
Evolution of Heme Peroxygenases: Ancient Roots and Later Evolved Branches



Abstract
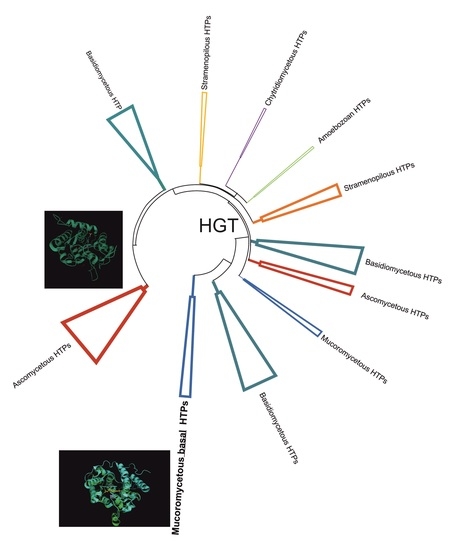
:
1. Introduction
2. Materials and Methods
2.1. Retrieval of Protein Sequences from Databases
2.2. Multiple Sequence Alignment
2.3. Reconstruction of Evolutionary Relationships
2.4. Analysis of Transcription Regulatory Regions of HTP Genes
2.5. Comparison of 3D Structures of Various Heme Peroxygenases
3. Results
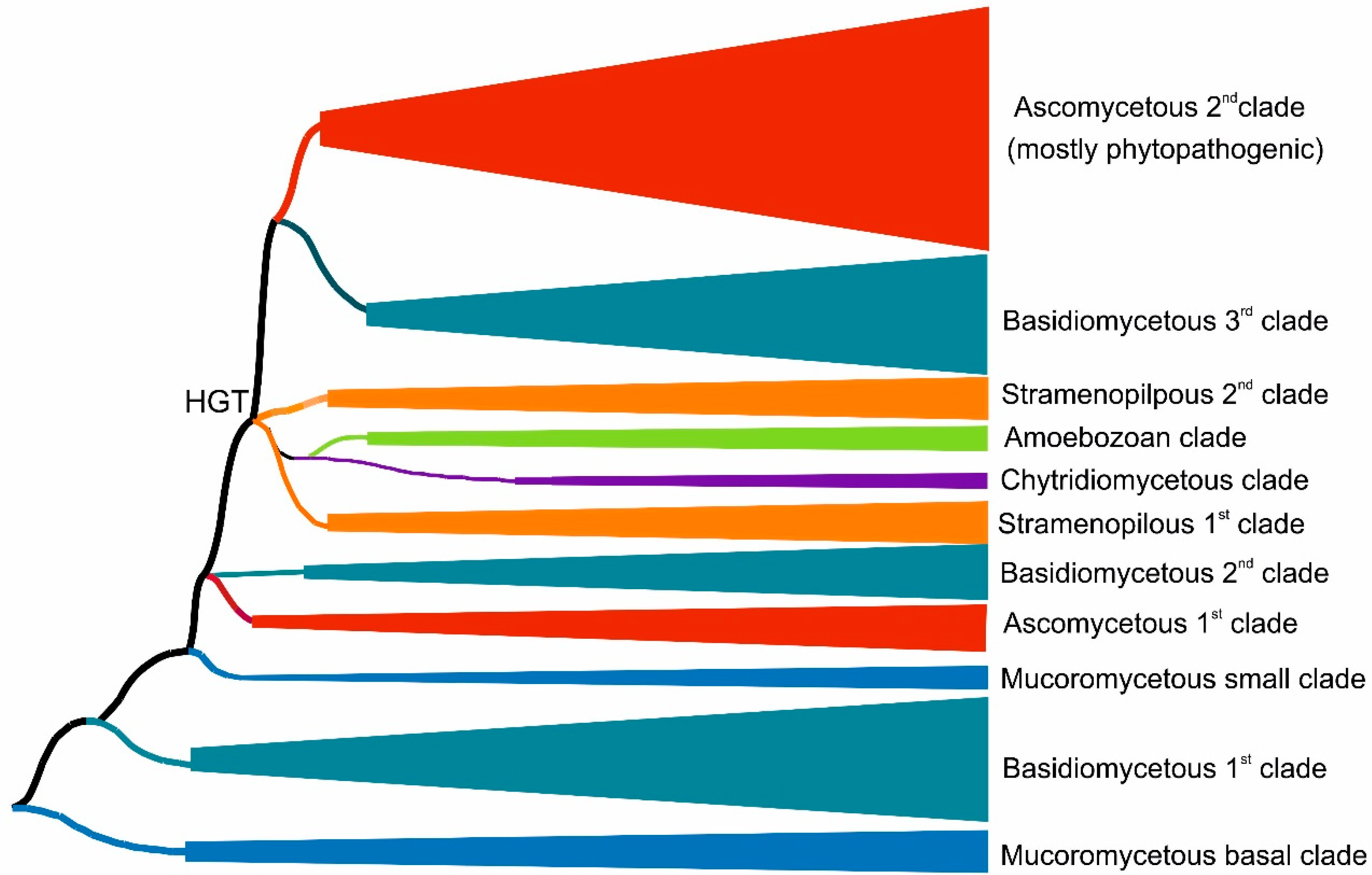
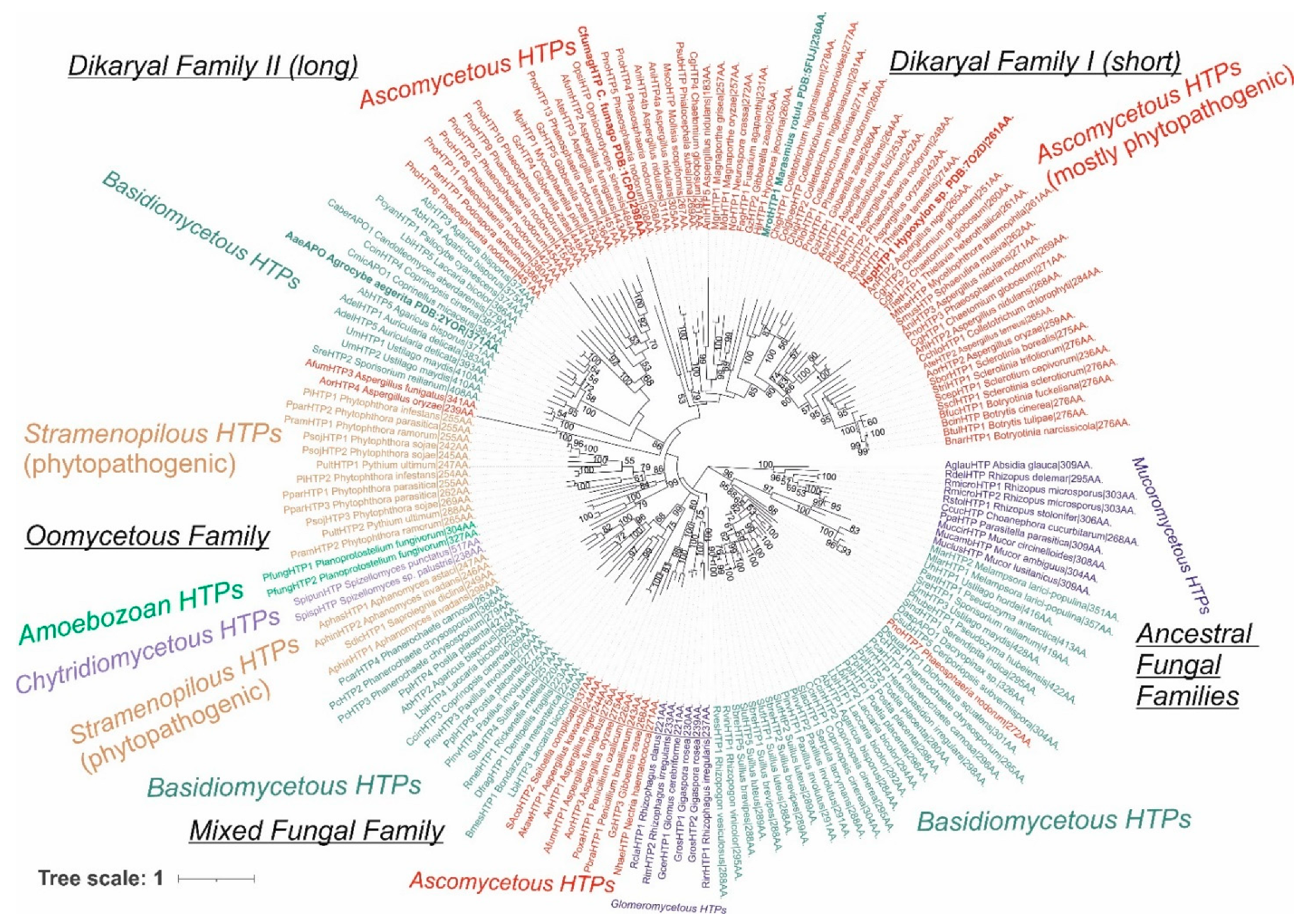
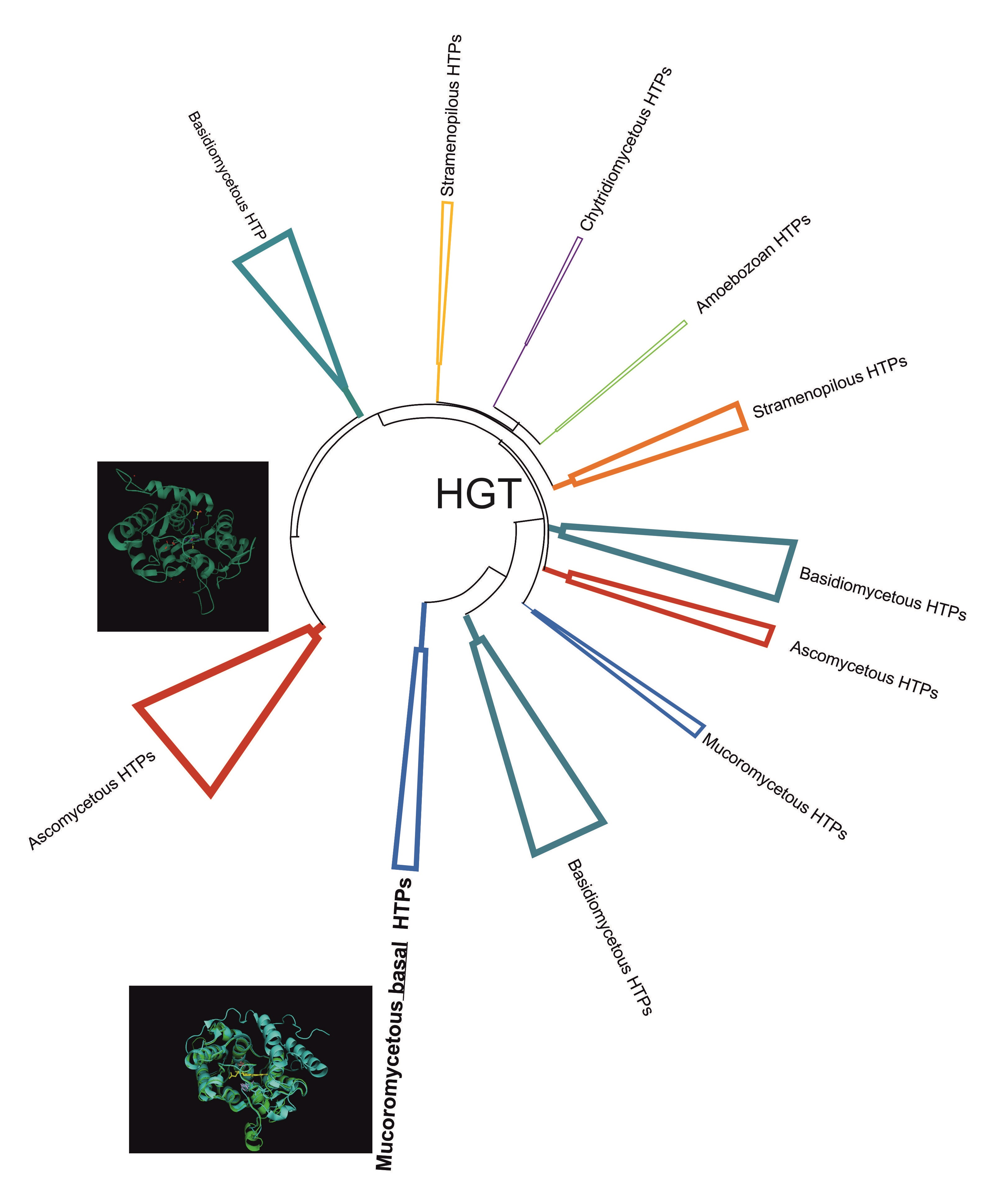
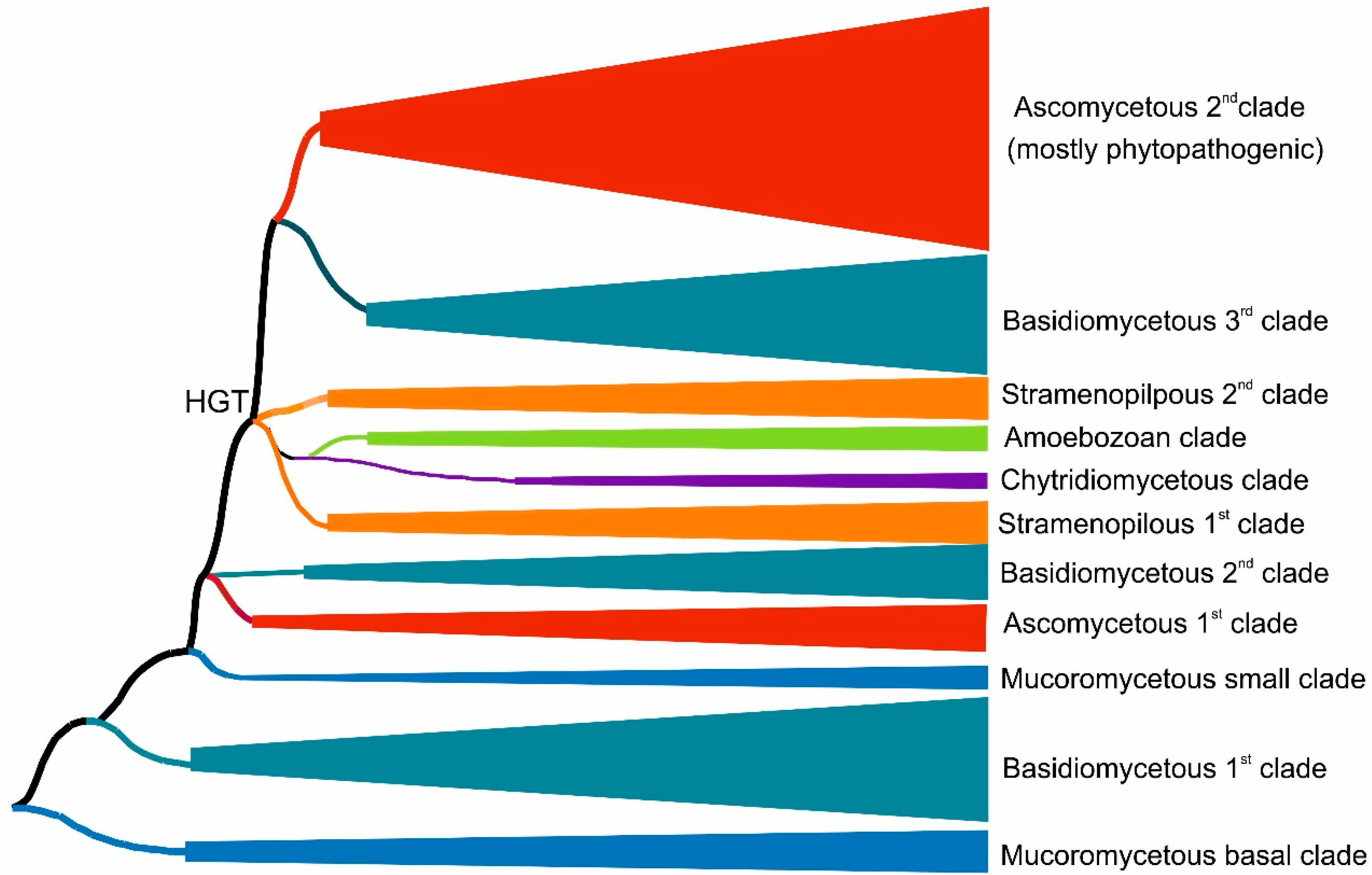
3.1. Reconstructed Phylogenetic Tree of the Peroxidase–Peroxygenase Superfamily
3.1.1. Fungal Representatives of Heme Peroxygenases
3.1.2. Non-Fungal Representatives of Heme Peroxygenases and Their Phylogenetic Position
3.2. Prediction of Signal Sequences in Heme Peroxygenases
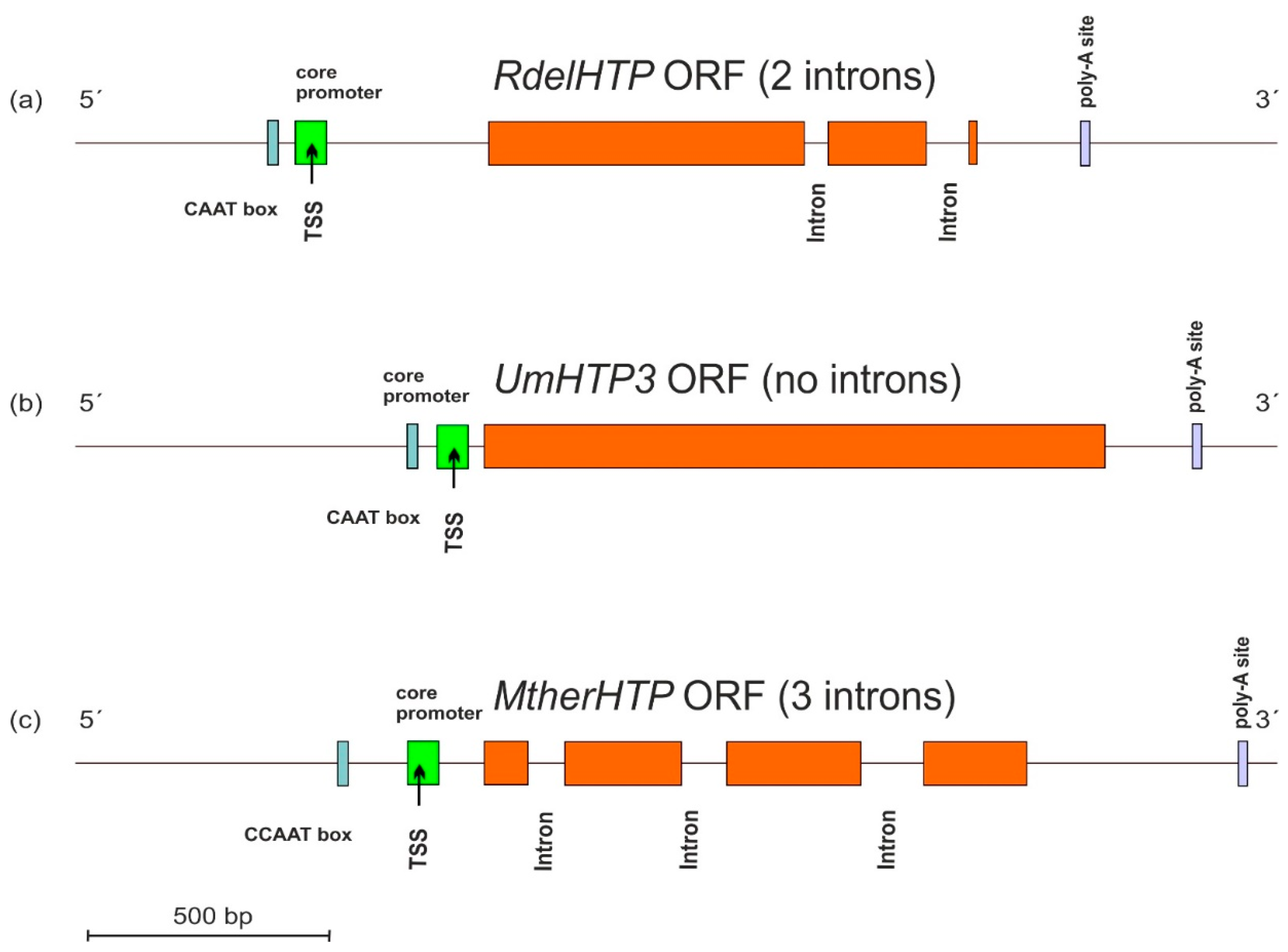
3.3. Gene Architecture of HTP Genes
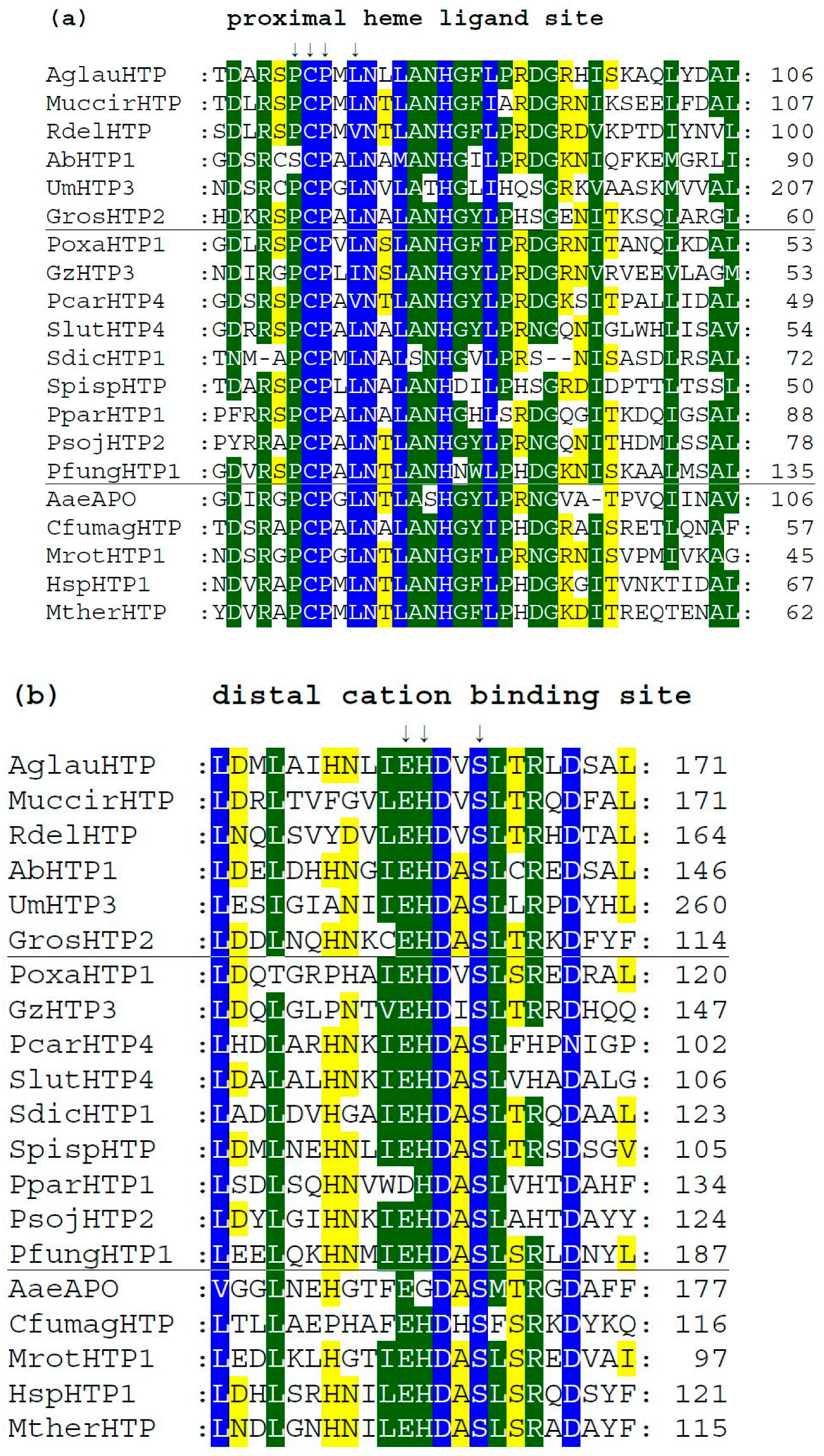
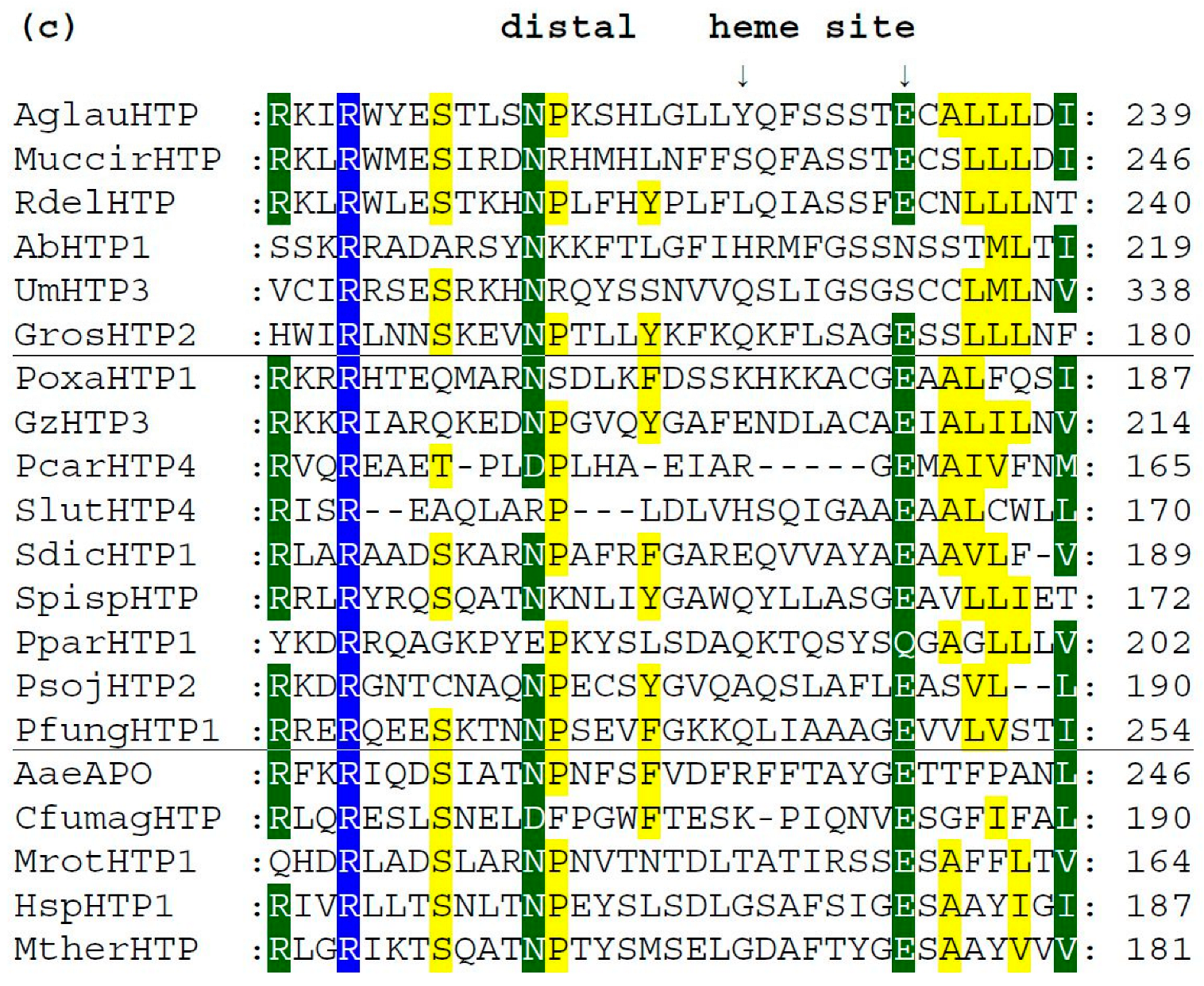
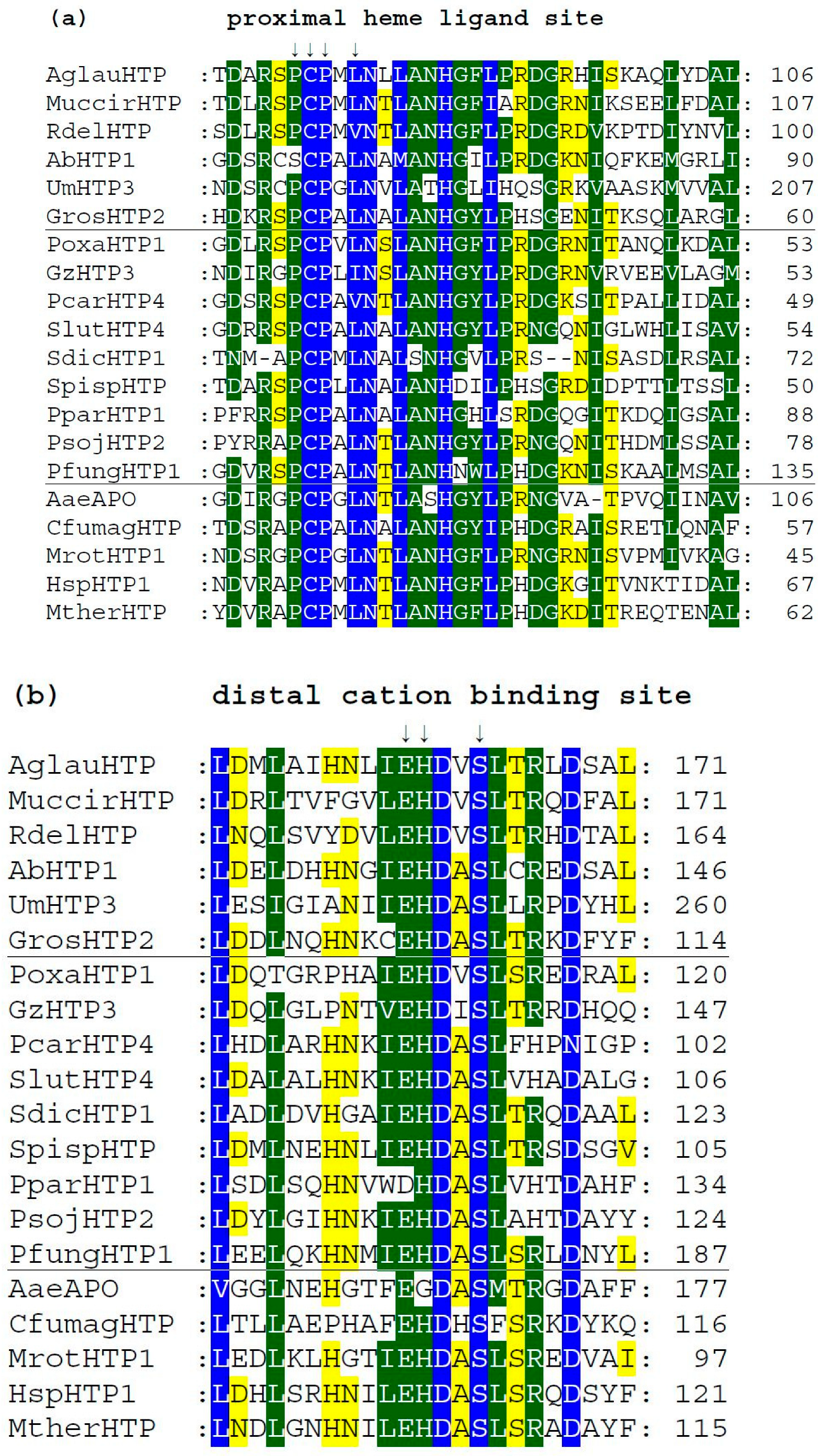
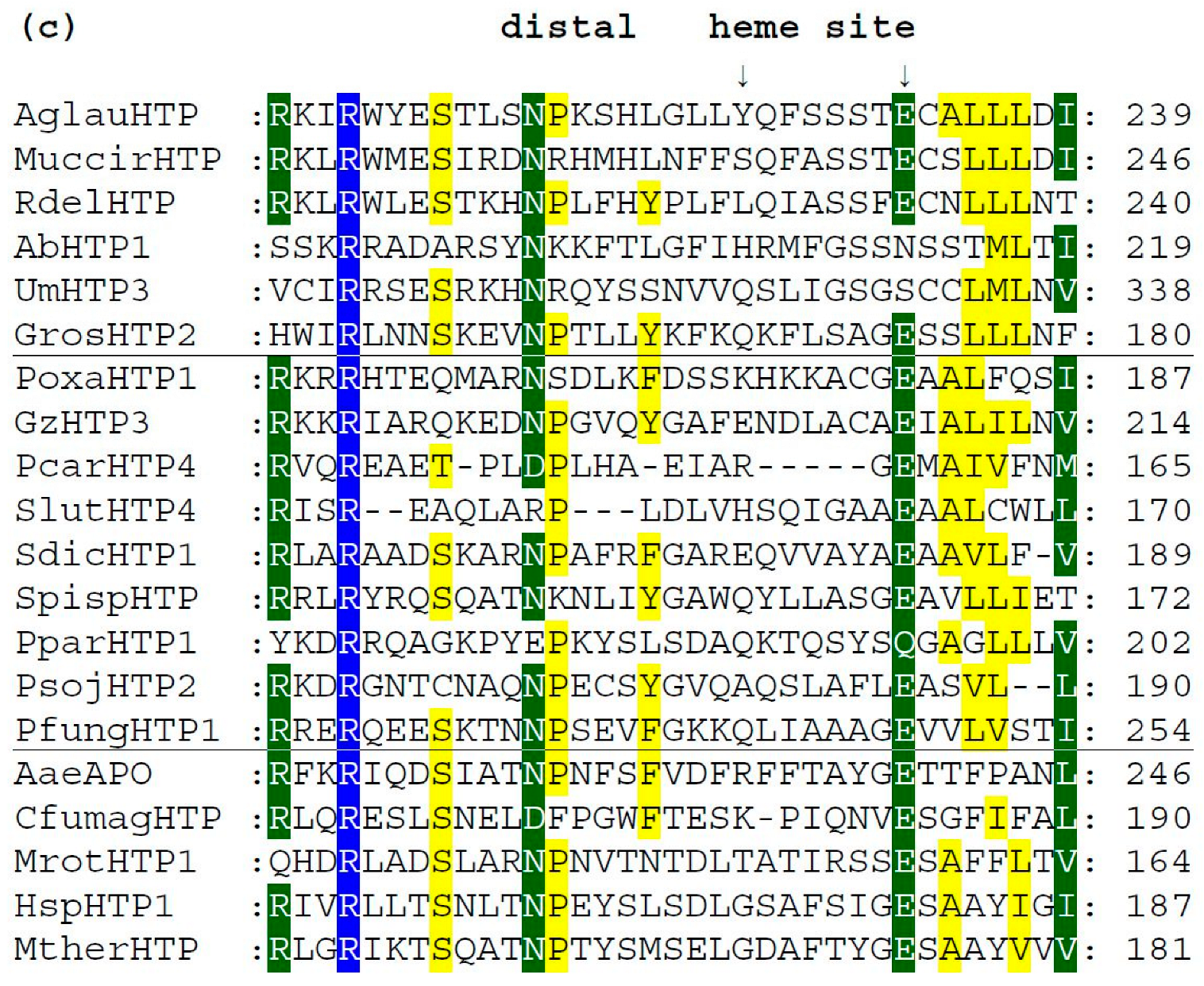
3.4. Conserved Protein Sequence Motifs in Aligned Heme Peroxygenases
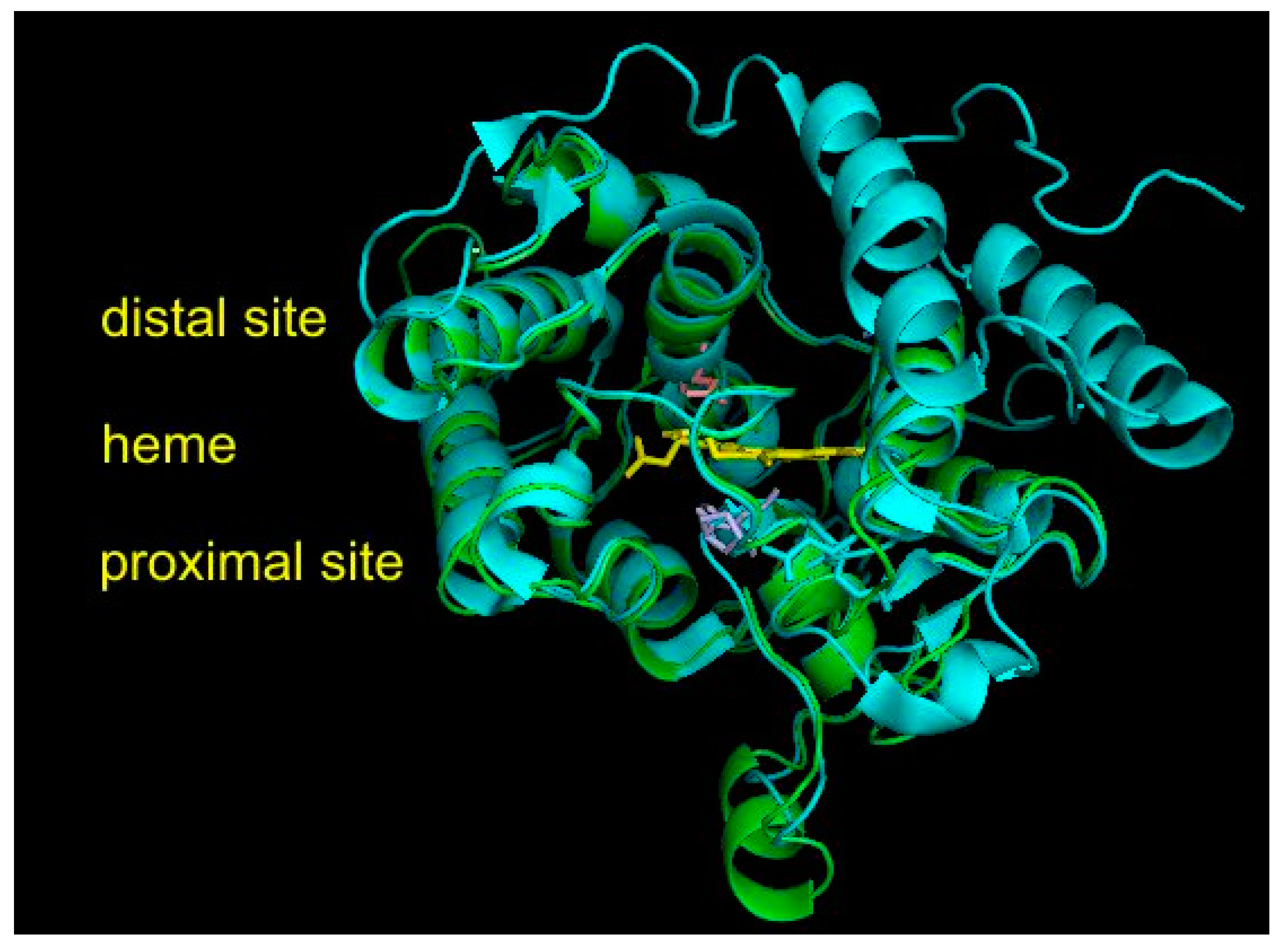
3.5. Structural Comparisons of Conserved Protein Architecture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kinner, A.; Rosenthal, K.; Lütz, S. Identification and Expression of New Unspecific Peroxygenases—Recent Advances, Challenges and Opportunities. Front. Bioeng. Biotechnol. 