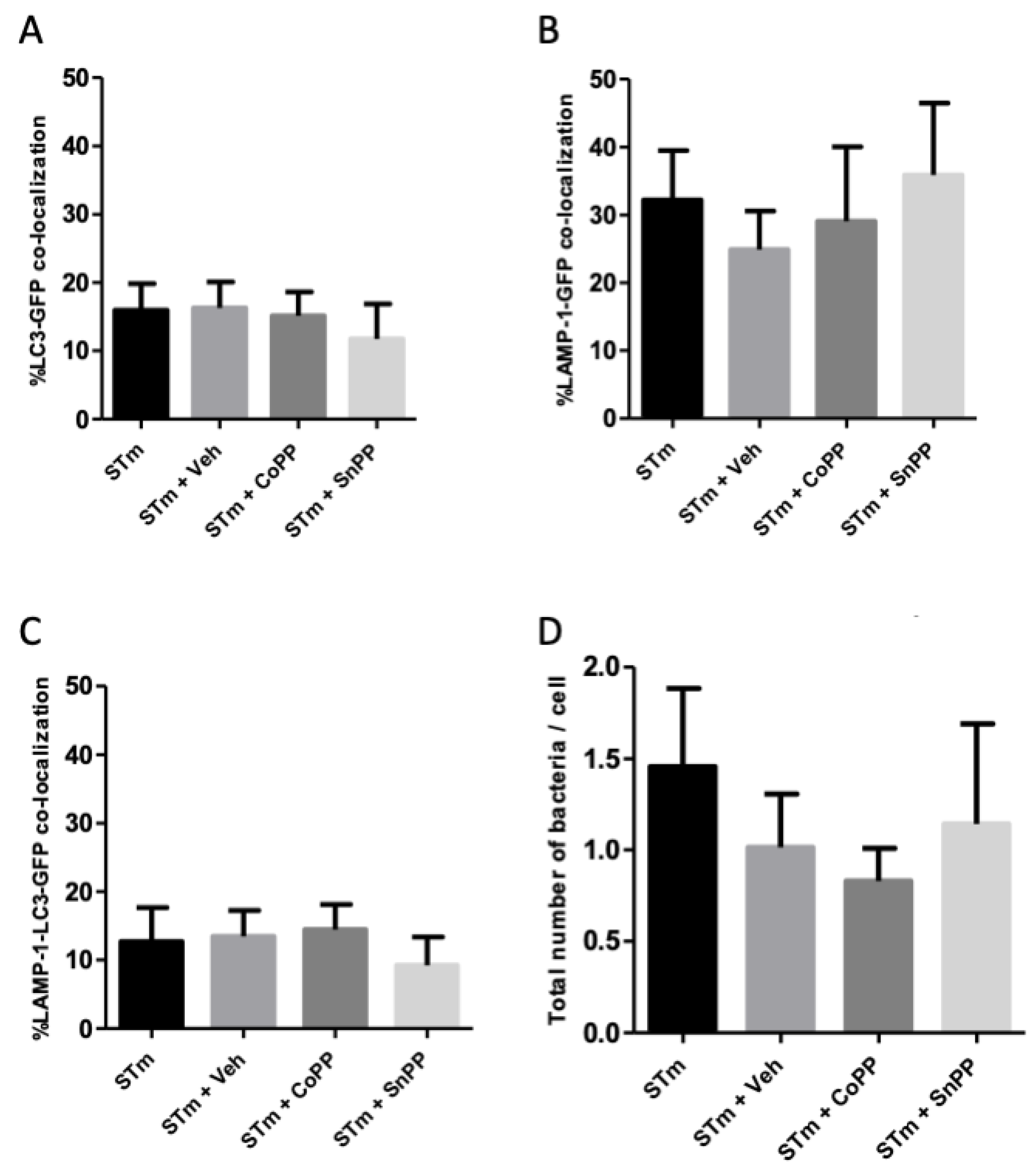
3.5. CoPP and SnPP Treatments Modulate S. Typhimurium Survival in RAW264.7 Cells in a CO-Independent Manner
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observations | Score | |
|---|---|---|
| Physiological parameters: -weight loss | Normal (no weight loss) | 0 |
| Weight loss under 10% | 1 | |
| Weight loss between 10–20% | 2 | |
| Weight loss greater than 20% | 3 | |
| Appearance parameters: -posture | Normal (erect, intense activity, clean fur) | 0 |
| Lower activity, hirsute fur, presence, or absence of porphyric secretion in eyes and nose | 1 | |
| Bowed position, very low activity, stays at the bottom of the cage, hirsute fur, porphyric secretion in eyes and nose | 2 | |
| Prostate, lateral decubitus, obvious dehydration, hollow eyes | 3 | |
| Spontaneous behavior: -response to stimuli | Normal (watchful, interacts with mates, groom) | 0 |
| Little changes: less groom and motion, accelerated breathing, tends to stay at the bottom of the cage | 1 | |
| Wobbly motion, inactive or stays at the bottom of the cage, abdominal breathing | 2 | |
| Motionless, prostrate, lateral decubitus, open its mouth to breath | 3 |

References
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States-Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Wang, Y.; Su, L.H.; Chiu, C.H. Nontyphoid Salmonella infection: Microbiology, clinical features, and antimicrobial therapy. Pediatr. Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helms, M.; Ethelberg, S.; Mølbak, K.; Lightfoot, D.; Powling, J.; Berghold, C.; Kornshober, C.; Wybo, I.; Collard, J.M.; Godard, C.; et al. International Salmonella typhimurium DT104 infections, 1992–2001. Emerg. Infect. Dis. 2005, 11, 859–867. [Google Scholar] [CrossRef]
- Bueno, S.M.; Riquelme, S.; Riedel, C.A.; Kalergis, A.M. Mechanisms used by virulent Salmonella to impair dendritic cell function and evade adaptive immunity. Immunology 2012, 137, 28–36. [Google Scholar] [CrossRef]
- de Jong, H.K.; Parry, C.M.; van der Poll, T.; Wiersinga, W.J. Host-Pathogen Interaction in Invasive Salmonellosis. PLoS Pathog. 2012, 8, e1002933. [Google Scholar] [CrossRef] [Green Version]
- Salazar, G.A.; Peñaloza, H.F.; Pardo-Roa, C.; Schultz, B.M.; Muñoz-Durango, N.; Gómez, R.S.; Salazar, F.J.; Pizarro, D.P.; Riedel, C.A.; González, P.A.; et al. Interleukin-10 production by T and B cells is a key factor to promote systemic Salmonella enterica serovar typhimurium infection in mice. Front. Immunol. 2017, 8, 889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DelGiorno, K.E.; Tam, J.W.; Hall, J.C.; Thotakura, G.; Crawford, H.C.; van der Velden, A.W.M. Persistent salmonellosis causes pancreatitis in a murine model of infection. PLoS ONE 2014, 9, e92807. [Google Scholar] [CrossRef] [Green Version]
