Abstract
Free radicals are formed as a part of normal metabolic activities but are neutralized by the endogenous antioxidants present in cells/tissue, thus maintaining the redox balance. This redox balance is disrupted in certain neuropathophysiological conditions, causing oxidative stress, which is implicated in several progressive neurodegenerative diseases. Following neuronal injury, secondary injury progression is also caused by excessive production of free radicals. Highly reactive free radicals, mainly the reactive oxygen species (ROS) and reactive nitrogen species (RNS), damage the cell membrane, proteins, and DNA, which triggers a self-propagating inflammatory cascade of degenerative events. Dysfunctional mitochondria under oxidative stress conditions are considered a key mediator in progressive neurodegeneration. Exogenous delivery of antioxidants holds promise to alleviate oxidative stress to regain the redox balance. In this regard, natural and synthetic antioxidants have been evaluated. Despite promising results in preclinical studies, clinical translation of antioxidants as a therapy to treat neurodegenerative diseases remains elusive. The issues could be their low bioavailability, instability, limited transport to the target tissue, and/or poor antioxidant capacity, requiring repeated and high dosing, which cannot be administered to humans because of dose-limiting toxicity. Our laboratory is investigating nanoparticle-mediated delivery of antioxidant enzymes to address some of the above issues. Apart from being endogenous, the main advantage of antioxidant enzymes is their catalytic mechanism of action; hence, they are significantly more effective at lower doses in detoxifying the deleterious effects of free radicals than nonenzymatic antioxidants. This review provides a comprehensive analysis of the potential of antioxidant therapy, challenges in their clinical translation, and the role nanoparticles/drug delivery systems could play in addressing these challenges.
1. Introduction
Free radicals are generated during pivotal biological processes such as metabolic reactions, cell signaling, and gene transcription [1]. Cellular organelles such as mitochondria, peroxisomes, lysosomes, microsomes, endoplasmic reticulum, plasma membrane, and phagocytic cells are also the source of free radical production [2,3]. External factors such as environmental pollutants, radiation, smoking, heavy-metal exposure, diet, and physical exercise also contribute to the production of free radicals [4,5]. Under normal conditions, the innate antioxidative defense system that includes various enzymatic and nonenzymatic antioxidants neutralize free radicals, thus maintaining the redox balance [6]. This balance is disrupted under certain pathological conditions such as genetic mutations, inflammation, injury, ischemia/reperfusion, etc. [7,8,9]. Excessive free radicals formed overwhelm the endogenous antioxidant defense mechanism, thus causing oxidative stress which downregulates the endogenous defense system [10,11]. Neuronal cells are particularly susceptible to damage due to free radicals, as they contain high levels of unsaturated lipids that are susceptible to oxidation and the presence of high levels of redox-active transition metals that catalyze the formation of free radicals [12]. The central nervous system (CNS) has high metabolic activity and, hence, a high oxygen demand, which favors free radical formation [13]. Metabolism of neurotransmitters also produces free radicals [14]. The CNS also has a relatively weaker antioxidant defense than other organs (e.g., liver) which makes it more susceptible to oxidative stress than other organs [15,16]. Under oxidative stress condition, dysfunctional mitochondria are unable to meet the high energy need of neuronal cells for their normal biochemical and physiological functions; hence they become vulnerable to rapid cell death [17].
Pro-oxidants or free radicals are usually those atoms or molecules that contain an unpaired electron in their outermost orbit and can be formed when oxygen interacts with certain molecules [18]. These free radicals are very unstable but highly reactive and, when they interact with other molecules, create additional free radicals, initiating a self-propagating chain reaction of free radical formation [18]. Free radicals contain reactive oxygen species (ROS) and reactive nitrogen species (RNS). ROS are chemically reactive molecules containing oxygen, whereas RNS includes nitrogen (N) and oxygen (O) atoms. The ROS and RNS produced in cells comprise both free radical and non-free radical species and include hydrogen peroxide (H2O2), nitric oxide (•NO), nitrogen dioxide (•NO2), hydroxyl radical (•OH), superoxide anion (O2•−), peroxynitrite (OONO−), hypochlorous acid (HClO), etc. The •OH radical, produced from H2O2 in the metal-catalyzed (free Fe and Cu) redox reactions such as Fenton reaction, is particularly unstable and reacts rapidly and nonspecifically with most biological molecules [3].
1.1. Endogenous and Exogenous Sources of Free Radicals
There are multiple cellular processes and biochemical reactions that produce free radicals as a part of normal cellular function. For e.g., during Electron Transport Chain (ETC) and its five integrated mitochondrial complexes (I, II, III, IV, and V), reduction of O2 to H2O by cytochrome c oxidase prematurely generates ROS such as singlet oxygen (1O2), O2•−, •OH, and H2O2 [19,20,21]. Intracellular organelle, peroxisomes, responsible for degradation of fatty acids, generate H2O2 as a byproduct [22]. Neutrophils that contain myeloperoxidase (MPO) uses H2O2 and halides (Cl−, Br−, and I−) or pseudohalide (SCN−) ions to catalyze the production of free radicals [23]. Phagocytic cells (neutrophils, macrophages, and monocytes) while defending the CNS against invading microorganisms or clearing the dead cell debris produces ROS [24]. Cytochrome P450 is another intracellular enzyme present in microsomes and the endoplasmic reticulum catalyzes the ROS formation [25]. Cytosolic enzymes such as xanthine oxidase (XO) during the catalytic oxidation of hypoxanthine to xanthine and Prostaglandin H Synthase (PHS) from arachidonic acid to prostaglandin generate ROS [24]. In addition, environmental pollutants; ionizing radiation (UV-rays, X-rays, γ-rays, and infrared or electromagnetic waves); smoking; long-term chemical exposure like pesticides, insecticides, or industrial solvents; heavy or transition metals (Cu, Fe, Mn, As, Cd, Pb, and Hg); diet; and physical exercise contribute to the production of ROS/RNS [26,27,28,29,30,31,32,33,34,35,36,37,38,39].
