
Membrane-Free Stem Cells and Pyridoxal 5′-Phosphate Synergistically Enhance Cognitive Function in Alzheimer’s Disease Mouse Model

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of MFSCE
2.2. Animals and Experimental Protocols
2.3. Aβ25–35-Infused Mouse Model
2.4. Behavioral Tests
2.4.1. T-Maze Test
2.4.2. NOR Test
2.4.3. MWM Test
2.5. Measurement of MDA Generation
2.6. Measurement of NO Generation
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
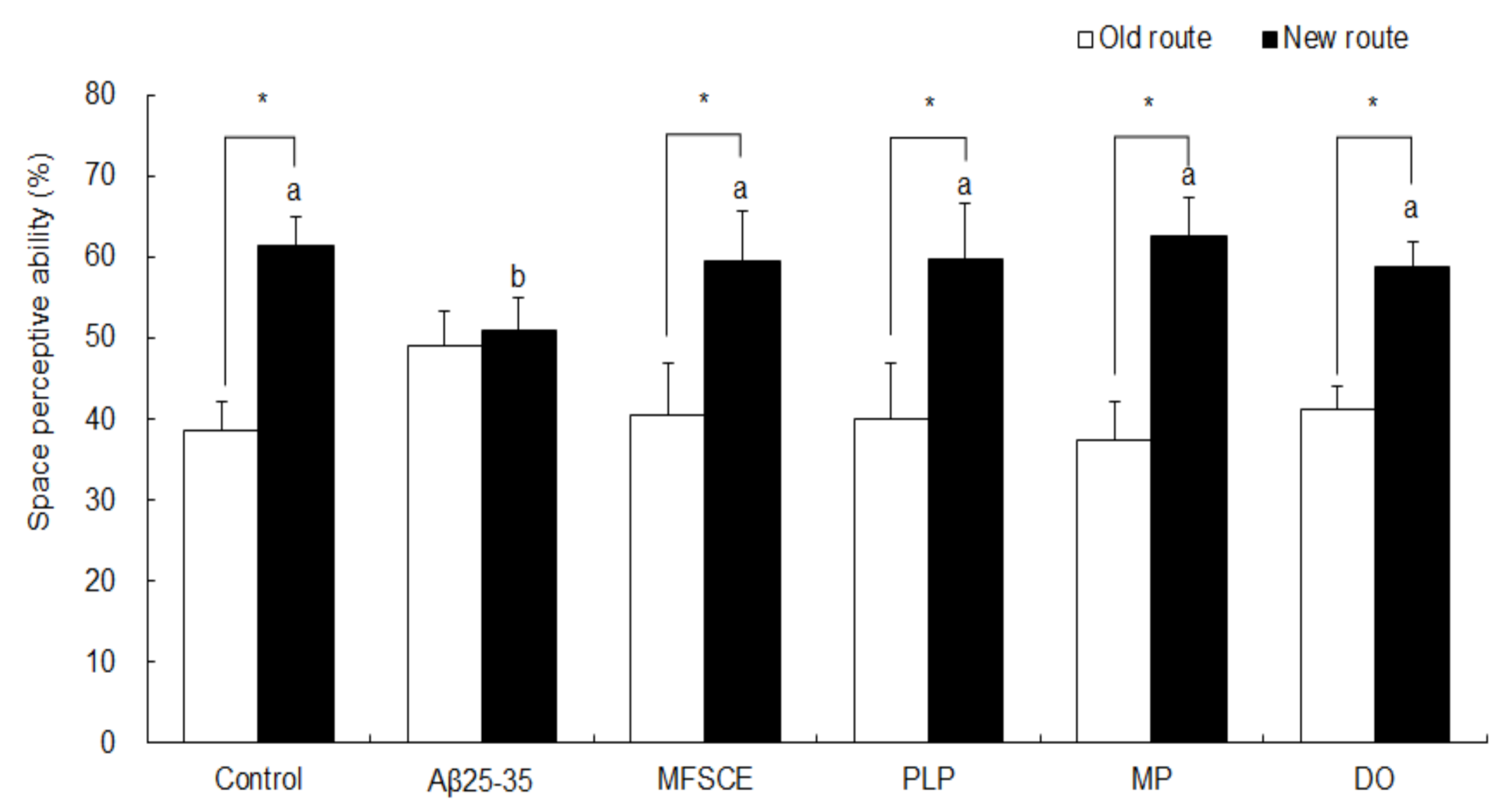
3.1. T-Maze Test
3.2. Novel Object Recognition (NOR) Test
3.3. Morris Water Maze (MWM) Test
3.4. Effect of MFSCE on the Levels of Malondialdehyde (MDA) and Nitric Oxide (NO) in Aβ25–35-Injected Mice Brain
3.5. Effect of MFSCE on Neuronal Apoptosis in Aβ25–35-Injected Mice Brain
3.6. Effect of MFSCE on the Amyloidogenic Pathway in Aβ25–35-Injected Mice Brain
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, H.C.; Jiang, Z.F. Accumulated amyloid-beta peptide and hyperphosphorylated tau protein: Relationship and links in Alzheimer’s disease. J. Alzheimers Dis. 2009, 16, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Robert, D.; Terry, M.D. Cell death or synaptic loss in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2000, 59, 1118–1119. [Google Scholar]
- Selkoe, D.J. The molecular pathology of Alzheimer’s disease. Neuron 1991, 6, 487. [Google Scholar] [CrossRef]
- Guo, T.; Hobbs, D.W. Development of BACE1 inhibitors for Alzheimer’s disease. Curr. Med. Chem. 2006, 13, 1811–1829. [Google Scholar] [CrossRef]
- Floyd, R.A.; Hensley, K. Oxidative stress in brain aging: Implications for therapeutics of neurodegenerative diseases. Neurobiol. Aging 2002, 23, 795–807. [Google Scholar] [CrossRef]
- Rogers, J.; Webster, S.; Lue, L.F.; Brachova, L.; Civin, W.H.; Emmerling, M.; Shivers, B.; Walker, D.; McGeer, P. Inflammation and Alzheimer’s disease pathogenesis. Neurobiol. Aging 1996, 17, 681–686. [Google Scholar] [CrossRef]
- Prabhakaran, M.P.; Venugopal, J.R.; Ramakrishna, S. Mesenchymal stem cell differentiation to neuronal cells on electrospun nanofibrous substrates for nerve tissue engineering. Biomaterials 2009, 30, 4996–5003. [Google Scholar] [CrossRef]
- Planat-Bénard, V.; Menard, C.; André, M.; Puceat, M.; Perez, A.; Garcia-Verdugo, J.M.; Pénicaud, L.; Casteilla, L. Spontaneous cardiomyocyte differentiation from adipose tissue stroma cells. Circ. Res. 2004, 94, 223–229. [Google Scholar] [CrossRef]
- Lou, G.; Chen, Z.; Zheng, M.; Liu, Y. Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases. Exp. Mol. Med. 2017, 49, e346. [Google Scholar] [CrossRef]