- Schultz, B.M.; Salazar, G.A.; Paduro, C.A.; Pardo-Roa, C.; Pizarro, D.P.; Salazar-Echegarai, F.J.; Torres, J.; Riedel, C.A.; Kalergis, A.M.; Álvarez-Lobos, M.M.; et al. Persistent Salmonella enterica serovar Typhimurium infection increases the susceptibility of mice to develop intestinal inflammation. Front. Immunol. 2018, 9, 1166. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, P.; Regoes, R.R.; Dolowschiak, T.; Wotzka, S.Y.; Lengefeld, J.; Slack, E.; Grant, A.J.; Ackermann, M.; Hardt, W.D. Cecum Lymph Node Dendritic Cells Harbor Slow-Growing Bacteria Phenotypically Tolerant to Antibiotic Treatment. PLoS Biol. 2014, 12, e1001793. [Google Scholar] [CrossRef]
- Monack, D.M.; Bouley, D.M.; Falkow, S. Salmonella typhimurium persists within macrophages in the mesenteric lymph nodes of chronically infected Nramp1+/+ mice and can be reactivated by IFNgamma neutralization. J. Exp. Med. 2004, 199, 231–241. [Google Scholar] [CrossRef]
- Goldberg, M.F.; Roeske, E.K.; Ward, L.N.; Pengo, T.; Dileepan, T.; Kotov, D.I.; Jenkins, M.K. Salmonella Persist in Activated Macrophages in T Cell-Sparse Granulomas but Are Contained by Surrounding CXCR3 Ligand-Positioned Th1 Cells. Immunity 2018, 49, 1090–1102.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryter, S.W.; Alam, J.; Choi, A.M.K. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef] [PubMed]
- Schuller, D.J.; Wilks, A.; Ortiz de Montellano, P.R.; Poulos, T.L. Crystal structure of human heme oxygenase-1. Nat. Struct. Biol. 1999, 6, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Ferrandiz, M.L.; Devesa, I. Inducers of Heme Oxygenase-1. Curr. Pharm. Des. 2008, 14, 473–486. [Google Scholar] [CrossRef]
- Vareille, M.; Rannou, F.; Thélier, N.; Glasser, A.L.; de Sablet, T.; Martin, C.; Gobert, A.P. Heme Oxygenase-1 Is a Critical Regulator of Nitric Oxide Production in Enterohemorrhagic Escherichia coli-Infected Human Enterocytes. J. Immunol. 2008, 180, 5720–5726. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, J.; León, M.; Céspedes, P.; Gómez, P.; Canedo-Marroquín, G.; Riquelme, S.; Salazar-Echegarai, F.; Blancou, P.; Simon, T.; Kollias, G.; et al. Heme Oxygenase-1 modulates Human Respiratory Syncytial Virus replication and lung pathogenesis during infection. J. Immunol. 2017, 199, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Sebastian, V.P.; Salazar, G.A.; Coronado-Arrazola, I.; Schultz, B.M.; Vallejos, O.P.; Berkowitz, L.; Alvarez-Lobos, M.M.; Riedel, C.A.; Kalergis, A.M.; Bueno, S.M. Heme Oxygenase-1 as a Modulator of Intestinal Inflammation Development and Progression. Front. Immunol. 2018, 9, 1956. [Google Scholar] [CrossRef]
- Sachar, M.; Anderson, K.E.; Ma, X. Protoporphyrin IX: The good, the bad, and the ugly. J. Pharmacol. Exp. Ther. 2016, 356, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Shan, Y.; Lambrecht, R.W.; Donohue, S.E.; Bonkovsky, H.L. Role of Bach1 and Nrf2 in up-regulation of the heme oxygenase-1 gene by cobalt protoporphyrin. FASEB J. 2006, 20, 2651–2653. [Google Scholar] [CrossRef] [Green Version]
