
Gingerenone A Induces Antiproliferation and Senescence of Breast Cancer Cells

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Extraction, and Isolation
2.2. Gin A Chemical Profile
2.3. Cell Culture and Chemicals
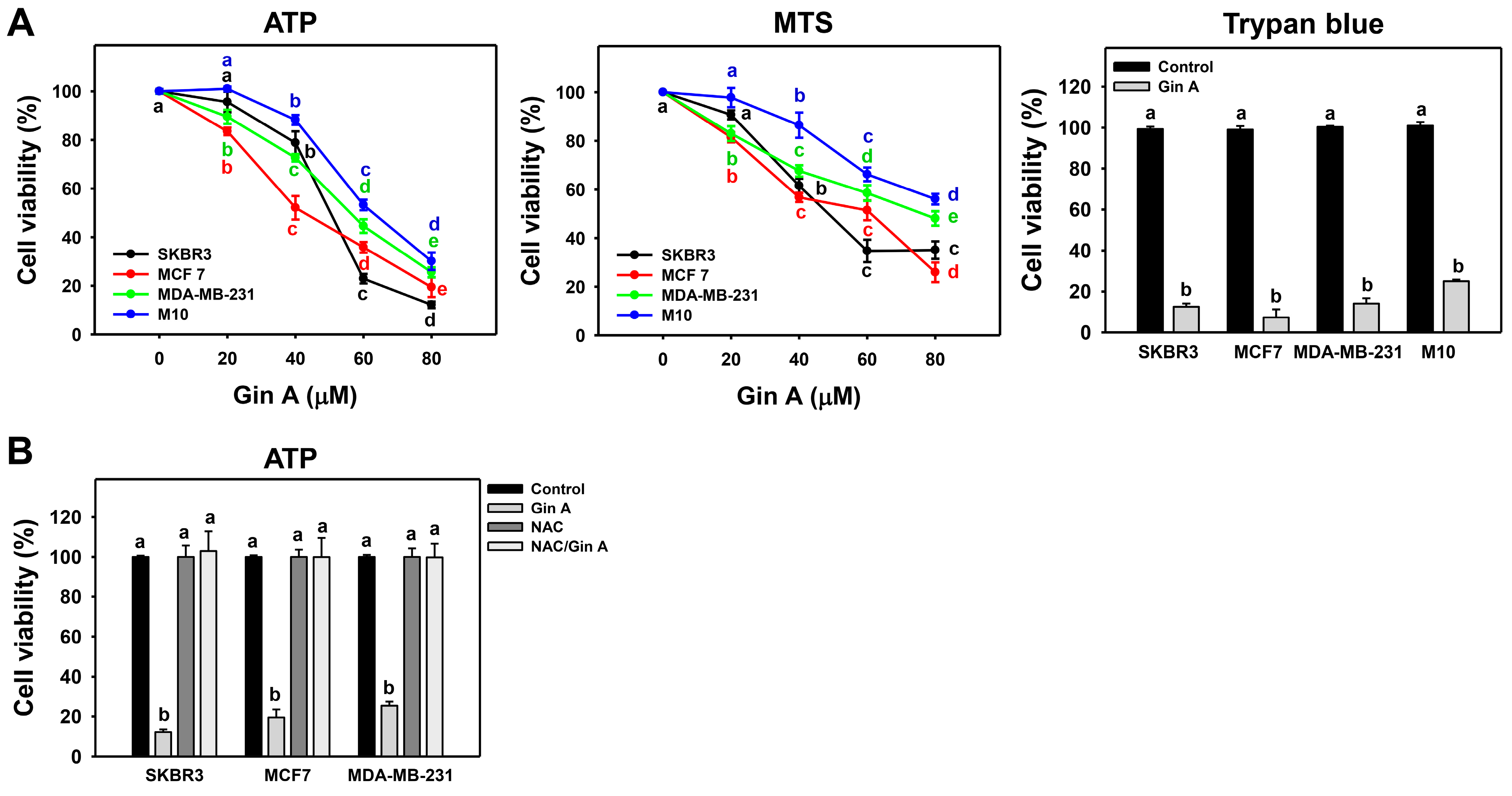
2.4. Cell Viability
2.5. Cell Cycle Detection
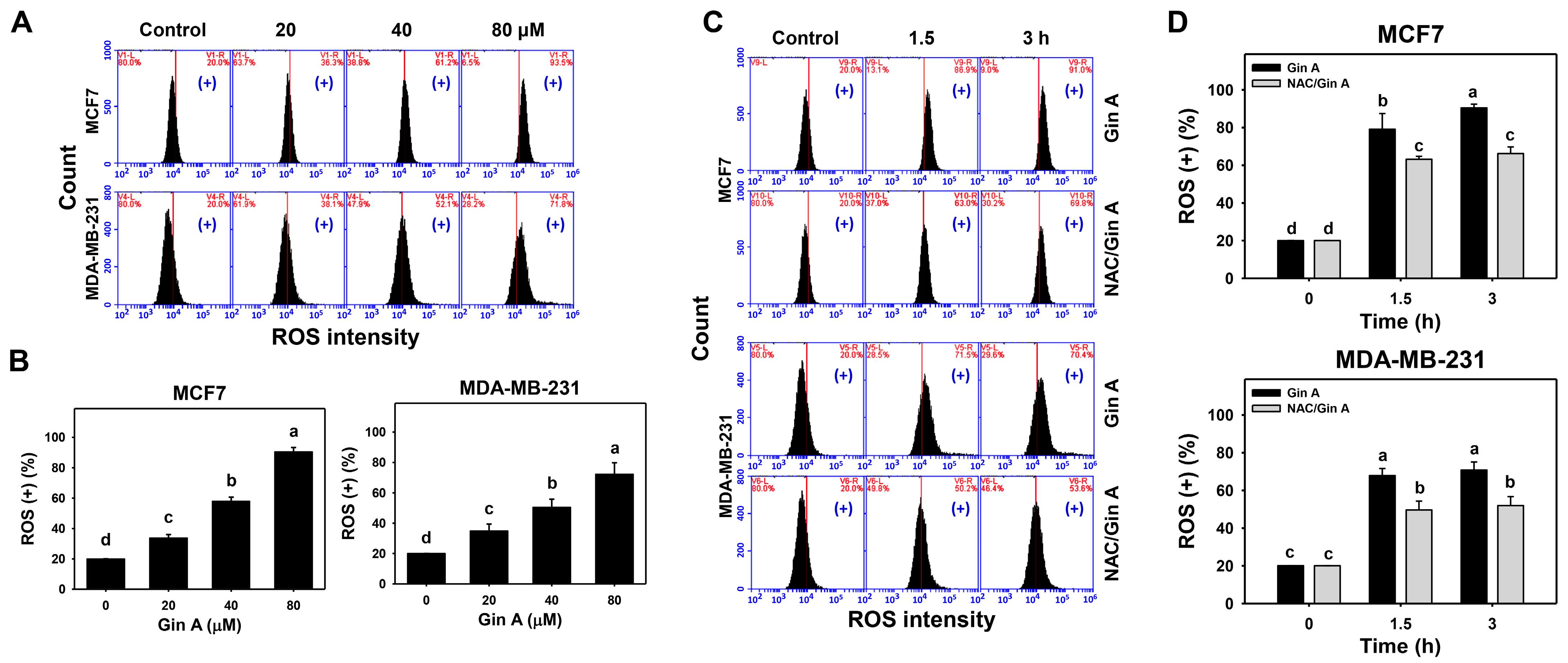
2.6. ROS Detection
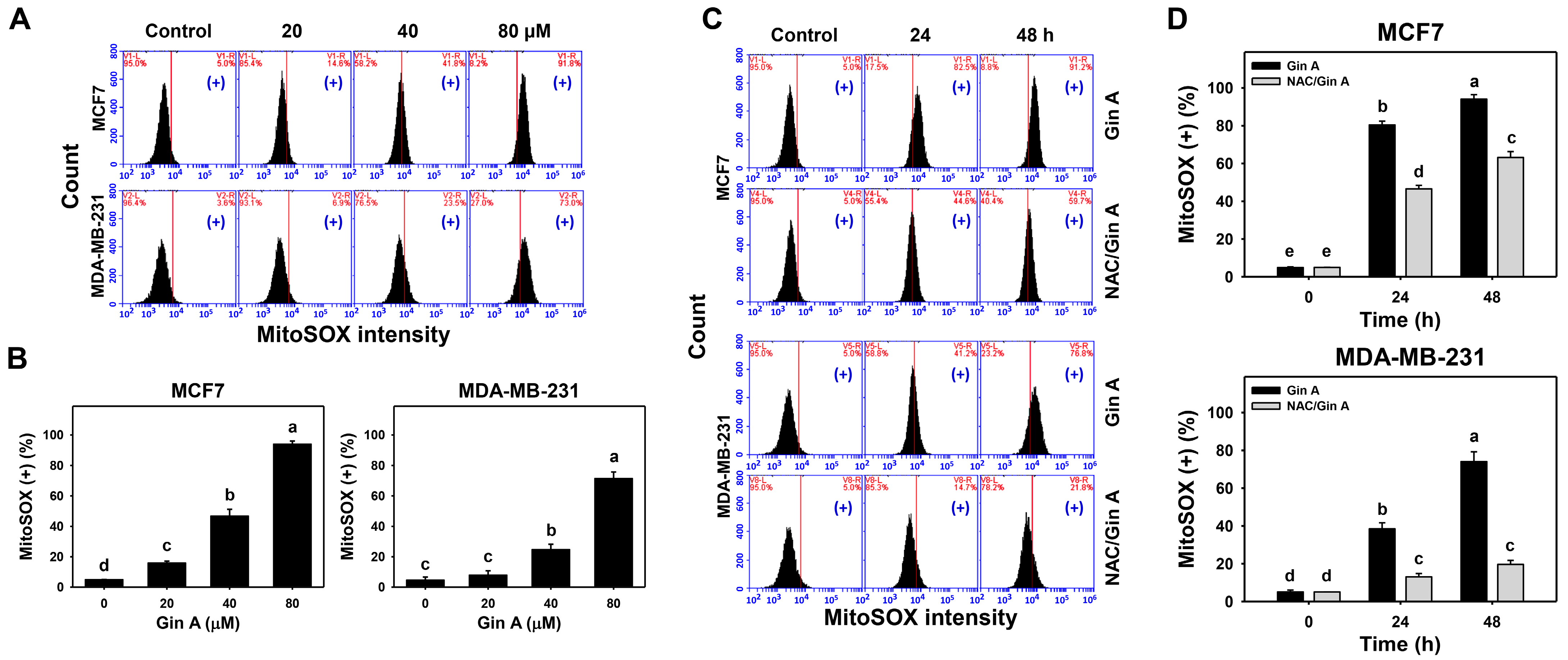
2.7. Mitochondrial Superoxide (MitoSOX) Detection by Flow Cytometry
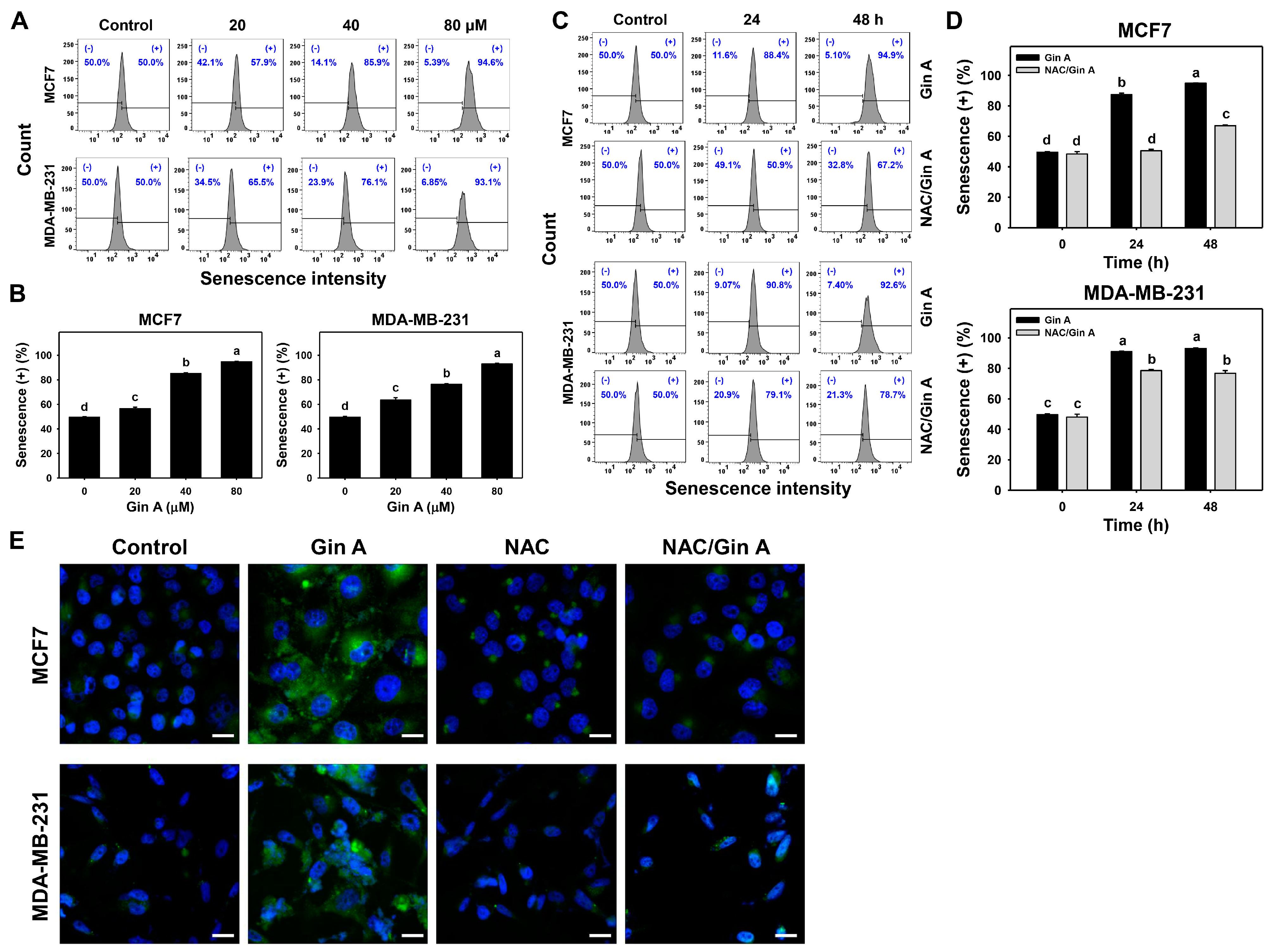
2.8. Senescence Detection
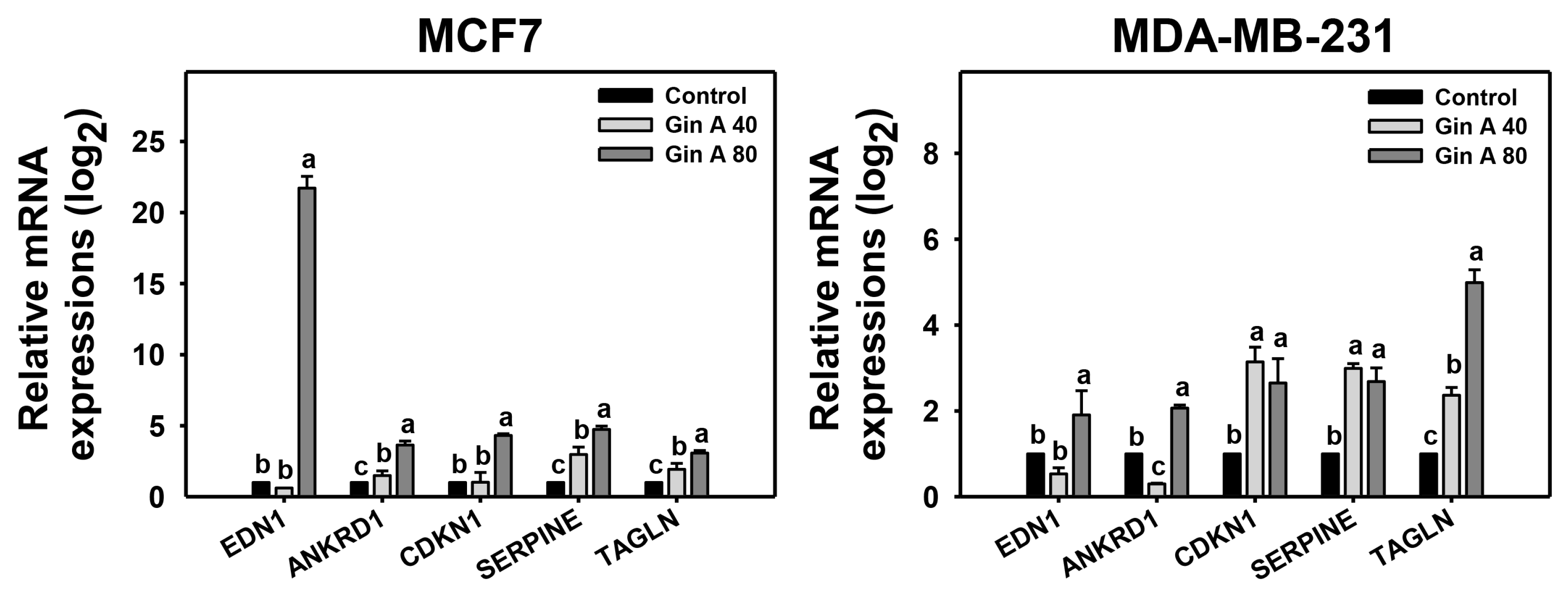
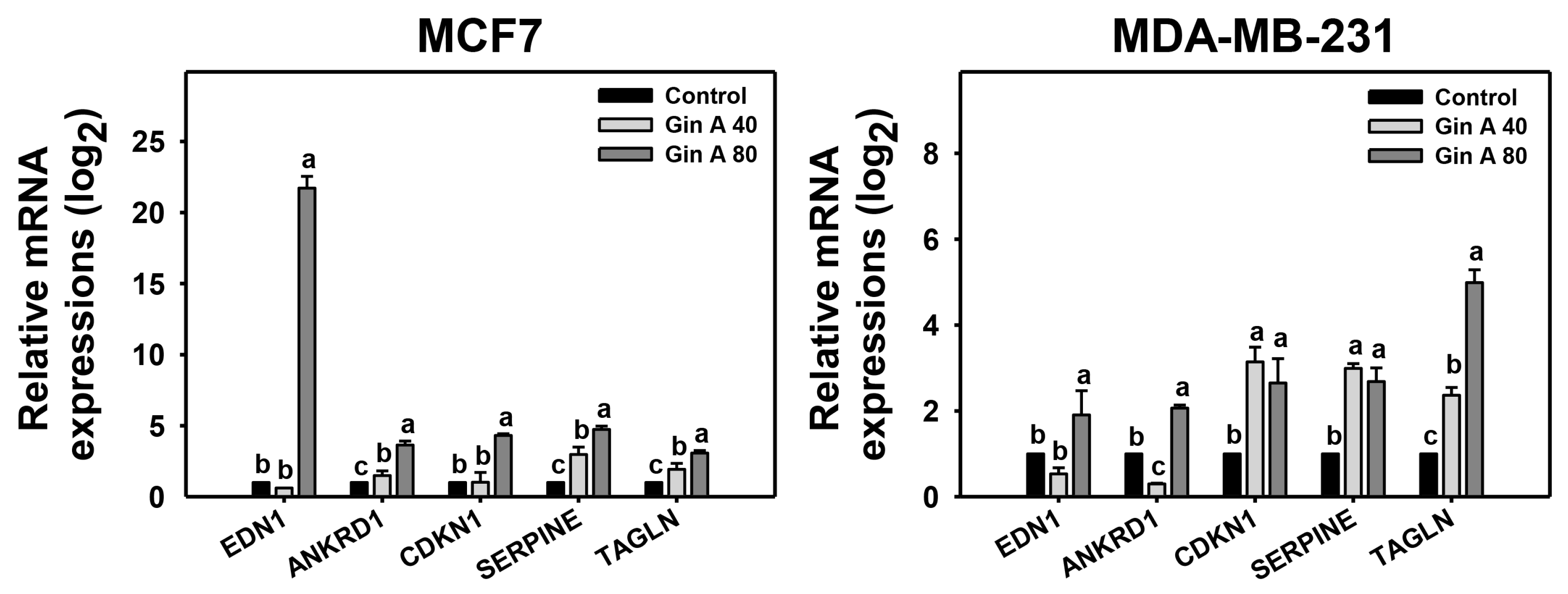
2.9. Senescence Gene Expression Detection
2.10. DNA Damage Detection
2.11. 8-Hydroxyl-2′-Deoxyguanosine (8-OHdG) Detection
2.12. Statistics
3. Results
3.1. Gin A Diminished Proliferation of Breast Cancer Cells
3.2. Gin A Delayed G2/M Progression of Breast Cancer Cells
3.3. Gin A Provoked ROS Increment in Breast Cancer Cells
3.4. Gin A Provoked MitoSOX Increment of Breast Cancer Cells
3.5. Gin A Provoked the Appearance of Senescence in Breast Cancer Cells
3.6. Gin A Provoked Senescence-Associated Gene Expressions of Breast Cancer Cells
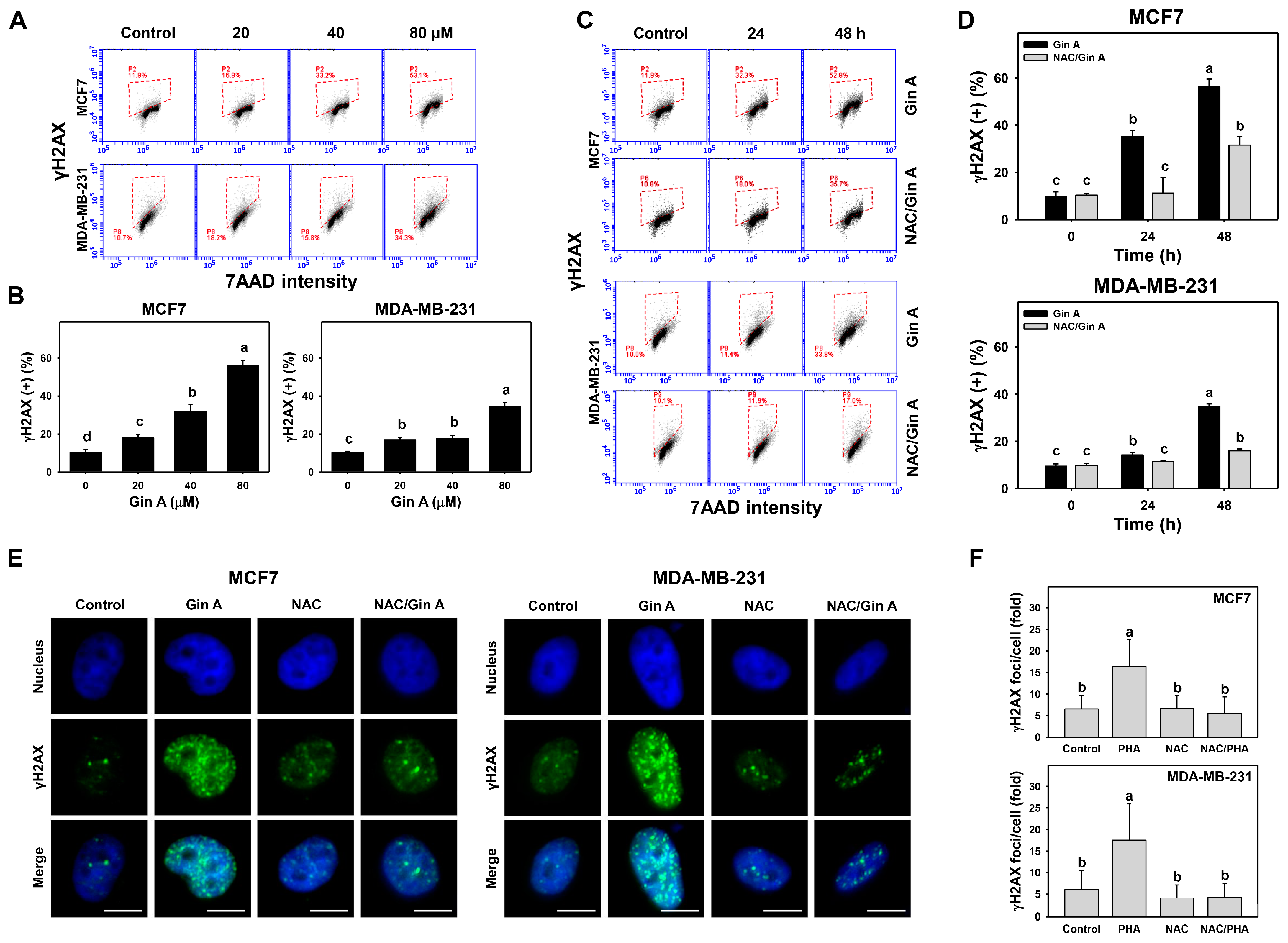
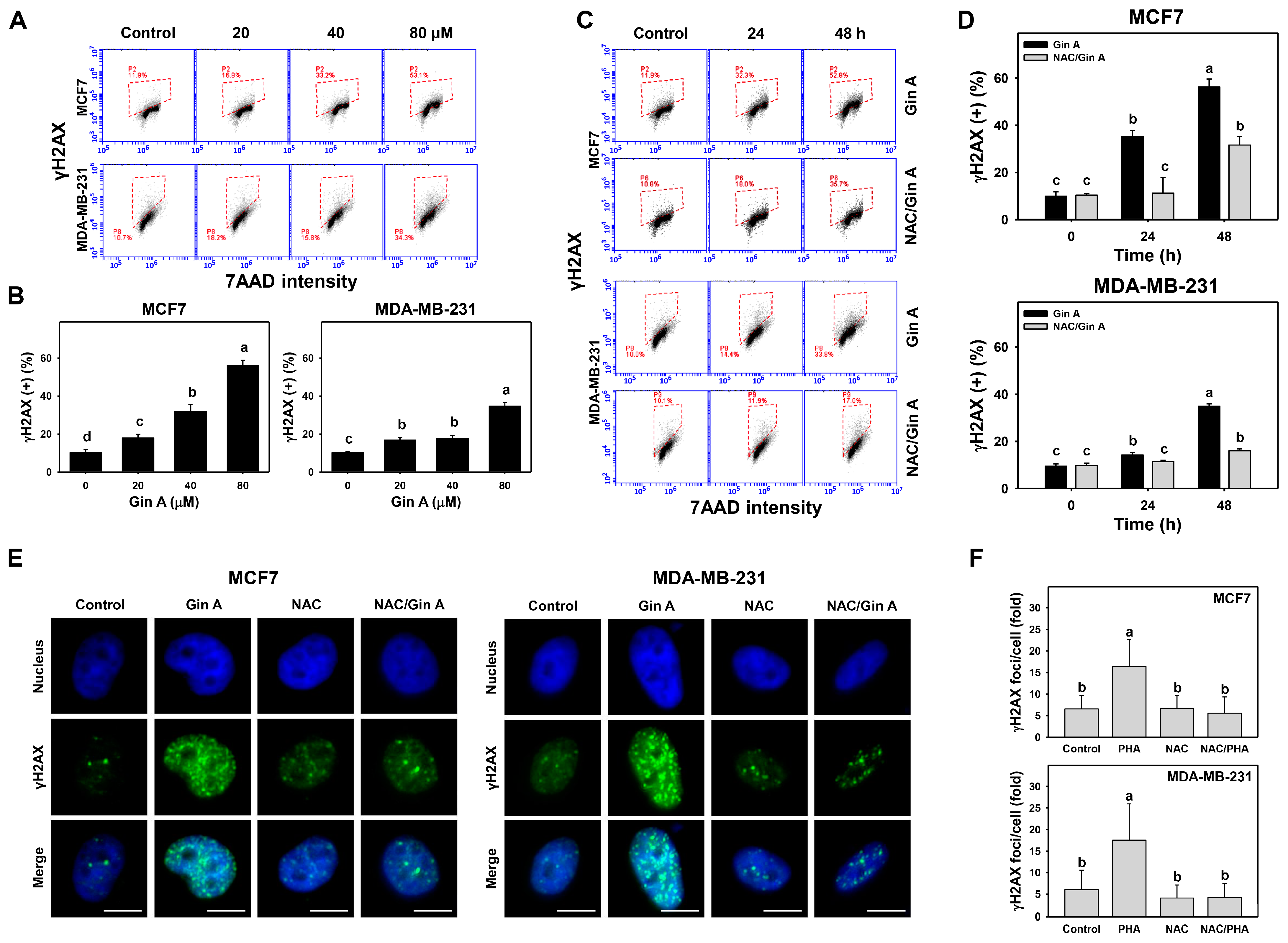
3.7. Gin A Provoked γH2AX Appearance of Breast Cancer Cells
3.8. Gin A Provoked 8-OHdG Increment of Breast Cancer Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLO-BOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. 2011, 13, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, B.D.; Pietenpol, J.A. Identification and use of biomarkers in treatment strategies for triple-negative breast cancer subtypes. J. Pathol. 2014, 232, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Burstein, M.D.; Tsimelzon, A.; Poage, G.M.; Covington, K.R.; Contreras, A.; Fuqua, S.A.; Savage, M.I.; Osborne, C.K.; Hilsenbeck, S.G.; Chang, J.C.; et al. Comprehensive Genomic Analysis Identifies Novel Subtypes and Targets of Triple-Negative Breast Cancer. Clin. Cancer Res. 2015, 21, 1688–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagano, E.; Souto, E.B.; Durazzo, A.; Sharifi-Rad, J.; Lucarini, M.; Souto, S.B.; Salehi, B.; Zam, W.; Montanaro, V.; Lucariello, G.; et al. Ginger (Zingiber officinale Roscoe) as a nutraceutical: Focus on the metabolic, analgesic, and antiinflammatory effects. Phytother. Res. 2021, 35, 2403–2417. [Google Scholar] [CrossRef]