1.2. Free Radicals: A Double Edge Sword
Under normal physiological conditions, low levels of ROS are essential for the regulation of critical signaling pathways involved in cell growth, proliferation, differentiation, survival, regulation of blood pressure, cognitive function, immunity, and maintaining normal antioxidant defense mechanisms of the body [40]. RNS in the CNS regulate cerebral blood flow and memory and plays a significant role in maintaining the immune system and cytokine production [41]. However, excess ROS and RNS, which are the byproducts of the oxygen and nitrogen-rich tissue environment in the body, if not neutralized by the endogenous antioxidants, results in oxidative/nitrosative stress [42]. Such conditions can damage cells by starting a chemical chain reaction and modifying biomolecules, i.e., lipids, proteins, and DNA [43]. The ROS produced by mitochondria can accelerate the oxidation of polyunsaturated fatty acids in the cell membrane lipids, a process known as lipid peroxidation (LPO) that changes the cell membrane structure, impairing its integrity, thus affecting cell signaling. The LPO products such as F2-isoprostanes, malondialdehyde (MDA), 4-hydroxynonenal (4-HNE), and oxidized low-density lipoproteins (LDL) can further damage proteins and nucleic acid bases [44]. With oxidative stress, multiple changes can occur such as mitochondrial DNA mutation, impairment in the mitochondrial respiratory chain, and change in membrane permeability influencing Ca2+ homeostasis [20,45,46,47].
2. Oxidative Stress and Neurodegenerative Diseases
Cell damage triggers a cascade of degenerative events via mitochondrial dysfunction, neuroinflammation, apoptosis, and tissue necrosis [20,48,49]. Oxidative stress-induced homeostatic dysregulation remains a central component of several neurodegenerative diseases such as Alzheimer’s disease (AD), Parkinson’s disease (PD), and Amyotrophic Lateral Sclerosis (ALS) [7]. Examples of injury-triggered neurodegenerative diseases include stroke, spinal cord injury (SCI), peripheral nerve injury (PNI), etc. [8,9]. The common link between these neurodegeneration conditions is oxidative stress, ineffective antioxidant defense, and mitochondrial dysfunction (Figure 1).
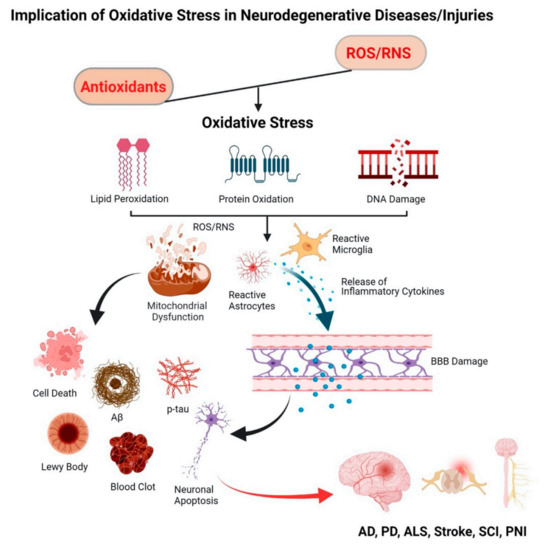
Figure 1.
Schematic representing the effect of oxidative stress in neurodegenerative diseases. Imbalance in the level of ROS/RNS and antioxidants leads to an oxidative stress condition that causes damage to cellular biomolecules, i.e., lipids, proteins, and DNA. Mitochondrial dysfunction and accumulation of activated astrocytes and microglia release inflammatory cytokines and chemokines, promoting cellular apoptosis and tissue death.
2.1. Progressive Neurodegenerative Diseases
2.1.1. Alzheimer’s Disease (AD)
AD, a leading cause of dementia, is characterized by a progressive decline in cognitive function [48]. Amyloid beta (Aβ) plaques, neurofibrillary tangles (NFTs), hyperphosphorylated microtubule-associated protein tau, and neuronal loss within the brain are specific histopathological hallmarks of the AD [49]. Prior to the development of plaque pathology, oxidative stress has been recognized as the key player in the etiology of AD, contributing to mitochondrial dysfunction in synapses and neurons, and in Aβ production [50,51]. In fact, the concept of oxidative stress in AD was originally derived from the “free radical theory of aging”, meaning that free radicals play a central role in the aging process [52]. Mitochondrial dysfunction in AD includes impaired mitochondrial complexes [53,54,55,56], malfunctioning of F1Fo adenosine triphosphate (ATP) synthase, which is involved in oxidative phosphorylation [57,58], and damage to the promoter of the mitochondrial ATP synthase gene that controls ATP generation [59,60]. Further, dysfunctional mitochondria produce 4-HNE that upregulates γ-secretase complex and promotes cleavage of the amyloid precursor protein (APP), leading to Aβ accumulation [61,62]. In addition, increased Ca2+ and ROS levels lead to a buildup of p-tau aggregates which are toxic and are considered as one of the defining pathological hallmarks of the AD [63]. ROS also play a pivotal role in the stress kinases like the phospho-c-Jun N-terminal kinase 1 (p-JNK) pathway which is linked to tau hyperphosphorylation and cell death in response to Aβ accumulation [64]. Further, oxidative stress reduces the activities of antioxidants, i.e., superoxide dismutase (SOD), catalase (CAT), and glutathione S-transferase (GST), thus weakening the endogenous antioxidant defense of the CNS [65]. The increased levels of LPO under oxidative stress are strongly associated with neurotoxicity in AD [50] as it leads to an increase in amyloidogenesis through upregulation of β-secretase expression [66]. Although there are several downstream degenerative events, it appears that mitochondrial dysfunction and oxidative stress are the key triggering factors in the pathogenesis of AD.
2.1.2. Parkinson’s Disease (PD)
PD is the second-most common neurodegenerative disease after AD that causes both motor and nonmotor symptoms [67]. The pathology of PD is driven by the accumulation and aggregation of α-synuclein, a presynaptic neuronal protein in the nervous system [68]. The mechanisms associated with the pathogenesis of PD include aberrant protein homeostasis, bioenergetic impairment, and oxidative stress [69]. Oxidative stress is associated with α-synuclein protein aggregation [64]. The cascade of events leading to degeneration of dopaminergic neurons in PD is also linked to oxidative stress [70]. Analysis of the postmortem brain tissue of the victims of PD shows elevated levels of oxidative stress markers such as 4-HNE, protein carbonyl, 8-hydroxy-2′-deoxyguanosine, and 8-hydroxy-guanosine [71]. In addition, oxidative stress is associated with the formation of Lewy bodies, which are the clumps of protein in the PD brain [72]. Experimental evidence in PD models suggests that oxidative stress in the dopaminergic neurons activates p38 mitogen-activated protein kinase (p38 MAPK) pathway that ultimately leads to apoptosis of the brain cells [73].