- Rosa, A.O.; Egea, J.; Lorrio, S.; Rojo, A.I.; Cuadrado, A.; López, M.G. Nrf2-mediated haeme oxygenase-1 up-regulation induced by cobalt protoporphyrin has antinociceptive effects against inflammatory pain in the formalin test in mice. Pain 2008, 137, 332–339. [Google Scholar] [CrossRef]
- Sardana, M.K.; Kappas, A. Dual control mechanism for heme oxygenase: Tin(IV)-protoporphyrin potently inhibits enzyme activity while markedly increasing content of enzyme protein in liver. Proc. Natl. Acad. Sci. USA 1987, 84, 2464–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riquelme, S.A.; Carreño, L.J.; Espinoza, J.A.; Mackern-Oberti, J.P.; Alvarez-Lobos, M.M.; Riedel, C.A.; Bueno, S.M.; Kalergis, A.M. Modulation of antigen processing by haem-oxygenase 1. Implications on inflammation and tolerance. Immunology 2016, 149, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferretti, L.; Ledda, A.; Wymant, C.; Zhao, L.; Ledda, V.; Abeler- Dorner, L.; Kendall, M.; Nurtay, A.; Cheng, H.-Y.; Ng, T.-C.; et al. The timing of COVID-19 transmission. medRxiv 2020. [Google Scholar] [CrossRef]
- Lin, H.W.; Chen, T.C.; Yeh, J.H.; Tsou, S.C.; Wang, I.; Shen, T.J.; Chuang, C.J.; Chang, Y.Y. Suppressive Effect of Tetrahydrocurcumin on Pseudomonas aeruginosa Lipopolysaccharide-Induced Inflammation by Suppressing JAK/STAT and Nrf2/HO-1 Pathways in Microglial Cells. Oxid. Med. Cell. Longev. 2022, 2022, 4978556. [Google Scholar] [CrossRef]
- Ahmad, I.M.; Dafferner, A.J.; O’connell, K.A.; Mehla, K.; Britigan, B.E.; Hollingsworth, M.A.; Abdalla, M.Y. Heme oxygenase-1 inhibition potentiates the effects of nab-paclitaxel-gemcitabine and modulates the tumor microenvironment in pancreatic ductal adenocarcinoma. Cancers 2021, 13, 2264. [Google Scholar] [CrossRef]
- Maines, M.D.; Kappas, A. Enzymatic Oxidation of Cobalt Protoporphyrin IX: Observations on the Mechanism of Heme Oxygenase Action. Biochemistry 1977, 16, 419–423. [Google Scholar] [CrossRef]
- Berglund, L.; Angelin, B.; Blomstrand, R.; Drummond, G.; Kappas, A. Sn-protoporphyrin lowers serum bilirubin levels, decreases biliary bilirubin output, enhances biliary heme excretion and potently inhibits hepatic heme oxygenase activity in normal human subjects. Hepatology 1988, 8, 625–631. [Google Scholar] [CrossRef]
- Jözkowicz, A.; Huk, I.; Nigisch, A.; Weigel, G.; Dietrich, W.; Motterlini, R.; Dulak, J. Heme Oxygenase and Angiogenic Activity of Endothelial Cells: Stimulation by Carbon Monoxide and Inhibition by Tin Protoporphyrin-IX. Antioxid. Redox Signal. 2004, 5, 155–162. [Google Scholar] [CrossRef]
