
Phoenix dactilyfera L. Pits Extract Restored Bone Homeostasis in Glucocorticoid-Induced Osteoporotic Animal Model through the Antioxidant Effect and Wnt5a Non-Canonical Signaling

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of Date Pits Extract
2.3. Characterization of Date Pits Extract
2.4. Experimental Design
2.5. Collection and Preparation of Blood and Tissues
2.6. Bone Density Test
2.7. Biochemical Analysis and Spectrophotometry Evaluations
2.8. Sandwich ELISA
2.9. Total RNA Isolation and Quantitative Real-Time Reverse Transcription PCR Analysis (qRT-PCR)
2.10. Histopathological Study
2.11. Statistical Analysis
3. Results
3.1. Characterization of Date Pits Extract
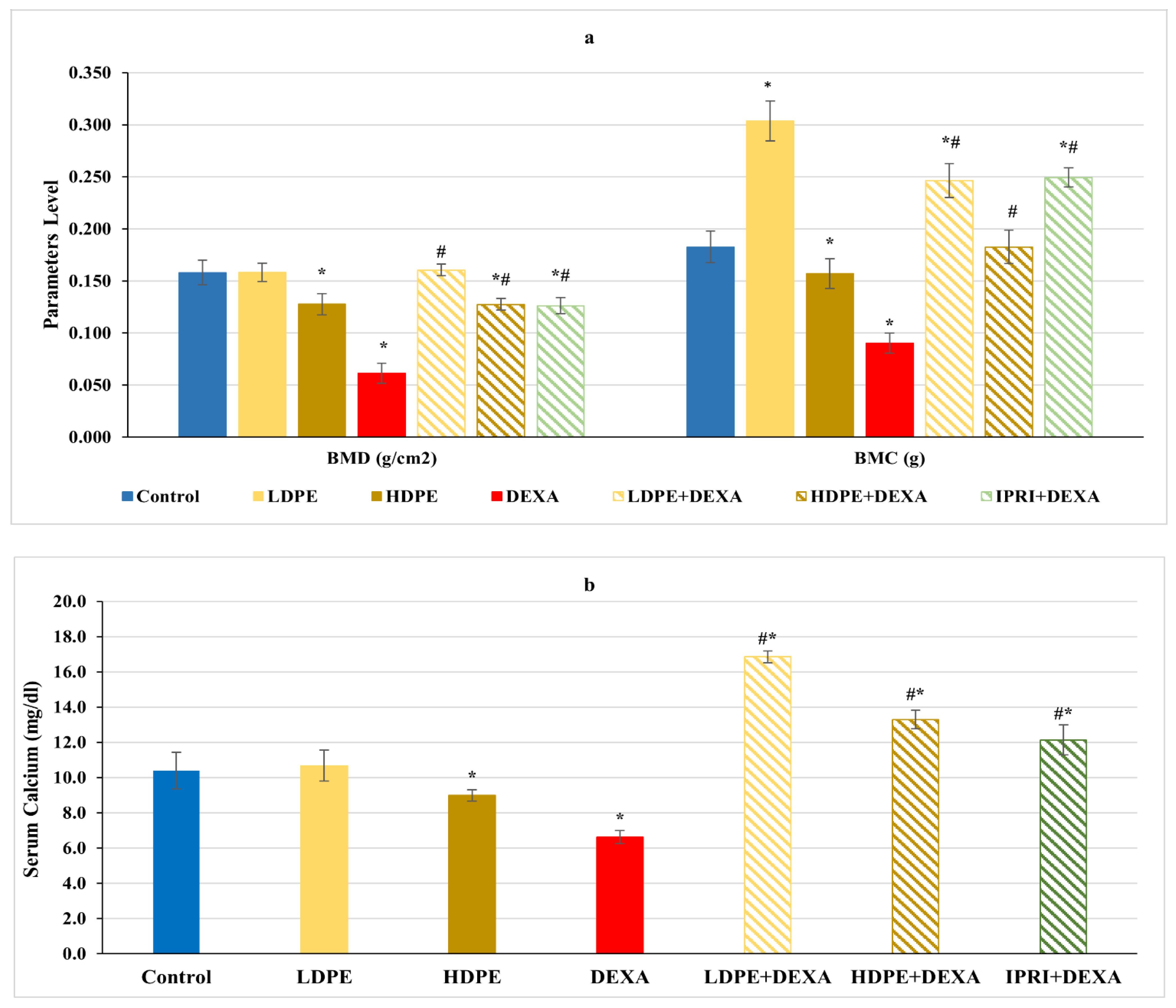
3.2. BMD, BMC, and Serum Calcium Level
3.3. Oxidative Stress Indices
3.4. Estimation of MSC Proliferation Markers
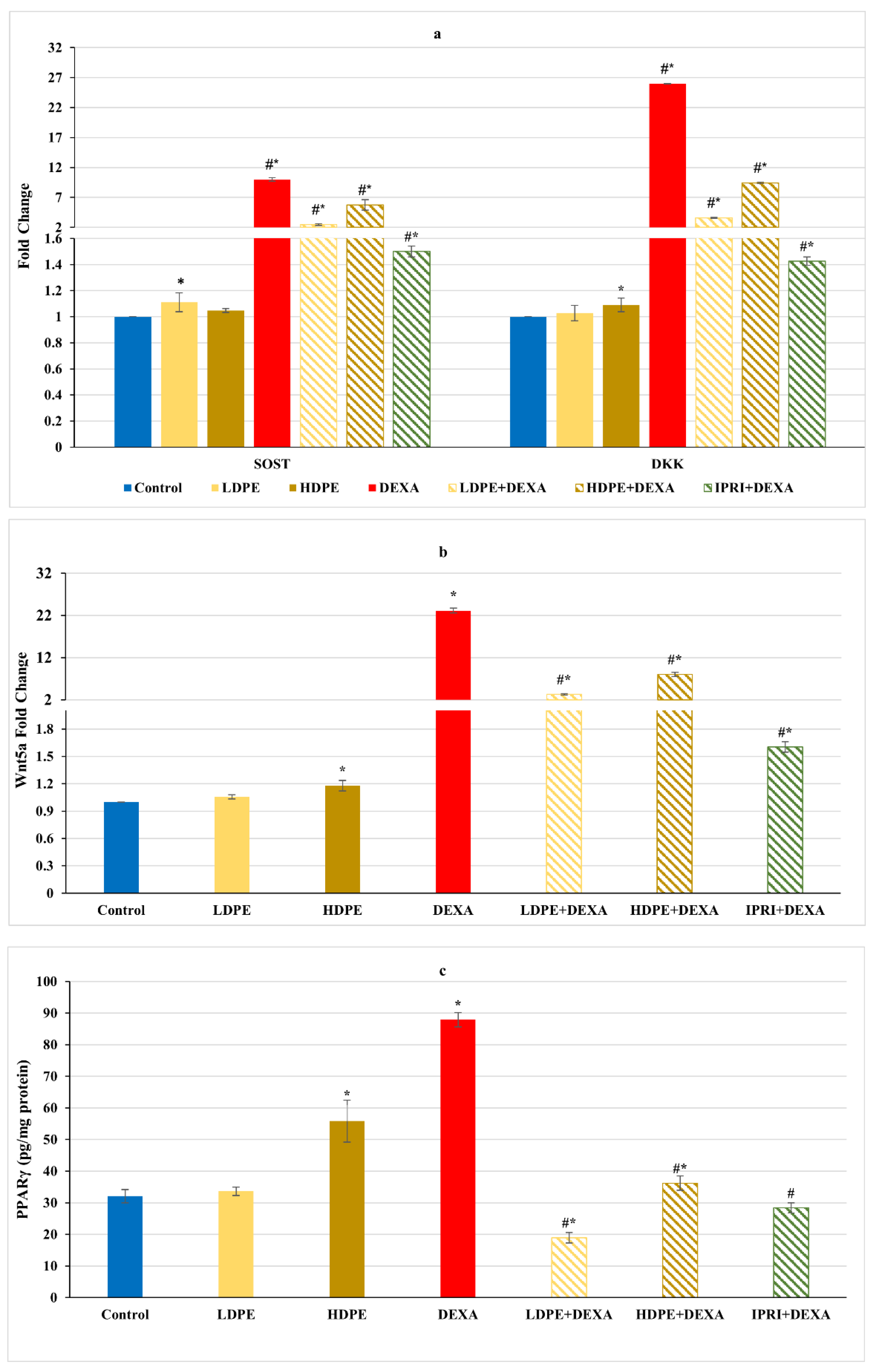
3.5. Osteoblast’s Differentiation Indices
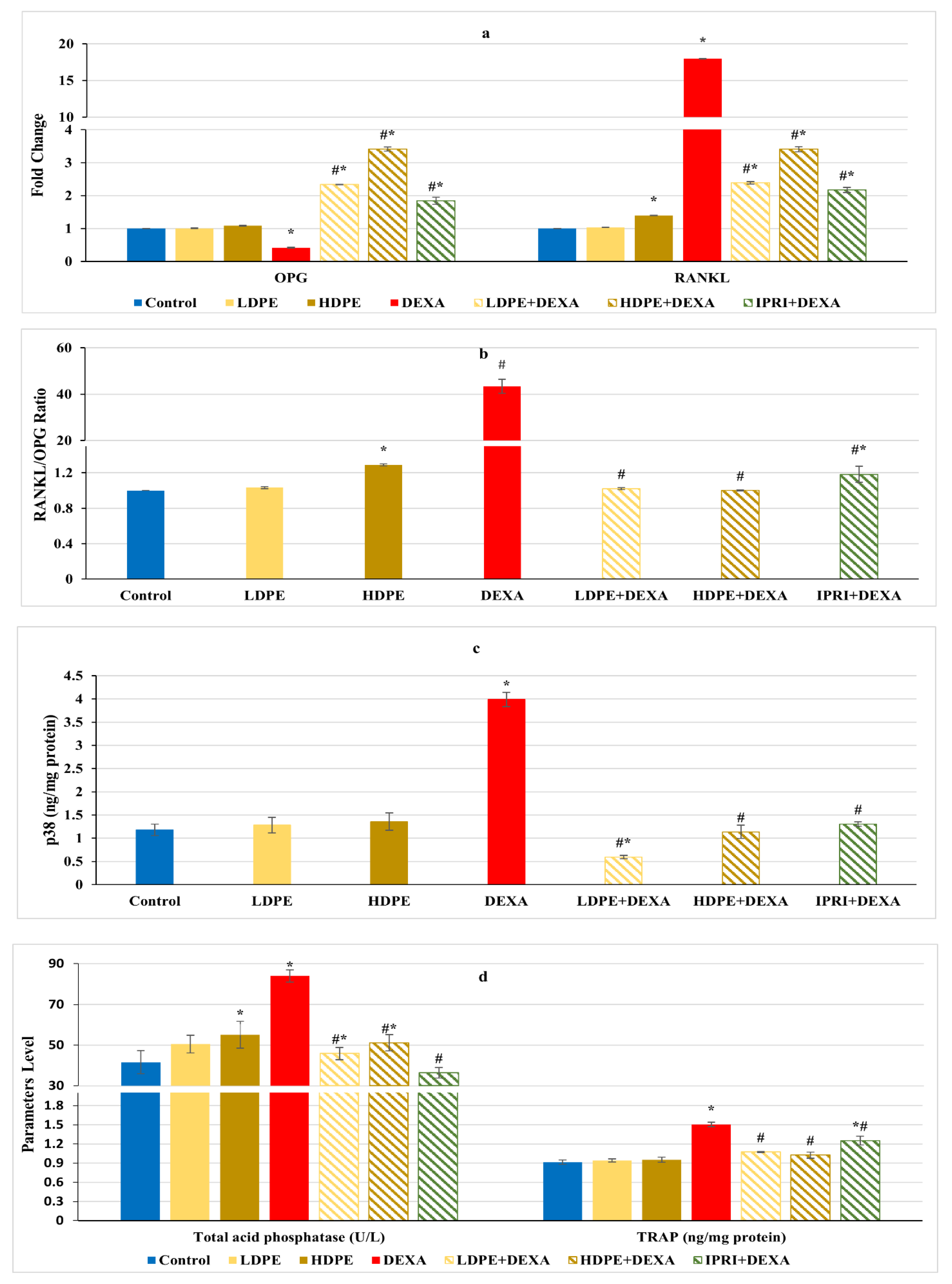
3.6. Osteoclast’s Differentiation Markers
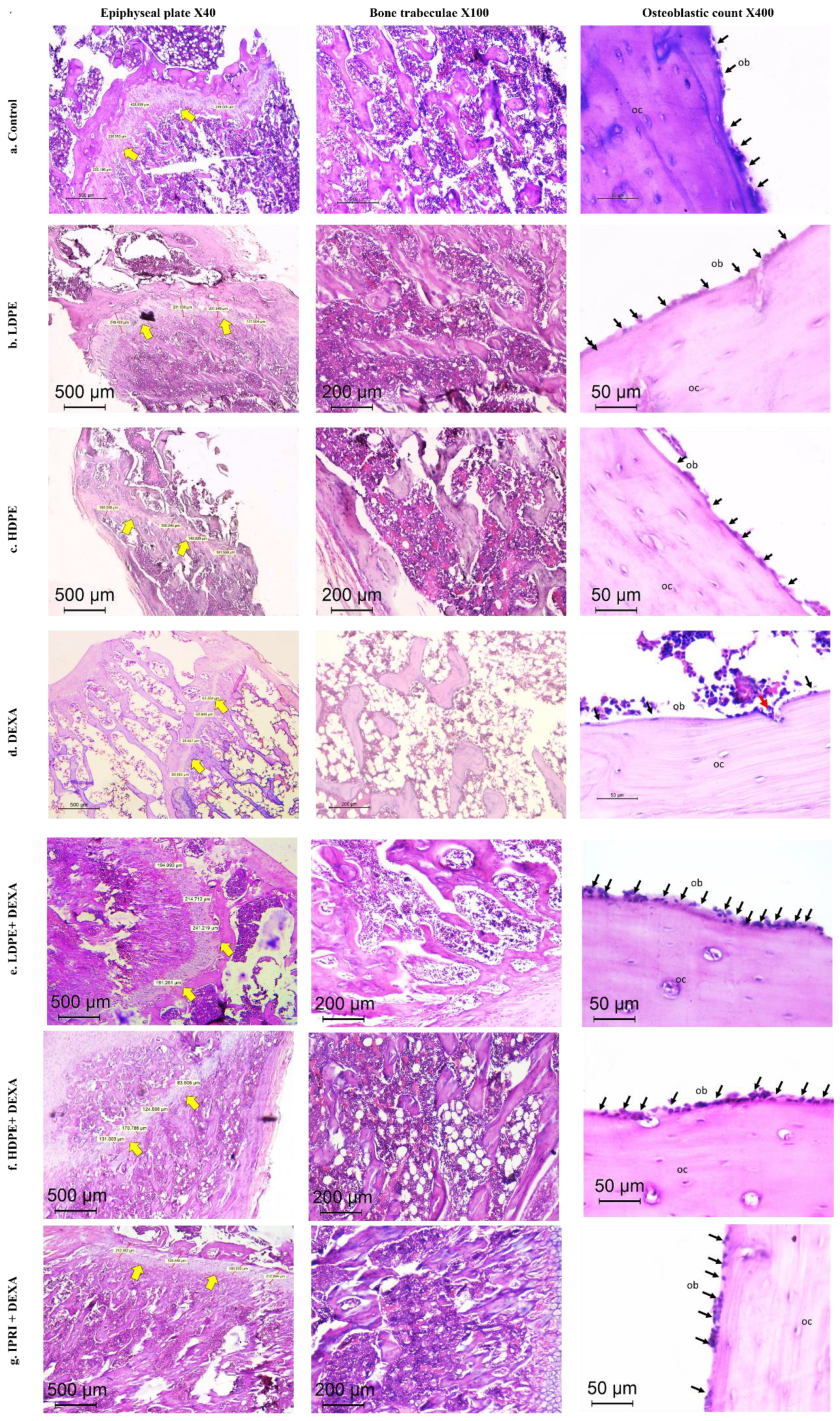
3.7. Histopathologic Outcomes
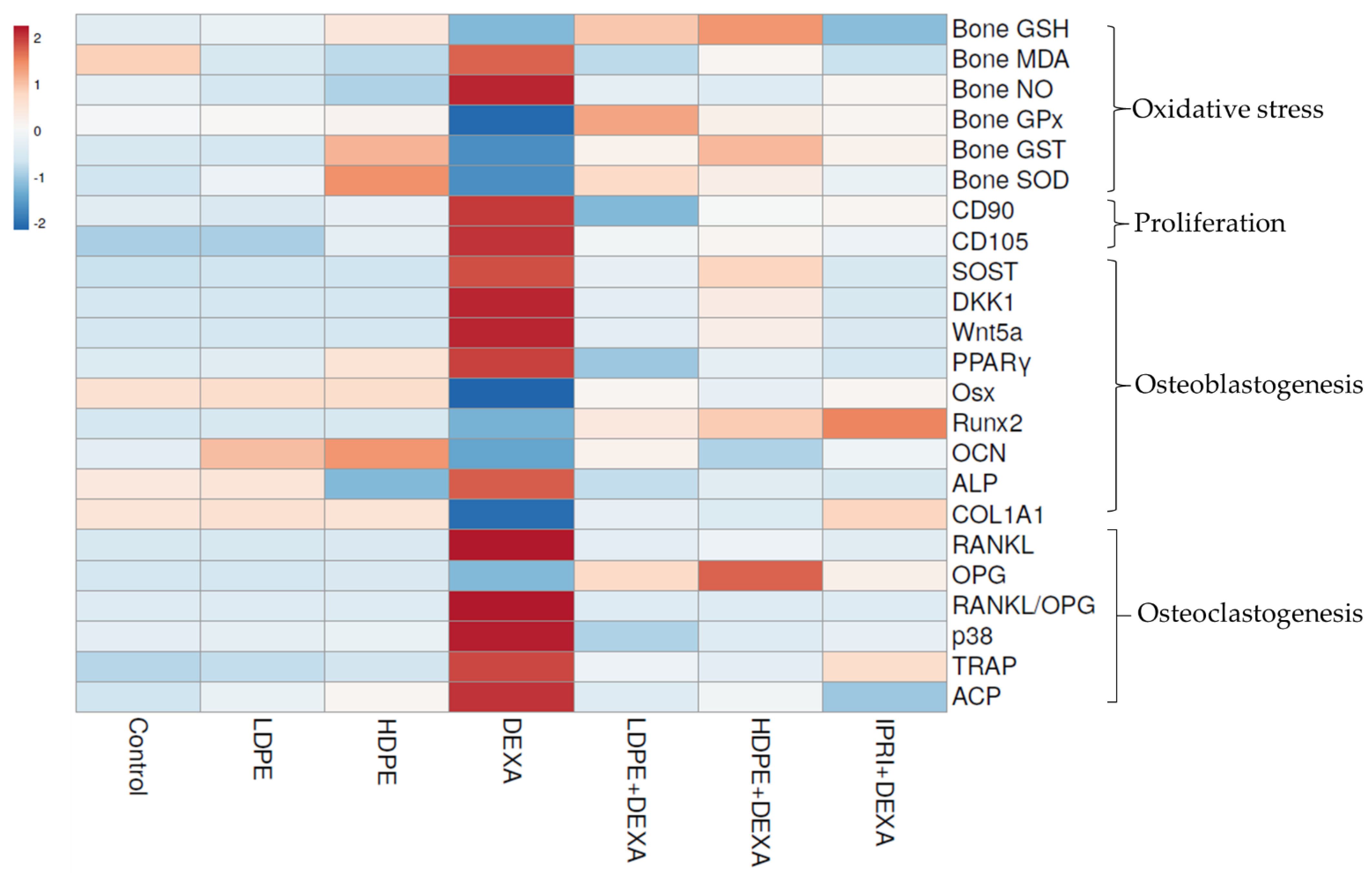
3.8. Heat Map Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrabet, A.; Jiménez-Araujo, A.; Guillén-Bejarano, R.; Rodríguez-Arcos, R.; Sindic, M. Date Seeds: A Promising Source of Oil with Functional Properties. Foods 2020, 9, 787. [Google Scholar] [CrossRef] [PubMed]
- Bentrad, N.; Gaceb-Terrak, R. Evaluation of the level of biomolecules isolated from date palm seeds (Phoenix dactylifera L.) and in vitro Antioxidant property. BioMedicine 2020, 10, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Khattak, M.N.K.; Shanableh, A.; Hussain, M.I.; Khan, A.A.; Abdulwahab, M.; Radeef, W.; Samreen, M.H. Anticancer activities of selected Emirati Date (Phoenix dactylifera L.) varieties pits in human triple negative breast cancer MDA-MB-231 cells. Saudi J. Biol. Sci. 2020, 27, 3390–3396. [Google Scholar] [CrossRef] [PubMed]
- Saleh, S.R.; Masry, A.M.; Ghareeb, D.A.; Newairy, A.-S.A.; Sheta, E.; Maher, A.M. Trichoderma reesei fungal degradation boosted the potentiality of date pit extract in fighting scopolamine-induced neurotoxicity in male rats. Sci. Rep. 2021, 11, 14872. [Google Scholar] [CrossRef]
- Gensler, L.S. Glucocorticoids: Complications to anticipate and prevent. Neurohospitalist 2013, 3, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef]
- Han, L.; Wang, B.; Wang, R.; Gong, S.; Chen, G.; Xu, W. The shift in the balance between osteoblastogenesis and adipogenesis of mesenchymal stem cells mediated by glucocorticoid receptor. Stem Cell Res. Ther. 2019, 10, 377. [Google Scholar] [CrossRef] [Green Version]
- Ardura, J.A.; Álvarez-Carrión, L.; Gortázar, A.R.; Alonso, V. Linking bone cells, aging, and oxidative stress: Osteoblasts, osteoclasts, osteocytes, and bone marrow cells. In Aging; Academic Press: Cambridge, MA, USA, 2020; pp. 61–71. [Google Scholar]
- Takada, I.; Yogiashi, Y.; Kato, S. Signaling Crosstalk between PPARγ and BMP2 in Mesenchymal Stem Cells. PPAR Res. 2012, 2012, 607141. [Google Scholar] [CrossRef] [Green Version]
- Gennari, C. Differential Effect of Glucocorticoids on Calcium Absorption and Bone Mass. Rheumatology 1993, 32, 11–14. [Google Scholar] [CrossRef]
- Ackers, I.; Malgor, R. Interrelationship of canonical and non-canonical Wnt signalling pathways in chronic metabolic diseases. Diabetes Vasc. Dis. Res. 2017, 15, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meszaros, K.; Patocs, A. Glucocorticoids Influencing Wnt/β-Catenin Pathway; Multiple Sites, Heterogeneous Effects. Molecules 2020, 25, 1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, U.H.; Ohlsson, C. The WNT system: Background and its role in bone. J. Intern. Med. 2015, 277, 630–649. [Google Scholar] [CrossRef] [PubMed]