- Cheema, H.S.; Singh, M.P. The Use of Medicinal Plants in Digestive System Related Disorders—A Systematic Review. J. Ayurvedic Herb. Med. 2021, 7, 182–187. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, R.; Wang, D.; Wang, L.; Zhang, Q.; Wei, S.; Lu, F.; Peng, W.; Wu, C. Ginger (Zingiber officinale Roscoe) and its bioactive components are potential resources for health beneficial agents. Phytother. Res. 2021, 35, 711–742. [Google Scholar] [CrossRef] [PubMed]
- Kubra, I.R.; Rao, L.J.M. An Impression on Current Developments in the Technology, Chemistry, and Biological Activities of Ginger (Zingiber officinale Roscoe). Crit. Rev. Food Sci. Nutr. 2012, 52, 651–688. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-M.; Kao, C.-L.; Tseng, Y.-T.; Lo, Y.-C.; Chen, C.-Y. Ginger Phytochemicals Inhibit Cell Growth and Modulate Drug Resistance Factors in Docetaxel Resistant Prostate Cancer Cell. Molecules 2017, 22, 1477. [Google Scholar] [CrossRef]
- Oboh, G.; Akinyemi, A.J.; Ademiluyi, A.O. Antioxidant and inhibitory effect of red ginger (Zingiber officinale var. Rubra) and white ginger (Zingiber officinale Roscoe) on Fe2+ induced lipid peroxidation in rat brain in vitro. Exp. Toxicol. Pathol. 2012, 64, 31–36. [Google Scholar] [CrossRef]
- Mann, T.F.; Chen, C.Y.; Chern, C.L.; Liou, S.S.; Yiin, S.J. Gingernone A-induced apoptosis in SK-Hep-1 cells is mediated via increased reactive oxygen species (ROS) production and the mitochondria-associated apoptotic mechanisms. Tajen J. 2015, 47, 1–14. [Google Scholar]
- Wang, B.; Kohli, J.; Demaria, M. Senescent Cells in Cancer Therapy: Friends or Foes? Trends Cancer 2020, 6, 838–857. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, P.G.; Citrin, D.E.; Hildesheim, J.; Ahmed, M.M.; Venkatachalam, S.; Riscuta, G.; Xi, D.; Zheng, G.; van Deursen, J.V.; Goronzy, J.; et al. Therapy-Induced Senescence: Opportunities to Improve Anticancer Therapy. JNCI J. Natl. Cancer Inst. 2021, 113, 1285–1298. [Google Scholar] [CrossRef]