- Rana, N.; McLean, S.; Mann, B.E.; Poole, R.K. Interaction of the carbon monoxide-releasing molecule Ru(CO)3Cl(glycinate) (CORM-3) with Salmonella enterica serovar Typhimurium: In situ measurements of CO binding by integrating cavity dual beam spectrophotometry. Microbiology 2014, 160, 2771–2779. [Google Scholar] [CrossRef]
- Onyiah, J.C.; Sheikh, S.Z.; Maharshak, N.; Steinbach, E.C.; Russo, S.M.; Kobayashi, T.; Mackey, L.C.; Hansen, J.J.; Moeser, A.J.; Rawls, J.F.; et al. Carbon monoxide and heme oxygenase-1 prevent intestinal inflammation in mice by promoting bacterial clearance. Gastroenterology 2013, 144, 789–798. [Google Scholar] [CrossRef] [Green Version]
- Wegiel, B.; Larsen, R.; Gallo, D.; Chin, B.Y.; Harris, C.; Mannam, P.; Kaczmarek, E.; Lee, P.J.; Zuckerbraun, B.S.; Flavell, R.; et al. Macrophages sense and kill bacteria through carbon monoxide—Dependent inflammasome activation. J. Clin. Investig. 2014, 124, 4926–4940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitterstiller, A.-M.; Haschka, D.; Dichtl, S.; Nairz, M.; Demetz, E.; Talasz, H.; Soares, M.; Einwallner, E.; Esterbauer, H.; Fang, F.C.; et al. Heme oxygenase 1 controls early innate immune response of macrophages to Salmonella typhimurium infection. Cell. Microbiol. 2016, 18, 1374–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deretic, V. Autophagy in Immunity and Cell-Autonomous Defense against Intracellular Microbes. Immunol. Rev. 2011, 240, 92–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliva, L.; Cenci, S. Autophagy in plasma cell pathophysiology. Front. Immunol. 2014, 5, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cemma, M.; Brumell, J.H.H. Interactions of Pathogenic Bacteria with Autophagy Systems. Curr. Biol. 2012, 22, R540–R545. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition) 1. Autophagy 2021, 17, 1–382. [Google Scholar] [CrossRef]
- Sahlberg Bang, C.; Kruse, R.; Johansson, K.; Persson, K. Carbon monoxide releasing molecule-2 (CORM-2) inhibits growth of multidrug-resistant uropathogenic Escherichia coli in biofilm and following host cell colonization. BMC Microbiol. 2016, 16, 64. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, S.A.; Bueno, S.M.; Kalergis, A.M. Carbon monoxide down-modulates Toll-like receptor 4/MD2 expression on innate immune cells and reduces endotoxic shock susceptibility. Immunology 2015, 144, 321–332. [Google Scholar] [CrossRef]
- Su, W.C.; Liu, X.; Macias, A.A.; Baron, R.M.; Perrella, M.A. Heme oxygenase-1-derived carbon monoxide enhances the host defense response to microbial sepsis in mice. J. Clin. Investig. 2008, 118, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Wang, B. Toward carbon monoxide based therapeutics: Carbon monoxide in a pill. Pharm. Pat. Anal. 2017, 6, 171–177. [Google Scholar] [CrossRef]
