
A Dynamic Model for Estimating the Interaction of ROS–PUFA–Antioxidants in Rabbit

 ,
,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
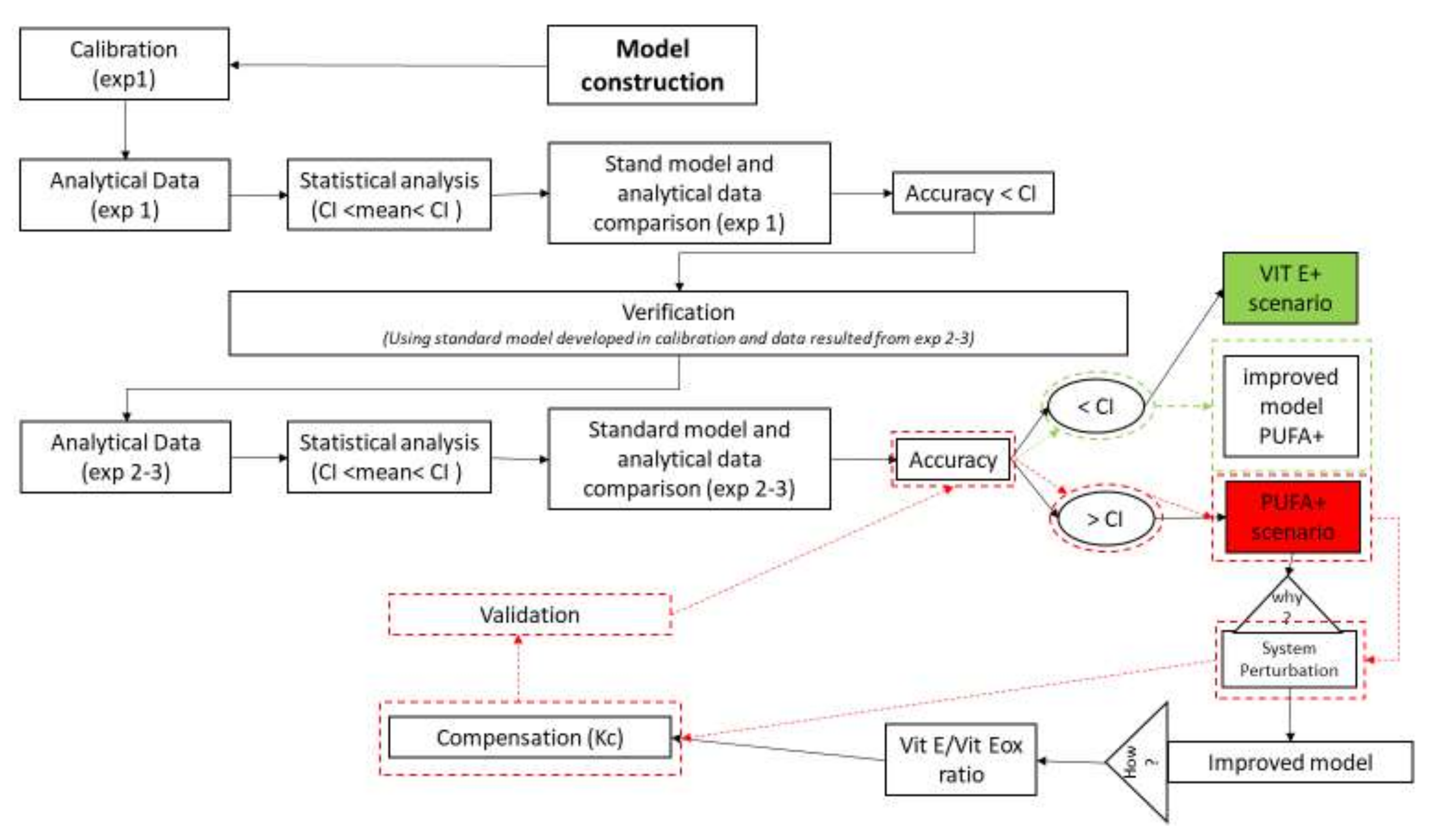
2.1. Cognitive Map Used for Definition, Calibration and Validation of the Model
2.1.1. Problem Entity
2.1.2. Conceptual Model
2.1.3. Structural Model
Sub–Model 1: ROS Production
Sub–Model 2: Oxidative Thrust
Sub–Model 3: Antioxidant Defence
2.2. Verification and Validation of the Model
2.3. Experimental Plan: Animals and Dietary Treatments
- ○
- High VITAMIN E (VIT E+): 10 rabbits of the same age and breed mentioned above were fed ad libitum the standard diet (50 mg/kg alpha–tocopheryl acetate by vitamin–mineral premix), including 200 mg/kg alpha–tocopheryl acetate.