- Pole, A.; Dimri, M.; Dimri, G.P. Oxidative stress, cellular senescence and ageing. AIMS Mol. Sci. 2016, 3, 300–324. [Google Scholar] [CrossRef]
- Li, Y.; Li, W.; Ying, Z.; Tian, H.; Zhu, X.; Li, J.; Li, M. Metastatic Heterogeneity of Breast Cancer Cells Is Associated with Expression of a Heterogeneous TGFβ-Activating miR424–503 Gene Cluster. Cancer Res. 2014, 74, 6107–6118. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Cuadrado, L.; Tracey, N.; Ma, R.; Qian, B.; Brunton, V.G. Mouse models of metastasis: Progress and prospects. Dis. Model. Mech. 2017, 10, 1061–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.W.; Tang, J.Y.; Ou-Yang, F.; Wang, H.R.; Guan, P.Y.; Huang, C.Y.; Chen, C.Y.; Hou, M.F.; Sheu, J.H.; Chang, H.W. Sinularin selectively kills breast cancer cells showing G2/M arrest, apoptosis, and oxidative DNA damage. Molecules 2018, 23, 849. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.H.; Shiao, N.H.; Lu, P.Z. CdSe quantum dots induce apoptosis in human neuroblastoma cells via mitochondri-al-dependent pathways and inhibition of survival signals. Toxicol. Lett. 2006, 167, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.-H.; Chen, C.-Y.; Omar, H.A.; Huang, K.-Y.; Tsao, C.-C.; Chiu, C.-C.; Chen, Y.-L.; Chen, P.-H.; Teng, Y.-N. Reactive oxygen species mediate Terbufos-induced apoptosis in mouse testicular cell lines via the modulation of cell cycle and pro-apoptotic proteins. Environ. Toxicol. 2015, 31, 1888–1898. [Google Scholar] [CrossRef]
- Huang, C.-H.; Yeh, J.-M.; Chan, W.-H. Hazardous impacts of silver nanoparticles on mouse oocyte maturation and fertilization and fetal development through induction of apoptotic processes. Environ. Toxicol. 2018, 33, 1039–1049. [Google Scholar] [CrossRef]
- Wang, T.-S.; Lin, C.-P.; Chen, Y.-P.; Chao, M.-R.; Li, C.-C.; Liu, K.-L. CYP450-mediated mitochondrial ROS production involved in arecoline N -oxide-induced oxidative damage in liver cell lines. Environ. Toxicol. 2018, 33, 1029–1038. [Google Scholar] [CrossRef]