2021, 9, 705630. [Google Scholar] [CrossRef] [PubMed]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.; Liers, C.; Lundell, T. New and classical families of secreted fungal heme peroxidases. Appl. Microbiol. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef] [PubMed]
- Hofrichter, M.; Ullrich, R. Oxidations catalyzed by fungal peroxygenases. Curr. Opinion Chem. Biol. 2014, 19, 116–125. [Google Scholar] [CrossRef]
- Hiner, A.N.P.; Raven, E.L.; Thorneley, R.N.F.; García-Cánovas, F.; Rodriguéz-López, J.N. Mechanisms of Compound I Formation in Heme Peroxidases. J. Inorg. Biochem. 2002, 91, 27–34. [Google Scholar] [CrossRef]
- Zámocký, M.; Hofbauer, S.; Schaffner, I.; Gasselhuber, B.; Nicolussi, A.; Soudi, M.; Pirker, K.F.; Furtmüller, P.G.; Obinger, C. Independent evolution of four heme peroxidase superfamilies. Arch. Biochem. Biophys. 2015, 574, 108–119. [Google Scholar] [CrossRef]
- Zámocký, M.; Jakopitsch, C.; Furtmüller, P.G.; Dunand, C.; Obinger, C. The Peroxidase-Cyclooxygenase Superfamily: Reconstructed Evolution of Critical Enzymes of the Innate Immune System. Proteins 2008, 72, 589–605. [Google Scholar] [CrossRef]
- Karich, A.; Scheibner, K.; Ulrich, R.; Hofrichter, M. Exploring the Catalase Activity of Unspecific Peroxygenases and the Mechanism of Peroxide-dependent Heme Destruction. J. Mol. Catal. B Enzym. 2016, 134, 238–246. [Google Scholar] [CrossRef]