2.1.3. Amyotrophic Lateral Sclerosis (ALS)
ALS is also known as Lou Gehrig’s disease, in which motor neurons in the brain, brain stem, and spinal cord are damaged, resulting in muscle weakness, atrophy, paralysis, and premature death [74]. Oxidative stress, mitochondrial dysfunction, and mutations in the genes that act on mitochondrial processes are involved in the pathophysiology of the ALS [75,76]. Most of the familial ALS patients (15–20%) have mutations in the superoxide dismutase 1 (SOD1) gene, which plays an important role in the defense mechanism against oxidative stress [77]. More than 150 ALS-related SOD1 gene mutations have been discovered in various parts of the enzyme, which result in protein misfolding and aggregation, increased ROS production, and redox system disequilibrium, ultimately resulting in nerve cell loss [77,78]. ALS is also linked to several interrelated risk factors, such as neuroinflammation, excitotoxicity, mitochondrial dysfunction/dysregulation, and endoplasmic reticulum stress [79,80,81]. Considerably high oxidative stress biomarkers such as MDA, 8-hydroxyguanosine, and advanced oxidation protein products are found in ALS patients [82]. In sporadic ALS patients, cystine/glutamate antiporter overexpression was observed that causes increased oxidative stress and extracellular glutamate accumulation [83]. In addition, dysregulation of the retinoic acid (RA) signaling pathway, a product of vitamin A, contributes to the death of motor neurons [84].
2.2. Injury-Induced Oxidative Stress
Neuronal tissue injury, physical or due to ischemic condition, is known to induce oxidative stress that triggers progressive degeneration, known as secondary injury.
2.2.1. Stroke
In stroke, thrombus formation in cerebral blood vessels creates an ischemic condition, triggering free radical formation and tissue damage (Figure 2) [85]. Resumption of blood supply to the ischemic region further exuberates the condition as more free radicals are formed, termed “reperfusion injury or reoxygenation injury” [86]. Collectively, it is referred to as the ischemia/reperfusion (I/R) injury [86]. Oxidative stress leads to mitochondrial dysfunction, neuroinflammation, and glutamate excitotoxicity, resulting in the blood-brain barrier (BBB) damage, apoptosis/necrosis of neurons, and supporting cellular elements (glial cells and vessels) [87,88,89]. These are the prominent features of neurodegeneration in the stroke-related cerebral pathology [90,91,92,93]. Further, excessive ROS production or impaired ROS degradation [94,95] stimulates vasoconstriction, increased platelet aggregation, and endothelial cell permeability, thereby affecting cerebral blood circulation [96]. Activation of matrix metalloproteinases (MMPs) disrupts the cerebral extracellular matrix (ECM), which causes immunocyte infiltration and neuroinflammation, culminating in the breakdown of the neurovascular unit (NVU), leading to hemorrhage and edema [97,98].
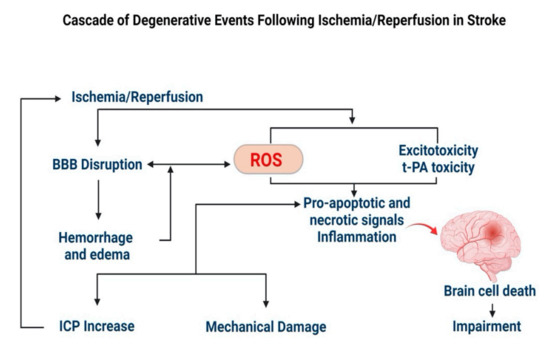
Figure 2.
ROS-mediated degenerative events during a stroke. Excessive production of ROS during I/R injury leads to mechanical damage to the brain due to breakdown of the BBB, hemorrhage and edema, causing a build-up of intracranial pressure (ICP). The biochemical changes lead to inflammation and progression of apoptosis. Therefore, excess ROS formed during I/R is considered a target to inhibit the progression of secondary brain damage.
2.2.2. Spinal Cord Injury (SCI)
SCI is another common form of neuronal injury that causes neurological dysfunctions [99] and is characterized by an initial primary injury followed by the secondary phase of injury (Figure 3) [100]. Primary injury results immediately from the initial trauma causing damage to the blood vessels and axons [101]. In contrast, secondary injury is the indirect result of the primary injury that involves inflammation and oxidative stress [10]. The secondary injury progression occurs not only at the site of impact, but it spreads along the entire spinal cord, including the faraway segments of the spinal cord that are not impacted, making the condition more devastating and debilitating with time [101]. Following injury, the elevated levels of ROS and the consequent oxidative stress are considered critical events associated with the secondary injury progression [102]. Under oxidative stress condition, dysfunctional mitochondria become the source of ROS [103] that cause a further cascade of degenerative processes, particularly curtailing ATP production required for normal cellular functioning, thus promoting apoptosis [103]. The excess ROS alters cell functions by modulating ion channels, followed by excessive accumulation of intracellular calcium ions that eventually causes excitotoxicity [104]. Oxidative stress damages the microvascular endothelium that reduces the spinal cord white matter blood flow, resulting in ischemic injury [105].
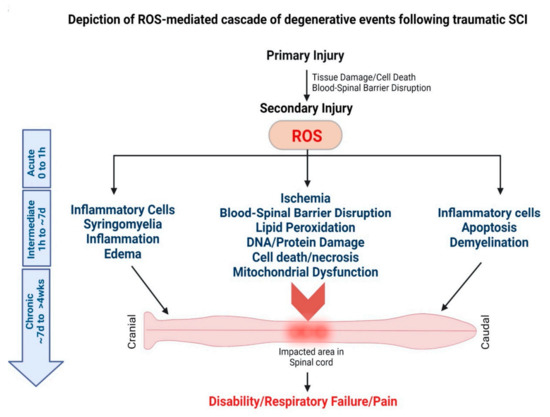
Figure 3.
Secondary injury cascade following spinal cord injury. Traumatic injury to the spinal cord leads to secondary injury progression that affects the lesion site and the entire spinal cord, including the cranial and caudal segments of the spinal cord. Following injury, excessive production of ROS is considered to trigger the secondary injury cascade of progressive degeneration that affects the entire spinal cord.
2.2.3. Peripheral Nerve Injury (PNI)
The peripheral nervous system (PNS) is a bundle of long nerve fibers that connect different parts of the body with the CNS. Damage to the peripheral nerves due to trauma and compression can cause impairment in the brain’s communication with the target organs [106]. These injuries affect motor and sensory behaviors, perception, consciousness, and sensations of the skin and joints [106]. The most common symptoms of PNI are the defects in sensory and motor function that can lead to complete paralysis of the affected limb or the development of an intractable neuropathic pain [107]. Many surgical procedures, such as oral and maxillofacial surgery, can also cause injury to the peripheral nerves [108]. The major component of the mechanism and pathogenies of PNI involves oxidative stress and inflammation that exacerbates neural damages and plays a negative role in the regeneration process [109]. Experimental evidence at the preclinical level has demonstrated that inhibiting oxidative stress could help improve functional recovery by accelerating the repair processes [110,111,112,113,114].