- Chen, C.Y.; Yen, C.Y.; Wang, H.R.; Yang, H.P.; Tang, J.Y.; Huang, H.W.; Hsu, S.H.; Chang, H.W. Tenuifolide B from Cin-namomum tenuifolium stem selectively inhibits proliferation of oral cancer cells via apoptosis, ROS generation, mitochondrial depolarization, and DNA damage. Toxins 2016, 8, 319. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-W.; Li, R.-N.; Wang, H.-R.; Liu, J.-R.; Tang, J.-Y.; Huang, H.-W.; Chan, Y.-H.; Yen, C.-Y. Withaferin A Induces Oxidative Stress-Mediated Apoptosis and DNA Damage in Oral Cancer Cells. Front. Physiol. 2017, 8, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, C.C.; Chang, H.W.; Chuang, D.W.; Chang, F.R.; Chang, Y.C.; Cheng, Y.S.; Tsai, M.T.; Chen, W.Y.; Lee, S.S.; Wang, C.K.; et al. Fern plant-derived protoapigenone leads to DNA damage, apoptosis, and G(2)/M arrest in lung cancer cell line H1299. DNA Cell Biol. 2009, 28, 501–506. [Google Scholar] [CrossRef]
- Tang, J.Y.; Wu, C.Y.; Shu, C.W.; Wang, S.C.; Chang, M.Y.; Chang, H.W. A novel sulfonyl chromen-4-ones (CHW09) prefer-entially kills oral cancer cells showing apoptosis, oxidative stress, and DNA damage. Environ. Toxicol. 2018, 33, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Beltran, M.; Tavares, M.; Justin, N.; Khandelwal, G.; Ambrose, J.; Foster, B.M.; Worlock, K.B.; Tvardovskiy, A.; Kunzelmann, S.; Herrero, J.; et al. G-tract RNA removes Polycomb repressive complex 2 from genes. Nat. Struct. Mol. Biol. 2019, 26, 899–909. [Google Scholar] [CrossRef]
- Yen, C.-Y.; Huang, C.-Y.; Hou, M.-F.; Yang, Y.-H.; Chang, C.-H.; Huang, H.-W.; Chen, C.-H.; Chang, H.-W. Evaluating the performance of fibronectin 1 (FN1), integrin α4β1 (ITGA4), syndecan-2 (SDC2), and glycoprotein CD44 as the potential biomarkers of oral squamous cell carcinoma (OSCC). Biomarkers 2012, 18, 63–72. [Google Scholar] [CrossRef]
- Hooten, N.N.; Evans, M.K. Techniques to Induce and Quantify Cellular Senescence. J. Vis. Exp. 2017, e55533. [Google Scholar] [CrossRef]
