
Antioxidant Genetic Profile Modifies Probability of Developing Neurological Sequelae in Long-COVID


 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
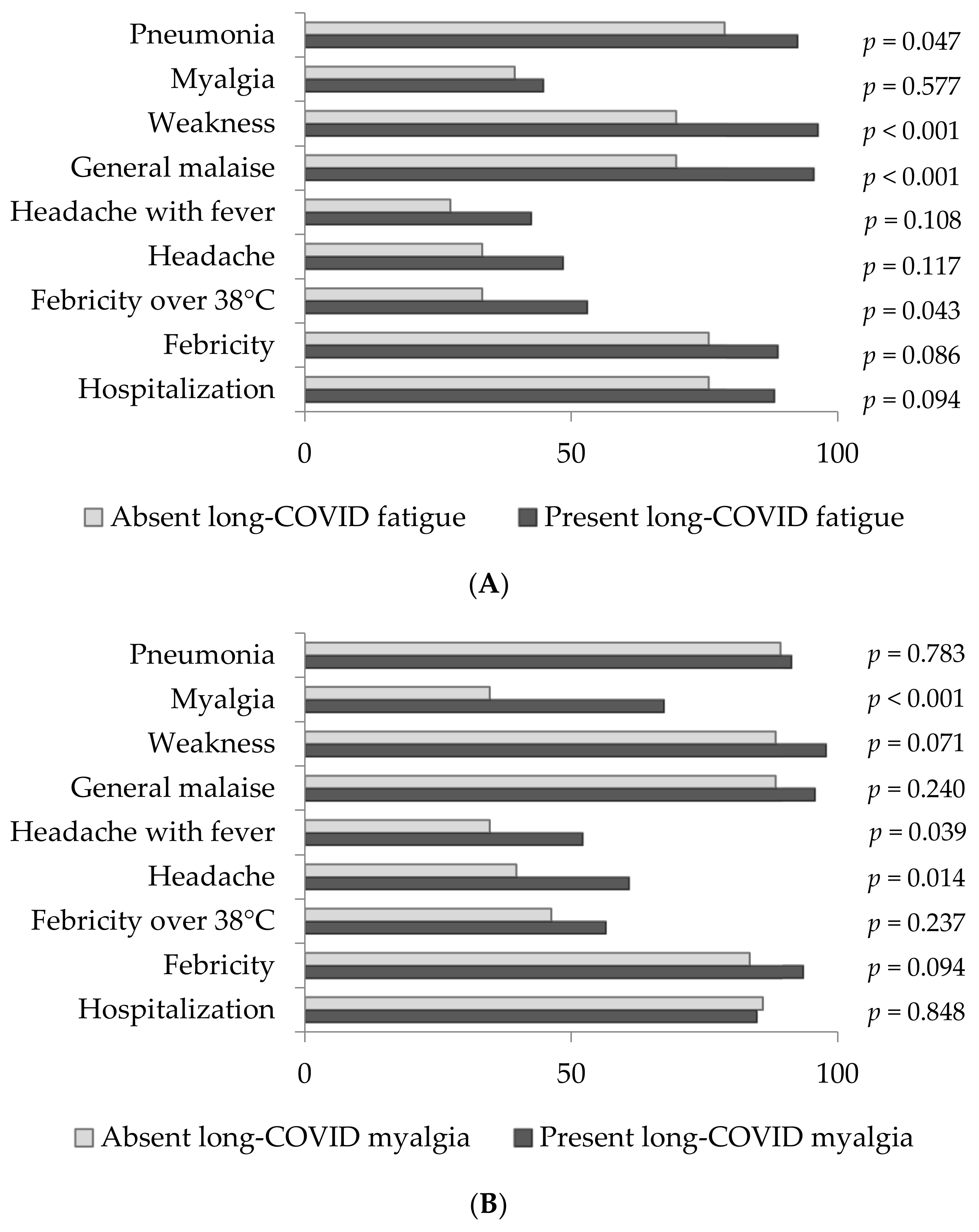
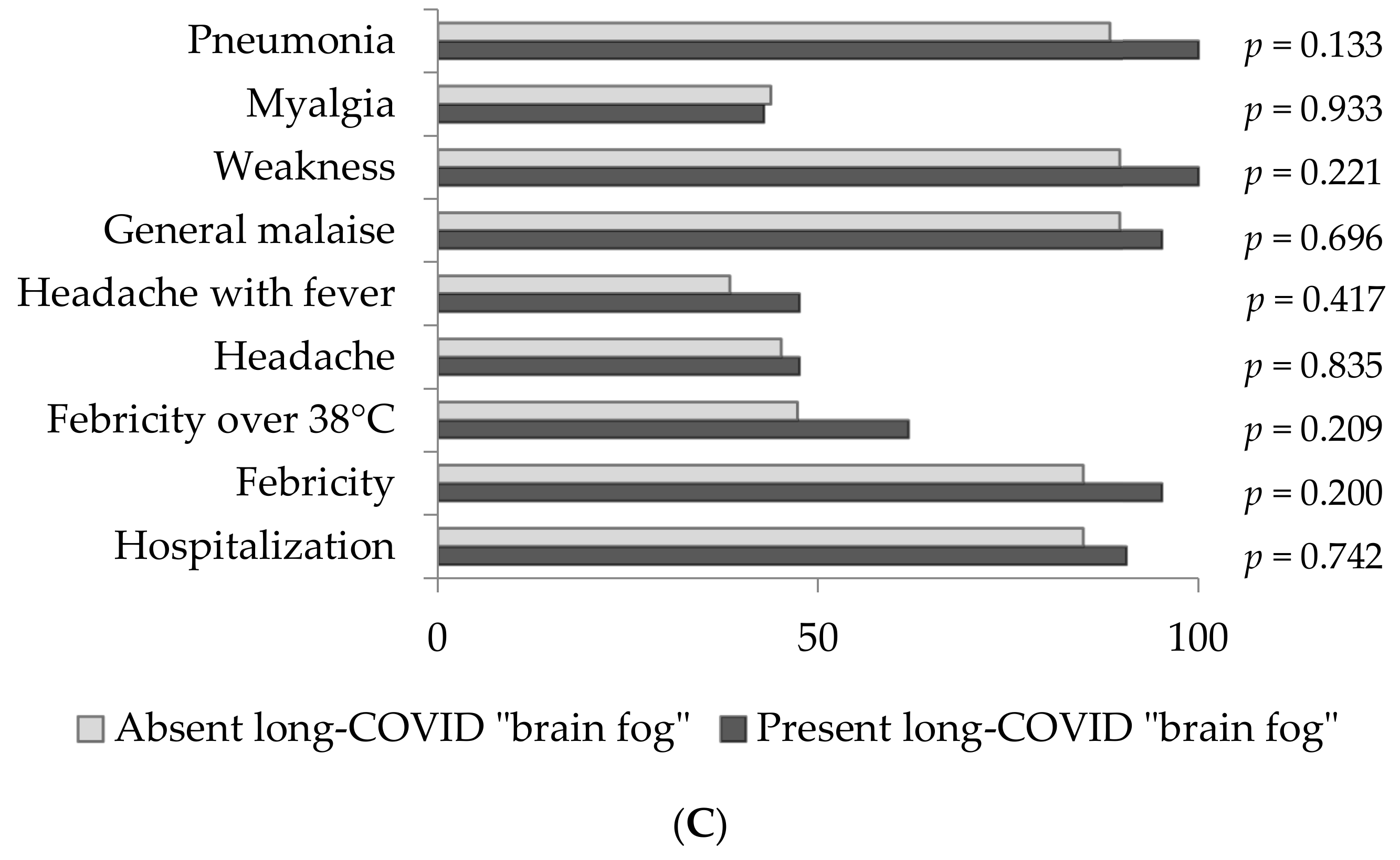
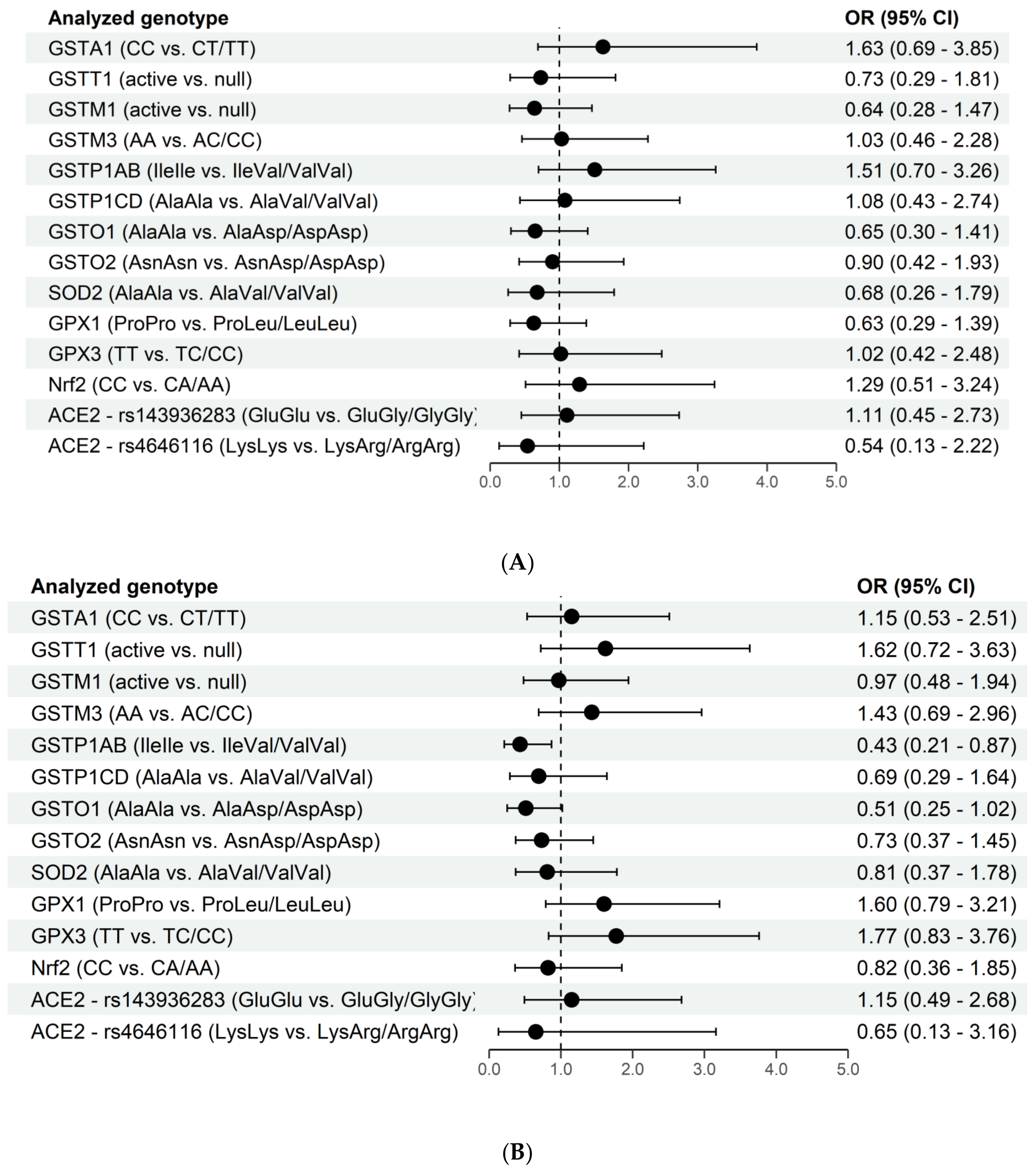
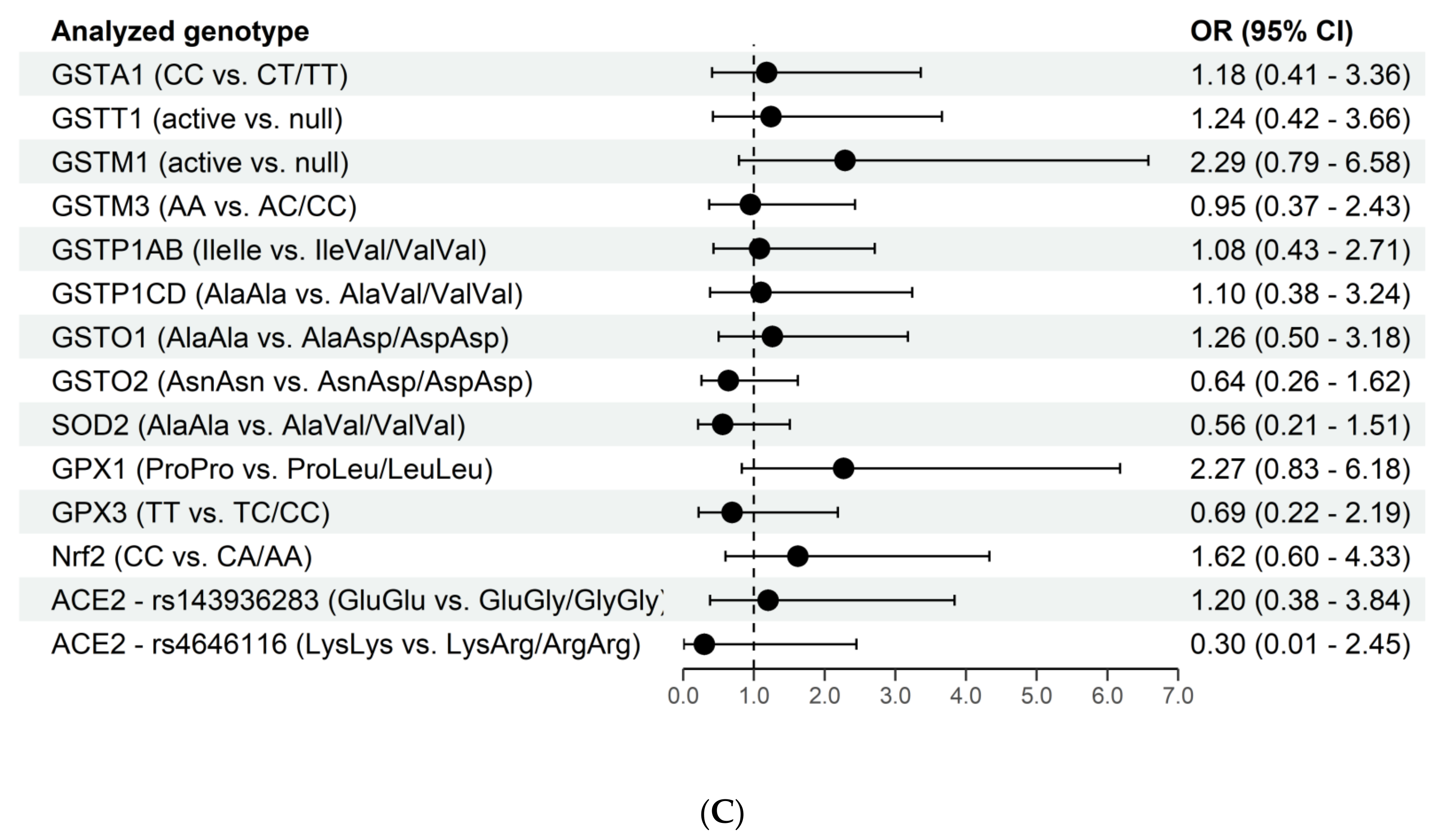
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stefanou, M.-I.; Palaiodimou, L.; Bakola, E.; Smyrnis, N.; Papadopoulou, M.; Paraskevas, G.P.; Rizos, E.; Boutati, E.; Grigoriadis, N.; Krogias, C. Neurological manifestations of long-COVID syndrome: A narrative review. Ther. Adv. Chronic Dis. 2022, 13, 20406223221076890. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.J.; Sarangi, A.; Bangash, F. Neurological sequelae of COVID-19: A review. Egypt. J. Neurol. Psychiatry Neurosurg. 2021, 57, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nolen, L.T.; Mukerji, S.S.; Mejia, N.I. Post-acute neurological consequences of COVID-19: An unequal burden. Nat. Med. 2022, 28, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Sivan, M.; Taylor, S. NICE guideline on long COVID. BMJ 2020, 371, m4938. [Google Scholar] [CrossRef] [PubMed]
- Callard, F.; Perego, E. How and why patients made Long COVID. Soc. Sci. Med. 2021, 268, 113426. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, F.; Shen, Y.; Zhang, X.; Cen, Y.; Wang, B.; Zhao, S.; Zhou, Y.; Hu, B.; Wang, M. Symptoms and health outcomes among survivors of COVID-19 infection 1 year after discharge from hospitals in Wuhan, China. JAMA Netw. Open 2021, 4, e2127403. [Google Scholar] [CrossRef]
- Fernández-de-Las-Peñas, C.; Palacios-Ceña, D.; Gómez-Mayordomo, V.; Cuadrado, M.L.; Florencio, L.L. Defining post-COVID symptoms (post-acute COVID, long COVID, persistent post-COVID): An integrative classification. Int. J. Environ. Res. Public Health 2021, 18, 2621. [Google Scholar] [CrossRef]
- Long-Term-Effects-of-Coronavirus. Available online: https://www.nhs.uk/conditions/coronavirus-covid-19 (accessed on 24 March 2022).