- Houschyar, K.S.; Tapking, C.; Borrelli, M.R.; Popp, D.; Duscher, D.; Maan, Z.N.; Chelliah, M.P.; Li, J.; Harati, K.; Wallner, C.; et al. Wnt Pathway in Bone Repair and Regeneration—What Do We Know So Far. Front. Cell Dev. Biol. 2019, 6, 170. [Google Scholar] [CrossRef]
- Kim, J.H.; Liu, X.; Wang, J.; Chen, X.; Zhang, H.; Kim, S.H.; Cui, J.; Li, R.; Zhang, W.; Kong, Y.; et al. Wnt signaling in bone formation and its therapeutic potential for bone diseases. Ther. Adv. Musculoskelet. Dis. 2013, 5, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, J.; Chen, J.; Dong, C.; Yan, X.; Zhu, Z.; Lu, P.; Song, Z.; Liu, H.; Chen, S. Daphnetin ameliorates glucocorticoid-induced osteoporosis via activation of Wnt/GSK-3β/β-catenin signaling. Toxicol. Appl. Pharmacol. 2020, 409, 115333. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Uehara, S.; Udagawa, N. Roles of non-canonical Wnt signaling pathways in bone resorption. J. Oral Biosci. 2018, 60, 31–35. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Uehara, S.; Koide, M.; Takahashi, N. The regulation of osteoclast differentiation by Wnt signals. BoneKEy Rep. 2015, 4, 713. [Google Scholar] [CrossRef] [Green Version]
- De Winter, T.J.J.; Nusse, R. Running Against the Wnt: How Wnt/β-Catenin Suppresses Adipogenesis. Front. Cell Dev. Biol. 2021, 9, 140. [Google Scholar] [CrossRef]
- Gao, A.-G.; Zhou, Y.-C.; Hu, Z.-J.; Lu, B.-B. Ipriflavone promotes osteogenesis of MSCs derived from osteoporotic rats. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4669–4676. [Google Scholar]
- John, A.; Konkodi, K. Revisiting Ipriflavone: A Potential Isoflavone for the Management of Postmenopausal Osteoporosis. Rev. Bras. Farm. 2021, 10, 1–8. [Google Scholar] [CrossRef]
- Geng, M.; Ren, M.; Liu, Z.; Shang, X. Free radical scavenging activities of pigment extract from Hibiscus syriacus L. petals in vitro. Afr. J. Biotechnol. 2012, 11, 429–435. [Google Scholar]
- Taga, M.S.; Miller, E.E.; Pratt, D.E. Chia seeds as a source of natural lipid antioxidants. J. Am. Oil Chem. Soc. 1984, 61, 928–931. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- U.S. EPA. Method 200.7 trace elements in water, solids, and biosolids, by inductively coupled plasma-atomic emission spectrometry. In Methods for Chemical Analysis of Water and Wastes; U.S. EPA: Washington, DC, USA, 1983; Volume 5, p. 821-01-010. [Google Scholar]
- U.S. EPA. Method 6010C (SW-846): Inductively Coupled Plasma—Atomic Emission Spectrometry. 2000. Available online: https://www.epa.gov/sites/production/files/2015-07/documents/epa-6010c.pdf (accessed on 18 May 2021).
- Takahashi, M.; Ushijima, K.; Hayashi, Y.; Maekawa, T.; Ando, H.; Tsuruoka, S.-I.; Fujimura, A. Dosing-time dependent effect of dexamethasone on bone density in rats. Life Sci. 2010, 86, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Tappel, A.; Zalkin, H. Inhibition of lipide peroxidation in mitochondria by vitamin E. Arch. Biochem. Biophys. 1959, 80, 333–336. [Google Scholar] [CrossRef]
- Montgomery, H.; Dymock, J. The determination of nitrite in water. Analyst 1961, 86, 414–416. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Paglia, E.; Valentine, N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Chiu, D.T.; Stults, F.H.; Tappel, A.L. Purification and properties of rat lung soluble glutathione peroxidase. Biochim. Biophys. Acta (BBA)—Enzymol. 1976, 445, 558–566. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the Superoxide Anion Radical in the Autoxidation of Pyrogallol and a Convenient Assay for Superoxide Dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Maher, A.M.; Saleh, S.R.; Elguindy, N.; Hashem, H.M.; Yacout, G.A. Exogenous melatonin restrains neuroinflammation in high fat diet induced diabetic rats through attenuating indoleamine 2,3-dioxygenase 1 expression. Life Sci. 2020, 247, 117427. [Google Scholar] [CrossRef] [PubMed]