- Chen, J.Y.-F.; Hwang, C.-C.; Chen, W.-Y.; Lee, J.-C.; Fu, T.-F.; Fang, K.; Chu, Y.-C.; Huang, Y.-L.; Lin, J.-C.; Tsai, W.-H.; et al. Additive effects of C2-ceramide on paclitaxel-induced premature senescence of human lung cancer cells. Life Sci. 2010, 87, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Yoshihashi, K.; Suzuki, H.; Tsutsumi, S.; Mutoh, H.; Maeda, S.; Yamagata, Y.; Seto, Y.; Aburatani, H.; Hatakeyama, M. CDX1 confers intestinal phenotype on gastric epithelial cells via induction of stemness-associated reprogramming factors SALL4 and KLF5. Proc. Natl. Acad. Sci.USA 2012, 109, 20584–20589. [Google Scholar] [CrossRef] [Green Version]
- Laddha, N.C.; Dwivedi, M.; Mansuri, M.S.; Singh, M.; Patel, H.H.; Agarwal, N.; Shah, A.M.; Begum, R. Association of Neuropeptide Y (NPY), Interleukin-1B (IL1B) Genetic Variants and Correlation of IL1B Transcript Levels with Vitiligo Susceptibility. PLoS ONE 2014, 9, e107020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.Y.; Huang, H.W.; Wang, H.R.; Chan, Y.C.; Haung, J.W.; Shu, C.W.; Wu, Y.C.; Chang, H.W. 4beta-Hydroxywithanolide E selectively induces oxidative DNA damage for selective killing of oral cancer cells. Environ. Toxicol. 2018, 33, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Bahreyni-Toossi, M.T.; Azimian, H.; Aghaee-Bakhtiari, S.H.; Mahmoudi, M.; Sadat-Darbandi, M.; Zafari, N. Radiation-induced DNA damage and altered expression of p21, cyclin D1 and Mre11 genes in human fibroblast cell lines with different radio-sensitivity. Mutat. Res. 2021, 823, 111760. [Google Scholar] [CrossRef] [PubMed]
- Omari Shekaftik, S.; Nasirzadeh, N. 8-Hydroxy-2′-deoxyguanosine (8-OHdG) as a biomarker of oxidative DNA damage in-duced by occupational exposure to nanomaterials: A systematic review. Nanotoxicology 2021, 15, 850–864. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′ -deoxyguanosine (8-OHdG): A Critical Biomarker of Oxidative Stress and Carcinogenesis. J. Environ. Sci. Health Part C 2009, 27, 120–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, K.; Virta, H. Growing Madin-Darby Canine Kidney Cells for Studying Epithelial Cell Biology. In Cell Biology; Celis, J.E., Ed.; Academic Press: Burlington, VT, USA; Elsevier BV: Amsterdam, The Netherlands, 2006; pp. 127–131. [Google Scholar]