- ○
- High PUFA (PUFA+): 10 rabbits of the same age and breed mentioned above were fed ad libitum a standard diet supplemented with 10% extruded flaxseed and 200 mg/kg alpha–tocopheryl acetate, following the rabbit requirements for a flaxseed enriched diet [53]. This diet had a 1.34–fold higher concentration of ALA than the standard diet.
2.4. Samples Collection
2.5. Analytical Determinations
2.5.1. Determination of the Fatty Acids
2.5.2. Determination of Vit E and AA
2.5.3. Determination of ROS, Malondialdehyde, F2–Isoprostane, F3–Isoprostane, and F4–Neuroprostane
2.6. Statistical Evaluation
3. Results
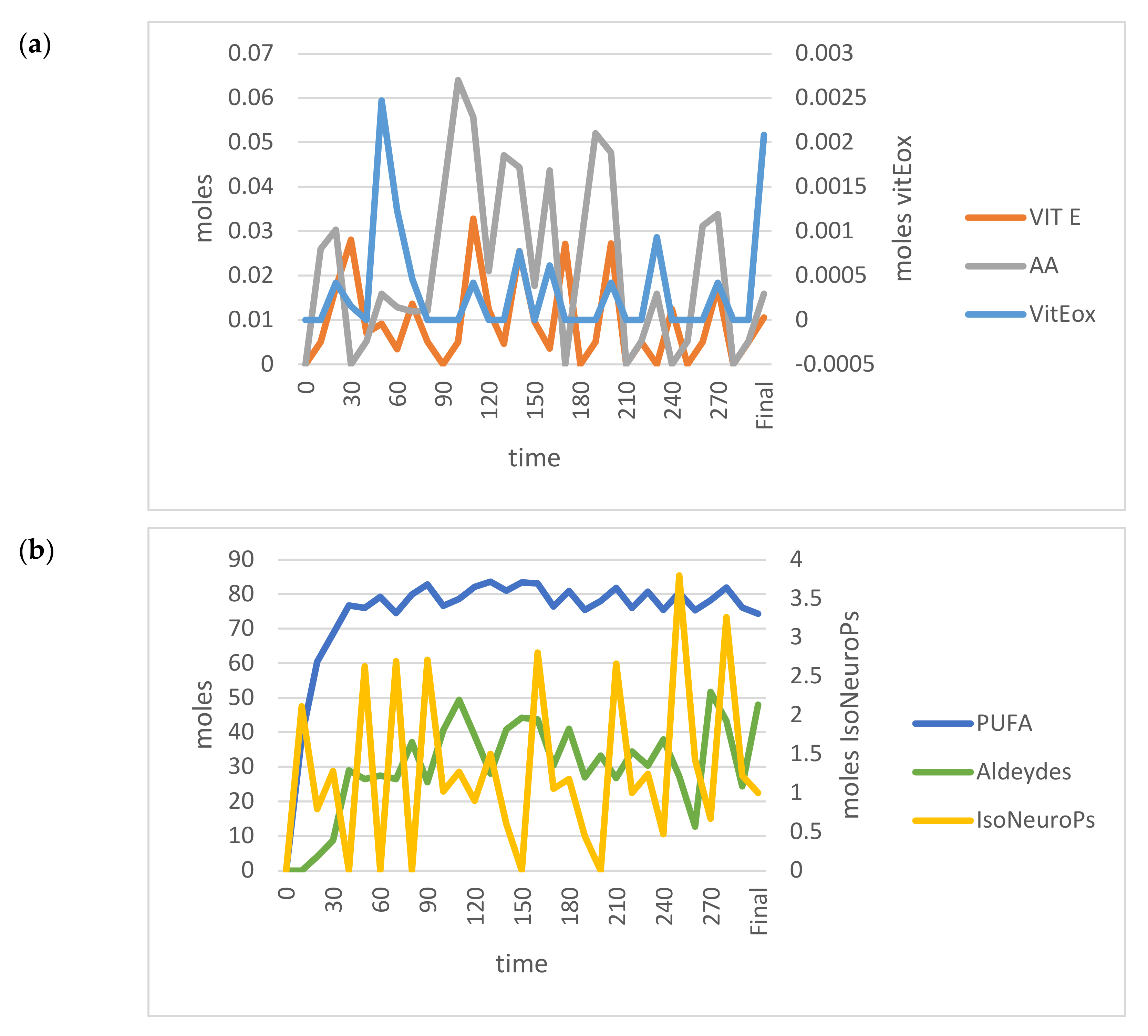
3.1. Calibration of the Standard Model
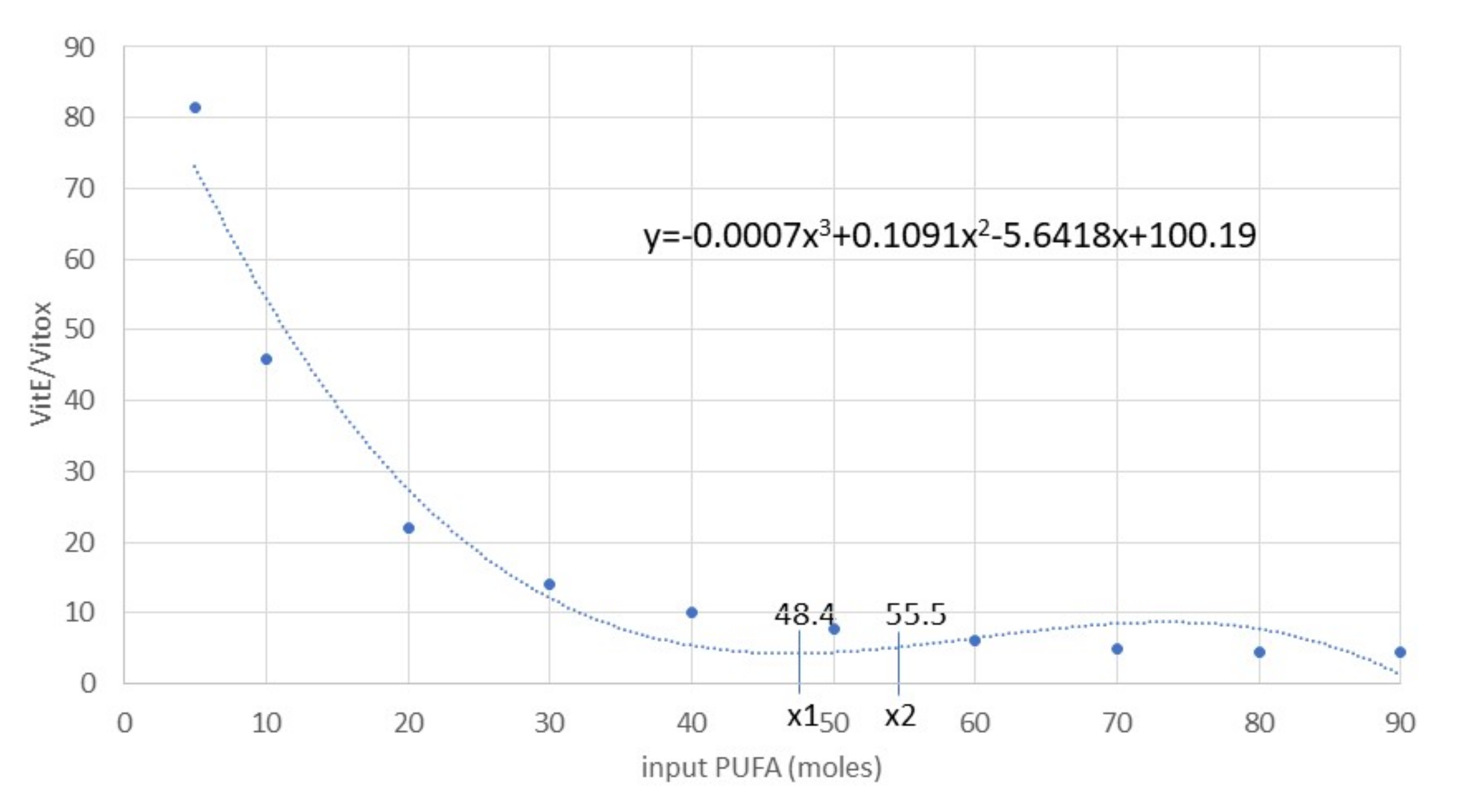
3.2. Model Validation in the VIT E+ and PUFA+ Scenarios
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Units | Standard | VIT E+ | PUFA + | |
|---|---|---|---|---|
| Ingredients | ||||
| Dehydrated alfalfa meal | g/kg | 300 | 300 | 380 |
| Soybean meal 44% | g/kg | 150 | 150 | 100 |
| Barley meal | g/kg | 410 | 410 | 310 |
| Wheat bran | g/kg | 52 | 52 | 52 |
| Soybean oil | g/kg | 30 | 30 | – |
| Extruded flaxseed | g/kg | – | – | 100 |
| Beet molasses | g/kg | 20 | 20 | 10 |
| Calcium carbonate | g/kg | 7 | 7 | 7 |
| Calcium diphosphate | g/kg | 13.5 | 13.5 | 13.5 |
| Salt | g/kg | 7 | 7 | 7 |
| DL–methionine | g/kg | 0.5 | 0.5 | 0.5 |
| Vitamin–mineral premix 1 | g/kg | 10 | 10 | 10 |
| α–tocopheryl acetate | mg/kg | – | 200 | 200 |
| Proximate composition | ||||
| Crude protein | g/kg | 174 | 174 | 174 |
| Ether extract | g/kg | 47.7 | 47.7 | 47.2 |
| Crude fibre | g/kg | 122 | 122 | 137 |
| Ash | g/kg | 89 | 89 | 84 |
| Fatty acids provided by flaxseed | ||||
| C18:3 ALA | g/kg | 9.23 | 9.05 | 12.08 |
References
- Murphy, M.P.; Holmgren, A.; Larsson, N.G.; Halliwell, B.; Chang, C.J.; Kalyanaraman, B.; Rhee, S.G.; Thornalley, P.J.; Partridge, L.; Gems, D.; et al. Unraveling the biological roles of reactive oxygen species. Cell Metab. 2011, 13, 361–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauridsen, C. From oxidative stress to inflammation: Redox balance and immune system. Poult. Sci. 2019, 98, 4240–4246. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar] [CrossRef]
