
Evaluation of Label-Free Confocal Raman Microspectroscopy for Monitoring Oxidative Stress In Vitro in Live Human Cancer Cells




Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Preparation for Microspectroscopy
2.2. Fluorescence Data Acquisition and Analysis
2.3. Raman Data Acquisition and Analysis
3. Results
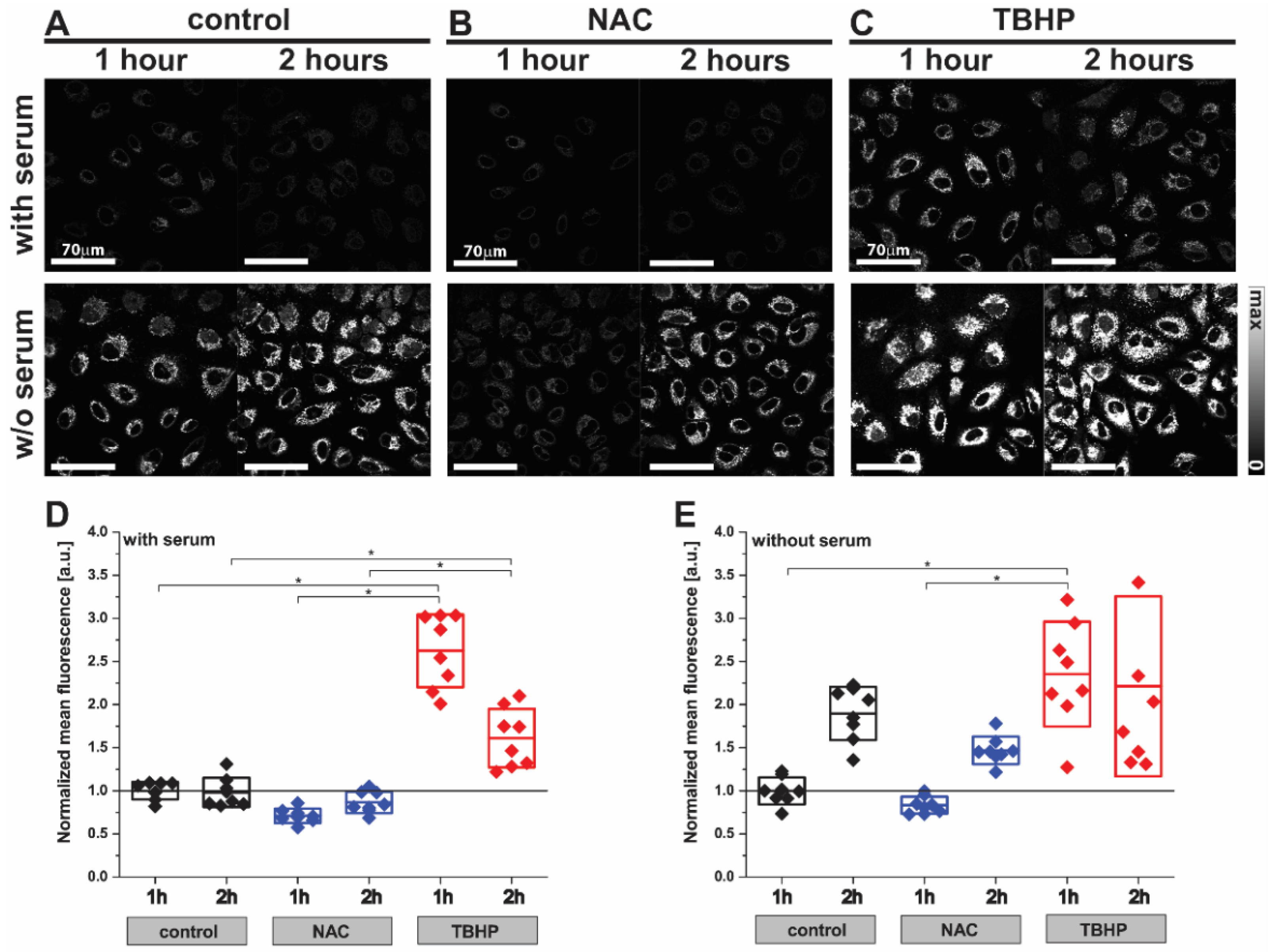
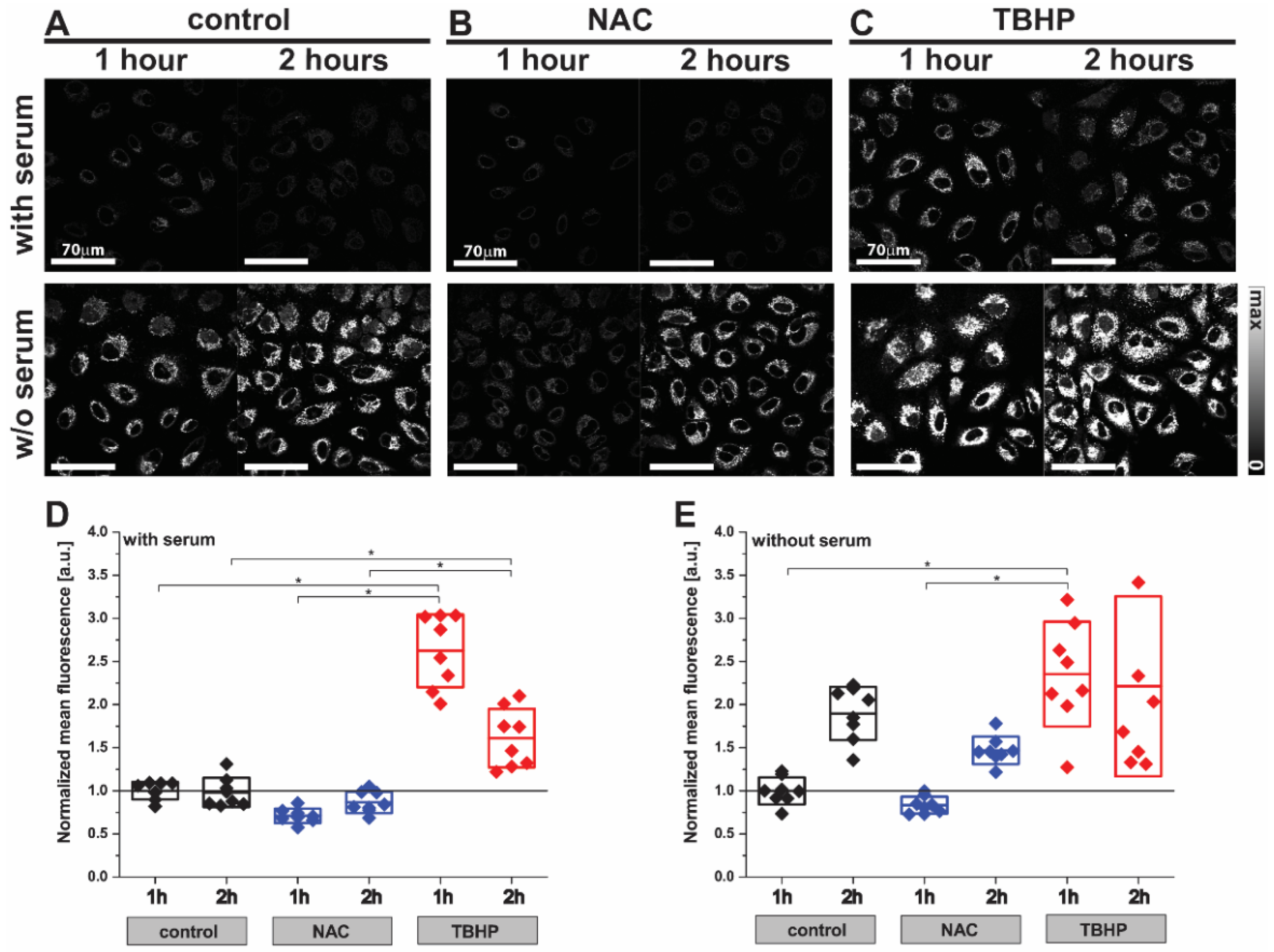
3.1. Fluorescence Imaging Reveals Changes in the Levels of Oxidative Stress in A549 Cells with Antioxidant or Pro-Oxidant Treatment
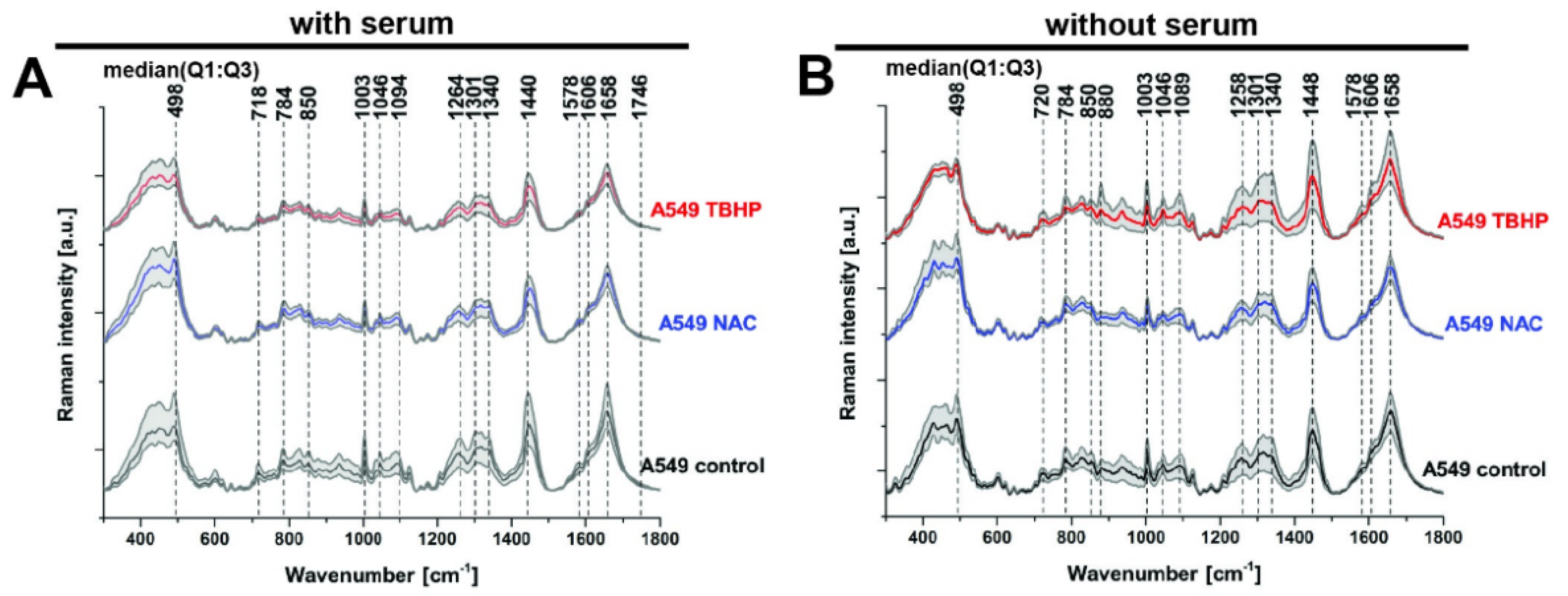
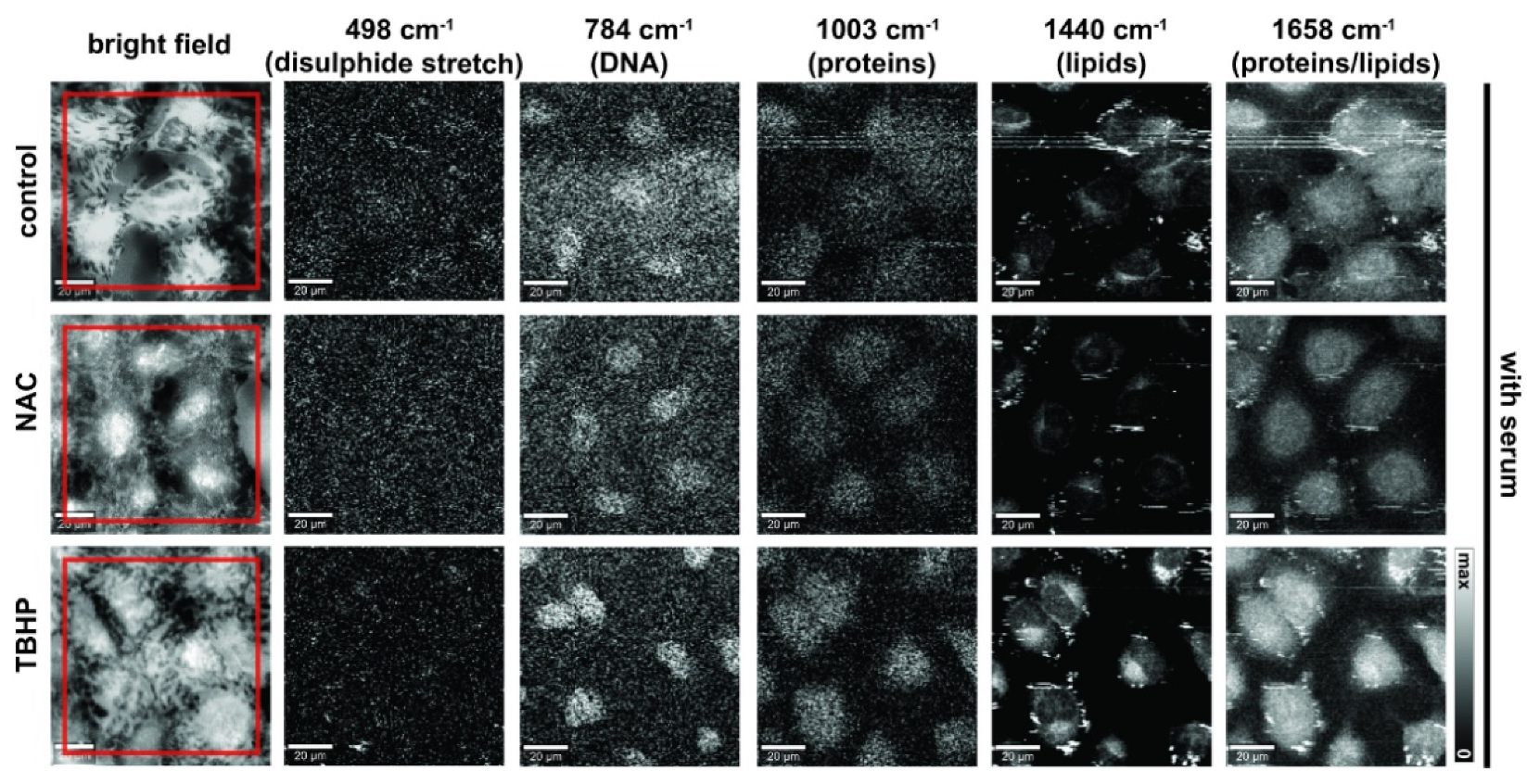
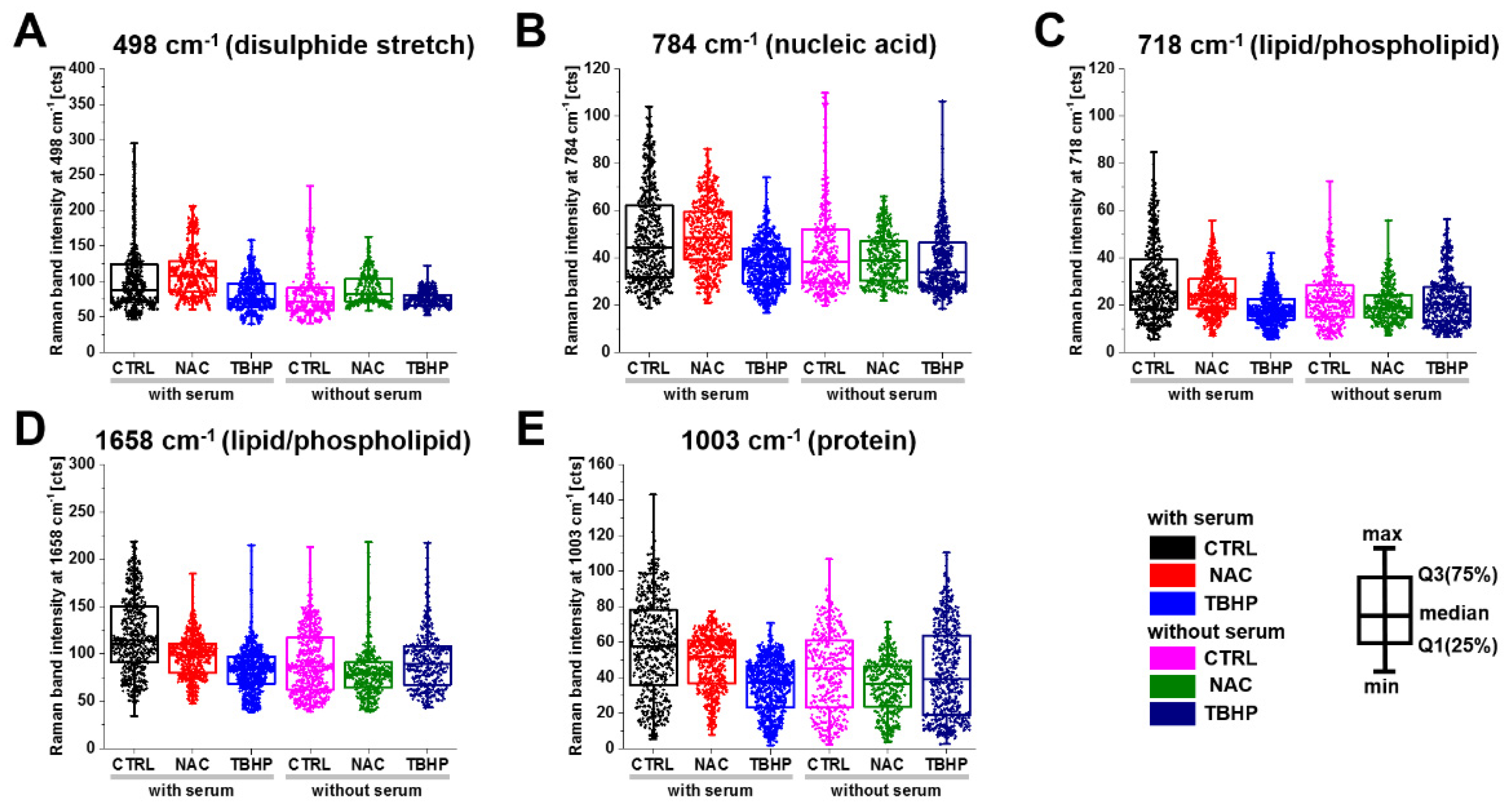
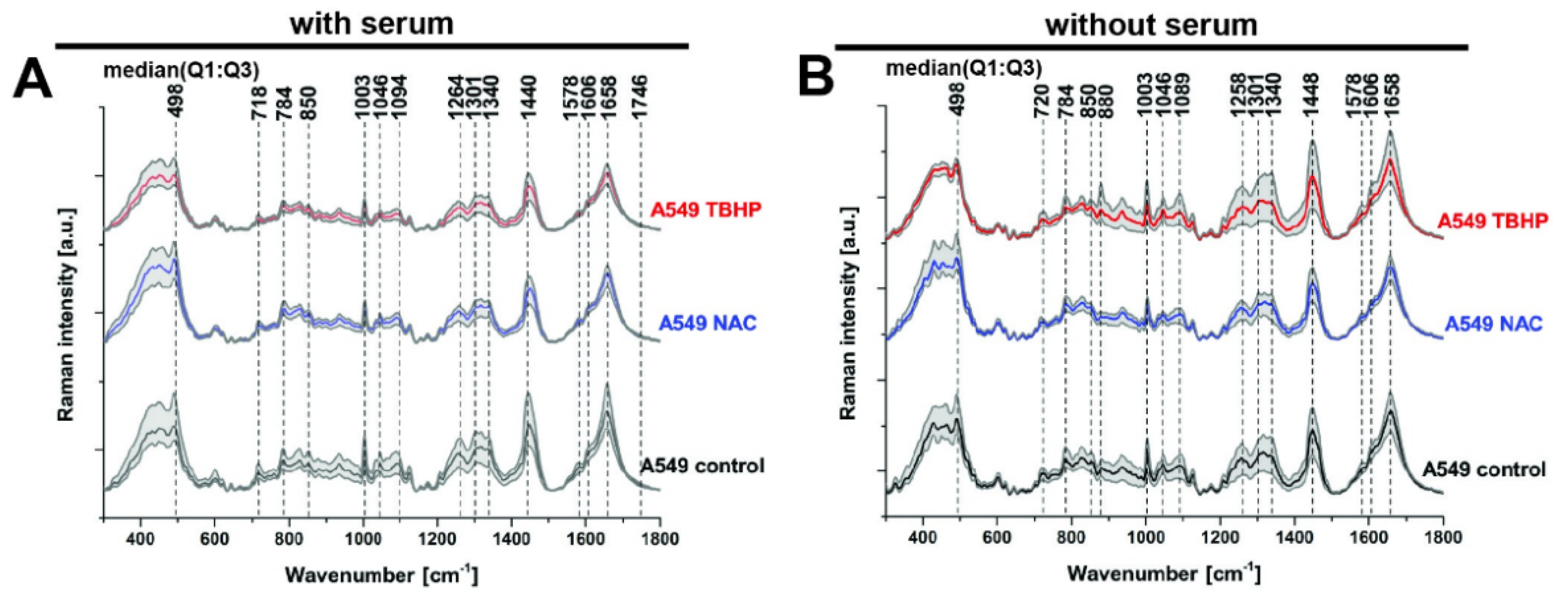
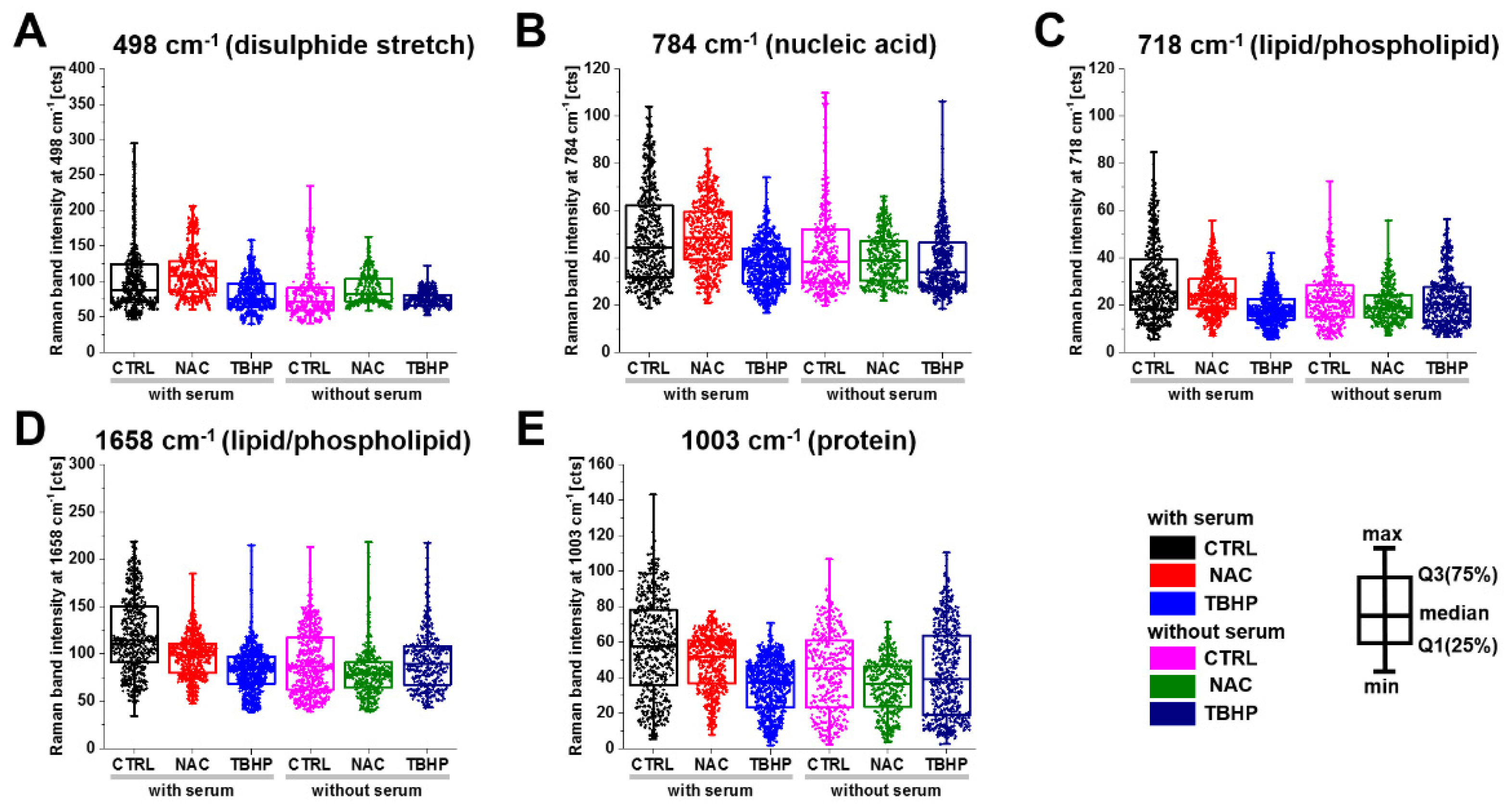
3.2. Raman Microspectroscopy and Imaging Indicate the Major Vibrational Modes That Are Altered under Different Levels of Oxidative Stress
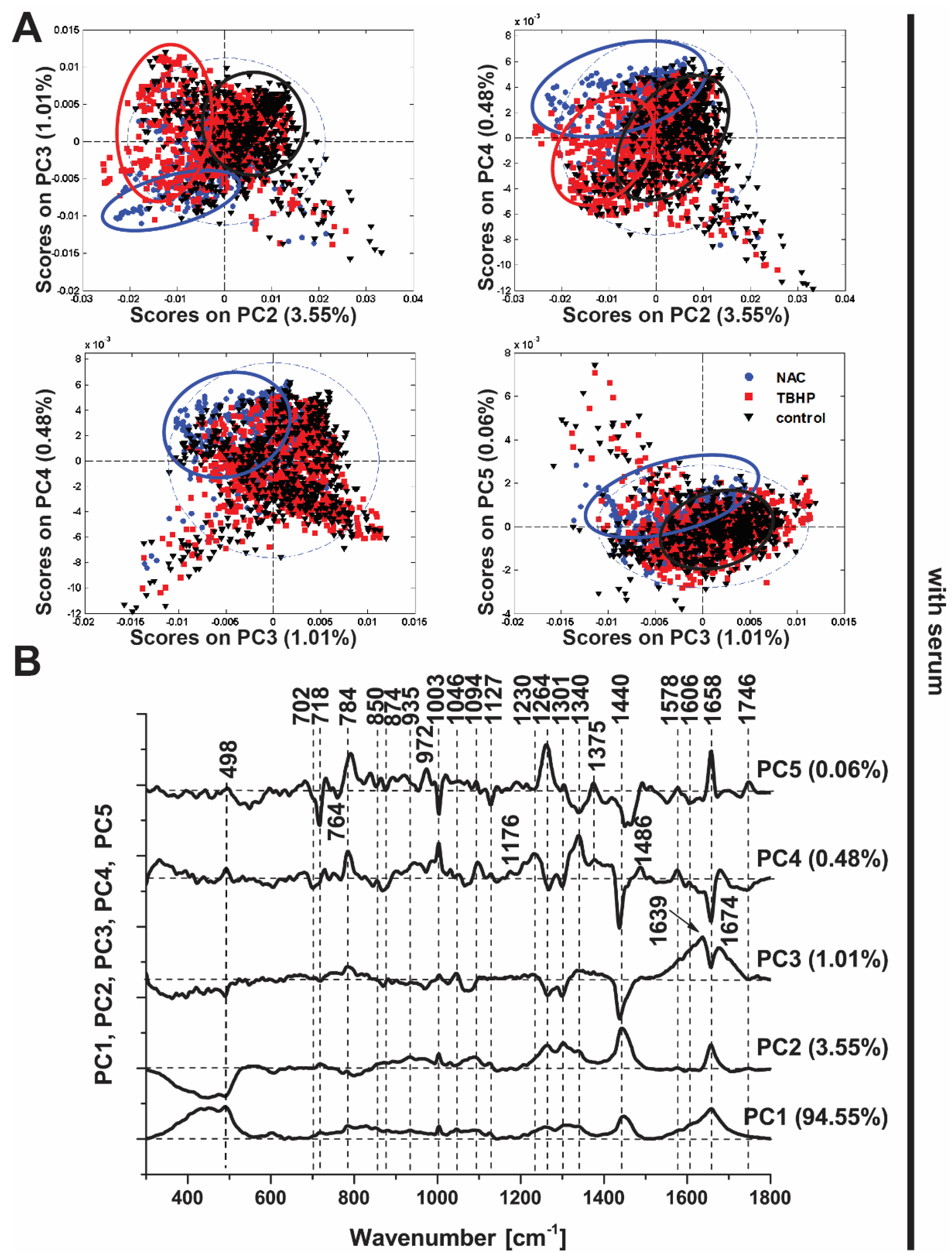
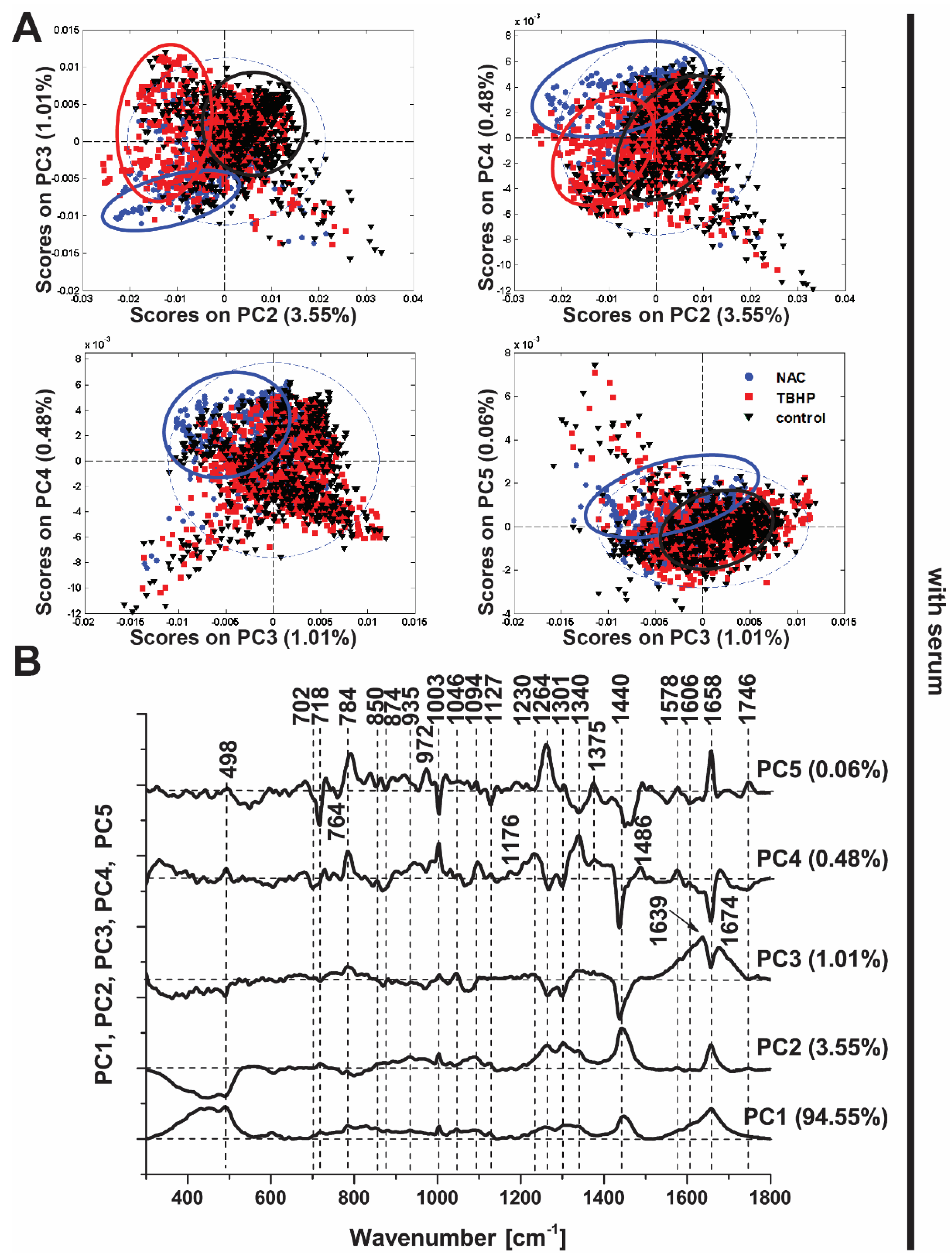
3.3. Principal Components Analysis Confirms the Discrimination Power of Raman Spectroscopy
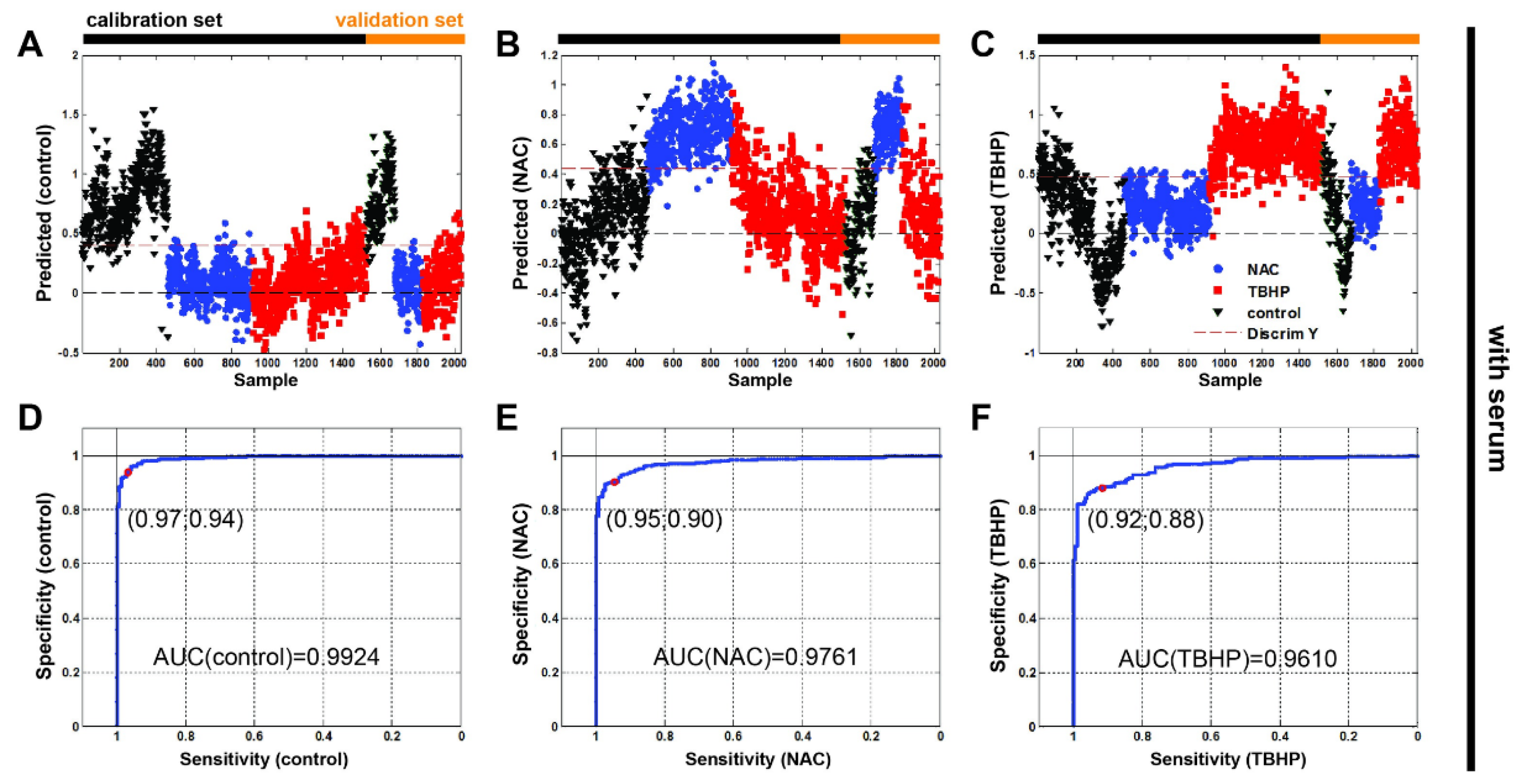