Other neurodegenerative diseases implicated due to oxidative stress are: vascular dementia [115], Down syndrome [116], Autism [117], attention-deficit/hyperactivity disorder (ADHD) [118], Huntington’s disease (HD) [119], multiple sclerosis (MS) [120], depression [121], and epilepsy [122]. Similarly, in traumatic brain injury (TBI) [123], progressive degeneration occurs due to the accumulation of excessive free radicals, glutamate release, Ca2+ overload, mitochondrial dysfunction, leading to apoptosis/necrosis [123].
3. Antioxidants
From the above review of the etiology of different neurodegenerative diseases, oxidative stress is considered as the key component, whether these are chronic neurodegenerative conditions such as AD, PD, or ALS or caused by neuronal tissue injury, such as in stroke, SCI, or PNI. Dysfunctional mitochondria under oxidative stress become the main source of free radical formation and deplete the energy needed for normal cellular function, leading to inflammation and cell death [124]. Another set of literature data indicates that dysfunctional mitochondria cause oxidative stress [125]. Thus, there is a complexity in understanding the root cause, whether oxidative stress leads to mitochondrial dysfunction, or it is mitochondrial dysfunction that leads to oxidative stress [49,126,127]. Despite ambiguity on the root cause of oxidative stress, it is hypothesized that an effective treatment based on antioxidants can alleviate oxidative stress and regain the redox balance that can attenuate mitochondrial dysfunction and curtail the downstream cascade of degeneration [126]. It is also contemplated that oxidative stress-free condition can promote regeneration and healing by the endogenous mechanisms, such as by promoting migration and differentiation of progenitor and stem cells [127]. In addition, an oxidative stress-free environment could promote differentiation of macrophages preferentially to M2 phenotype, which contains growth factors and can promote healing, rather than to M1 phenotype, which contains degenerative inflammatory cytokines [128]. With this in consideration, natural and synthetic antioxidants have been evaluated in preclinical model studies and clinical trials [129].
Antioxidants can reduce oxidative stress by quenching/scavenging free radical intermediates, thereby preventing oxidative chain reactions from propagating [4]. These antioxidants predominantly include various endogenous antioxidant enzymes with their substrates or coenzymes and nonenzymatic antioxidants, along with exogenous (natural and synthetic) antioxidant sources that maintain the redox equilibrium in the biological system [130]. Endogenous antioxidant activity is directly regulated by nuclear factor erythroid 2-related factor 2 (Nrf2). It is a ubiquitous redox-sensitive transcription factor that stimulates the expression of antioxidant response element (ARE)-containing gene promoters involved in ROS detoxification. These promoters are heme oxygenase 1 (HO-1), glutathione s-transferase (GST), and NADPH quinine oxidoreductase 1 (NQO1) (Figure 4) [131]. Thus, the Nrf2 pathway is an important aspect of the cellular defense mechanism against oxidative stress [132].
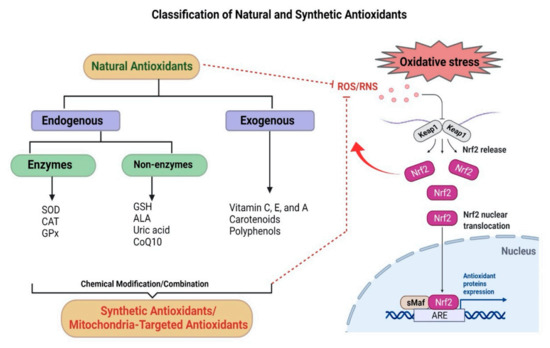
Figure 4.
Natural and synthetic antioxidants: Classification of natural and synthetic antioxidants and the endogenous Nrf2 pathway, which regulates the activation of ARE genes. Kelch-like ECH-associated protein 1 (Keap1) represents a negative regulator of Nrf2. Under physiological conditions, Keap1 forms a ubiquitin E3 ligase complex with Cullin3 in the cytoplasm that targets Nrf2 for polyubiquitination and rapid proteasomal degradation. During oxidative stress, cysteines in Keap1 are modified and inactivated, and Nrf2 can quickly translocate into the nucleus, where it binds to small musculoaponeurotic fibrosarcoma oncogene homolog (sMaf) proteins, upregulates downstream ARE genes, and maintains redox homeostasis.
3.1. Endogenous Antioxidants
The inherent antioxidative protective mechanism is composed of antioxidant enzymes such as superoxide dismutase (SOD), catalases (CAT), and glutathione peroxidases (GPx-1) [130]. In addition, low-molecular-weight nonenzymatic antioxidants include thiol antioxidants (Glutathione, α-lipoic acid), uric acid, and coenzyme Q10 (CoQ10) [133]. By scavenging excess ROS and limiting further generation of free radical species, these antioxidants collectively can prevent the detrimental effects of oxidative stress [134]. Antioxidants can also neutralize any free radical or a reactive species that can produce new free radicals [135].
3.1.1. Antioxidant Enzymes
Superoxide Dismutase (SOD): SOD is a heterogeneous metalloprotein enzyme having four different types of metals at the center, i.e., Cu, Zn, Fe, Mg, and Ni with a crystalline nature. In the presence of these metal ion cofactors, SOD located in the cytosol and mitochondria catalytically converts O2•− into O2 and H2O2 [136]. O2•− is detoxified to yield H2O2 by Mn-SOD in the mitochondrial matrix or by Cu/Zn-SOD in the cytosol and intermembrane space, and H2O2 can also be transformed to •OH in the presence of reduced transition metals [40]. Cu/Zn-SOD enzymes play a critical function in the first line of antioxidant defense [135].
Catalase (CAT): It is a tetrameric porphyrin-containing enzyme found mostly in the peroxisome that protects cells by converting H2O2 into H2O and O2 using either a Fe or Mn cofactor [137]. This mechanism prevents the formation of H2O2 and lowers the level of ROS; both are important mechanisms in the development of oxidative stress tolerance [135].
Glutathione Peroxidases (GPx): GPx is another intracellular enzyme that reduces ROS levels by conversion of H2O2 into H2O while oxidizing glutathione (GSH) to produce H2O and glutathione disulfide (GSSG) [138]. Several isoforms of GPx contain either five selenium cofactors or three noncysteine residues, which is important for enzyme activity. Most of these enzymes are found in the mitochondrial matrix, with a little quantity in the cytoplasm [139].