- Reeg, S.; Grune, T. Protein Oxidation in Aging: Does It Play a Role in Aging Progression? Antioxid. Redox Signal. 2015, 23, 239–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanvah, S.; Joseph, J.; Schuster, G.B.; Barnett, R.N.; Cleveland, C.L.; Landman, U.Z.I. Oxidation of DNA: Damage to nucleobases. Acc. Chem. Res. 2010, 43, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Milne, G.L.; Dai, Q.; Roberts, L.J. The isoprostanes-25 years later. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2015, 1851, 433–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.S.; Ahsan, H.; Zia, M.K.; Siddiqui, T.; Khan, F.H. Understanding oxidants and antioxidants: Classical team with new players. J. Food Biochem. 2020, 44, e13145. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of Oxidative Stress on Male Reproduction. World J. Mens. Health 2014, 32, 1. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B. Antioxidant defence mechanisms: From the beginning to the end (of the beginning). Free Radic. Res. 1999, 31, 261–272. [Google Scholar] [CrossRef]
- Sies, H. Glutathione and its role in cellular functions. Free Radic. Biol. Med. 1999, 27, 916–921. [Google Scholar] [CrossRef]
- Mattioli, S.; Collodel, G.; Signorini, C.; Cotozzolo, E.; Noto, D.; Cerretani, D.; Micheli, L.; Fiaschi, A.I.; Brecchia, G.; Menchetti, L.; et al. Tissue antioxidant status and lipid peroxidation are related to dietary intake of n-3 polyunsaturated acids: A rabbit model. Antioxidants 2021, 10, 681. [Google Scholar] [CrossRef] [PubMed]
- Griveau, J.F.; Le Lannou, D. Reactive oxygen species and human spermatozoa: Physiology and pathology. Int. J. Androl. 1997, 20, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Castellini, C.; Mattioli, S.; Signorini, C.; Cotozzolo, E.; Noto, D.; Moretti, E.; Brecchia, G.; Dal Bosco, A.; Belmonte, G.; Durand, T.; et al. Effect of Dietary n-3 Source on Rabbit Male Reproduction. Oxid. Med. Cell. Longev. 2019, 2019, 3279670. [Google Scholar] [CrossRef] [Green Version]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Angel Torres, M.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef]
- Sauer, H.; Rahimi, G.; Hescheler, J.; Wartenberg, M. Role of reactive oxygen species and phosphatidylinositol 3-kinase in cardiomyocyte differentiation of embryonic stem cells. FEBS Lett. 2000, 476, 218–223. [Google Scholar] [CrossRef]