3.4. Partial Least Squares–Discriminant Analysis Shows Good Classification Performance for Antioxidant and Pro-Oxidant Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacob, C.; Doering, M.; Burkholz, T. The Chemical Basis of Biological Redox Control. In Redox Signaling and Regulation in Biology and Medicine; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2009; pp. 63–122. ISBN 978-3-527-62758-5. [Google Scholar]
- Sies, H.; Jones, D.P. Reactive Oxygen Species (ROS) as Pleiotropic Physiological Signalling Agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R. Redox Metabolism and Life. In Redox Biochemistry; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; pp. 1–9. ISBN 978-0-470-17733-4. [Google Scholar]
- Iannitti, T.; Palmieri, B. Antioxidant Therapy and Its Effectiveness in Oxidative Stress-Mediated Disorders. In Oxidative Stress in Vertebrates and Invertebrates; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; pp. 209–234. [Google Scholar]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindoli, A.; Rigobello, M.P. Principles in Redox Signaling: From Chemistry to Functional Significance. Antioxid. Redox Signal. 2013, 18, 1557–1593. [Google Scholar] [CrossRef] [PubMed]
- Benz, C.C.; Yau, C. Ageing, Oxidative Stress and Cancer: Paradigms in Parallax. Nat. Rev. Cancer 2008, 8, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim Rizvi, S.; Bhooshan Pandey, K. Oxidative Stress and Aging: A Comparison between Vertebrates and Invertebrates. In Oxidative Stress in Vertebrates and Invertebrates; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011; pp. 167–174. ISBN 978-1-118-14814-3. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015; ISBN 978-0-19-871747-8. [Google Scholar]
- Cole, S.P.; Downes, H.F.; Mirski, S.E.; Clements, D.J. Alterations in Glutathione and Glutathione-Related Enzymes in a Multidrug-Resistant Small Cell Lung Cancer Cell Line. Mol. Pharmacol. 1990, 37, 192–197. [Google Scholar] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative Stress, Inflammation, and Cancer: How Are They Linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, R.A.; Harris, I.S.; Mak, T.W. Regulation of Cancer Cell Metabolism. Nat. Rev. Cancer 2011, 11, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balendiran, G.K.; Dabur, R.; Fraser, D. The Role of Glutathione in Cancer. Cell Biochem. Funct. 2004, 22, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Hevia, D.; Rey, S.; Riego, M.; Gomez, E.; Muñoz-Cimadevilla, H. P-133—Chronic Heart Failure (CHF): Thiol Oxidation and Antioxidant Capacity as Possible Biomarker (e-BQC). Free Radic. Biol. Med. 2018, 120, S85. [Google Scholar] [CrossRef]