- Chovanová, K.; Böhmer, M.; Poljovka, A.; Budiš, J.; Harichová, J.; Szemeš, T.; Zámocký, M. Parallel Molecular Evolution of Catalases and Superoxide Dismutases—Focus on Thermophilic Fungal Genomes. Antioxidants 2020, 9, 1047. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Savelli, B.; Li, Q.; Webber, M.; Jemmat, A.M.; Robitaille, A.; Zámocký, M.; Mathe, C.; Dunand, C. RedoxiBase: A Database for ROS Homeostasis Regulated Proteins. Redox Biol. 2019, 26, 101247. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Eolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Armenteros, J.J.A.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 6.0 Predicts all Five Types of Signal Peptides Using Protein Language Models. Nat. Biotechnol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Le, S.Q.; Gascuel, O. An Improved General Amino Acid Replacement Matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucl. Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Solovyev, V.V.; Kosarev, P.; Seledsov, I.; Vorobyev, D. Automatic annotation of eukaryotic genes, pseudogenes and promoters. Genome Biol. 2006, 7, S10. [Google Scholar] [CrossRef] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction, and Analysis. Nat. Prot. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Sundaramoorthy, M.; Terner, J.; Poulos, T.L. The crystal structure of chloroperoxidase: A heme peroxidase—Cytochrome P450 functional hybrid. Structure 1995, 3, 1367–1377. [Google Scholar] [CrossRef] [Green Version]
- Piontek, K.; Ullrich, R.; Liers, C.; Diederichs, K.; Plattner, D.A.; Hofrichter, M. Crystallization of a 45 kDa Peroxygenase/Peroxidase from the Mushroom Agrocybe Aegerita and Structure Determination by Sad Utilizing Only the Haem Iron. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 693–698. [Google Scholar] [CrossRef] [Green Version]