- Niethammer, P.; Grabher, C.; Look, A.T.; Mitchison, T.J. A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in zebrafish. Nature 2009, 459, 996–999. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Visweswariah, S.S. Intramacrophage ROS Primes the Innate Immune System via JAK/STAT and Toll Activation. Cell Rep. 2020, 33, 108368. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Torquato, P.; Bartolini, D.; Giusepponi, D.; Piroddi, M.; Sebastiani, B.; Saluti, G.; Galarini, R.; Galli, F. Increased plasma levels of the lipoperoxyl radical-derived vitamin E metabolite α-tocopheryl quinone are an early indicator of lipotoxicity in fatty liver subjects. Free Radic. Biol. Med. 2019, 131, 115–125. [Google Scholar] [CrossRef]
- Suburu, J.; Gu, Z.; Chen, H.; Chen, W.; Zhang, H.; Chen, Y.Q. Fatty acid metabolism: Implications for diet, genetic variation, and disease. Food Biosci. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simopoulos, A.P. Human requirement for N-3 polyunsaturated fatty acids. Poult. Sci. 2000, 79, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Genetic variation and dietary response: Nutrigenetics/nutrigenomics. Asia Pac. J. Clin. Nutr. 2002, 11, S117–S128. [Google Scholar] [CrossRef]
- Castellini, C.; Mourvaki, E.; Dal Bosco, A.; Galli, F. Vitamin E biochemistry and function: A case study in male rabbit. Reprod. Domest. Anim. 2007, 42, 248–256. [Google Scholar] [CrossRef]
- Mattioli, S.; Dal Bosco, A.; Duarte, J.M.M.; D’Amato, R.; Castellini, C.; Beone, G.M.; Fontanella, M.C.; Beghelli, D.; Regni, L.; Businelli, D.; et al. Use of Selenium-enriched olive leaves in the feed of growing rabbits: Effect on oxidative status, mineral profile and Selenium speciation of Longissimus dorsi meat. J. Trace Elem. Med. Biol. 2019. [Google Scholar] [CrossRef]
- Signorini, C.; Cardile, V.; Pannuzzo, G.; Graziano, A.C.E.; Durand, T.; Galano, J.M.; Oger, C.; Leoncini, S.; Cortelazzo, A.; Lee, J.C.Y.; et al. Increased isoprostanoid levels in brain from murine model of Krabbe disease–Relevance of isoprostanes, dihomo-isoprostanes and neuroprostanes to disease severity. Free Radic. Biol. Med. 2019, 139, 46–54. [Google Scholar] [CrossRef]
- Ahmed, O.; Galano, J.-M.; Pavlickova, T.; Revol-Cavalier, J.; Vigor, C.; Lee, J.-Y.; Oger, C.; Durand, T. Moving forward with isoprostanes, neuroprostanes and phytoprostanes: Where are we now? Essays Biochem. 2020, 64, 463–484. [Google Scholar] [CrossRef]