- Kauffman, M.E.; Kauffman, M.K.; Traore, K.; Zhu, H.; Trush, M.A.; Jia, Z.; Li, Y.R. MitoSOX-Based Flow Cytometry for Detecting Mitochondrial ROS. React. Oxyg. Species 2016, 2, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Kiani-Esfahani, A.; Tavalaee, M.; Deemeh, M.R.; Hamiditabar, M.; Nasr-Esfahani, M.H. DHR123: An Alternative Probe for Assessment of ROS in Human Spermatozoa. Syst. Biol. Reprod. Med. 2012, 58, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhong, Z.; Xu, Z.; Chen, L.; Wang, Y. 2′,7′-Dichlorodihydrofluorescein as a Fluorescent Probe for Reactive Oxygen Species Measurement: Forty Years of Application and Controversy. Free Radic. Res. 2010, 44, 587–604. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, A.I.; Panova, A.S.; Kokova, A.D.; Kotova, D.A.; Maltsev, D.I.; Podgorny, O.V.; Belousov, V.V.; Bilan, D.S. In Vivo Imaging with Genetically Encoded Redox Biosensors. Int. J. Mol. Sci. 2020, 21, E8164. [Google Scholar] [CrossRef]
- de Bittner, G.C.V.; Dubikovskaya, E.A.; Bertozzi, C.R.; Chang, C.J. In Vivo Imaging of Hydrogen Peroxide Production in a Murine Tumor Model with a Chemoselective Bioluminescent Reporter. Proc. Natl. Acad. Sci. USA 2010, 107, 21316–21321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, J.; Bollepalli, L.; Belenguer, A.M.; Antonio, M.D.; De Mitri, N.; Joseph, J.; Balasubramanian, S.; Hunter, C.A.; Bohndiek, S.E. An Activatable Cancer-Targeted Hydrogen Peroxide Probe for Photoacoustic and Fluorescence Imaging. Cancer Res. 2019, 79, 5407–5417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.; Wright, K.L.; Ashton, L. Raman Spectroscopy: An Evolving Technique for Live Cell Studies. Analyst 2016, 141, 3590–3600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, H.J.; Ashton, L.; Bird, B.; Cinque, G.; Curtis, K.; Dorney, J.; Esmonde-White, K.; Fullwood, N.J.; Gardner, B.; Martin-Hirsch, P.L.; et al. Using Raman Spectroscopy to Characterize Biological Materials. Nat. Protoc. 2016, 11, 664–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surmacki, J.M.; Woodhams, B.J.; Haslehurst, A.; Ponder, B.A.J.; Bohndiek, S.E. Raman Micro-Spectroscopy for Accurate Identification of Primary Human Bronchial Epithelial Cells. Sci. Rep. 2018, 8, 12604. [Google Scholar] [CrossRef] [PubMed]