- Piontek, K.; Strittmatter, E.; Ullrich, R.; Grobe, G.; Pecyna, M.J.; Kluge, M.; Scheibner, K.; Hofrichter, M.; Plattner, D.A. Structural Basis of Substrate Conversion in a New Aromatic Peroxygenase: P450 Functionality with Benefits. J. Biol. Chem. 2013, 288, 34767. [Google Scholar] [CrossRef] [Green Version]
- Rotilio, L.; Swoboda, A.; Ebner, K.; Rinnofner, C.; Glieder, A.; Kroutil, W.; Mattevi, A. Structural and biochemical studies enlighten the unspecific peroxygenase from Hypoxylon sp. EC38 as an efficient oxidative biocatalyst. ACS Catal. 2021, 11, 11511–11525. [Google Scholar] [CrossRef] [PubMed]
- Naranjo-Ortiz, M.; Gabaldón, T. Fungal Evolution: Diversity, Taxonomy and Phylogeny of the Fungi. Biol. Rev. 2019, 94, 2101–2137. [Google Scholar] [CrossRef]
- Berbee, M.L.; James, T.Y.; Strullu-Derrien, C. Early Diverging Fungi: Diversity and Impact at the Dawn of Terrestrial Life. Ann. Rev. Microbiol. 2017, 71, 41–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, P.T.; Probst, A.J.; Grigoriev, I.V.; Thomas, B.C.; Banfield, J.F. Genome-reconstruction for Eukaryotes from Complex Natural Microbial Communities. Genome Res. 2018, 28, 569–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lastovetsky, O.A.; Gaspar, M.L.; Mondo, S.J.; LaButti, K.M.; Sandor, L.; Grigoriev, I.V.; Henry, S.A.; Pawlowska, T.E. Lipid Metabolic Changes in an Early Divergent Fungus govern the Establishment of a Mutualistic Symbiosis with Endobacteria. Proc. Natl. Acad. Sci. USA 2016, 113, 15102–15107. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Wang, S.; Sekimoto, S.; Aerts, A.L.; Choi, C.; Clum, A.; LaButti, K.M.; Lindquist, E.A.; Ngan, C.Y.; Ohm, R.A.; et al. Phylogenomic Analyses Indicate that Early Fungi Evolved Digesting Cell Walls of Algal Ancestors of Land Plants. Genome Biol. Evol. 2015, 7, 1590–1601. [Google Scholar] [CrossRef] [PubMed]
- Pambulka, G.T.; Kinge, T.R.; Ghosh, S.; Cason, E.D.; Nyaga, M.M.; Gryzenhout, M. Baseline Data of the Fungal Phytobiome of Three Sorghum (Sorghum bicolor) Cultivars in South Africa using Targeted Environmental Sequencing. J. Fungi 2021, 7, 978. [Google Scholar] [CrossRef]
- Hofrichter, M.; Kellner, H.; Herzog, R.; Karich, A.; Kiebist, J.; Scheibner, K.; Ullrich, R. Peroxide-Mediated Oxygenation of Organic Compounds by Fungal Peroxygenases. Antioxidants 2022, 11, 163. [Google Scholar] [CrossRef]
- Meng, Y.; Zhang, Q.; Ding, W.; Shan, W. Phytophtora parasitica: A Model Oomycete Plant Pathogen. Mycology 2014, 5, 43–51. [Google Scholar] [CrossRef]
- Fernández-Benéitez, M.J.; Ortiz-Santaliestra, M.E.; Lizana, M.; Diéguez-Uribeondo, J. Saprolegnia diclina: Another Species Responsible for the Emergent Disease “Saprolegnia infections” in Amphibians. FEMS Microbiol. Lett. 2008, 279, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Jiao, F. Effectors of Phytophtora Pathogens are Powerful Weapons for Manipulating Host Immunity. Planta 