- Birmingham, C.L.; Smith, A.C.; Bakowski, M.A.; Yoshimori, T.; Brumell, J.H. Autophagy controls Salmonella infection in response to damage to the Salmonella-containing vacuole. J. Biol. Chem. 2006, 281, 11374–11383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tattoli, I.; Sorbara, M.T.; Vuckovic, D.; Ling, A.; Soares, F.; Carneiro, L.A.M.; Yang, C.; Emili, A.; Philpott, D.J.; Girardin, S.E. Amino acid starvation induced by invasive bacterial pathogens triggers an innate host defense program. Cell Host Microbe 2012, 11, 563–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robey, R.B.; Ruiz, O.; Santos, A.V.P.; Ma, J.; Kear, F.; Wang, L.J.; Li, C.J.; Bernardo, A.A.; Arruda, J.A.L. pH-Dependent Fluorescence of a Heterologously Expressed Aequorea Green Fluorescent Protein Mutant: In Situ Spectral Characteristics and Applicability to Intracellular pH Estimation. Biochemistry 1998, 37, 9894–9901. [Google Scholar] [CrossRef] [PubMed]
- Griffin, A.J.; Li, L.X.; Voedisch, S.; Pabst, O.; McSorley, S.J. Dissemination of persistent intestinal bacteria via the mesenteric lymph nodes causes typhoid relapse. Infect. Immun. 2011, 79, 1479–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamal Uddin, M.; Joe, Y.; Kim, S.-K.; Oh Jeong, S.; Ryter, S.W.; Pae, H.-O.; Taeg Chung, H. IRG1 induced by heme oxygenase-1/carbon monoxide inhibits LPS-mediated sepsis and pro-inflammatory cytokine production. Cell. Mol. Immunol. 2015, 13, 170–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, T.; Uchiyama, K.; Naito, Y. The therapeutic potential of carbon monoxide for inflammatory bowel disease. Digestion 2015, 91, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Neves, P.; Lampropoulou, V.; Calderon-Gomez, E.; Roch, T.; Stervbo, U.; Shen, P.; Kühl, A.A.; Loddenkemper, C.; Haury, M.; Nedospasov, S.A.; et al. Signaling via the MyD88 adaptor protein in B cells suppresses protective immunity during salmonella typhimurium infection. Immunity 2010, 33, 777–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.-S.; Chau, L.-Y. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice. Nat. Med. 2002, 8, 240–246. [Google Scholar] [CrossRef]
- Chen, S.; Kapturczak, M.H.; Wasserfall, C.; Glushakova, O.Y.; Campbell-thompson, M.; Deshane, J.S.; Joseph, R.; Cruz, P.E.; Hauswirth, W.W.; Madsen, K.M.; et al. Interleukin 10 attenuates neointimal proliferation and inflammation in aortic allografts by a heme oxygenase-dependent pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 7251–7256. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, D.W. Pharmacokinetics of cobalt chloride and cobalt-protoporphyrin. Drug Metab. Dispos. 1993, 21, 846–849. [Google Scholar]