- An, J.-J.; Han, D.-H.; Kim, D.-M.; Kim, S.-H.; Rhee, Y.; Lee, E.-J.; Lim, S.-K. Expression and Regulation of Osteoprotegerin in Adipose Tissue. Yonsei Med. J. 2007, 48, 765–772. [Google Scholar] [CrossRef] [Green Version]
- Caricasole, A.; Copani, A.; Caraci, F.; Aronica, E.; Rozemuller, A.J.; Caruso, A.; Storto, M.; Gaviraghi, G.; Terstappen, G.C.; Nicoletti, F. Induction of Dickkopf-1, a negative modulator of the Wnt pathway, is associated with neuronal degeneration in Alzheimer’s brain. J. Neurosci. 2004, 24, 6021–6027. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Galfo, F.; Oteri, G.; Atteritano, M.; Pallio, G.; Mannino, F.; D’Amore, A.; Pellegrino, E.; Aliquò, F.; et al. Adenosine Receptor Stimulation Improves Glucocorticoid-Induced Osteoporosis in a Rat Model. Front. Pharmacol. 2017, 8, 558. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Xue, J.; Shen, T.; Mu, S.; Fu, Q. Curcumin alleviates glucocorticoid-induced osteoporosis through the regulation of the Wnt signaling pathway. Int. J. Mol. Med. 2016, 37, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Xu, X.; Zhang, B.; Chen, B.; Hang, W. Expression of ATF4 and RUNX2 in periodontal tissue of pressure side during orthodontic tooth movement in rat. Int. J. Clin. Exp. Med. 2015, 8, 934–938. [Google Scholar]
- Liao, X.; Feng, B.; Zhang, D.; Liu, P.; Zhou, X.; Li, R.; Ye, L. The Sirt6 gene: Does it play a role in tooth development? PLoS ONE 2017, 12, e0174255. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques. Am. J. Dermatopathol. 2009, 31, 514. [Google Scholar] [CrossRef]
- Bitto, A.; Polito, F.; Burnett, B.; Levy, R.; Di Stefano, V.; Armbruster, M.A.; Marini, H.R.; Minutoli, L.; Altavilla, D.; Squadrito, F. Protective effect of genistein aglycone on the development of osteonecrosis of the femoral head and secondary osteoporosis induced by methylprednisolone in rats. J. Endocrinol. 2009, 201, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, D.-K.; Disha, C.; Razdan, R.; Mahapatra, D.-R.; Bhat, M.-R. Comparative evaluation of zoledronic acid, alfacalcidol, and propranolol in pharmacological correction of experimental osteoporosis. Lat. Am. J. Pharm. 2013, 32, 968–976. [Google Scholar]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Hachemi, Y.; Rapp, A.E.; Picke, A.-K.; Weidinger, G.; Ignatius, A.; Tuckermann, J. Molecular mechanisms of glucocorticoids on skeleton and bone regeneration after fracture. J. Mol. Endocrinol. 2018, 61, R75–R90. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, S.R.; Soliman, A.M.; Sayed, A.A.; Marzouk, M. Possible antiosteoporotic mechanism of Cicer arietinum extract in ovariectomized rats. Int. J. Clin. Exp. Pathol. 2015, 8, 3477–3490. [Google Scholar] [PubMed]
- Samir, S.M.; Malek, H.A. Effect of cannabinoid receptors 1 modulation on osteoporosis in a rat model of different ages. J. Physiol. Pharmacol. 2014, 65, 687–694. [Google Scholar] [PubMed]
- Ahmed, H.; Morcos, N.Y.S.; Eskander, E.; Seoudi, D.; Shalby, A. Potential role of leptin against glucocorticoid-induced secondary osteoporosis in adult female rats. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 1446–1452. [Google Scholar]
- Hozayen, W.G.; El-Desouky, M.A.; Soliman, H.A.; Ahmed, R.R.; Ali, M. Antiosteoporotic effect of Petroselinum crispum, Ocimum basilicum and Cichorium intybus L. in glucocorticoid-induced osteoporosis in rats. BMC Complement. Altern. Med. 2016, 16, 165. [Google Scholar] [CrossRef] [Green Version]