2019, 250, 413–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillmann, F.; Forbes, G.; Novohradska, S.; Ferling, I.; Riege, K.; Groth, M.; Glockner, G. Multiple Roots of Fruiting Body Formation in Amoebozoa. Genome Biol. Evol. 2018, 10, 591–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radosa, S.; Sprague, J.L.; Lau, S.-H.; Toth, R.; Linde, J.; Kruger, T.; Sprenger, T.; Kasper, L.; Westermann, M.; Kniemeyer, O.; et al. The Fungivorous Amoeba Protostelium aurantium Targets Redox Homeostasis and Cell Wall Integrity during Intracellular Killing of Candida parapsilosis. Cell. Microbiol. 2021, 23, e13389. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ruiz, M.I.; Ayuso-Fernández, I.; Rencoret, J.; González-Ramírez, A.M.; Linde, D.; Davó-Siguero, I.; Romero, A.; Gutiérrez, A.; Martínez, A.T.; Ruiz-Dueñas, F.J. Agaricales Mushroom Lignin Peroxidase: From Structure–Function to Degradative Capabilities. Antioxidants 2021, 10, 1446. [Google Scholar] [CrossRef]
- Zámocký, M.; Musil, M.; Danchenko, M.; Ferianc, P.; Chovanová, K.; Baráth, P.; Poljovka, A.; Bednář, D. Deep Insights into the Specific Evolution of Fungal Hybrid B Heme Peroxidases. Biology 2022, 11, 459. [Google Scholar] [CrossRef]
- Faiza, M.; Huang, S.; Lan, D.; Wang, Y. New Insights on Unspecific Peroxygenases: Superfamily Reclassification and Evolution. BMC Evol. Biol. 2019, 19, 76. [Google Scholar] [CrossRef] [Green Version]
- Faiza, M.; Lan, D.; Huang, S.; Wang, Y. UPObase: An Online Database of Unspecific Peroxygenases. Database 2019, 2019, baz122. [Google Scholar] [CrossRef] [Green Version]