- Signorini, C.; Moretti, E.; Noto, D.; Mattioli, S.; Castellini, C.; Pascarelli, N.A.; Durand, T.; Oger, C.; Galano, J.M.; De Felice, C.; et al. F4-neuroprostanes: A role in sperm capacitation. Life 2021, 11, 655. [Google Scholar] [CrossRef]
- Galli, F.; Azzi, A.; Birringer, M.; Cook-Mills, J.M.; Eggersdorfer, M.; Frank, J.; Cruciani, G.; Lorkowski, S.; Özer, N.K. Vitamin E: Emerging aspects and new directions. Free Radic. Biol. Med. 2017, 102, 16–36. [Google Scholar] [CrossRef]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Torquato, P.; Giusepponi, D.; Galarini, R.; Bartolini, D.; Piroddi, M.; Galli, F. Analysis of vitamin E metabolites. In Vitamin E: Chemistry and Nutritional Benefits; Royal Society of Chemistry: London, UK, 2019; pp. 208–227. [Google Scholar]
- Meunier, B.; de Visser, S.P.; Shaik, S. Mechanism of oxidation reactions catalyzed by cytochrome P450 enzymes. Chem. Rev. 2004, 104, 3947–3980. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Ursini, F. Homeostatic control of redox status and health. IUBMB Life 2022, 74, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.L. Protein Oxidation and Implications for Muscle Food Quality. In Antioxidants in Muscle Foods: Nutritional Strategies to Improve Quality; John Wiley & Sons: New York, NY, USA, 2000; ISBN 0-471-31454-4. [Google Scholar]
- Xiong, J. Fatty Acid Oxidation in Cell Fate Determination. Trends Biochem. Sci. 2018, 43, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Suckow, M.A.; Stevens, K.A.; Wilson, R.P. (Eds.) The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Sargent, R.G. Verification and validation of simulation models. Proc.-Winter Simul. Conf. 2010, 166–183. [Google Scholar] [CrossRef] [Green Version]
- Trucano, T.G.; Swiler, L.P.; Igusa, T.; Oberkampf, W.L.; Pilch, M. Calibration, validation, and sensitivity analysis: What’s what. Reliab. Eng. Syst. Saf. 2006, 91, 1331–1357. [Google Scholar] [CrossRef]
- Dimokas, G.; Tchamitchian, M.; Kittas, C. Calibration and validation of a biological model to simulate the development and production of tomatoes in Mediterranean greenhouses during winter period. Biosyst. Eng. 2009, 103, 217–227. [Google Scholar] [CrossRef]
- VENSIM, Vensim(R) PLE, Version 9.0.1, Ventana Systems, Inc. Available online: https://vensim.com/vensim-personal-learning-edition (accessed on 30 November 2021).