3.1.2. Antioxidant Non-Enzymes
Glutathione (GSH): GSH is a tripeptide composed of amino acids, i.e., glycine, cysteine, and glutamic acid, and is the most abundant endogenous water-soluble antioxidant. GSH can directly neutralize ROS and is an important factor in the xenobiotic metabolism [140]. To maintain an intracellular reducing environment and counteract excessive generation of ROS, GSH works with three groups of detoxification enzymes. These enzymes include glutathione peroxidase (GPx), glutathione reductase (GR), and glutathione oxidase [139].
α-Lipoic Acid (ALA): ALA is categorized as sulfur-containing molecules that catalyze the oxidative decarboxylation of α-keto acids, such as pyruvate and α-ketoglutarate. As a universal antioxidant, oxidized lipoic acid and its reduced counterpart, dihydrolipoic acid (DHLA), can quench free radicals in both lipid and aqueous environments [141].
Uric Acid: Uric acid is a hydrophilic antioxidant produced during purine nucleotide metabolism that accounts for about 60% of the total blood serum-free radical scavenging activity. Uric acid is an effective electron donor and scavenger of a variety of ROS, including •OH, O2•−, OONO−, HClO, and lipid peroxides. Complete scavenging of such species requires the participation of ascorbic acid and thiols in its cycle [142].
Coenzyme Q 10 (CoQ10): Coenzyme Q10 (CoQ10) or ubiquinol is another antioxidant enzyme cofactor involved in the mitochondrial ETC, which transfers electrons in complex I and complex II to complex III. CoQ10 is a lipid-soluble antioxidant present in all the cell membranes and inhibits lipid peroxidation [143]. In addition, other antioxidants, such as Vitamin E and C, require CoQ10 for their recycling and regeneration [144].
3.2. Exogenous Antioxidants
Dietary sources contain complex systems of multiple antioxidants that include vitamins (C, E, and A); carotenoids; and various polyphenols that the human body cannot synthesize. These antioxidants inhibit the initiation of the chain reactions or break the chain reactions by donating an electron to radicals, resulting in nonharmful species [4]. Furthermore, these exogenous antioxidants aid in the reinforcement and replenishment of the endogenous antioxidant, allowing the elimination of excess ROS/RNS [145].
Vitamins: Vitamin C (ascorbic acid) represents an efficient electron donor, converting free radicals to stable entities in the aqueous phase of the cytoplasm [146]. Tocopherols and tocotrienols are lipid-soluble forms of vitamin E, protecting the membrane lipids by inhibiting lipid peroxidation caused by oxidative and inflammatory reactions [147]. Vitamin A designates a family of unsaturated lipid-soluble organic compounds that include retinol, retinal, retinoic acid, retinyl palmitate, and many provitamin-A carotenoids, such as β-carotene [148]. Vitamin supplements are commonly used with an anticipation that they will protect cells and tissue from oxidative stress [149,150,151].
Carotenoids: Carotenoids are fat-soluble terpenoids containing conjugated trans double bonds. Carotenes (lycopene, β-carotene, or α-carotene) and xanthophylls (lutein, astaxanthin, fucoxanthin, capsanthin, zeaxanthin, and canthaxanthin) belong to the carotenoid family, widely present in red, orange, and yellow pigments in carrots; sweet potatoes; papaya; mangos; tomatoes; and oranges [152]. Carotenoids, acting as free radical scavengers and singlet oxygen quenchers, play a key role in inhibiting the oxidation of lipids [135]. In addition, these carotenoids inhibit apoptosis by preventing oxidative stress and display antioxidant and neuroprotective roles [153,154,155].
Polyphenols: Polyphenolic compounds are present in various fruits; vegetables; and beverages, such as grape juice, green tea, or coffee, and possess antioxidative, anti-inflammatory, and neuroprotective properties by scavenging free radicals [156]. Polyphenols have a wide range of aromatic structures, but the basic monomer in polyphenols is the phenolic ring. Depending on the strength of the phenolic ring into phenolic acids, they can be classified into phenolic acids, flavonoids, stiblins, phenolic alcohols, and lignans [157]. These polyphenols commonly include anthocyanins from berries, resveratrol found in grape skin or seeds, catechins from green tea, and curcumin isolated from the rhizome of the Indian spice turmeric Curcuma longa Linn [158]. Other most studied polyphenolic chemicals include chalcones, epigallocatechin gallates (EGCG), and quercetin [159]. These antioxidants detoxify various free radicals by scavenging or trapping them and by upregulating the activities of endogenous antioxidants [160]. These natural polyphenols also prevent oxidation of proteins, LPO, and show neuroprotective and neuroregenerative effects [133].
3.3. Synthetic Antioxidants
Synthetic antioxidants, modifications of natural antioxidants, or conjugates with other effective molecules have been prepared for better scavenging activity, bioavailability, and metabolic stability than natural antioxidants [161]. Synthetic antioxidants such as butylated hydroxyanisole (BHA), butylated hydroxytoluene (BHT), propyl gallate (PG), and tert-butyl hydroquinone (TBHQ) are widely used in the food industry to prevent lipid oxidation [162].
Recent research on synthetic antioxidant derivatives provides promising data against oxidative stress and multiple targets in neurodegenerative diseases. For example, synthetic compound 4-((5-(Tert-butyl)-3-chloro-2-hydroxy benzyl) amino)-2-hydroxybenzoic acid [163] and 1,3,4 oxadiazole compound A3 [164] showed significant antioxidative and neuroprotective effects. Synthesized docosahexaenoic acid (DHA)-acylated astaxanthin diesters (AST-DHA) showed substantially better effects than astaxanthin in reducing oxidative stress tau protein, enhanced learning and memory [165], and suppressing apoptosis of the dopaminergic neurons [166]. Similarly, the synthetic pyrazole derivative of curcumin (CNB-001) was demonstrated to suppress RNS generation with anti-inflammatory effect [167]. Synthetic derivatives of a natural phenolic compound such as caffeic acid phenethyl ester (CAPE) [168] or coumarin [169] also demonstrated to protect dopaminergic neurons by inhibiting p38 phosphorylation, increasing cell viability, and promoting antioxidant response.
Combination of novel synthetic pyrazole-containing compound 5-amino-1- phenyl-1H-pyrazole-4-carbonitrile (APPC) with lipoic acid, i.e., UPEI-800, showed synergistic neuroprotection both an in vitro hypoxia model and in vivo stroke model by reducing infarct volume [170]. A synthetic hybrid of antioxidants, i.e., coumarin and licochalcone A (Lico A), i.e., LM-031, has shown to inhibit Aβ aggregation in Aβ-GFP SH-SY5Y cells, scavenge ROS, promote neurite outgrowth, and activate the Nrf2-related antioxidant and antiapoptotic pathways [171]. Another hybrid compound (Dlx-23) developed by conjugating ALA and 3-n-butylphthalide (NBP), was shown to protect neuronal cell death, restore redox homeostasis, and synergistically prevent mitochondrial damage in a stroke model [172].