- Armour, K.J.; Armour, K.E.; Van’t Hof, R.J.; Reid, D.M.; Wei, X.Q.; Liew, F.Y.; Ralston, S.H. Activation of the inducible nitric oxide synthase pathway contributes to inflammation-induced osteoporosis by suppressing bone formation and causing osteoblast apoptosis. Arthritis Rheum. 2001, 44, 2790–2796. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Nitric oxide and bone. Ann. N. Y. Acad. Sci. 2010, 1192, 391–403. [Google Scholar] [CrossRef]
- Whirledge, S.; Cidlowski, J.A. Glucocorticoids, stress, and fertility. Minerva Endocrinol. 2010, 35, 109–125. [Google Scholar] [PubMed]
- Manolagas, S.C. Steroids and osteoporosis: The quest for mechanisms. J. Clin. Investig. 2013, 123, 1919–19121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, T.J. Battle of the sex steroids in the male skeleton: And the winner is. J. Clin. Investig. 2016, 126, 829–832. [Google Scholar] [CrossRef] [Green Version]
- Atashi, F.; Modarressi, A.; Pepper, M.S. The Role of Reactive Oxygen Species in Mesenchymal Stem Cell Adipogenic and Osteogenic Differentiation: A Review. Stem Cells Dev. 2015, 24, 1150–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Marsboom, G.; Toth, P.; Rehman, J. Mitochondrial Respiration Regulates Adipogenic Differentiation of Human Mesenchymal Stem Cells. PLoS ONE 2013, 8, e77077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobeiha, M.; Moghadasian, M.H.; Amin, N.; Jafarnejad, S. RANKL/RANK/OPG Pathway: A Mechanism Involved in Exercise-Induced Bone Remodeling. BioMed Res. Int. 2020, 2020, 6910312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luis, T.C.; Naber, B.A.; Roozen, P.P.; Brugman, M.H.; de Haas, E.F.; Ghazvini, M.; Fibbe, W.E.; van Dongen, J.J.; Fodde, R.; Staal, F.J. Canonical Wnt Signaling Regulates Hematopoiesis in a Dosage-Dependent Fashion. Cell Stem Cell 2011, 9, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manolagas, S.C. Wnt signaling and osteoporosis. Maturitas 2014, 78, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.-X.; Cheng, X.-Y.; Wang, Y.; Yin, W. Toosendanin inhibits adipogenesis by activating Wnt/β-catenin signaling. Sci. Rep. 2018, 8, 4626. [Google Scholar] [CrossRef]
- Ahmadzadeh, A.; Norozi, F.; Shahrabi, S.; Shahjahani, M.; Saki, N. Wnt/β-catenin signaling in bone marrow niche. Cell Tissue Res. 2016, 363, 321–335. [Google Scholar] [CrossRef]
- Liu, Q.; Li, M.; Wang, S.; Xiao, Z.; Xiong, Y.; Wang, G. Recent Advances of Osterix Transcription Factor in Osteoblast Differentiation and Bone Formation. Front. Cell Dev. Biol. 2020, 8, 601224. [Google Scholar] [CrossRef] [PubMed]
- Reggio, A.; Rosina, M.; Palma, A.; Cerquone Perpetuini, A.; Petrilli, L.L.; Gargioli, C.; Fuoco, C.; Micarelli, E.; Giuliani, G.; Cerretani, M.; et al. Adipogenesis of skeletal muscle fibro/adipogenic progenitors is affected by the WNT5a/GSK3/β-catenin axis. Cell Death Differ. 2020, 27, 2921–2941. [Google Scholar] [CrossRef] [PubMed]
- Colla, S.; Zhan, F.; Xiong, W.; Wu, X.; Xu, H.; Stephens, O.; Yaccoby, S.; Epstein, J.; Barlogie, B.; Shaughnessy, J.D. The oxidative stress response regulates DKK1 expression through the JNK signaling cascade in multiple myeloma plasma cells. Blood 2007, 109, 4470–4477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinzone, J.J.; Hall, B.M.; Thudi, N.K.; Vonau, M.; Qiang, Y.-W.; Rosol, T.J.; Shaughnessy, J.D., Jr. The role of Dickkopf-1 in bone development, homeostasis, and disease. Blood 2009, 113, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Suen, P.K.; Qin, L. Sclerostin, an emerging therapeutic target for treating osteoporosis and osteoporotic fracture: A general review. J. Orthop. Transl. 2016, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Boonanantanasarn, K.; Baek, K.; Woo, K.M.; Ryoo, H.-M.; Baek, J.-H.; Kim, G.