- Surmacki, J.M.; Gonzalez, I.Q.; Bohndiek, S.E. Application of Confocal Raman Micro-Spectroscopy for Label-Free Monitoring of Oxidative Stress in Living Bronchial Cells. In Biomedical Vibrational Spectroscopy 2018: Advances in Research and Industry; International Society for Optics and Photonics: Bellingham, WA, USA, 2018; Volume 10490, p. 1049002. [Google Scholar]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an Antioxidant and Disulphide Breaking Agent: The Reasons Why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Kučera, O.; Endlicher, R.; Roušar, T.; Lotková, H.; Garnol, T.; Drahota, Z.; Červinková, Z. The Effect of Tert-Butyl Hydroperoxide-Induced Oxidative Stress on Lean and Steatotic Rat Hepatocytes In Vitro. Oxid. Med. Cell. Longev. 2014, 2014, e752506. [Google Scholar] [CrossRef] [Green Version]
- Hix, S.; Kadiiska, M.B.; Mason, R.P.; Augusto, O. In Vivo Metabolism of Tert-Butyl Hydroperoxide to Methyl Radicals. EPR Spin-Trapping and DNA Methylation Studies. Chem. Res. Toxicol. 2000, 13, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide Metabolism in Mammalian Organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar] [CrossRef] [PubMed]