- Boyer, E.; Bergevin, I.; Malo, D.; Gros, P.; Cellier, M.F.M. Acquisition of Mn(II) in Addition to Fe(II) Is Required for Full Virulence of Salmonella enterica serovar Typhimurium. Infect. Immun. 2002, 70, 6032–6042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minasyan, H. Mechanisms and pathways for the clearance of bacteria from blood circulation in health and disease. Pathophysiology 2016, 23, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Pham, O.H.; McSorley, S.J. Protective host immune responses to Salmonella infection. Future Microbiol. 2015, 10, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojiljkovic, I.; Evavold, B.D.; Kumar, V. Antimicrobial properties of porphyrins. Expert Opin. Investig. Drugs 2001, 10, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Gyulkhandanyan, G.V.; Ghazaryan, R.K.; Paronyan, M.H.; Ulikhanyan, G.I.; Gyulkhandanyan, A.G.; Sahakyan, L.A. Antimicrobial activity of new porphyrins of synthetic and natural origin. In Proceedings of the Progress in Biomedical Optics and Imaging—Proceedings of SPIE, San Francisco, CA, USA, 21–26 January 2012; Volume 8211. [Google Scholar] [CrossRef]
- Runyen-Janecky, L.J. Role and regulation of heme iron acquisition in gram-negative pathogens. Front. Cell. Infect. Microbiol. 2013, 3, 55. [Google Scholar] [CrossRef] [Green Version]
- Stojiljkovic, I.; Kumar, V.; Srinivasan, N. Non-iron metalloporphyrins: Potent antibacterial compounds that exploit haem/Hb uptake systems of pathogenic bacteria. Mol. Microbiol. 1999, 31, 429–442. [Google Scholar] [CrossRef]
- Schmid, D.; Münz, C. Innate and Adaptive Immunity through Autophagy. Immunity 2007, 27, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yan, J.; Niu, H.; Huang, R.; Wu, S. Autophagy and ubiquitination in Salmonella infection and the related inflammatory responses. Front. Cell. Infect. Microbiol. 2018, 8, 78. [Google Scholar] [CrossRef]
- Kreibich, S.; Emmenlauer, M.; Fredlund, J.; Rämö, P.; Münz, C.; Dehio, C.; Enninga, J.; Hardt, W.D. Autophagy proteins promote repair of endosomal membranes damaged by the Salmonella type three secretion system 1. Cell Host Microbe 2015, 18, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Fujii, S.; Matsunaga, T.; Nishimura, A.; Ono, K.; Ida, T. Reactive Persulfides from Salmonella Typhimurium Downregulate Autophagy-Mediated Innate Immunity in Macrophages by Inhibiting Electrophilic Signaling Article Reactive Persulfides from Salmonella Typhimurium Downregulate Autophagy-Mediated Innate Immunity. Cell Chem. Biol. 2018, 25, 1403–1413.e4. [Google Scholar] [CrossRef]
- Nurmi, K.; Kareinen, I.; Virkanen, J.; Rajamäki, K.; Kouri, V.P.; Vaali, K.; Levonen, A.L.; Fyhrquist, N.; Matikainen, S.; Kovanen, P.T.; et al. Hemin and Cobalt Protoporphyrin Inhibit NLRP3 Inflammasome Activation by Enhancing Autophagy: A novel mechanism of inflammasome regulation. J. Innate Immun. 2017, 9, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Unuma, K.; Aki, T.; Matsuda, S.; Funakoshi, T.; Yoshida, K.-I.; Uemura, K. Inducer of heme oxygenase-1 cobalt protoporphyrin accelerates autophagy and suppresses oxidative damages during lipopolysaccharide treatment in rat liver. Hepatol. Res. 2013, 43, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Carr, J.F.; Garcia, D.; Scaffa, A.; Peterson, A.L.; Ghio, A.J.; Dennery, P.A. Heme Oxygenase-1 Supports Mitochondrial Energy Production and Electron Transport Chain Activity in Cultured Lung Epithelial Cells. Int. J. Mol. Sci. 2020, 21, 6941. [Google Scholar] [CrossRef] [PubMed]