-S. Hyperglycemia increases the expression levels of sclerostin in a reactive oxygen species- and tumor necrosis factor-alpha-dependent manner. J. Periodontal Implant Sci. 2015, 45, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Maleki, M.; Ghanbarvand, F.; Behvarz, M.R.; Ejtemaei, M.; Ghadirkhomi, E. Comparison of Mesenchymal Stem Cell Markers in Multiple Human Adult Stem Cells. Int. J. Stem Cells 2014, 7, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Kays, S.-K.; Kaufmann, K.B.; Abel, T.; Brendel, C.; Bönig, H.; Grez, M.; Buchholz, C.J.; Kneissl, S. CD105 Is a Surface Marker for Receptor-Targeted Gene Transfer into Human Long-Term Repopulating Hematopoietic Stem Cells. Stem Cells Dev. 2015, 24, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Zeve, D.; Suh, J.M.; Wang, X.; Du, Y.; Zerwekh, J.E.; Dechow, P.C.; Graff, J.M.; Wan, Y. Biphasic and Dosage-Dependent Regulation of Osteoclastogenesis by β-Catenin. Mol. Cell. Biol. 2011, 31, 4706–4719. [Google Scholar] [CrossRef] [Green Version]
- Kamga, P.T.; Bazzoni, R.; Collo, G.D.; Cassaro, A.; Tanasi, I.; Russignan, A.; Tecchio, C.; Krampera, M. The Role of Notch and Wnt Signaling in MSC Communication in Normal and Leukemic Bone Marrow Niche. Front. Cell Dev. Biol. 2021, 8, 599276. [Google Scholar] [CrossRef]
- Bruderer, M.; Richards, R.G.; Alini, M.; Stoddart, M.J. Role and regulation of RUNX2 in osteogenesis. Eur. Cell Mater. 2014, 28, 269–286. [Google Scholar] [CrossRef] [PubMed]
- Phunikom, N.; Boonmuen, N.; Kheolamai, P.; Suksen, K.; Manochantr, S.; Tantrawatpan, C.; Tantikanlayaporn, D. Andrographolide promotes proliferative and osteogenic potentials of human placenta-derived mesenchymal stem cells through the activation of Wnt/β-catenin signaling. Stem Cell Res. Ther. 2021, 12, 241. [Google Scholar] [CrossRef] [PubMed]
- Weivoda, M.M.; Ruan, M.; Hachfeld, C.M.; Pederson, L.; Howe, A.; Davey, R.A.; Zajac, J.D.; Kobayashi, Y.; Williams, B.O.; Westendorf, J.J.; et al. Wnt Signaling Inhibits Osteoclast Differentiation by Activating Canonical and Noncanonical cAMP/PKA Pathways. J. Bone Miner. Res. 2015, 31, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolba, M.F.; El-Serafi, A.T.; Omar, H.A. Caffeic acid phenethyl ester protects against glucocorticoid-induced osteoporosis in vivo: Impact on oxidative stress and RANKL/OPG signals. Toxicol. Appl. Pharmacol. 2017, 324, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Choi, Y.G.; Baik, J.Y.; Han, S.Y.; Jeong, D.-W.; Bae, Y.S.; Kim, N.; Lee, S.Y. A crucial role for reactive oxygen species in RANKL-induced osteoclast differentiation. Blood 2005, 106, 852–859. [Google Scholar] [CrossRef] [Green Version]
- Briot, K.; Roux, C. Biochemical markers of bone remodeling. Gynecol. Obstet. Fertil. 2005, 33, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Bull, H.; Murray, P.G.; Thomas, D.; Fraser, A.M.; Nelson, P.N. Acid phosphatases. Mol. Pathol. 2002, 55, 65–72. [Google Scholar] [CrossRef]
- Yun, S.I.; Yoon, H.Y.; Jeong, S.Y.; Chung, Y.S. Glucocorticoid induces apoptosis of osteoblast cells through the activation of glycogen synthase kinase 3beta. J. Bone Miner. Metab. 2009, 27, 140–148. [Google Scholar] [CrossRef]
- Chotiyarnwong, P.; McCloskey, E.V. Pathogenesis of glucocorticoid-induced osteoporosis and options for treatment. Nat. Rev. Endocrinol. 2020, 16, 437–447. [Google Scholar] [CrossRef]
- Bai, S.-C.; Xu, Q.; Li, H.; Qin, Y.-F.; Song, L.-C.; Wang, C.-G.; Cui, W.-H.; Zheng, Z.; Yan, D.-W.; Li, Z.-J.; et al. NADPH Oxidase Isoforms Are Involved in Glucocorticoid-Induced Preosteoblast Apoptosis. Oxid. Med. Cell. Longev. 2019, 2019, 9192413. [Google Scholar] [CrossRef]
- Farley, J.R.; Stilt-Coffing, B. Apoptosis may determine the release of skeletal alkaline phosphatase activity from human osteoblast-line cells. Calcif. Tissue Int. 2001, 68, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Słupski, W.; Jawień, P.; Nowak, B. Botanicals in Postmenopausal Osteoporosis. Nutrients 2021, 13, 1609. [Google Scholar] [CrossRef] [PubMed]