- Hofrichter, M.; Kellner, H.; Herzog, R.; Karich, A.; Liers, C.; Scheibner, K.; Kimani, V.W.; Ullrich, R. Fungal Peroxygenases: A Phylogenetically Old Superfamily of Heme Enzymes with Promiscuity for Oxygen Transfer Reactions. In Grand Challenges in Fungal Biotechnology; Nevalainen, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 369–403. [Google Scholar]
- Döhlemann, G.; Hemetsberger, C. Apoplastic Immunity and its Suppression by Filamentous Plant Pathogens. New Phytol. 2013, 198, 1001–1016. [Google Scholar] [CrossRef]
- Zámocký, M.; Kamlárová, A.; Maresch, D.; Chovanová, K.; Harichová, J.; Furtmüller, P.G. Hybrid Heme Peroxidases from Rice Blast Fungus Magnaporthe oryzae Involved in Defence against Oxidative Stress. Antioxidants 2020, 9, 655. [Google Scholar] [CrossRef]
- Rich, M.K.; Nouri, E.; Courty, P.-E.; Reinhardt, D. Diet of Arbuscular Mycorrhizal Fungi: Bread and Butter? Trends Plant Sci. 2017, 22, 652–660. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Abbreviation 1 | Signal Sequence Length (AA) | Prediction Probability |
|---|---|---|
| AbHTP1 (B) | 0 | |
| AbHTP2 (B) | 0 | |
| AbHTP3 (B) | 20 | 0.9781 |
| AbHTP4 (B) | 20 | 0.9806 |
| AbHTP5 (B) | 20 | 0.9879 |
| BcinHTP (A) | 19 | 0.7056 |
| CaberAPO1 (B) | 24 | 0.9767 |
| CcinHTP1 (B) | 0 | |
| CcinHTP2 (B) | 0 | |
| CcinHTP3 (B) | 0 | |
| CcinHTP4 (B) | 25 | 0.9377 |
| CgHTP2 (A) | 17 | 0.9762 |
| DaspAPO1 (B) | 0 | |
| FagHTP1 (A) | 17 | 0.9763 |
| GcerHTP1 (G) | 0 | |