- MacGarvey, N.C.; Suliman, H.B.; Bartz, R.R.; Fu, P.; Withers, C.M.; Welty-Wolf, K.E.; Piantadosi, C.A. Activation of mitochondrial biogenesis by heme oxygenase-1-mediated NF-E2-related factor-2 induction rescues mice from lethal Staphylococcus aureus sepsis. Am. J. Respir. Crit. Care Med. 2012, 185, 851–861. [Google Scholar] [CrossRef] [Green Version]
- Müllebner, A.; Dorighello, G.G.; Kozlov, A.V.; Duvigneau, J.C. Interaction between Mitochondrial Reactive Oxygen Species, Heme Oxygenase, and Nitric Oxide Synthase Stimulates Phagocytosis in Macrophages. Front. Med. 2018, 4, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, R.J.; Vreman, H.J.; Schulz, S.; Kalish, F.S.; Pierce, N.W.; Stevenson, D.K. In vitro inhibition of heme oxygenase isoenzymes by metalloporphyrins. J. Perinatol. 2011, 31 (Suppl. S1), S35–S41. [Google Scholar] [CrossRef] [Green Version]
- Intagliata, S.; Salerno, L.; Ciaffaglione, V.; Leonardi, C.; Fallica, A.N.; Carota, G.; Amata, E.; Marrazzo, A.; Pittalà, V.; Romeo, G. Heme Oxygenase-2 (HO-2) as a therapeutic target: Activators and inhibitors. Eur. J. Med. Chem. 2019, 183, 111703. [Google Scholar] [CrossRef]
- Naito, Y.N.A.; Takagi, T.; Yoshikawa, T. Heme oxygenase-1: A new therapeutic target for inflammatory bowel disease. Aliment. Pharmacol. Ther. 2004, 20, 177–184. [Google Scholar] [CrossRef]
- Protzer, U.; Seyfried, S.; Quasdorff, M.; Sass, G.; Svorcova, M.; Webb, D.; Bohne, F.; Hösel, M.; Schirmacher, P.; Tiegs, G. Antiviral activity and hepatoprotection by heme oxygenase-1 in hepatitis B virus infection. Gastroenterology 2007, 133, 1156–1165. [Google Scholar] [CrossRef]
- Silva-Gomes, S.; Appelberg, R.; Larsen, R.; Soares, M.P.; Gomes, M.S. Heme catabolism by heme oxygenase-1 confers host resistance to Mycobacterium infection. Infect. Immun. 2013, 81, 2536–2545. [Google Scholar] [CrossRef] [Green Version]


| Salmonella Typhimurium Load 1 | |||||
|---|---|---|---|---|---|
| Experimental Group | Mouse | Liver (CFU/mg) | Spleen (CFU/mg) | mLN (CFU/mg) | Blood (CFU/mL) |
| STm pre-Veh | 1 | >5000 | 1.44 | - | - |
| 2 | - | - | - | - | |
| 3 | >5000 | >5000 | 4.4 | 333.3 | |
| 4 | - | - | - | - | |
| 5 | - | - | - | - | |
| 6 | - | - | - | - | |
| 7 | - | - | - | - | |
| 8 | - | - | - | - | |
| STm pre-CoPP | 1 | 41.7 | |||
| 2 | - | - | - | - | |
| 3 | - | - | - | - | |
| 4 | - | - | - | - | |
| 5 | - | - | - | - | |
| 6 | - | - | - | - | |
| 7 | - | - | - | - | |
| 8 | - | - | - | - | |