Synthetic nitrones are effective inhibitors of short-lived free radicals [173]. Due to their ability to react with free radicals to form a persistent nitroxide spin adduct; they can be used as an analytical tool for the detection and characterization of free radicals using Electron Paramagnetic Resonance (EPR) spectroscopy [174]. Synthetic nitrone derivatives showed antioxidative and neuroprotective effects in various neurodegenerative disease conditions [175,176].
Synthetic edaravone scavenges free •OH radicals and OONO− radicals, which are highly associated with neuronal damage/death in cerebrovascular disorders such as ischemic strokes and degenerative neurological disorders such as ALS [177]. It exerts neuroprotective and antioxidant effects and delays disease progression by limiting the extent of lipid peroxidation and cell membrane damage from oxidative stress [178].
In recent years, mitochondrial-targeted antioxidants have been successfully developed. For e.g., synthetic analogs of CoQ10, idebenone, and mitoquinone (MitoQ) demonstrated effective amelioration of mitochondrial ROS [179], DNA damage, neuroinflammation, and prevented neuronal degradation [180]. Idebenone is characterized by a shorter and less lipophilic tail than CoQ10, and MitoQ is composed of ubiquinone and triphenylphosphonium (TPP+) [181]. Plastoquinone derivatives, i.e., SkQ1 and SkQR1 molecules that contain an antioxidant moiety linked to a lipophilic cation, also demonstrated a neuroprotective effect [182]. Synthetic arylidenmalonate derivative 5-(3,4-dihydroxybenzylidene)-2,2-dimethyl-1,3-dioxane-4,6-dione (KM-34) also showed significant antioxidant property, mitoprotection and neuroprotection in vitro and in vivo models [183].
4. Preclinical Studies with Antioxidant Agents
4.1. Antioxidant-Based Therapy in Neurodegenerative Diseases
With a strong rationale that oxidative stress is a key component of neurodegenerative diseases, antioxidants of different types, either alone or in combination, natural and synthetic have been tested in neurodegenerative disease models. In general, in AD models, the treatment with antioxidants produced favorable outcomes. For e.g., the treatment with CoQ10 or lipoic acid increased the levels of ATP and SOD and reduced the levels of Apolipoprotein E (ApoE) and Aβ fragments [184]. The treatments also reduced the levels of phosphorylated tau and neuroinflammatory factors [185] and improved hippocampal synaptic plasticity [186]. Similarly, the treatment with carotenoids inhibited the markers of oxidative stress [137,187] and the AD marker proteins, improved memory loss, and reduced inflammation [188,189,190]. Polyphenols such as resveratrol [191], curcumin [192,193], and anthocyanin [121] have been shown to attenuate glutamate-induced excitotoxicity, increased antioxidant capacity and mitophagy [194], and rescue cell death in AD models [195,196,197]. The nutritious mushroom, hericium erinaceus is a source of exogenous antioxidants and has been shown to possess neuroprotective and anti-inflammatory properties [198]. In a sporadic AD model, hericium erinaceus treatment reduced behavioral abnormalities, hippocampus neuronal degeneration, and AD markers [199]. A few combination therapies such as ubiquinol and ascorbic acid [200], lycopene with vitamin E [201], CoQ10 and Omega-3 [202], and resveratrol and curcumin [203] reported to having a synergistic beneficial effect on reducing amyloid plaques and tau hyperphosphorylation in transgenic or sporadic models of AD. A synthetic derivative of CAPE termed FA-97 was developed by Wan et al. and has been shown to attenuate H2O2-induced apoptosis and suppress the levels of ROS, MDA, and protein carbonyl; and induce the cellular antioxidant levels in an in vitro study [204].
In PD models, supplements of vitamins E [205] and C [206,207] and CoQ10 [208] have been shown to restore corticostriatal synaptic plasticity, reduce dopaminergic cell death in the substantia nigra, microglial activation and astrogliosis, and improve behavioral parameters. ALA was shown to suppress oxidative stress, mitochondrial dysfunction, and glutamate-induced toxicity [209,210]. The treatment with other antioxidants, crocin [211] or fucoxanthin [212], was also shown to suppress autophagy and improve behavioral alterations, homeostasis, and mitochondrial enzyme function. In the pesticide-induced PD model, treatment with resveratrol [213] improved lifespan and behavioral deficits [214] via Nrf2 activation [215]. Further, the combinatorial treatment of quercetin and piperine (bioavailability enhancer) significantly improved behavioral abnormalities [216]. A recent study has reported that the treatments with synthetic chalcone derivate and 2-Hydroxy-40-methoxychalcone (AN07) reduced ROS level, stimulated Nrf2 pathways, increased GSH levels, and decreased inflammatory factors, thus favoring recovery [217]. In dopaminergic catecholaminergic (CATH.a) cells, a novel synthetic morpholine-containing chalcone (KMS99220) was shown to reduce oxidative stress effectively and protein aggregation, potentiate the Nrf2 mechanism and lower intracellular aggregation of α-synuclein [218]. In another example, Drummond et al. reported the antioxidant ability of a novel synthetic flavonoid, Proxison (7-decyl-3-hydroxy-2-(3,4,5-trihydroxyphenyl)-4-chromenone), and demonstrated enhanced cellular uptake, radical scavenging capabilities and neuroprotection against cell loss in a zebrafish model of dopaminergic neurodegeneration [219].
In ALS, mutation of the SOD1 genes reduces the antioxidant enzyme activity and hence is ineffective in lowering the ROS levels [220]. Curcumin has been shown to inhibit aggregation and fibrillation of SOD1 amyloid fibrils, lowering amyloidogenicity and neurotoxicity [221]. In a mouse model of ALS, treatment with anthocyanin-enriched extracts from strawberries was found to delay the disease onset, improve grip strength, reduce spinal motor neuron death, and preserve neuromuscular junctions (NMJs) [222]. Zhao et al. discovered that EGCG treatment stabilizes SOD1 conformation against misfolding and inhibits apo-SOD1 aggregation [223]. EGCG was found to have a substantial binding affinity for mutant SOD1, which reduces its toxic aggregate formation [224]. Phenolic compounds, quercitrin, quercetin 3-β-d-glucoside, and EGCG have been found to inhibit H2O2-induced misfolding and aggregation of A4V SOD1 [225]. Kaempferide and kaempferol are active ingredients of Brazilian green propolis that possess antioxidative properties and were shown to prevent SOD1 intracellular aggregates in a mutant SOD1-induced N2A cellular model [226]. Other studies also reported that the treatment with antioxidants (e.g., fisetin or protocatechuic acid) improves survival rate, attenuates motor impairment, reduces astrogliosis and microgliosis in the spinal cord, protects the spinal motor neurons from apoptosis, and regulates redox homeostasis by lowering the levels of both mutant and wild-type human SOD1 [227,228].