- Brieger, K.; Schiavone, S.; Miller, F.J.; Krause, K.H. Reactive oxygen species: From health to disease. Swiss Med. Wkly. 2012, 142, w13659. [Google Scholar] [CrossRef]
- Turchini, G.M.; Francis, D.S.; De Silva, S.S. A whole body, in vivo, fatty acid balance method to quantify PUFA metabolism (desaturation, elongation and beta-oxidation). Lipids 2007, 42, 1065–1071. [Google Scholar] [CrossRef]
- Bargut, T.C.L.; Frantz, E.D.C.; Mandarim-De-Lacerda, C.A.; Aguila, M.B. Effects of a diet rich in n-3 polyunsaturated fatty acids on hepatic lipogenesis and beta-oxidation in mice. Lipids 2014, 49, 431–444. [Google Scholar] [CrossRef]
- Mouchlis, V.D.; Hayashi, D.; Vasquez, A.M.; Cao, J.; McCammon, J.A.; Dennis, E.A. Lipoprotein-associated phospholipase A2: A paradigm for allosteric regulation by membranes. Proc. Natl. Acad. Sci. USA 2022, 119, 1–8. [Google Scholar] [CrossRef]
- Min, B.; Ahn, D.U. Mechanism of lipid peroxidation in meat and meat products-A review. Food Sci. Biotechnol. 2005, 14, 152–163. [Google Scholar]
- Fereidoon, S.; Ying, Z. Lipid oxidation and improving the oxidative stability. Chem. Soc. Rev. 2010, 39, 4067–4079. [Google Scholar] [CrossRef]
- Galli, F.; Polidori, M.C.; Stahl, W.; Mecocci, P.; Kelly, F.J. Vitamin E biotransformation in humans. Vitam. Horm. 2007, 76, 263–280. [Google Scholar] [PubMed]
- Torquato, P.; Giusepponi, D.; Bartolini, D.; Barola, C.; Marinelli, R.; Sebastiani, B.; Galarini, R.; Galli, F. Pre-analytical monitoring and protection of oxidizable lipids in human plasma (vitamin E and ω−3 and ω−6 fatty acids): An update for redox-lipidomics methods. Free Radic. Biol. Med. 2021, 176, 142–148. [Google Scholar] [CrossRef]
- Meister, A. Glutathione-ascorbic acid antioxidant system in animals. J. Biol. Chem. 1994, 269, 9397–9400. [Google Scholar] [CrossRef]
- Combs, G.F. Biomarkers of selenium status. Nutrients 2015, 7, 2209–2236. [Google Scholar] [CrossRef] [Green Version]
- De Blas, C.; Wisewan, J. Nutrition of the Rabbit; Cabi: Wallingford, UK, 2020. [Google Scholar]
- Castellini, C.; Lattaioli, P.; Dal Bosco, A.; Beghelli, D. Effect of supranutritional level of dietary α-tocopheryl acetate and selenium on rabbit semen. Theriogenology 2002, 58, 1723–1732. [Google Scholar] [CrossRef]
- Rodríguez, M.; Rebollar, P.G.; Mattioli, S.; Castellini, C. n-3 PUFA sources (precursor/products): A review of current knowledge on rabbit. Animals 2019, 9, 806. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar] [CrossRef]
- Mattioli, S.; Machado Duarte, J.M.; Castellini, C.; D’Amato, R.; Regni, L.; Proietti, P.; Businelli, D.; Cotozzolo, E.; Rodrigues, M.; Dal Bosco, A. Use of olive leaves (whether or not fortified with sodium selenate) in rabbit feeding: Effect on performance, carcass and meat characteristics, and estimated indexes of fatty acid metabolism. Meat Sci. 2018, 143, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.A. Determination of ascorbic acid and uric acid in plasma by high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1994, 657, 197–200. [Google Scholar] [CrossRef]