- Ou-Yang, F.; Tsai, I.H.; Tang, J.Y.; Yen, C.Y.; Cheng, Y.B.; Farooqi, A.A.; Chen, S.R.; Yu, S.Y.; Kao, J.K.; Chang, H.W. Anti-proliferation for breast cancer cells by ethyl acetate extract of Nepenthes thorellii × (ventricosa × maxima). Int. J. Mol. Sci. 2019, 20, 3238. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.J.; Tang, J.Y.; Lin, L.C.; Lien, W.J.; Cheng, Y.B.; Chang, F.R.; Ou-Yang, F.; Chang, H.W. Withanolide C inhibits prolifer-ation of breast cancer cells via oxidative stress-mediated apoptosis and DNA damage. Antioxidants 2020, 9, 876. [Google Scholar] [CrossRef]
- Kabała-Dzik, A.; Rzepecka-Stojko, A.; Kubina, R.; Wojtyczka, R.D.; Buszman, E.; Stojko, J. Caffeic Acid Versus Caffeic Acid Phenethyl Ester in the Treatment of Breast Cancer MCF-7 Cells: Migration Rate Inhibition. Integr. Cancer Ther. 2018, 17, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Hamid, S.S.; Motaghed, M.; Al-Hassan, F.M. Cellular responses with thymoquinone treatment in human breast cancer cell line MCF-7. Pharmacogn. Res. 2013, 5, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Jing, Z.; Li, Y.; Mao, W. Berberine in combination with cisplatin suppresses breast cancer cell growth through in-duction of DNA breaks and caspase-3-dependent apoptosis. Oncol. Rep. 2016, 36, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Badran, A.; Tul-Wahab, A.; Zafar, H.; Mohammad, N.; Imad, R.; Ashfaq Khan, M.; Baydoun, E.; Choudhary, M.I. Antipsy-chotics drug aripiprazole as a lead against breast cancer cell line (MCF-7) in vitro. PLoS ONE 2020, 15, e0235676. [Google Scholar] [CrossRef]
- Duarte, D.; Vale, N. New Trends for Antimalarial Drugs: Synergism between Antineoplastics and Antimalarials on Breast Cancer Cells. Biomolecules 2020, 10, 1623. [Google Scholar] [CrossRef] [PubMed]