4.2. Antioxidant-Based Therapy in Neurological Injury
This review selected stroke, SCI, and PNI as examples where oxidative stress plays a key role in the early pathological and progressive degeneration following the acute event; this mechanism is also relevant to the acute TBI [123]. Treatments with several types of antioxidants, including α-lipoic acid (ALA) [229], α-tocopherol [230], vitamin C [231], crocin [232], resveratrol [233], and (−)-Epicatechin [144], have been shown to significantly reduce infarct volume, brain edema, oxidative damage, and apoptosis. In addition, the treatments protected the BBB integrity and promoted neurological recovery in stroke model studies. In other studies, pretreatment with natural free radical scavenger (e.g., ginkgo biloba extracts (Egb-761) [234] and astaxanthin [235]) has been shown to significantly ameliorate ischemic injury and reduce infarct volumes and brain edema, accompanied by alleviated oxidative stress, and upregulation of expression of brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF) mRNA.
In several studies, the efficacy of antioxidant treatment has been examined in animal models of acute SCI. They have been found to inhibit the expression of proapoptotic proteins (Bax and Caspase-3), increase the level of antiapoptotic protein (Bcl-2), reduce the level of MDA, and improve the activities of SOD and GSH (e.g., CoQ10) [236]. For example, the rats with SCI treated with vitamin E-enriched diet showed accelerated bladder recovery and improved locomotor function [237]. Treatment either with β-carotene or lycopene was also shown to reduce oxidative damage, mitochondrial dysfunction, cell apoptosis, and hind limb motor disturbances [238]. The treatment also inhibited inflammation by blocking the nuclear factor kappa B (NF-κB) pathway [239]. Antioxidants such as curcumin derivative, EGCG, or astaxanthin have been shown to reduce inflammation [240], promote regeneration, provide neuroprotection, and ultimately improve functional recovery [241,242]. Similarly, the treatment with resveratrol was shown to reduce the levels of inflammatory cytokines and inhibit cell death [243], improve motor function [244] via activation of the Sirtuin 1 (SIRT-1)/NF-κB signaling pathway [245], Beclin-1 and LC3-B, key proteins of autophagy [246], or the SIRT1/Adenosine 5′ monophosphate-activated protein kinase (AMPK) signaling pathway [247]. Quercetin treatment in SCI models was shown to reduce necroptosis of oligodendrocytes, which prevented axonal loss [248] and also suppressed macrophages/microglia polarization to proinflammatory M1 phenotype [249]. The combination treatment with ascorbic acid and taurine (nonproteogenic essential amino acid) showed synergistic protection against apoptotic, inflammatory, and oxidative stress markers in SCI-induced rats [250]. In PNI, compounds having antioxidative properties such as vitamins, carotenoids, enzymes, and proteins have been demonstrated to facilitate the process of nerve repair [127].
4.3. Clinical Trials with Antioxidants
Promising data from preclinical studies led clinical trials to determine the efficacy of antioxidants in different neurodegenerative diseases/injuries, primarily with few commonly used antioxidants such as curcumin, vitamin E, lipoic acid, and CoQ (Table 1 and Table 2).

Table 1.
Clinical trials of antioxidants in neurodegenerative diseases.
Table 2.
Clinical trials with antioxidants in neurological injury.
A phase II clinical trial study with oral dosing of curcumin in AD patients was shown to reduce cognitive deterioration but did not improve cognition [251]. Curcumin oral supplementation also demonstrated a slight slowdown in the disease progression in ALS patients [252]. Treatments with resveratrol [253,254] and EGCG [255] have been shown to attenuate Aβ1–40 and slow cognitive decline in AD patients, and in stroke patients, reduce the levels of matrix metalloproteinase-9 (MMP-9) and matrix metalloproteinase-2 (MMP-2). However, intranasal administration of GSH did not show the effect of the treatment on motor scores in PD patients [256,257]. High-dose treatment of CoQ10 in the idiopathic PD participants showed improved unified PD rating scale (UPDRS); however, it was indicated that the high-dose of CoQ10 (2400 mg/day) could increase the risk of oxidative damage in the long run [258]. In another clinical trial with CoQ10, although its dose was found safe and well-tolerated, it did not show any therapeutic benefits; hence, the study was terminated [259]. Similarly, a high dose of CoQ10 treatment showed a decrease in the ALS Functional Rating Scale-revised (ALSFRSr) score; however, the subsequent analyses revealed no significant differences compared to the placebo control [260]. Ginkgo biloba treatment in older patients with cognitive impairment also did not improve cognition compared to placebo [261]. Clinical trial on edaravone demonstrated a significant reduction in the ALSFRSr score in ALS patients compared to placebo group [262,263]. Furthermore, edaravone has been shown effective in recovery in stroke patients and reduce the MMP-9 levels [264,265]. In addition, the combination treatment, edaravone with (+)- borneol, a food additive, has been proven to be safe and well-tolerated in stroke patients [266] and is currently under a phase II clinical trial in patients suffering from intracerebral hemorrhage [267].
The combination treatments such as lipoic acid and omega-3 fatty acids, i.e., fish oil, given to AD patients were found to slow the cognitive and functional decline as compared to the placebo [268]. Similarly, vitamin E, the AD drug memantine (brand name: Namenda), or their combination was shown to slow down the clinical progression of AD. Interestingly vitamin E treatment resulted in a slower functional deterioration than the combination [269].