- Liguori, C.; Pierantozzi, M.; Spanetta, M.; Sarmati, L.; Cesta, N.; Iannetta, M.; Ora, J.; Mina, G.G.; Puxeddu, E.; Balbi, O. Subjective neurological symptoms frequently occur in patients with SARS-CoV2 infection. Brain Behav. Immun. 2020, 88, 11–16. [Google Scholar] [CrossRef]
- Nuzzo, D.; Picone, P. Potential neurological effects of severe COVID-19 infection. Neurosci. Res. 2020, 158, 1–5. [Google Scholar] [CrossRef]
- Paterson, R.W.; Brown, R.L.; Benjamin, L.; Nortley, R.; Wiethoff, S.; Bharucha, T.; Jayaseelan, D.L.; Kumar, G.; Raftopoulos, R.E.; Zambreanu, L. The emerging spectrum of COVID-19 neurology: Clinical, radiological and laboratory findings. Brain 2020, 143, 3104–3120. [Google Scholar] [CrossRef]
- Chou, S.H.; Beghi, E.; Helbok, R. GCS-NeuroCOVID Consortium and ENERGY Consortium: Global incidence of neurological manifestations among patients hospitalized with COVID-19-a report for the GCS-NeuroCOVID Consortium and the ENERGY Consortium. JAMA Netw. Open 2021, 4, e2112131. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.L.R.; Hardwick, M.; Jeyanantham, A.; White, L.M.; Deb, S.; Burnside, G.; Joy, H.M.; Smith, C.J.; Pollak, T.A.; Nicholson, T.R. Spectrum, risk factors and outcomes of neurological and psychiatric complications of COVID-19: A UK-wide cross-sectional surveillance study. Brain Commun. 2021, 3, fcab168. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Jin, H.; Wang, M.; Hu, Y.; Chen, S.; He, Q.; Chang, J.; Hong, C.; Zhou, Y.; Wang, D.; et al. Neurologic Manifestations of Hospitalized Patients with Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol. 2020, 77, 683–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baig, A.M. Deleterious outcomes in long-hauler COVID-19: The effects of SARS-CoV-2 on the CNS in chronic COVID syndrome. ACS Chem. Neurosci. 2020, 11, 4017–4020. [Google Scholar] [CrossRef]
- Fiani, B.; Covarrubias, C.; Desai, A.; Sekhon, M.; Jarrah, R. A contemporary review of neurological sequelae of COVID-19. Front. Neurol. 2020, 11, 640. [Google Scholar] [CrossRef]
- Tavassoly, O.; Safavi, F.; Tavassoly, I. Seeding brain protein aggregation by SARS-CoV-2 as a possible long-term complication of COVID-19 infection. ACS Chem. Neurosci. 2020, 11, 3704–3706. [Google Scholar] [CrossRef]
- Sun, B.; Tang, N.; Peluso, M.J.; Iyer, N.S.; Torres, L.; Donatelli, J.L.; Munter, S.E.; Nixon, C.C.; Rutishauser, R.L.; Rodriguez-Barraquer, I. Characterization and biomarker analyses of post-COVID-19 complications and neurological manifestations. Cells 2021, 10, 386. [Google Scholar] [CrossRef]
- Fernandes, I.G.; De Brito, C.A.; Dos Reis, V.M.S.; Sato, M.N.; Pereira, N.Z. SARS-CoV-2 and other respiratory viruses: What does oxidative Stress have to do with it? Oxid. Med. Cell. Longev. 2020, 2020, 8844280. [Google Scholar] [CrossRef]
- Chatterjee, S. Oxidative stress, inflammation, and disease. In Oxidative Stress and Biomaterials; Elsevier: Amsterdam, The Netherlands, 2016; pp. 35–58. [Google Scholar]
- Cheng, Y.; Sun, F.; Wang, L.; Gao, M.; Xie, Y.; Sun, Y.; Liu, H.; Yuan, Y.; Yi, W.; Huang, Z. Virus-induced p38 MAPK activation facilitates viral infection. Theranostics 2020, 10, 12223. [Google Scholar] [CrossRef]
- Kosanovic, T.; Sagic, D.; Djukic, V.; Pljesa-Ercegovac, M.; Savic-Radojevic, A.; Bukumiric, Z.; Lalosevic, M.; Djordjevic, M.; Coric, V.; Simic, T. Time Course of Redox Biomarkers in COVID-19 Pneumonia: Relation with Inflammatory, Multiorgan Impairment Biomarkers and CT Findings. Antioxidants 2021, 10, 1126. [Google Scholar] [CrossRef]
- Wu, B.; Dong, D. Human cytosolic glutathione transferases: Structure, function, and drug discovery. Trends Pharmacol. Sci. 2012, 33, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Pljesa-Ercegovac, M.; Savic-Radojevic, A.; Matic, M.; Coric, V.; Djukic, T.; Radic, T.; Simic, T. Glutathione transferases: Potential targets to overcome chemoresistance in solid tumors. Int. J. Mol. Sci. 2018, 19, 3785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, D.; Innes, A.; Oakley, A.J.; Dahlstrom, J.E.; Jensen, L.M.; Brüstle, A.; Tummala, P.; Rooke, M.; Casarotto, M.G.; Baell, J.B. GSTO1-1 plays a pro-inflammatory role in models of inflammation, colitis and obesity. Sci. Rep. 2017, 7, 17832. [Google Scholar] [CrossRef] [PubMed]
- Djukic, T.; Stevanovic, G.; Coric, V.; Bukumiric, Z.; Pljesa-Ercegovac, M.; Matic, M.; Jerotic, D.; Todorovic, N.; Asanin, M.; Ercegovac, M. GSTO1, GSTO2 and ACE2 Polymorphisms Modify Susceptibility to Developing COVID-19. J. Pers. Med. 2022, 12, 458. [Google Scholar] [CrossRef] [PubMed]