- Mansingh, D.P.; Sunanda, O.J.; Sali, V.K.; Vasanthi, H.R. [6]-Gingerol-induced cell cycle arrest, reactive oxygen species generation, and disruption of mitochondrial membrane potential are associated with apoptosis in human gastric cancer (AGS) cells. J. Biochem. Mol. Toxicol. 2018, 32, e22206. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.-H.; Ni, Z.-J.; Zhang, F.; Zhang, Y.-Y.; Liu, M.-M.; Thakur, K.; Zhang, J.-G.; Wang, S.Y.; Wei, Z.-J. 6-Shogaol mediated ROS production and apoptosis via endoplasmic reticulum and mitochondrial pathways in human endometrial carcinoma Ishikawa cells. J. Funct. Foods 2020, 74, 104178. [Google Scholar] [CrossRef]
- Su, P.; Veeraraghavan, V.P.; Krishna Mohan, S.; Lu, W. A ginger derivative, zingerone-a phenolic compound-induces ROS-mediated apoptosis in colon cancer cells (HCT-116). J. Biochem. Mol. Toxicol. 2019, 33, e22403. [Google Scholar] [CrossRef]
- Bohr, V.A.; Dianov, G.L. Oxidative DNA damage processing in nuclear and mitochondrial DNA. Biochimie 1999, 81, 155–160. [Google Scholar] [CrossRef]
- Salehi, F.; Behboudi, H.; Kavoosi, G.; Ardestani, S.K. Oxidative DNA damage induced by ROS-modulating agents with the ability to target DNA: A comparison of the biological characteristics of citrus pectin and apple pectin. Sci. Rep. 2018, 8, 13902. [Google Scholar] [CrossRef]
- Zhu, M.-J.; Wang, X.; Shi, L.; Liang, L.-Y.; Wang, Y. Senescence, oxidative stress and mitochondria dysfunction. Cancer Med. Res. Innov. 2018, 2, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Te Poele, R.H.; Okorokov, A.L.; Jardine, L.; Cummings, J.; Joel, S.P. DNA damage is able to induce senescence in tumor cells in vitro and in vivo. Cancer Res. 2002, 62, 1876–1883. [Google Scholar]
- Moiseeva, O.; Bourdeau, V.; Roux, A.; Deschenes-Simard, X.; Ferbeyre, G. Mitochondrial dysfunction contributes to onco-gene-induced senescence. Mol. Cell. Biol. 2009, 29, 4495–4507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaewtunjai, N.; Wongpoomchai, R.; Imsumran, A.; Pompimon, W.; Athipornchai, A.; Suksamrarn, A.; Lee, T.R.; Tuntiwechapikul, W. Ginger Extract Promotes Telomere Shortening and Cellular Senescence in A549 Lung Cancer Cells. ACS Omega 2018, 3, 18572–18581. [Google Scholar] [CrossRef] [PubMed]