- Rajput, R.; Wairkar, S.; Gaud, R. Nutraceuticals for better management of osteoporosis: An overview. J. Funct. Foods 2018, 47, 480–490. [Google Scholar] [CrossRef]
- Arjmandi, B.H.; Khalil, D.A.; Hollis, B.W. Ipriflavone, a Synthetic Phytoestrogen, Enhances Intestinal Calcium Transport In Vitro. Calcif. Tissue Int. 2000, 67, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.-J.; Wang, J.; Song, J.; Wang, C.-F.; Gu, J.-F.; Yuan, J.-R.; Zhang, L.; Jiang, J.; Feng, L.; Jia, X.-B. Beneficial Effects of a Flavonoid Fraction of Herba Epimedii on Bone Metabolism in Ovariectomized Rats. Planta Med. 2016, 82, 322–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, I.; Shino, A.; Shimizu, Y.; Tsukuda, R.; Shirakawa, Y.; Kinoshita, M. Effect of ipriflavone on glucocorticoid-induced osteoporosis in rats. Life Sci. 1986, 38, 951–958. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, A.; Xin, X.Z.; Peng, L.J.; Wang, J.; Cao, L.Y.; Jiang, X.Q. Osteogenic Induction of Low-dose Ipriflavone on Bone Marrow Mesenchymal Stem Cells Extracted from Osteoporosis Rats. Chin. J. Dent. Res. 2021, 24, 153–158. [Google Scholar]
- Hussien, H.M.; Ghareeb, D.A.; Ahmed, H.E.A.; Hafez, H.S.; Saleh, S.R. Pharmacological implications of ipriflavone against environmental metal–induced neurodegeneration and dementia in rats. Environ. Sci. Pollut. Res. 2021, 28, 65349–65362. [Google Scholar] [CrossRef]
- Savjani, K.T.; Gajjar, A.K.; Savjani, J.K. Drug Solubility: Importance and Enhancement Techniques. ISRN Pharm. 2012, 2012, 195727. [Google Scholar] [CrossRef] [Green Version]
- Sansai, K.; Na Takuathung, M.; Khatsri, R.; Teekachunhatean, S.; Hanprasertpong, N.; Koonrungsesomboon, N. Effects of isoflavone interventions on bone mineral density in postmenopausal women: A systematic review and meta-analysis of randomized controlled trials. Osteoporos. Int. 2020, 31, 1853–1864. [Google Scholar] [CrossRef]
- Hoppert, M. Metalloenzymes. In Encyclopedia of Geobiology; Reitner, J., Thiel, V., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 558–563. [Google Scholar]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-Dependent Antioxidant Enzymes: Actions and Properties of Selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominiak, A.; Wilkaniec, A.; Wroczyński, P.; Adamczyk, A. Selenium in the Therapy of Neurological Diseases. Where is it Going? Curr. Neuropharmacol. 2016, 14, 282–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castiglioni, S.; Cazzaniga, A.; Albisetti, W.; Maier, J.A.M. Magnesium and Osteoporosis: Current State of Knowledge and Future Research Directions. Nutrients 2013, 5, 3022–3033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, J.; Kim, S.-A.; Lim, K.; Shin, S. The association of potassium intake with bone mineral density and the prevalence of osteoporosis among older Korean adults. Nutr. Res. Pract. 2020, 14, 55–61. [Google Scholar] [CrossRef]
- Szentmihályi, K.; Kéry, Á.; Then, M.; Lakatos, B.; Sándor, Z.; Vinkler, P. Potassium-sodium ratio for the characterization of medicinal plant extracts with diuretic activity. Phytother. Res. 1998, 12, 163–166. [Google Scholar] [CrossRef]
- Loughrill, E.; Wray, D.; Christides, T.; Zand, N. Calcium to phosphorus ratio, essential elements and vitamin D content of infant foods in the UK: Possible implications for bone health. Matern. Child Nutr. 2017, 13, e12368. [Google Scholar] [CrossRef] [Green Version]
- Everett, D.W. Phosphorus and Calcium. 2021. Available online: https://www.encyclopedia.com/food/encyclopedias-almanacs-transcripts-and-maps/phosphorus-and-calcium (accessed on 24 April 2021).