- Saralamma, V.V.G.; Vetrivel, P.; Kim, S.M.; Ha, S.E.; Lee, H.J.; Lee, S.J.; Kim, Y.S.; Pak, J.E.; Lee, H.J.; Heo, J.D.; et al. Proteome profiling of membrane-free stem cell components by nano-LS/MS analysis and its anti-inflammatory activity. Evid.-Based Complement. Alternat. Med. 2019, 2019, 4683272. [Google Scholar]
- Kim, M.J.; Kim, J.H.; Park, H.S.; Kim, Y.S.; Cho, E.J. Protective effect of membrane-free stem cell extract against oxidative stress in LLC-PK1 cells. J. Korea Acad. Ind. Coop. Soc. 2019, 20, 303–312. [Google Scholar]
- He, M.T.; Kim, J.H.; Kim, Y.S.; Park, H.S.; Cho, E.J. Protective effects of membrane-free stem cell extract from H2O2-induced inflammation responses in human periodontal ligament fibroblasts. J. Korea Acad. Ind. Coop. Soc. 2019, 20, 95–103. [Google Scholar]
- Park, H.S.; Pang, Q.Q.; Kim, Y.S.; Kim, J.H.; Cho, E.J. Neuroprotective effect of membrane-free stem cell extract against amyloid beta 25-35-indcued neurotoxicity in SH-SY5Y cells. Appl. Sci. 2021, 11, 2219. [Google Scholar] [CrossRef]
- Mooney, S.; Leuendorf, J.E.; Hendrickson, C.; Hellmann, H. Vitamin B6: A long known compound of surprising complexity. Molecules 2009, 14, 329–351. [Google Scholar] [CrossRef]
- Hellmann, H.; Mooney, S. Vitamin B6: A molecule for human health. Molecules 2010, 15, 442–459. [Google Scholar] [CrossRef]
- Riede, U.N.; Sandritter, W.; Pietzxch, A.; Rohrbach, R. Reaction patterns of cell organelles in vitamin B6 deficiency: Ultrastructural-morphometric analysis of the liver parenchymal cell. Pathol. Res. Pract. 1980, 170, 376–387. [Google Scholar] [CrossRef]
- Parra, M.; Stahl, S.; Hellmann, H. Vitamin B6 and its role in cell metabolism and physiology. Cells 2018, 7, 84. [Google Scholar] [CrossRef]
- Maurice, T.; Lockhart, B.P.; Privat, A. Amnesia induced in mice by centrally administered β-amyloid peptides involves cholinergic dysfunction. Brain Res. 1996, 706, 181–193. [Google Scholar] [CrossRef]
- Laursen, S.E.; Belknap, J.K. Intracerebroventricular injections in mice. Some methodological refinements. J. Pharmacol. Method 1986, 16, 355–357. [Google Scholar] [CrossRef]
- Montgomery, K.C. A test of two explanations of spontaneous alternation. J. Comp. Physiol. Psych. 1952, 45, 287–293. [Google Scholar] [CrossRef]
- Bevins, R.A.; Besheer, J. Object recognition in rats and mice: A one-trial non-matching-to-sample learning task to study ‘recognition memory’. Nat. Protoc. 2006, 1, 1306–1311. [Google Scholar] [CrossRef] [PubMed]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Meth. 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Schmidt, H.H.; Warner, T.D.; Nakane, M.; Förstermann, U.; Murad, F. Regulation and subcellular location of nitrogen oxide synthases in RAW264.7 macrophages. Mol. Pharmacol. 1992, 41, 615–624. [Google Scholar] [PubMed]