- Masaki, N.; Kyle, M.E.; Farber, J.L. Tert-Butyl Hydroperoxide Kills Cultured Hepatocytes by Peroxidizing Membrane Lipids. Arch. Biochem. Biophys. 1989, 269, 390–399. [Google Scholar] [CrossRef]
- Castilho, R.F.; Kowaltowski, A.J.; Meinicke, A.R.; Bechara, E.J.; Vercesi, A.E. Permeabilization of the Inner Mitochondrial Membrane by Ca2+ Ions Is Stimulated by T-Butyl Hydroperoxide and Mediated by Reactive Oxygen Species Generated by Mitochondria. Free Radic. Biol. Med. 1995, 18, 479–486. [Google Scholar] [CrossRef]
- Sandström, B.E.; Marklund, S.L. Effects of Variation in Glutathione Peroxidase Activity on DNA Damage and Cell Survival in Human Cells Exposed to Hydrogen Peroxide and T-Butyl Hydroperoxide. Biochem. J. 1990, 271, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, M.; Coombs, K.M. Serum-Reduced Media Impacts on Cell Viability and Protein Expression in Human Lung Epithelial Cells. J. Cell. Physiol. 2019, 234, 7718–7724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Generating and Detecting Reactive Oxygen Species—Section 18.2—PL. Available online: //www.thermofisher.com/uk/en/home/references/molecular-probes-the-handbook/probes-for-reactive-oxygen-species-including-nitric-oxide/generating-and-detecting-reactive-oxygen-species.html (accessed on 29 November 2021).