- Mohd Sahardi, N.F.N.; Jaafar, F.; Mad Nordin, M.F.; Makpol, S. Zingiber officinale Roscoe prevents cellular senescence of my-oblasts in culture and promotes muscle regeneration. Evid. Based Complement. Altern. Med. 2020, 2020, 1787342. [Google Scholar] [CrossRef] [PubMed]
- Han, H.S.; Kim, K.B.; Jung, J.H.; An, I.S.; Kim, Y.-J.; An, S. Anti-apoptotic, antioxidant and anti-aging effects of 6-shogaol on human dermal fibroblasts. Biomed. Dermatol. 2018, 2, 27. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primers (5′→3′) | Reverse Primers (5′→3′) | Length |
|---|---|---|---|
| EDN1 | CAGCAGTCTTAGGCGCTGAG | ACTCTTTATCCATCAGGGACGAG | 126 bp |
| ANKRD1 | AGTAGAGGAACTGGTCACTGG | TGGGCTAGAAGTGTCTTCAGAT | 138 bp |
| CDKN1A | GACACCACTGGAGGGTGACT | CAGGTCCACATGGTCTTCCT | 172 bp |
| SERPINE1 | GTGTTTCAGCAGGTGGCGC | CCGGAACAGCCTGAAGAAGTG | 300 bp |
| TAGLN | TGGCGTGATTCTGAGCAAG | ACTGCCAAGCTGCCCAA | 240 bp |
| GAPDH | CCTCAACTACATGGTTTACATGTTCC | CAAATGAGCCCCAGCCTTCT | 220 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, T.-J.; Tang, J.-Y.; Shiau, J.-P.; Hou, M.-F.; Yen, C.-H.; Ou-Yang, F.; Chen, C.-Y.; Chang, H.-W. Gingerenone A Induces Antiproliferation and Senescence of Breast Cancer Cells. Antioxidants 2022, 11, 587. https://doi.org/10.3390/antiox11030587
Yu T-J, Tang J-Y, Shiau J-P, Hou M-F, Yen C-H, Ou-Yang F, Chen C-Y, Chang H-W. Gingerenone A Induces Antiproliferation and Senescence of Breast Cancer Cells. Antioxidants. 2022; 11(3):587. https://doi.org/10.3390/antiox11030587
Chicago/Turabian StyleYu, Tzu-Jung, Jen-Yang Tang, Jun-Ping Shiau, Ming-Feng Hou, Chia-Hung Yen, Fu Ou-Yang, Chung-Yi Chen, and Hsueh-Wei Chang. 2022. "Gingerenone A Induces Antiproliferation and Senescence of Breast Cancer Cells" Antioxidants 11, no. 3: 587. https://doi.org/10.3390/antiox11030587
APA StyleYu, T.-J., Tang, J.-Y., Shiau, J.-P., Hou, M.-F., Yen, C.-H., Ou-Yang, F., Chen, C.-Y., & Chang, H.-W. (2022). Gingerenone A Induces Antiproliferation and Senescence of Breast Cancer Cells. Antioxidants, 11(3), 587. https://doi.org/10.3390/antiox11030587