- Muraoka, K.; Shingo, T.; Yasuhara, T.; Kameda, M.; Yuen, W.J.; Uozumi, T.; Matsui, T.; Miyoshi, Y.; Date, I. Comparison of the therapeutic potential of adult and embryonic neural precursor cells in a rat model of Parkinson disease. J. Neurosurg. 2008, 108, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.P.; Lang, B.T.; Baskaya, M.K.; Dempsey, R.J.; Vemuganti, R. The potential of neural stem cells to repair stroke-induced brain damage. Acta Neuropathol. 2009, 117, 469–480. [Google Scholar] [CrossRef]
- Kim, S.; Chang, K.A.; Kim, J.A.; Park, H.G.; Ra, J.C.; Kim, H.S.; Suh, Y.H. The preventive and therapeutic effects of intravenous human adipose-derived stem cells in Alzheimer’s disease mice. PLoS ONE 2012, 7, e45757. [Google Scholar] [CrossRef] [PubMed]
- Blurton-Jones, M.; Kitazawa, M.; Martinez-Coria, H.; Castello, N.A.; Muller, F.J.; Loring, J.F.; Yamasaki, T.R.; Poon, W.W.; Green, K.N.; LaFerla, F.M. Neural stem cells improve cognition via BDNF in a transgenic model of Alzheimer disease. Proc. Natl. Acad. Sci. USA 2009, 106, 13594–13599. [Google Scholar] [CrossRef]
- Lee, J.K.; Jin, H.K.; Endo, S.; Schuchman, E.H.; Carter, J.E.; Bae, J.S. Intracerebral transplantation of bone marrow-derived mesenchymal stem cells reduces amyloid-beta deposition and rescues memory deficits in Alzheimer’s disease mice by modulation of immune responses. Stem Cells 2010, 28, 329–343. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Chen, Y.; Yuan, L.; Liu, H.; Wang, J.; Liu, Q.; Zhang, Y. Adipose-derived stem cells: Current applications and future directions in the regeneration of multiple tissues. Stem Cell Int. 2020, 2020, 8810813. [Google Scholar] [CrossRef]
- Huang, T.; He, D.; Kleiner, G.; Kuluz, J. Neuron-like differentiation of adipose-derived stem cells from infant piglets in vitro. J. Spinal Cord. Med. 2007, 30, S35–S40. [Google Scholar] [CrossRef]
- Nakamura, S.; Murayama, N.; Noshita, T.; Annoura, H.; Ohno, T. Progressive brain dysfunction following intracerebroventricular infusion of beta1–42-amyloid peptide. Brain Res. 2001, 912, 128–136. [Google Scholar] [CrossRef]
- Chistensen, R.; Marcussen, A.B.; Wörtwein, G.; Knudsen, G.M.; Aznar, S. Aβ(1–42) injection causes memory impairment, lowered cortical and serum BDNF levels, and decreased hippocampal 5-HT2A levels. Exp. Neurol. 2008, 210, 164–171. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lee, D.K.; Chung, B.R.; Kim, H.J.; Kim, Y.S. Intracerebroventricular injection of amyloid-β peptides in normal mice to acutely induce Alzheimer-like cognitive deficits. J. Vis. Exp. 2016, 109, e53308. [Google Scholar] [CrossRef]