- Shara, M.A.; Dickson, P.H.; Bagchi, D.; Stohs, S.J. Excretion of formaldehyde, malondialdehyde, acetaldehyde and acetone in the urine of rats in response to 2,3,7,8-tetrachlorodibenzo-p-dioxin, paraquat, endrin and carbon tetrachloride. J. Chromatogr. B Biomed. Sci. Appl. 1992, 576, 221–233. [Google Scholar] [CrossRef]
- Longini, M.; Moretti, E.; Signorini, C.; Noto, D.; Iacoponi, F.; Collodel, G. Relevance of seminal F2-dihomo-IsoPs, F2-IsoPs and F4-NeuroPs in idiopathic infertility and varicocele. Prostaglandins Other Lipid Mediat. 2020, 149, 106448. [Google Scholar] [CrossRef]
- SAS. Proc GLM in SAS, Release 9.4; SAS Institute Inc.: Cary, NC, USA, 2015. [Google Scholar]
- Feature, T. A Living System By Monya Baker. Nature 2011, 471, 661–665. [Google Scholar]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, D.; Torquato, P.; Barola, C.; Russo, A.; Rychlicki, C.; Giusepponi, D.; Bellezza, G.; Sidoni, A.; Galarini, R.; Svegliati-Baroni, G.; et al. Nonalcoholic fatty liver disease impairs the cytochrome P-450-dependent metabolism of α-tocopherol (vitamin E). J. Nutr. Biochem. 2017, 47, 120–131. [Google Scholar] [CrossRef]
- Chakraborty, P.; Dugmonits, K.N.; Végh, A.G.; Hollandi, R.; Horváth, P.; Maléth, J.; Hegyi, P.; Németh, G.; Hermesz, E. Failure in the compensatory mechanism in red blood cells due to sustained smoking during pregnancy. Chem. Biol. Interact. 2019, 313, 108821. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Yin, H. Role of lipid peroxidation derived 4-hydroxynonenal (4-HNE) in cancer: Focusing on mitochondria. Redox Biol. 2015, 4, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, D.; Marinelli, R.; Giusepponi, D.; Galarini, R.; Barola, C.; Stabile, A.M.; Sebastiani, B.; Paoletti, F.; Betti, M.; Rende, M.; et al. Alpha-tocopherol metabolites (the vitamin E metabolome) and their interindividual variability during supplementation. Antioxidants 2021, 10, 173. [Google Scholar] [CrossRef]
- Musesti, A.; Giusteri, G.G.; Marzocchi, A. Predicting ageing: On the mathematical modelization of ageing muscle tissue. In 821 Active Ageing and Healthy Living; IOS Press: Amsterdam, The Netherlands, 2014; pp. 185–192. [Google Scholar]
- Giantesio, G.; Marzocchi, A.; Musesti, A. Loss of mass and performance in skeletal muscle tissue: A continuum model. Commun. Appl. Ind. Math. 2018, 9, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Mattioli, S.; Dal Bosco, A.; Maranesi, M.; Petrucci, L.; Rebollar, P.G.; Castellini, C. Dietary fish oil and flaxseed for rabbit does: Fatty acids distribution and Δ6-desaturase enzyme expression of different tissues. Animal 2019, 13, 1934–1942. [Google Scholar] [CrossRef] [PubMed]




| VENSIM INPUT 1 | OUTPUT 2 | Mean ± SE (Analytic Determination) | Confidence Interval (95%) | VENSIM Values | |