- Silvagno, F.; Vernone, A.; Pescarmona, G.P. The role of glutathione in protecting against the severe inflammatory response triggered by COVID-19. Antioxidants 2020, 9, 624. [Google Scholar] [CrossRef] [PubMed]
- Coric, V.; Milosevic, I.; Djukic, T.; Bukumiric, Z.; Savic-Radojevic, A.; Matic, M.; Jerotic, D.; Todorovic, N.; Asanin, M.; Ercegovac, M. GSTP1 and GSTM3 variant alleles affect susceptibility and severity of COVID-19. Front. Mol. Biosci. 2021, 8, 747493. [Google Scholar] [CrossRef]
- Jerotic, D.; Ranin, J.; Bukumiric, Z.; Djukic, T.; Coric, V.; Savic-Radojevic, A.; Todorovic, N.; Asanin, M.; Ercegovac, M.; Milosevic, I. SOD2 rs4880 and GPX1 rs1050450 polymorphisms do not confer risk of COVID-19, but influence inflammation or coagulation parameters in Serbian cohort. Redox Rep. 2022, 27, 85–91. [Google Scholar] [CrossRef]
- Camargo-Martínez, W.; Lozada-Martínez, I.; Escobar-Collazos, A.; Navarro-Coronado, A.; Moscote-Salazar, L.; Pacheco-Hernández, A.; Janjua, T.; Bosque-Varela, P. Post-COVID 19 neurological syndrome: Implications for sequelae’s treatment. J. Clin. Neurosci. 2021, 88, 219–225. [Google Scholar] [CrossRef]
- Taquet, M.; Geddes, J.R.; Husain, M.; Luciano, S.; Harrison, P.J. 6-month neurological and psychiatric outcomes in 236,379 survivors of COVID-19: A retrospective cohort study using electronic health records. Lancet Psychiatry 2021, 8, 416–427. [Google Scholar] [CrossRef]
- Qin, Y.; Wu, J.; Chen, T.; Li, J.; Zhang, G.; Wu, D.; Zhou, Y.; Zheng, N.; Cai, A.; Ning, Q. Long-term microstructure and cerebral blood flow changes in patients recovered from COVID-19 without neurological manifestations. J. Clin. Investig. 2021, 131, e147329. [Google Scholar] [CrossRef]
- Shi, M.; Bradner, J.; Bammler, T.K.; Eaton, D.L.; Zhang, J.; Ye, Z.; Wilson, A.M.; Montine, T.J.; Pan, C.; Zhang, J. Identification of glutathione S-transferase pi as a protein involved in Parkinson disease progression. Am. J. Pathol. 2009, 175, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Piacentini, S.; Polimanti, R.; Squitti, R.; Mariani, S.; Migliore, S.; Vernieri, F.; Rossini, P.M.; Manfellotto, D.; Fuciarelli, M. GSTO1*E155del polymorphism associated with increased risk for late-onset Alzheimer’s disease: Association hypothesis for an uncommon genetic variant. Neurosci. Lett. 2012, 506, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Ercegovac, M.; Jovic, N.; Sokic, D.; Savic-Radojevic, A.; Coric, V.; Radic, T.; Nikolic, D.; Kecmanovic, M.; Matic, M.; Simic, T. GSTA1, GSTM1, GSTP1 and GSTT1 polymorphisms in progressive myoclonus epilepsy: A Serbian case-control study. Seizure 2015, 32, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eum, K.-D.; Seals, R.M.; Taylor, K.M.; Grespin, M.; Umbach, D.M.; Hu, H.; Sandler, D.P.; Kamel, F.; Weisskopf, M.G. Modification of the association between lead exposure and amyotrophic lateral sclerosis by iron and oxidative stress related gene polymorphisms. Amyotroph. Lateral Scler. Front. Degener. 2015, 16, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allocati, N.; Masulli, M.; Di Ilio, C.; Federici, L. Glutathione transferases: Substrates, inihibitors and pro-drugs in cancer and neurodegenerative diseases. Oncogenesis 2018, 7, 8. [Google Scholar] [CrossRef]
- Chartoumpekis, D.V.; Wakabayashi, N.; Kensler, T.W. Keap1/Nrf2 pathway in the frontiers of cancer and non-cancer cell metabolism. Biochem. Soc. Trans. 2015, 43, 639–644. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-Y.; Marzec, J.; Kleeberger, S.R. Functional polymorphisms in Nrf2: Implications for human disease. Free Radic. Biol. Med. 2015, 88, 362–372. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, X.; Ding, Y.; Zhou, W.; Tao, L.; Lu, P.; Wang, Y.; Hu, R. Nuclear factor E2-related factor-2 negatively regulates NLRP3 inflammasome activity by inhibiting reactive oxygen species-induced NLRP3 priming. Antioxid. Redox Signal. 2017, 26, 28–43. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An overview of Nrf2 signaling pathway and its role in inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Michielsen, H.J.; De Vries, J.; Van Heck, G.L. Psychometric qualities of a brief self-rated fatigue measure: The Fatigue Assessment Scale. J. Psychosom. Res. 2003, 54, 345–352. [Google Scholar] [CrossRef]
- Ross, A.J.; Medow, M.S.; Rowe, P.C.; Stewart, J.M. What is brain fog? An evaluation of the symptom in postural tachycardia syndrome. Clin. Auton. Res. 2013, 23, 305–311. [Google Scholar] [CrossRef]