- Varadarajan, S.; Kanski, J.; Aksenova, M.; Lauderback, C.; Butterfield, D.A. Different mechanisms of oxidative stress and neurotoxicity for Alzheimer’s A beta (1-42) and A beta (25-35). J. Am. Chem. Soc. 2001, 123, 5625–5631. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Reed, T.; Newman, S.F.; Sultana, R. Roles of amyloid β-peptide-associated oxidative stress and brain protein modifications in the pathogenesis of Alzheimer’s disease and mild cognitive impairment. Free Radic. Biol. Med. 2007, 43, 658–677. [Google Scholar] [CrossRef]
- Cheignon, C.; Tomas, M.; Bonnefont-Rousselot, D.; Faller, P.; Hueau, C.; Collin, F. Oxidative stress and the amyloid beta peptide in Alzheimer’s disease. Redox Biol. 2018, 14, 450–464. [Google Scholar] [CrossRef]
- Montine, T.J.; Neely, M.D.; Quinn, J.F.; Beal, M.L.; Markesbery, W.R.; Roberts, L.J.; Morrow, J.D. Lipid peroxidation in aging brain and Alzheimer’s disease. Free Radic. Biol. Med. 2002, 33, 620–626. [Google Scholar] [CrossRef]
- Shukla, L.; Morrison, W.A.; Shayan, R. Adipose-derived stem cells in radiotherapy injury: A new frontier. Front. Surg. 2015, 28, 1. [Google Scholar] [CrossRef]
- Ge, Y.; Zhang, Q.; Jiao, Z.; Li, H.; Bai, G.; Wang, H. Adipose-derived stem cells reduce liver oxidative stress and autophagy induced by ischemia-reperfusion and hepatectomy injury in swine. Life Sci. 2018, 214, 62–69. [Google Scholar] [CrossRef]
- Hashim, A.; Wang, L.; Juneja, K.; Ye, Y.; Zhao, Y.; Ming, L.J. Vitamin B6s inhibit oxidative stress caused by Alzheimer’s disease-related CuII-β-amyloid complexes-cooperative action of phospho-moiety. Bioorg. Med. Chem. Lett. 2011, 21, 6430–6432. [Google Scholar] [CrossRef] [PubMed]
- Forloni, G.; Chiesa, R.; Smiroldo, S.; Verga, L.; Salmona, M.; Tagliavini, F.; Angeretti, N. Apoptosis mediated neurotoxicity induced by chronic application of beta amyloid fragment 25–35. Neuroreport 1993, 4, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Crews, L.; Masliah, E. Molecular mechanisms of neurodegeneration in Alzheimer’s disease. Hum. Mol. Gen. 2010, 19, R12–R20. [Google Scholar] [CrossRef] [PubMed]
- Morishima, Y.; Gotoh, Y.; Zieg, J.; Barrett, T.; Takano, H.; Flavell, R.; Davis, R.J.; Shirasaki, Y.; Greenberg, M.E. β-Amyloid induces neuronal apoptosis via a mechanism that involves the c-Jun N-terminal kinase pathway and the induction of Fas ligand. J. Neurosci. 2001, 21, 7551–7560. [Google Scholar] [CrossRef]
- Loo, D.T.; Copani, A.; Pike, C.J.; Whittemore, E.R.; Walencewicz, A.J.; Cotman, C.W. Apoptosis is induced by beta-amyloid in cultured central nervous system neurons. Proc. Natl. Acad. Sci. USA 1993, 90, 7951–7955. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.W.; Eldadah, B.A.; Huang, X.; Knoblach, S.M.; Faden, A.I. Multiple caspases are involved in β-amyloid-induced neuronal apoptosis. J. Neurosci. Res. 2001, 65, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Ferreiro, E.; Cardoso, S.M.; de Oliveira, C.R. Cell degeneration induced by amyloid-β peptides. J. Mol. Neurosci. 2004, 23, 97–104. [Google Scholar] [CrossRef]