| 9 | - | - | - | - | |
| 10 | - | - | - | - | |
| STm pre-SnPP | 1 | 181.8 | - | - | - |
| 2 | - | - | - | - | |
| 3 | - | - | - | - | |
| 4 | - | >5000 | - | - | |
| 5 | - | - | - | - | |
| 6 | - | >5000 | - | 466.7 | |
| 7 | - | - | - | - | |
| 8 | - | - | - | - | |
| 9 | - | - | - | - | |
| 10 | - | - | - | - | |
| 11 | - | - | - | - | |
| 12 | - | - | - | - | |
| 13 | - | - | - | - | |
| 14 | 6.41 | 12.9 | - | - | |
| Salmonella Typhimurium Load 1 | |||||
|---|---|---|---|---|---|
| Experimental Group | Mouse | Liver (CFU/mg) | Spleen (CFU/mg) | mLN (CFU/mg) | Blood (CFU/mL) |
| STm post-Veh | 1 | - | 171.8 | - | - |
| 2 | - | 61.7 | - | - | |
| 3 | - | 61.7 | - | - | |
| 4 | - | - | - | - | |
| 5 | - | - | - | - | |
| STm post-CoPP | 1 | - | - | - | - |
| 2 | - | - | - | - | |
| 3 | - | - | - | - | |
| 4 | - | - | - | - | |
| 5 | - | - | - | - | |
| 6 | - | - | - | - | |
| STm post-SnPP | 1 | - | 13.6 | - | - |
| 2 | - | 95.5 | - | - | |
| 3 | - | - | - | - | |
| 4 | - | >5000 | >5000 | >5000 | |
| 5 | - | 11 | 40.9 | - | |
| 6 | - | - | - | - | |
| 7 | - | - | - | - | |
| 8 | - | - | - | - | |
| 9 | - | - | - | - | |
| 10 | - | - | - | - | |
| 11 | - | - | - | - | |
| 12 | - | - | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sebastián, V.P.; Moreno-Tapia, D.; Melo-González, F.; Hernández-Cáceres, M.P.; Salazar, G.A.; Pardo-Roa, C.; Farías, M.A.; Vallejos, O.P.; Schultz, B.M.; Morselli, E.; et al. Limited Heme Oxygenase Contribution to Modulating the Severity of Salmonella enterica serovar Typhimurium Infection. Antioxidants 2022, 11, 1040. https://doi.org/10.3390/antiox11061040
Sebastián VP, Moreno-Tapia D, Melo-González F, Hernández-Cáceres MP, Salazar GA, Pardo-Roa C, Farías MA, Vallejos OP, Schultz BM, Morselli E, et al. Limited Heme Oxygenase Contribution to Modulating the Severity of Salmonella enterica serovar Typhimurium Infection. Antioxidants. 2022; 11(6):1040. https://doi.org/10.3390/antiox11061040
Chicago/Turabian StyleSebastián, Valentina P., Daniela Moreno-Tapia, Felipe Melo-González, María P. Hernández-Cáceres, Geraldyne A. Salazar, Catalina Pardo-Roa, Mónica A. Farías, Omar P. Vallejos, Bárbara M. Schultz, Eugenia Morselli, and et al. 2022. "Limited Heme Oxygenase Contribution to Modulating the Severity of Salmonella enterica serovar Typhimurium Infection" Antioxidants 11, no. 6: 1040. https://doi.org/10.3390/antiox11061040
APA StyleSebastián, V. P., Moreno-Tapia, D., Melo-González, F., Hernández-Cáceres, M. P., Salazar, G. A., Pardo-Roa, C., Farías, M. A., Vallejos, O. P., Schultz, B. M., Morselli, E., Álvarez-Lobos, M. M., González, P. A., Kalergis, A. M., & Bueno, S. M. (2022). Limited Heme Oxygenase Contribution to Modulating the Severity of Salmonella enterica serovar Typhimurium Infection. Antioxidants, 11(6), 1040. https://doi.org/10.3390/antiox11061040