- Shimoyama, Y.; Mitsuda, Y.; Tsuruta, Y.; Hamajima, N.; Niwa, T. Polymorphism of Nrf2, an antioxidative gene, is associated with blood pressure and cardiovascular mortality in hemodialysis patients. Int. J. Med. Sci. 2014, 11, 726. [Google Scholar] [CrossRef] [Green Version]
- Jarrott, B.; Head, R.; Pringle, K.G.; Lumbers, E.R.; Martin, J.H. “LONG COVID”—A hypothesis for understanding the biological basis and pharmacological treatment strategy. Pharmacol. Res. Perspect. 2022, 10, e00911. [Google Scholar] [CrossRef]
- Cuadrado, A.; Pajares, M.; Benito, C.; Jiménez-Villegas, J.; Escoll, M.; Fernández-Ginés, R.; Yagüe, A.J.G.; Lastra, D.; Manda, G.; Rojo, A.I. Can activation of NRF2 be a strategy against COVID-19? Trends Pharmacol. Sci. 2020, 41, 598–610. [Google Scholar] [CrossRef]
- Nalbandian, A.; Sehgal, K.; Gupta, A.; Madhavan, M.V.; McGroder, C.; Stevens, J.S.; Cook, J.R.; Nordvig, A.S.; Shalev, D.; Sehrawat, T.S. Post-acute COVID-19 syndrome. Nat. Med. 2021, 27, 601–615. [Google Scholar] [CrossRef]
- Libby, P.; Lüscher, T. COVID-19 is, in the end, an endothelial disease. Eur. Heart J. 2020, 41, 3038–3044. [Google Scholar] [CrossRef]
- Fogarty, H.; Townsend, L.; Morrin, H.; Ahmad, A.; Comerford, C.; Karampini, E.; Englert, H.; Byrne, M.; Bergin, C.; O’Sullivan, J.M. Persistent endotheliopathy in the pathogenesis of long COVID syndrome. J. Thromb. Haemost. 2021, 19, 2546–2553. [Google Scholar] [CrossRef]
- von Meijenfeldt, F.A.; Havervall, S.; Adelmeijer, J.; Lundström, A.; Magnusson, M.; Mackman, N.; Thalin, C.; Lisman, T. Sustained prothrombotic changes in COVID-19 patients 4 months after hospital discharge. Blood Adv. 2021, 5, 756–759. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, Y.; Fan, X. Microvascular pericytes in brain-associated vascular disease. Biomed. Pharmacother. 2020, 121, 109633. [Google Scholar] [CrossRef]
- Østergaard, L. SARS CoV-2 related microvascular damage and symptoms during and after COVID-19: Consequences of capillary transit-time changes, tissue hypoxia and inflammation. Physiol. Rep. 2021, 9, e14726. [Google Scholar] [CrossRef]
- Marshall, M. COVID and the brain: Researchers zero in on how damage occurs. Nature 2021, 595, 484–485. [Google Scholar] [CrossRef] [PubMed]
- Checconi, P.; De Angelis, M.; Marcocci, M.E.; Fraternale, A.; Magnani, M.; Palamara, A.T.; Nencioni, L. Redox-modulating agents in the treatment of viral infections. Int. J. Mol. Sci. 2020, 21, 4084. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Itoh, K.; Takahashi, S.; Sato, H.; Yanagawa, T.; Katoh, Y.; Bannai, S.; Yamamoto, M. Transcription factor Nrf2 coordinately regulates a group of oxidative stress-inducible genes in macrophages. J. Biol. Chem. 2000, 275, 16023–16029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippi, G.; Lavie, C.J.; Henry, B.M.; Sanchis-Gomar, F. Do genetic polymorphisms in angiotensin converting enzyme 2 (ACE2) gene play a role in coronavirus disease 2019 (COVID-19)? Clin. Chem. Lab. Med. 2020, 58, 1415–1422. [Google Scholar] [CrossRef]
- Devaux, C.A.; Rolain, J.-M.; Raoult, D. ACE2 receptor polymorphism: Susceptibility to SARS-CoV-2, hypertension, multi-organ failure, and COVID-19 disease outcome. J. Microbiol. Immunol. Infect. 2020, 53, 425–435. [Google Scholar] [CrossRef]
- Paniri, A.; Hosseini, M.M.; Akhavan-Niaki, H. First comprehensive computational analysis of functional consequences of TMPRSS2 SNPs in susceptibility to SARS-CoV-2 among different populations. J. Biomol. Struct. Dyn. 2021, 39, 3576–3593. [Google Scholar] [CrossRef]
- Suzuki, T.; Shibata, T.; Takaya, K.; Shiraishi, K.; Kohno, T.; Kunitoh, H.; Tsuta, K.; Furuta, K.; Goto, K.; Hosoda, F. Regulatory nexus of synthesis and degradation deciphers cellular Nrf2 expression levels. Mol. Cell. Biol. 2013, 33, 2402–2412. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, D.; Galli, F. The functional interactome of GSTP: A regulatory biomolecular network at the interface with the Nrf2 adaption response to oxidative stress. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1019, 29–44. [Google Scholar] [CrossRef]
- Thévenin, A.F.; Zony, C.L.; Bahnson, B.J.; Colman, R.F. GSTpi modulates JNK activity through a direct interaction with JNK substrate, ATF2. Protein Sci. 2011, 20, 834–848. [Google Scholar] [CrossRef] [Green Version]
- Board, P.G.; Menon, D. Glutathione transferases, regulators of cellular metabolism and physiology. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2013, 1830, 3267–3288. [Google Scholar] [CrossRef] [PubMed]