- Wei, M.C.; Zong, W.X.; Cheng, E.H.; Lindsten, T.; Panoutsakopoulou, V.; Ross, A.J.; Roth, K.A.; MacGregor, G.R.; Thompson, C.B.; Korsmeyer, S.J. Proapoptotic BAX and BAK: A requisite gateway to mitochondrial dysfunction and death. Science 2001, 292, 727–730. [Google Scholar] [CrossRef]
- Deng, G.; Su, J.H.; Ivins, K.L.; van Houten, B.; Cotman, C.W. Bcl-2 facilitates recovery from DNA damage after oxidative stress. Exp. Neurol. 1999, 159, 309–318. [Google Scholar] [CrossRef]
- Putcha, G.V.; Deshmukh, M.; Johnson, E.M., Jr. BAX translocation is a critical event in neuronal apoptosis: Regulation by neuroprotectants, BCL-2, and caspases. J. Neurosci. 1999, 19, 7476–7485. [Google Scholar] [CrossRef]
- Wang, H.; Xu, Y.; Yan, J.; Zhao, X.; Sun, X.; Zhang, Y.; Guo, J.; Zhu, C. Acteoside protects human neuroblastoma SH-SY5Y cells against β-amyloid-induced cell injury. Brain Res. 2009, 1283, 139–147. [Google Scholar] [CrossRef]
- Marin, N.; Romero, B.; Bosch-Morell, F.; Llansola, M.; Felipo, V.; Roma, J.; Romero, F.J. β-Amyloid-induced activation of Caspase-3 in primary cultures of rat neurons. Mech. Ageing Dev. 2000, 119, 63–67. [Google Scholar] [CrossRef]
- Mohebali, N.; Fazeli, S.A.S.; Ghafoori, H.; Farahmand, Z.; MohammadKhani, E.; Vakhshiteh, F.; Ghamarian, A.; Farhangniya, M.; Sanati, M.H. Effect of flavonoids rich extract of Capparis spinosa on inflammatory involved genes in amyloid-beta peptide injected rat model of Alzheimer’s disease. Nurt. Neurosci. 2016, 21, 143–150. [Google Scholar]
- Amon, F.U.; Shah, S.A.; Kim, M.O. Vanillic acid attenuates Aβ1-42-induced oxidative stress and cognitive impairment in mice. Sci. Rep. 2017, 7, 40753. [Google Scholar] [CrossRef]
- Lee, A.Y.; Lee, M.H.; Lee, S.; Cho, E.J. Alpha-linolenic acid from Perilla frutescens var. japonica oil protects Aβ-induced cognitive impairment through regulation of APP processing and Aβ degradation. J. Agric. Food Chem. 2017, 65, 10719–10729. [Google Scholar] [CrossRef]
- Chu, J.; Li, J.G.; Joshi, Y.B.; Giannopoulos, P.F.; Hoffman, N.E.; Madesh, M.; Pratico, D. Gamma secretase-activating protein is a substrate for caspase-3: Implications for Alzheimer’s disease. Biol. Psychiatry 2015, 77, 720–728. [Google Scholar] [CrossRef][Green Version]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Lee, M.S.; Su, T.C.; Huang, Y.C.; Yang, R.C.; Kao, J.K.; Lee, C.H.; Tseng, J.J.; Hsu, C.S.; Hsu, C.L. Effects of vitamin B-6 supplementation on oxidative stress and inflammatory response in neonatal rats receiving hyperoxia therapy. J. Food Drug Anal. 2018, 26, 1086–1096. [Google Scholar] [CrossRef]