- Miura, T.; Takeuchi, H.; Harada, I. Characterization of Individual Tryptophan Side Chains in Proteins Using Raman Spectroscopy and Hydrogen-Deuterium Exchange Kinetics. Biochemistry 1988, 27, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Bastian, E.J.; Martin, R.B. Disulfide Vibrational Spectra in the Sulfur-Sulfur and Carbon-Sulfur Stretching Region. J. Phys. Chem. 1973, 77, 1129–1133. [Google Scholar] [CrossRef]
- Rygula, A.; Majzner, K.; Marzec, K.M.; Kaczor, A.; Pilarczyk, M.; Baranska, M. Raman Spectroscopy of Proteins: A Review. J. Raman Spectrosc. 2013, 44, 1061–1076. [Google Scholar] [CrossRef]
- Schulze, H.G.; Konorov, S.O.; Piret, J.M.; Blades, M.W.; Turner, R.F.B. Label-Free Imaging of Mammalian Cell Nucleoli by Raman Microspectroscopy. Analyst 2013, 138, 3416–3423. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.; Kendall, C.; Smith, J.; Crow, P.; Barr, H. Raman Spectroscopy for Identification of Epithelial Cancers. Faraday Discuss. 2004, 126, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Puppels, G.J.; de Mul, F.F.; Otto, C.; Greve, J.; Robert-Nicoud, M.; Arndt-Jovin, D.J.; Jovin, T.M. Studying Single Living Cells and Chromosomes by Confocal Raman Microspectroscopy. Nature 1990, 347, 301–303. [Google Scholar] [CrossRef] [PubMed]
- Boren, J.; Brindle, K.M. Apoptosis-Induced Mitochondrial Dysfunction Causes Cytoplasmic Lipid Droplet Formation. Cell Death Differ. 2012, 19, 1561–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolliffe, I.T.; Cadima, J. Principal Component Analysis: A Review and Recent Developments. Philos. Trans. R. Soc. Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, K.; Jakob, U. The Role of Thiols in Antioxidant Systems. Free Radic. Biol. Med. 2019, 140, 14–27. [Google Scholar] [CrossRef]
- Requejo, R.; Hurd, T.R.; Costa, N.J.; Murphy, M.P. Cysteine Residues Exposed on Protein Surfaces Are the Dominant Intramitochondrial Thiol and May Protect against Oxidative Damage. FEBS J. 2010, 277, 1465–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafarullah, M.; Li, W.Q.; Sylvester, J.; Ahmad, M. Molecular Mechanisms of N-Acetylcysteine Actions. Cell. Mol. Life Sci. CMLS 2003, 60, 6–20. [Google Scholar] [CrossRef]
- Kim, K.Y.; Rhim, T.; Choi, I.; Kim, S.S. N-Acetylcysteine Induces Cell Cycle Arrest in Hepatic Stellate Cells through Its Reducing Activity. J. Biol. Chem. 2001, 276, 40591–40598. [Google Scholar] [CrossRef] [Green Version]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA Damage: Mechanisms, Mutation, and Disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef] [Green Version]
- Dutta, A.; Gautam, R.; Chatterjee, S.; Ariese, F.; Sikdar, S.K.; Umapathy, S. Ascorbate Protects Neurons against Oxidative Stress: A Raman Microspectroscopic Study. ACS Chem. Neurosci. 2015, 6, 1794–1801. [Google Scholar] [CrossRef]
- Sahm, F.; Oezen, I.; Opitz, C.A.; Radlwimmer, B.; von Deimling, A.; Ahrendt, T.; Adams, S.; Bode, H.B.; Guillemin, G.J.; Wick, W.; et al. The Endogenous Tryptophan Metabolite and NAD + Precursor Quinolinic Acid Confers Resistance of Gliomas to Oxidative Stress. Cancer Res. 2013, 73, 3225–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ermakov, I.V.; Sharifzadeh, M.; Ermakova, M.R.; Gellermann, W. Resonance Raman Detection of Carotenoid Antioxidants in Living Human Tissue. J. Biomed. Opt. 2005, 10, 064028. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Castellano, A.; Díaz-Quintana, A.; Pérez-Mejías, G.; Elena-Real, C.A.; González-Arzola, K.; García-Mauriño, S.M.; la Rosa, M.A.D.; Díaz-Moreno, I. Oxidative Stress Is Tightly Regulated by Cytochrome c Phosphorylation and Respirasome Factors in Mitochondria. Proc. Natl. Acad. Sci. USA 2018, 115, 7955–7960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramczyk, H.; Brozek-Pluska, B.; Kopec, M.; Surmacki, J.; Błaszczyk, M.; Radek, M. Redox Imbalance and Biochemical Changes in Cancer by Probing Redox-Sensitive Mitochondrial Cytochromes in Label-Free Visible Resonance Raman Imaging. Cancers 2021, 13, 960. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raman Band Wavenumber (cm−1) | Raman Vibrational Mode Assignment |
|---|---|
| 498 | S-S disulphide stretching [35,36] |
| 718 | CN+-(CH3)3 symmetric stretching, phospholipids [37,38] |
| 784 | Cytosine, uracil, thymine, pyrimidine bases, ring breathing modes in DNA bases [37,38,39] |
| 880 | Indole ring mode of tryptophan [34] |
| 1003 | Phenylalanine, proline, symmetric stretching (ring breathing) mode of phenyl group [36,37] |
| 1094 | Symmetric PO2− stretching mode of the DNA backbone [38,39] |
| 1264 | =CH deformation, triglycerides (fatty acids), lipids [38] |
| 1301 | CH2 twist, triglycerides (fatty acids), lipids [38] |
| 1440 | CH2 and CH3 deformations, lipids [38] |
| 1606 | Tyrosine, phenylalanine ring vibration C=C bending, cytosine NH2, protein [36,38] |
| 1658 | Amide I, C=O stretching mode, peptide linkage; C=C stretching, lipids [36,38] |
| 1746 | C=O stretching, ester group of lipids and phospholipids [38] |
| Treatment Condition | Sensitivity | Specificity | RMSEC | RMSECV | RMSEP |
|---|---|---|---|---|---|
| Control | 96.7 | 93.5 | 0.2576 | 0.2629 | 0.2465 |
| NAC | 94.8 | 90.1 | 0.3059 | 0.3101 | 0.2930 |
| TBHP | 91.6 | 87.9 | 0.3262 | 0.3299 | 0.3278 |
| Control without serum | 89.0 | 91.7 | 0.2721 | 0.2755 | 0.2784 |
| NAC without serum | 97.0 | 99.6 | 0.2343 | 0.2375 | 0.2409 |
| TBHP without serum | 95.4 | 94.5 | 0.2744 | 0.2774 | 0.2791 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surmacki, J.M.; Quiros-Gonzalez, I.; Bohndiek, S.E. Evaluation of Label-Free Confocal Raman Microspectroscopy for Monitoring Oxidative Stress In Vitro in Live Human Cancer Cells. Antioxidants 2022, 11, 573. https://doi.org/10.3390/antiox11030573
Surmacki JM, Quiros-Gonzalez I, Bohndiek SE. Evaluation of Label-Free Confocal Raman Microspectroscopy for Monitoring Oxidative Stress In Vitro in Live Human Cancer Cells. Antioxidants. 2022; 11(3):573. https://doi.org/10.3390/antiox11030573
Chicago/Turabian StyleSurmacki, Jakub Maciej, Isabel Quiros-Gonzalez, and Sarah Elizabeth Bohndiek. 2022. "Evaluation of Label-Free Confocal Raman Microspectroscopy for Monitoring Oxidative Stress In Vitro in Live Human Cancer Cells" Antioxidants 11, no. 3: 573. https://doi.org/10.3390/antiox11030573
APA StyleSurmacki, J. M., Quiros-Gonzalez, I., & Bohndiek, S. E. (2022). Evaluation of Label-Free Confocal Raman Microspectroscopy for Monitoring Oxidative Stress In Vitro in Live Human Cancer Cells. Antioxidants, 11(3), 573. https://doi.org/10.3390/antiox11030573