- Rex, D.A.B.; Dagamajalu, S.; Kandasamy, R.K.; Raju, R.; Prasad, T.S. SARS-CoV-2 signaling pathway map: A functional landscape of molecular mechanisms in COVID-19. J. Cell Commun. Signal. 2021, 15, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Bartolini, D.; Commodi, J.; Piroddi, M.; Incipini, L.; Sancineto, L.; Santi, C.; Galli, F. Glutathione S-transferase pi expression regulates the Nrf2-dependent response to hormetic diselenides. Free Radic. Biol. Med. 2015, 88, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Wong, J.; Henry, B.M. Myalgia may not be associated with severity of coronavirus disease 2019 (COVID-19). World J. Emerg. Med. 2020, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Seixas, M.L.G.A.; Mitre, L.P.; Shams, S.; Lanzuolo, G.B.; Bartolomeo, C.S.; Silva, E.A.; Prado, C.M.; Ureshino, R.; Stilhano, R.S. Unraveling Muscle Impairment Associated With COVID-19 and the Role of 3D Culture in Its Investigation. Front. Nutr. 2022, 9, 115. [Google Scholar] [CrossRef] [PubMed]
- Board, P.G.; Coggan, M.; Chelvanayagam, G.; Easteal, S.; Jermiin, L.S.; Schulte, G.K.; Danley, D.E.; Hoth, L.R.; Griffor, M.C.; Kamath, A.V. Identification, characterization, and crystal structure of the Omega class glutathione transferases. J. Biol. Chem. 2000, 275, 24798–24806. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.M.; Hooftman, A.; Angiari, S.; Tummala, P.; Zaslona, Z.; Runtsch, M.C.; McGettrick, A.F.; Sutton, C.E.; Diskin, C.; Rooke, M. Glutathione transferase omega-1 regulates NLRP3 inflammasome activation through NEK7 deglutathionylation. Cell Rep. 2019, 29, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Di, B.; Xu, L. The NLRP3 inflammasome and COVID-19: Activation, pathogenesis and therapeutic strategies. Cytokine Growth Factor Rev. 2021, 61, 2–15. [Google Scholar] [CrossRef]
- McKee, C.M.; Coll, R.C. NLRP3 inflammasome priming: A riddle wrapped in a mystery inside an enigma. J. Leukoc. Biol. 2020, 108, 937–952. [Google Scholar] [CrossRef]
- Tanaka-Kagawa, T.; Jinno, H.; Hasegawa, T.; Makino, Y.; Seko, Y.; Hanioka, N.; Ando, M. Functional characterization of two variant human GSTO 1-1s (Ala140Asp and Thr217Asn). Biochem. Biophys. Res. Commun. 2003, 301, 516–520. [Google Scholar] [CrossRef]
- Phetsouphanh, C.; Darley, D.R.; Wilson, D.B.; Howe, A.; Munier, C.; Patel, S.K.; Juno, J.A.; Burrell, L.M.; Kent, S.J.; Dore, G.J. Immunological dysfunction persists for 8 months following initial mild-to-moderate SARS-CoV-2 infection. Nat. Immunol. 2022, 23, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Ekoue, D.N.; He, C.; Diamond, A.M.; Bonini, M.G. Manganese superoxide dismutase and glutathione peroxidase-1 contribute to the rise and fall of mitochondrial reactive oxygen species which drive oncogenesis. Biochim. Biophys. Acta (BBA) -Bioenerg. 2017, 1858, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, E.-K.; Kang, D.H.; Lee, J.; Hong, S.H.; Jeong, W.; Kang, S.W. Glutathione peroxidase-1 regulates ASK1-dependent apoptosis via interaction with TRAF2 in RIPK3-negative cancer cells. Exp. Mol. Med. 2021, 53, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Hsu, A.C.Y.; Wang, G.; Reid, A.T.; Veerati, P.C.; Pathinayake, P.S.; Daly, K.; Mayall, J.R.; Hansbro, P.M.; Horvat, J.C.; Wang, F. SARS-CoV-2 Spike protein promotes hyper-inflammatory response that can be ameliorated by Spike-antagonistic peptide and FDA-approved ER stress and MAP kinase inhibitors in vitro. Biorxiv 2020. [Google Scholar] [CrossRef]
- Notz, Q.; Herrmann, J.; Schlesinger, T.; Helmer, P.; Sudowe, S.; Sun, Q.; Hackler, J.; Roeder, D.; Lotz, C.; Meybohm, P. Clinical significance of micronutrient supplementation in critically ill COVID-19 patients with severe ARDS. Nutrients 2021, 13, 2113. [Google Scholar] [CrossRef]
- Blunt, K.; Jacobse, J.; Allaman, M.; Washington, M.; Goettel, J.; Wilson, K.; Williams, C.; Short, S. GPX3 IS REQUIRED TO MEDIATE PROTECTIVE EFFECTS OF GPX1 LOSS IN COLITIS. Gastroenterology 2022, 162, S63. [Google Scholar] [CrossRef]
- Moghaddam, A.; Heller, R.A.; Sun, Q.; Seelig, J.; Cherkezov, A.; Seibert, L.; Hackler, J.; Seemann, P.; Diegmann, J.; Pilz, M. Selenium deficiency is associated with mortality risk from COVID-19. Nutrients 2020, 12, 2098. [Google Scholar] [CrossRef]
- Heller, R.A.; Sun, Q.; Hackler, J.; Seelig, J.; Seibert, L.; Cherkezov, A.; Minich, W.B.; Seemann, P.; Diegmann, J.; Pilz, M. Prediction of survival odds in COVID-19 by zinc, age and selenoprotein P as composite biomarker. Redox Biol. 2021, 38, 101764. [Google Scholar] [CrossRef]
- Mihailovic, S.; Coric, V.; Radic, T.; Radojevic, A.S.; Matic, M.; Dragicevic, D.; Djokic, M.; Vasic, V.; Dzamic, Z.; Simic, T. The Association of Polymorphisms in Nrf2 and Genes Involved in Redox Homeostasis in the Development and Progression of Clear Cell Renal Cell Carcinoma. Oxid. Med. Cell. Longev. 2021, 2021, 6617969. [Google Scholar] [CrossRef]