- Pahlavanzadeh, F.; Bidadkosh, A.; Derakhshanfar, A.; Rastegar, A.M.; Rushanzamir, M. Antioxidant protecting effects of vitamin B6 at reducing hemodynamic toxicity of gentamicin in rat model of nephrotoxicity. Comp. Clin. Pathol. 2013, 22, 637–643. [Google Scholar] [CrossRef]
- Geng, M.Y.; Saito, H.; Katsuki, H. Effects of vitamin B6 and its related compounds on survival of cultured brain neurons. Neurosci. Res. 1995, 24, 61–65. [Google Scholar] [CrossRef]
- Christen, P.; Mehta, P.K. From cofactor to enzymes. The molecular evolution of pyridoxal-5′-phosphate-dependent enzymes. Chem. Rec. 2001, 1, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Percudani, R.; Peracchi, A. A genomic overview of pyridoxal-phosphate-dependent enzymes. EMBO Rep. 2003, 4, 850–854. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n | Treatment |
|---|---|---|
| Control | 10 | 0.9% NaCl saline i.c.v. + 0.9% NaCl saline i.p. |
| Aβ25–35 | 10 | Aβ25–35 i.c.v. + 0.9% NaCl saline i.p. |
| MFSCE | 10 | Aβ25–35 i.c.v. + MFSCE (100 mg/kg) i.p. |
| PLP | 10 | Aβ25–35 i.c.v. + PLP (1 mg/kg) i.p. |
| MP | 10 | Aβ25–35 i.c.v. + MFSCE (100 mg/kg) and PLP (1 mg/kg) i.p. |
| DO | 9 | Aβ25–35 i.c.v. + donepezil (5 mg/kg) i.p. |
| Group | Concentration | |
|---|---|---|
| MDA (μmol/mg Protein) | NO (nmol/mg Protein) | |
| Control | 74.05 ± 4.39 d | 9.68 ± 1.07 d |
| Aβ25–35 | 108.11 ± 7.47 a | 28.20 ± 6.56 a |
| MFSCE | 86.67 ± 8.64 bc | 14.33 ± 1.65 bc |
| PLP | 86.22 ± 2.20 b | 15.48 ± 1.88 b |
| MP | 80.43 ± 8.34 cd | 11.43 ± 3.15 cd |
| DO | 89.49 ± 9.57 bc | 15.25 ± 3.21 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.M.; Park, H.S.; He, M.T.; Kim, Y.S.; Kim, H.Y.; Lee, A.Y.; Cho, E.J. Membrane-Free Stem Cells and Pyridoxal 5′-Phosphate Synergistically Enhance Cognitive Function in Alzheimer’s Disease Mouse Model. Antioxidants 2022, 11, 601. https://doi.org/10.3390/antiox11030601
Choi JM, Park HS, He MT, Kim YS, Kim HY, Lee AY, Cho EJ. Membrane-Free Stem Cells and Pyridoxal 5′-Phosphate Synergistically Enhance Cognitive Function in Alzheimer’s Disease Mouse Model. Antioxidants. 2022; 11(3):601. https://doi.org/10.3390/antiox11030601
Chicago/Turabian StyleChoi, Ji Myung, Hye Sook Park, Mei Tong He, Young Sil Kim, Hyun Young Kim, Ah Young Lee, and Eun Ju Cho. 2022. "Membrane-Free Stem Cells and Pyridoxal 5′-Phosphate Synergistically Enhance Cognitive Function in Alzheimer’s Disease Mouse Model" Antioxidants 11, no. 3: 601. https://doi.org/10.3390/antiox11030601
APA StyleChoi, J. M., Park, H. S., He, M. T., Kim, Y. S., Kim, H. Y., Lee, A. Y., & Cho, E. J. (2022). Membrane-Free Stem Cells and Pyridoxal 5′-Phosphate Synergistically Enhance Cognitive Function in Alzheimer’s Disease Mouse Model. Antioxidants, 11(3), 601. https://doi.org/10.3390/antiox11030601