- Torre, E. Molecular signaling mechanisms behind polyphenol-induced bone anabolism. Phytochem. Rev. 2017, 16, 1183–1226. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Urquiaga, I.; Leighton, F. Plant Polyphenol Antioxidants and Oxidative Stress. Biol. Res. 2000, 33, 55–64. [Google Scholar] [CrossRef]
- Liu, S.; Yang, L.; Mu, S.; Fu, Q. Epigallocatechin-3-Gallate Ameliorates Glucocorticoid-Induced Osteoporosis of Rats In Vivo and In Vitro. Front. Pharmacol. 2018, 9, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veliká, B.; Kron, I. Antioxidant properties of benzoic acid derivatives against Superoxide radical. Free Radic. Antioxid. 2012, 2, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Prahalathan, P.; Raja, B. Syringic acid ameliorates l-NAME-induced hypertension by reducing oxidative stress. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2012, 385, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, N.; Orihuela-Campos, R.C.; Inagaki, Y.; Fukui, M.; Nagata, T.; Ito, H.O. Resveratrol improves oxidative stress and prevents the progression of periodontitis via the activation of the Sirt1/AMPK and the Nrf2/antioxidant defense pathways in a rat periodontitis model. Free Radic. Biol Med. 2014, 75, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Pugazhendhi, D.; Pope, G.S.; Darbre, P.D. Oestrogenic activity of p-hydroxybenzoic acid (common metabolite of paraben esters) and methylparaben in human breast cancer cell lines. J. Appl. Toxicol. 2005, 25, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Özatik, F.Y.; Erol, K.; Özatik, O. Estrogen modulating effects of resveratrol in female rats. Marmara Med. J. 2016, 29, 95. [Google Scholar] [CrossRef] [Green Version]
- Zych, M.; Folwarczna, J.; Trzeciak, H.I. Natural phenolic acids may increase serum estradiol level in ovariectomized rats. Acta Biochim. Pol. 2009, 56, 503–507. [Google Scholar] [CrossRef]
- Xiao, H.-H.; Gao, Q.-G.; Zhang, Y.; Wong, K.-C.; Dai, Y.; Yao, X.-S.; Wong, M.-S. Vanillic acid exerts oestrogen-like activities in osteoblast-like UMR 106 cells through MAP kinase (MEK/ERK)-mediated ER signaling pathway. J. Steroid Biochem. Mol. Biol. 2014, 144, 382–391. [Google Scholar] [CrossRef]
- Dai, Z.; Li, Y.; Quarles, L.D.; Song, T.; Pan, W.; Zhou, H.; Xiao, Z. Resveratrol enhances proliferation and osteoblastic differentiation in human mesenchymal stem cells via ER-dependent ERK1/2 activation. Phytomedicine 2007, 14, 806–814. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, L.-M.; Guo, C.; Han, J.-F. Resveratrol promotes osteoblastic differentiation in a rat model of postmenopausal osteoporosis by regulating autophagy. Nutr. Metab. 2020, 17, 29. [Google Scholar] [CrossRef] [Green Version]
- Melguizo-Rodríguez, L.; Manzano-Moreno, F.J.; Illescas-Montes, R.; Ramos-Torrecillas, J.; de Luna-Bertos, E.; Ruiz, C.; García-Martínez, O. Bone Protective Effect of Extra-Virgin Olive Oil Phenolic Compounds by Modulating Osteoblast Gene Expression. Nutrients 2019, 11, 1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, J.; Li, Q.; Luo, X.; Li, J.; Guo, L.; Xue, H.; Wu, G. Epigallocatechin-3-gallate protects against secondary osteoporosis in a mouse model via the Wnt/β-catenin signaling pathway. Mol. Med. Rep. 2018, 18, 4555–4562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-D.; Shi, Y.-M.; Li, L.; Guo, J.-D.; Zhang, Y.-P.; Hou, S.-X. Treatment with resveratrol attenuates sublesional bone loss in spinal cord-injured rats. Br. J. Pharmacol. 2013, 170, 796–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Onuma, H.; Shigihara, T.; Kimura, E.; Fukuta, Y.; Shirasaka, N.; Moriyama, T.; Homma, Y. Anti-osteoporotic effects of syringic acid and vanilic acid in the extracts of waste beds after mushroom cultivation. J. Biosci. Bioeng. 2019, 128, 622–629. [Google Scholar] [CrossRef]
- Shakibaei, M.; Buhrmann, C.; Mobasheri, A. Resveratrol-mediated SIRT-1 interactions with p300 modulate receptor activator of NF-kappaB ligand (RANKL) activation of NF-kappaB signaling and inhibit osteoclastogenesis in bone-derived cells. J. Biol. Chem. 2011, 286, 11492–11505. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-T.; Kang, L.; Wang, C.-Z.; Huang, P.-J.; Huang, H.-T.; Lin, S.-Y.; Chou, S.-H.; Lu, C.-C.; Shen, P.-C.; Lin, Y.-S.; et al. (−)-Epigallocatechin-3-Gallate Decreases Osteoclastogenesis via Modulation of RANKL and Osteoprotegrin. Molecules 2019, 24, 156. [Google Scholar] [CrossRef] [Green Version]
- Kwak, S.C.; Lee, C.; Kim, J.-Y.; Oh, H.M.; So, H.-S.; Lee, M.S.; Rho, M.C.; Oh, J. Chlorogenic Acid Inhibits Osteoclast Differentiation and Bone Resorption by Down-Regulation of Receptor Activator of Nuclear Factor Kappa-B Ligand-Induced Nuclear Factor of Activated T Cells c1 Expression. Biol. Pharm. Bull. 2013, 36, 1779–1786. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.P.; Lin, S.J.; Wan, W.B.; Zuo, H.L.; Yao, F.F.; Ruan, H.B.; Xu, J.; Song, W.; Zhou, Y.C.; Wen, S.Y.; et al. Chlorogenic Acid Prevents Osteoporosis by Shp2/PI3K/Akt Pathway in Ovariectomized Rats. PLoS ONE 2016, 11, e0166751. [Google Scholar] [CrossRef]
- Lin, X.; Yuan, G.; Li, Z.; Zhou, M.; Hu, X.; Song, F.; Shao, S.; Fu, F.; Zhao, J.; Xu, J.; et al. Ellagic acid protects ovariectomy-induced bone loss in mice by inhibiting osteoclast differentiation and bone resorption. J. Cell. Physiol. 2020, 235, 5951–5961. [Google Scholar] [CrossRef]
- Lee, H.-H.; Jang, J.-W.; Lee, J.-K.; Park, C.-K. Rutin Improves Bone Histomorphometric Values by Reduction of Osteoclastic Activity in Osteoporosis Mouse Model Induced by Bilateral Ovariectomy. J. Korean Neurosurg. Soc. 2020, 63, 433–443. [Google Scholar] [CrossRef] [Green Version]
- Folwarczna, J.; Zych, M.; Burczyk, J.; Trzeciak, H.; Trzeciak, H.I. Effects of Natural Phenolic Acids on the Skeletal System of Ovariectomized Rats. Planta Med. 2009, 75, 1567–1572. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Primer Sequence | Annealing Temperature (°C) | Number of Cycles | Ref. | |