- Wu, W.; Peden, D.; Diaz-Sanchez, D. Role of GSTM1 in resistance to lung inflammation. Free Radic. Biol. Med. 2012, 53, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Niemi, M.E.K.; Karjalainen, J.; Liao, R.G.; Neale, B.M.; Daly, M.; Ganna, A.; Pathak, G.A.; Andrews, S.J.; Kanai, M.; Veerapen, K. Mapping the human genetic architecture of COVID-19. Nature 2021, 600, 472–477. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COVID-19 Patients | |
|---|---|
| Age (years) a | 55.9 ± 12.3 |
| Gender, n (%) | |
| Male | 100 (60) |
| Female | 67 (40) |
| BMI (kg/m2) a | 29.0 ± 5.0 |
| Smoking, n (%) | |
| Never | 84 (53) |
| Former | 57 (36) |
| Ever | 17 (11) |
| Hypertension, n (%) | |
| No | 109 (65) |
| Yes | 58 (35) |
| Diabetes, n (%) | |
| No | 149 (89) |
| Yes | 18 (11) |
| Febricity, n (%) | |
| No | 23 (14) |
| Yes | 144 (86) |
| Febricity over 38 °C, n (%) | |
| No | 85 (51) |
| Yes | 82 (49) |
| Hospitalization, n (%) | |
| No | 24 (14) |
| Yes | 143 (86) |
| Pneumonia, n (%) | |
| No | 17 (10) |
| Yes | 150 (90) |
| O2 support, n (%) | |
| No | 103 (62) |
| Yes | 64 (38) |
| ACUTE COVID-19 | COVID-19 Patients |
|---|---|
| Loss of smell, n (%) | |
| No | 115 (69) |
| Yes | 52 (31) |
| Loss of taste, n (%) | |
| No | 119 (71) |
| Yes | 48 (29) |
| Myalgia, n (%) | |
| No | 94 (56) |
| Yes | 73 (44) |
| Weakness, n (%) | |
| No | 16 (10) |
| Yes | 151 (90) |
| General malaise, n (%) | |
| No | 15 (9) |
| Yes | 152 (91) |
| Headache, n (%) | |
| No | 91 (54) |
| Yes | 76 (46) |
| Headache with fever, n (%) | |
| No | 101 (60) |
| Yes | 66 (40) |
| LONG-COVID | COVID-19 Patients |
|---|---|
| Fatigue, n (%) | |
| No | 33 (20) |
| Yes | 134 (80) |
| Myalgia, n (%) | |
| No | 121 (72) |
| Yes | 46 (28) |
| “Brain fog”, n (%) | |
| No | 146 (87) |
| Yes | 21 (13) |
| Instability, n (%) | |
| No | 152 (91) |
| Yes | 15 (9) |
| Paresthesia, n (%) | |
| No | 150 (90) |
| Yes | 17 (10) |
| Variants | Genotype, n (%) | Variants | Genotype, n (%) |
|---|---|---|---|
| GSTM1 | GSTT1 | ||
| active a | 65 (39) | active a | 131 (79) |
| null b | 100 (61) | null b | 34 (21) |
| GSTM3 (rs1332018) | GSTA1 (rs3957357) | ||
| AA | 61 (37) | CC | 52 (37) |
| AC | 51 (31) | CT | 66 (46) |
| CC | 53 (32) | TT | 24 (17) |
| GSTP1 (rs1695) | GSTP1 (rs1138272) | ||
| IleIle | 82 (49) | AlaAla | 129 (78) |
| IleVal | 66 (40) | AlaVal | 36 (21) |
| ValVal | 18 (11) | ValVal | 1 (1) |
| GSTO1 (rs4925) | GSTO2 (rs156697) | ||
| AlaAla | 79 (48) | AsnAsn | 79 (47) |
| AlaAsp | 48 (29) | AsnAsp | 46 (28) |
| AspAsp | 38 (23) | AspAsp | 41 (25) |
| Nrf2 (rs672196) | SOD2 (rs4880) | ||
| CC | 124 (75) | AlaAla | 39 (23) |
| CA | 38 (23) | AlaVal | 89 (54) |
| AA | 3 (2) | ValVal | 39 (23) |
| GPX1 (rs1050450) | GPX3 (rs8177412) | ||
| ProPro | 75 (45) | TT | 126 (75) |
| ProLeu | 71 (43) | TC | 39 (24) |
| LeuLeu | 20 (12) | CC | 2 (1) |
| ACE2 (rs4646116) | ACE2 (rs143936283) | ||
| LysLys | 154 (94) | GluGlu | 37 (23) |
| LysArg | 9 (5) | GluGly | 126 (77) |
| ArgArg | 1 (1) | GlyGly | 0 (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ercegovac, M.; Asanin, M.; Savic-Radojevic, A.; Ranin, J.; Matic, M.; Djukic, T.; Coric, V.; Jerotic, D.; Todorovic, N.; Milosevic, I.; et al. Antioxidant Genetic Profile Modifies Probability of Developing Neurological Sequelae in Long-COVID. Antioxidants 2022, 11, 954. https://doi.org/10.3390/antiox11050954
Ercegovac M, Asanin M, Savic-Radojevic A, Ranin J, Matic M, Djukic T, Coric V, Jerotic D, Todorovic N, Milosevic I, et al. Antioxidant Genetic Profile Modifies Probability of Developing Neurological Sequelae in Long-COVID. Antioxidants. 2022; 11(5):954. https://doi.org/10.3390/antiox11050954
Chicago/Turabian StyleErcegovac, Marko, Milika Asanin, Ana Savic-Radojevic, Jovan Ranin, Marija Matic, Tatjana Djukic, Vesna Coric, Djurdja Jerotic, Nevena Todorovic, Ivana Milosevic, and et al. 2022. "Antioxidant Genetic Profile Modifies Probability of Developing Neurological Sequelae in Long-COVID" Antioxidants 11, no. 5: 954. https://doi.org/10.3390/antiox11050954
APA StyleErcegovac, M., Asanin, M., Savic-Radojevic, A., Ranin, J., Matic, M., Djukic, T., Coric, V., Jerotic, D., Todorovic, N., Milosevic, I., Stevanovic, G., Simic, T., Bukumiric, Z., & Pljesa-Ercegovac, M. (2022). Antioxidant Genetic Profile Modifies Probability of Developing Neurological Sequelae in Long-COVID. Antioxidants, 11(5), 954. https://doi.org/10.3390/antiox11050954