|---|---|---|---|---|---|
| Lower Limit | Upper Limit | ||||
| standard model | |||||
| 7000 | ROS | 5704.29 ± 101.53 a | 5474.59 | 5933.99 | 5600.00 |
| 5.10 | PUFA | 5.27 ± 0.08 a | 5.01 | 5.54 | 5.48 |
| 0.0024 | Vit E | 422.00 ± 6.95 × 10−5 a | 400.61 × 10−5 | 435.60 × 10−5 | 401.00 × 10−5 |
| 0.0026 | AA | 929.74 ± 34.66 × 10−5 | 851.80 × 10−5 | 1008.67 × 10−5 | 920.00 × 10−5 |
| – | IsoNeuroPs | 0.1494 ± 0.026 a | 0.1434 | 0.1554 | 0.1400 |
| – | Aldehydes | 10.10 ± 0.16 a | 9.72 | 10.48 | 9.00 |
| – | Vit Eox | 5.99 ± 0.69 × 10−5 a | 4.63 × 10−5 | 7.33 × 10−5 | 4.98 × 10−5 |
| VIT E + | |||||
| 7000 | ROS | 6690.82 ± 164.04 b | 6319.71 | 7061.93 | 6720.00 |
| 5.10 | PUFA | 6.62 ± 0.50 a | 5.49 | 7.75 | 5.50 |
| 0.0030 | Vit E | 458.41 ± 22.2 × 10−5 a | 408.82 × 10−5 | 501.00 × 10−5 | 457.00 × 10−5 |
| 0.0026 | AA | 937.48 ± 47.60 × 10−5 | 829.93 × 10−5 | 1045.73 × 10−5 | 928.00 × 10−5 |
| – | IsoNeuroPs | 0.1412 ± 0.00329 a | 0.1338 | 0.1487 | 0.140 |
| – | Aldehydes | 7.45 ± 0.27 b | 6.84 | 8.07 | 7.02 |
| – | Vit Eox | 16.42 ± 1.44 × 10−5 b | 11.71 × 10−5 | 20.50 × 10−5 | 12.60 × 10−5 |
| PUFA + | |||||
| 7000 | ROS | 7871.43 ± 227.15 b | 7357.57 | 8385.30 | 7416.00 |
| 600 | PUFA | 82.15 ± 3.58 b | 74.17 | 90.12 | 76.99 |
| 0.0030 | Vit E | 260.22 ± 8.75 × 10−5 b | 243.06 × 10−5 | 277.38 × 10−5 | 298.00 × 10−5 |
| 0.0026 | AA | 976.78 ± 31.99 × 10−5 | 904.46 × 10−5 | 1049.50 × 10−5 | 953.30 × 10−5 |
| – | IsoNeuroPs | 0.2566 ± 0.0608 b | 0.2412 | 0.2720 | 0.3000 |
| – | Aldehydes | 14.05 ± 0.40 c | 13.13 | 14.97 | 14.81 |
| – | Vit Eox | 4.67 ± 0.77 × 10−5 a | 2.92 × 10−5 | 6.19 × 10−5 | 4.69 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattioli, S.; Dimauro, C.; Cesarani, A.; Dal Bosco, A.; Bartolini, D.; Galli, F.; Migni, A.; Sebastiani, B.; Signorini, C.; Oger, C.; et al. A Dynamic Model for Estimating the Interaction of ROS–PUFA–Antioxidants in Rabbit. Antioxidants 2022, 11, 531. https://doi.org/10.3390/antiox11030531
Mattioli S, Dimauro C, Cesarani A, Dal Bosco A, Bartolini D, Galli F, Migni A, Sebastiani B, Signorini C, Oger C, et al. A Dynamic Model for Estimating the Interaction of ROS–PUFA–Antioxidants in Rabbit. Antioxidants. 2022; 11(3):531. https://doi.org/10.3390/antiox11030531
Chicago/Turabian StyleMattioli, Simona, Corrado Dimauro, Alberto Cesarani, Alessandro Dal Bosco, Desiree Bartolini, Francesco Galli, Anna Migni, Bartolomeo Sebastiani, Cinzia Signorini, Camille Oger, and et al. 2022. "A Dynamic Model for Estimating the Interaction of ROS–PUFA–Antioxidants in Rabbit" Antioxidants 11, no. 3: 531. https://doi.org/10.3390/antiox11030531
APA StyleMattioli, S., Dimauro, C., Cesarani, A., Dal Bosco, A., Bartolini, D., Galli, F., Migni, A., Sebastiani, B., Signorini, C., Oger, C., Collodel, G., & Castellini, C. (2022). A Dynamic Model for Estimating the Interaction of ROS–PUFA–Antioxidants in Rabbit. Antioxidants, 11(3), 531. https://doi.org/10.3390/antiox11030531