| GzHTP1 (A) | 18 | 0.9795 |
| GzHTP2 (A) | 14 | 0.9708 |
| GzHTP3 (A) | 0 | |
| GzHTP4 (A) | 0 | |
| GzHTP5 (A) | 20 | 0.9917 |
| HjHTP1 (A) | 19 | 0.7098 |
| MgrHTP1 (A) | 24 | 0.5823 |
| MoHTP1 (A) | 24 | 0.6147 |
| NhaeHTP (A) | 0 | |
| PcyanHTP1 (B) | 20 | 0.9847 |
| PfungHTP1 (Am) | 0 | |
| PfungHTP2 (Am) | 0 | |
| PficHTP1 (A) | 18 | 0.8108 |
| PparHTP1 (S) | 24 | 0.9815 |
| PparHTP2 (S) | 17 | 0.9860 |
| PparHTP3 (S) | 0 | 0.5008 |
| PramHTP1 (S) | 17 | 0.9752 |
| RdelHTP (M) | 39 | 0.8967 |
| RmicroHTP1 (M) | 0 | |
| RstolHTP (M) | 0 | |
| SborHTP1 (A) | 17 | 0.9894 |
| SsclHTP1 (A) | 19 | 0.6837 |
| SpipunHTP (C) | 0 | |
| ThetHTP1 (A) | 17 | 0.9738 |
| UmHTP1 (B) | 22 | 0.9940 |
| UmHTP2 (B) | 22 | 0.9834 |
| UmHTP3 (B) | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zámocký, M.; Harichová, J. Evolution of Heme Peroxygenases: Ancient Roots and Later Evolved Branches. Antioxidants 2022, 11, 1011. https://doi.org/10.3390/antiox11051011
Zámocký M, Harichová J. Evolution of Heme Peroxygenases: Ancient Roots and Later Evolved Branches. Antioxidants. 2022; 11(5):1011. https://doi.org/10.3390/antiox11051011
Chicago/Turabian StyleZámocký, Marcel, and Jana Harichová. 2022. "Evolution of Heme Peroxygenases: Ancient Roots and Later Evolved Branches" Antioxidants 11, no. 5: 1011. https://doi.org/10.3390/antiox11051011
APA StyleZámocký, M., & Harichová, J. (2022). Evolution of Heme Peroxygenases: Ancient Roots and Later Evolved Branches. Antioxidants, 11(5), 1011. https://doi.org/10.3390/antiox11051011