|---|---|---|---|---|---|---|
| GAPDH | NM_017008.4 | F | 5′-AGATCCACAACGGATACATT-3′ | 52 | 35 | [36] |
| R | 5′-TCCCTCAAGATTGTCAGCAA-3′ | |||||
| OPG | NM_012870.2 | F | 5′-GTTCTTGCACAGCTTCACCA-3′ | 54 | 40 | [37] |
| R | 5′-AAACAGCCCAGTGACCATTC-3′ | |||||
| RANKL | NM_057149.1 | F | 5′-ACCAGCATCAAAATCCCAAG-3′ | 52 | 35 | [37] |
| R | 5′-GGCCGCTAATTTCCTCACCA-3′ | |||||
| DKK1 | NM_001106350.1 | F | 5′-GCTGCATGAGGCACGCTAT-3′ | 55 | 35 | [38] |
| R | 5′-AGGGCATGCATATTCCGTTT-3′ | |||||
| Wnt5a | NM_022631.3 | F | 5′-CCATGAAGAAGCCCATTGGAATA-3′ | 60 | 40 | [39] |
| R | 5′-GGCCAAAGCCATTAGGAAGAA-3′ | |||||
| SOST | NM_030584.2 | F | 5′-GTGCAAGTGCAAGCGCCTCA-3′ | 60 | 40 | [40] |
| R | 5′-GCTCCGCCTGGTTGGCTTTG-3′ | |||||
| Runx2 | NM_001278483.1 | F | 5′-AGTGTGTGTGTCCGCATGAT-3′ | 56 | 40 | [41] |
| R | 5′-CCACTTGGGGTCTAAGAACG-3′ | |||||
| Osx | NM_181374.2 | F | 5′-TGAGGAAGAAGCCCATTCAC-3′ | 53.5 | 40 | [42] |
| R | 5′-ACTTCTTCTCCCGGGTGTG-3′ | |||||
| COL1A1 | NM_053304.1 | F | 5′-CAAGGACTATGAAGTTGATGC-3′ | 43 | 40 | [40] |
| R | 5′-ACCAGTAGAGAAATCGCAGT-3′ | |||||
| Scavenging Activity (IC50, µg/mL) | |||
|---|---|---|---|
| DPPH | 112.17 ± 5.11 | ||
| H2O2 radical | 44.38 ± 2.56 | ||
| Constituents | Concentration | ||
| Phytochemicals | µg equivalent/ mg extract | LDPE (150 mg/kg) | HDPE (300 mg/kg) |
| Total phenolics | 301.97 ± 5.16 | 45,295 ± 774 | 90,591 ± 1548 |
| Total flavonoids | 5.10 ± 2.75 | 765 ± 412.5 | 1530 ± 825 |
| HPLC Analysis of Phenolic Compounds | |||
| Constituents | (μg/g extract) | LDPE (μg/150 mg/kg) | HDPE (μg/300 mg/kg) |
| Quinol | 703.81 ± 8.5 | 105.6 ± 1.3 | 211.14 ± 2.55 |
| p-Hydroxybenzoic acid | 1042.01 ± 15.9 | 156.3 ± 2.25 | 312.6 ± 4.77 |
| Catechin | 70.76 ± 2.0 | 10.61 ± 0.3 | 21.23 ± 0.6 |
| Chlorogenic | 26.67 ± 1.5 | 4.0 ± 0.225 | 8.0 ± 0.45 |
| Vanillicacid | 625.43 ± 11.6 | 93.81 ± 1.74 | 187.63 ± 3.48 |
| Caffeic acid | 89.58 ± 7.0 | 13.44 ± 1.05 | 26.87 ± 2.1 |
| Syringic acid | 119.03 ± 3.9 | 17.85 ± 0.59 | 35.71 ± 1.17 |
| p-Coumaric acid | 19.24 ± 1.9 | 2.89 ± 0.29 | 5.77 ± 0.57 |
| Benzoic acid | 587.45 ± 17.6 | 88.12 ± 2.64 | 176.24 ± 5.28 |
| Ferulic acid | 21.16 ± 3.2 | 3.17 ± 0.48 | 6.35 ± 0.96 |
| Rutin | 17.55 ± 2.9 | 2.63 ± 0.44 | 5.27 ± 0.87 |
| Ellagic acid | 24.89 ± 5.0 | 3.73 ± 0.75 | 7.74 ± 1.5 |
| o-Coumaric acid | 10.72 ± 1.1 | 1.61 ± 0.17 | 3.22 ± 0.33 |
| Resveratrol | 822.57 ± 13.7 | 123.39 ± 2.1 | 246.77 ± 4.11 |
| Total | 4180.86 ± 18.5 | 627.13 ± 2.78 | 1254.26 ± 5.55 |
| Mineral Content | DPE (µg/g Extract) | LDPE (150 mg/kg) | HDPE (300 mg/kg) |
|---|---|---|---|
| Ca | 44,675.673 | 6701.35 | 13,402.7 |
| Cu | 1121.442 | 168.22 | 336.43 |
| Fe | 1488.990 | 223.35 | 446.7 |
| K | 236,567.019 | 35,485.053 | 70,970.11 |
| Mg | 37,368.077 | 5605.21 | 11,210.42 |
| Mn | 133.269 | 19.99 | 39.98 |
| Na | 35,755.288 | 5363.29 | 10,726.59 |
| P | 34,262.933 | 5139.44 | 10,278.88 |
| Se | 20.817 | 3.12 | 6.25 |
| Zn | 697.163 | 104.57 | 209.15 |
| MDA Level (µmol/mg Protein) | NO Level (µmol/mg Protein) | GSH Content (µmol/mg Protein) | GPx Activity (U/mg Protein) | GST Activity (u/mg Protein) | SOD Activity (U/mg Protein) | |
|---|---|---|---|---|---|---|
| Control | 1.9534 ± 0.231 | 369.22 ± 12.425 | 10.2953 ± 0.483 | 58.1272 ± 8.152 | 0.727 ± 0.041 | 1.337 ± 0.059 |
| LDPE | 1.2168 ± 0.055 * | 296.44 ± 50.414 * | 10.9006 ± 0.729 | 54.4612 ± 2.901 | 0.7637 ± 0.044 | 1.3448 ± 0.085 * |
| HDPE | 1.063 ± 0.129 * | 235.23 ± 12.130 * | 13.8351 ± 0.492 * | 56.0947 ± 4.179 | 1.3592 ± 0.044 * | 1.6993 ± 0.040 * |
| DEXA | 2.2015 ± 0.033 * | 861.26 ± 17.011 * | 6.287 ± 0.290 * | 23.257 ± 1.874 * | 0.389 ± 0.028 * | 0.8987 ± 0.057 * |
| LDPE + DEXA | 1.0653 ± 0.074 *# | 365.8 ± 24.901 # | 15.9467 ± 0.646 *# | 72.4151 ± 4.680 *# | 1.0359 ± 0.041 *# | 1.5423 ± 0.062 *# |
| HDPE + DEXA | 1.5578 ± 0.084 *# | 338.29 ± 22.747 # | 17.9689 ± 0.637 *# | 57.4594 ± 3.724 # | 1.34 ± 0.067 *# | 1.4421 ± 0.044 *# |
| IPRI + DEXA | 1.1083 ± 0.050 *# | 441.28 ± 22.444 *# | 6.608 ± 1.116 * | 55.2588 ± 3.020 # | 1.0398 ± 0.017 #* | 1.336 ± 0.125 # |
| Epiphyseal Plate Thickness (µm) | Trabecular Bone Quality Score * | Trabecular Bone Area Score ** | Osteoblast Mean Count/HPF | |
|---|---|---|---|---|
| Control | 277 | 0 | 0 | 18 |
| LDPE | 233 | 0 | 0 | 16 |
| HDPE | 229 | 0 | 0 | 16 |
| DEXA | 80 | 2 | 3 | 4 |
| LDPE + DEXA | 210 | 0 | 1 | 18 |
| HDPE + DEXA | 181 | 1 | 1 | 13 |
| IPRI + DEXA | 204 | 1 | 1 | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, S.R.; Ghareeb, D.A.; Masoud, A.A.; Sheta, E.; Nabil, M.; Masoud, I.M.; Maher, A.M. Phoenix dactilyfera L. Pits Extract Restored Bone Homeostasis in Glucocorticoid-Induced Osteoporotic Animal Model through the Antioxidant Effect and Wnt5a Non-Canonical Signaling. Antioxidants 2022, 11, 508. https://doi.org/10.3390/antiox11030508
Saleh SR, Ghareeb DA, Masoud AA, Sheta E, Nabil M, Masoud IM, Maher AM. Phoenix dactilyfera L. Pits Extract Restored Bone Homeostasis in Glucocorticoid-Induced Osteoporotic Animal Model through the Antioxidant Effect and Wnt5a Non-Canonical Signaling. Antioxidants. 2022; 11(3):508. https://doi.org/10.3390/antiox11030508
Chicago/Turabian StyleSaleh, Samar R., Doaa A. Ghareeb, Aliaa A. Masoud, Eman Sheta, Mohamed Nabil, Inas M. Masoud, and Adham M. Maher. 2022. "Phoenix dactilyfera L. Pits Extract Restored Bone Homeostasis in Glucocorticoid-Induced Osteoporotic Animal Model through the Antioxidant Effect and Wnt5a Non-Canonical Signaling" Antioxidants 11, no. 3: 508. https://doi.org/10.3390/antiox11030508
APA StyleSaleh, S. R., Ghareeb, D. A., Masoud, A. A., Sheta, E., Nabil, M., Masoud, I. M., & Maher, A. M. (2022). Phoenix dactilyfera L. Pits Extract Restored Bone Homeostasis in Glucocorticoid-Induced Osteoporotic Animal Model through the Antioxidant Effect and Wnt5a Non-Canonical Signaling. Antioxidants, 11(3), 508. https://doi.org/10.3390/antiox11030508