A combination treatment consisting of vitamin E, vitamin C, ALA, and CoQ did not show the effect on amyloid or tau pathology biomarkers in the cerebrospinal fluid (CSF). Furthermore, the treatment with CoQ did not improve oxidative stress or neurodegenerative indicators [270]. On the other hand, omega-3 fatty acids and vitamin E co-supplementation in PD patients resulted in a significant improvement in the UPDRS and favorable effect on total antioxidant capacity (TAC) and GSH levels compared to placebo but did not affect the oxidative stress indices or lipid profiles and inflammatory factors [271]. Daily treatment of the combination of ginkgo biloba (Egb761) and aspirin in stroke patients alleviated cognitive and neurological impairment after acute ischemic stroke without increasing the risk of vascular injury [276]. In SCI patients, dietary supplementation containing curcumin and omega-3, vegetation protein powder, antioxidant network capsule, and chlorella tablet were reported to reduce inflammatory mediators and improve depressive behavior [277]. Overall, the clinical trial results showed some trend towards a positive outcome, particularly the changes in the pathological markers and a few studies, improvements in functional outcome, thus indicating the potential of antioxidants to mitigate oxidative stress in humans. However, the results also highlighted the need to improve their therapeutic efficacy and make the clinical outcomes conclusive and reproducible, and importantly achieve functional recovery. To that end, in addition to developing more potent and target-specific antioxidants, drug delivery approaches have also been explored.
4.4. Drug Delivery Challenges
Despite promising results in preclinical models of neurodegenerative diseases, not all the clinical trial results were definitive. In general, antioxidant compounds were found to reduce clinical signs and symptoms only [252] but unable to stop the disease progression or reverse it [261]. Vitamins and flavonoids are still used but mostly as dietary supplements, which may act as prophylactic with long-term use. Edaravone (free radical scavenger) is the only Food and Drug Administration (FDA)-approved antioxidant treatment, and it is used to help people recover from stroke in Japan and is used to treat the early stages of ALS in the US and Japan, but it does not affect the disease progression in late-stage ALS, thus benefiting only 5% of ALS patients [278]. In Europe, the use of Radicava medication (active substance of edaravone) has been withdrawn from the marketing authorization, since the data did not show a positive benefit-risk balance [279]. There are various challenges associated with effective drug delivery of antioxidants that may be impeding their clinical translation.
- Low permeability to the CNS: The presence of a physiological barrier such as the BBB or spinal–blood barrier (SBB) restricts the accessibility of antioxidant compounds to the CNS and hence could not achieve a prolonged therapeutic dose to impart an antioxidant effect in chronic neurodegenerative diseases [280]. In certain pathological conditions, the BBB/SBB may be compromised due to inflammation or injury (e.g., stroke and spinal cord injury) but still may not be able to achieve the desired dose for a prolonged period due to transient and limited permeability of the BBB/SBB, giving a narrow time window for delivery of therapeutics [281].
- Low bioavailability: Most antioxidants are given orally, and they are insoluble or unstable in a gastric environment that could result in low bioavailability to provide high systemic levels for transport to the CNS at effective doses [282,283]. Antioxidant compounds that are administered via systemic routes have short half-lives [284], which could also limit their transport to the CNS.
- Low catalytic activity: High doses of antioxidant compounds are needed to detoxify the effect of free radicals, which could not be given to humans because of the dose-limiting toxicity [285]. Noncatalytic antioxidant becomes ineffective, once these molecules interact with free radicals [286], and hence, maintaining high antioxidant levels in the target tissue to counteract free radicals that are formed over a period of time in chronic conditions could be challenging.
- Toxicity: Due to toxicity concerns, human doses could have been significantly lower than those used in animal model studies. This could also constrain the duration of treatment necessary to see the beneficial outcome in clinical trials [287].
- Oxidative stress target and other factors: Although oxidative stress is considered as the driving force behind neurodegenerative diseases, there could be other cofounding pathological factors in humans that may not have been targeted solely by antioxidants [288,289]. In addition, the question raised is also how close animal models are to human pathology [290].
5. Antioxidant-Based Nanotherapy
To overcome the limitations of natural and synthetic antioxidants, significant efforts have been made to improve their efficacy using drug delivery approaches. These include exploring nanocarriers of different polymeric materials, conjugates, and complexes [291,292] to improve their stability, half-lives, transport to the CNS, and sustained their effect in the target tissue (Figure 5).
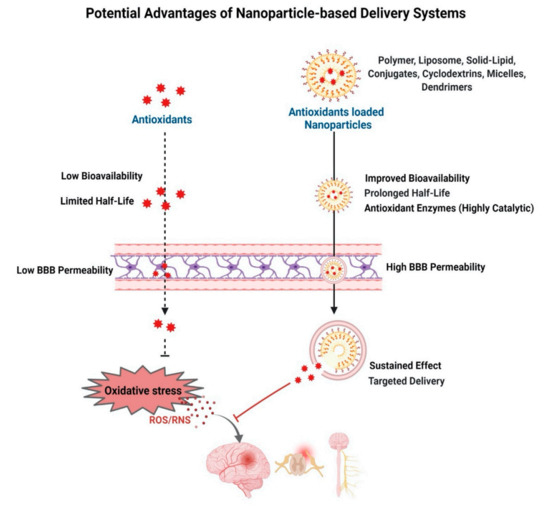
Figure 5.
Antioxidant-based nanotherapy. Schematic depicting advantages of delivery of antioxidant-loaded nanoparticles to improve half-life of antioxidants and their ability to cross the BBB, improve bioavailability, and sustain the effect, thus effectively neutralizing oxidative stress in neurodegenerative diseases.
Due to its broad pharmacological effects, including anti-inflammatory and antioxidant properties, curcumin has been widely investigated in clinical studies. To overcome its low water solubility, poor bioavailability, and rapid metabolism, curcumin is formulated as nanocurcumin using different nanocarriers, such as liposomes, polymers, conjugates, cyclodextrins, micelles, dendrimers, and nanoparticles [293]. Transferrin-conjugated poly (lactic co-glycolic acid) (PLGA) nanoparticles have been demonstrated to improve the bioavailability of curcumin to the brain and reduce Aβ deposition and tau hyperphosphorylation in the AD model [294]. Similarly, different formulations of nanoparticles have been shown to inhibit aggregation of Aβ and reduce depressive-like behavior and oxidative stress in AD models [295,296]. Intra-arterial administration of resveratrol (RES)-encapsulated nanoparticle (RES-NP) in a rat transient middle cerebral artery occlusion (t-MCAO) enhanced the resveratrol bioavailability and its brain-penetration, resulting in reduced infarct volume, and attenuated oxidative stress [297], brain edema, and neuronal apoptosis. The treatment also contributed to neurogenesis, leading to improved neurological recovery [298]. In a cerebral palsy rabbit model, intravenous treatment of dendrimer-based N-acetyl-l-cysteine (NAC) [299], a glutathione precursor with antioxidant and anti-inflammatory properties [300], reduced neuroinflammation and neurological injury, and improved motor function. In general, formulating antioxidants in nanocarriers has enhanced their efficacy due to better stability and/or improved transport to the CNS than free antioxidants [301,302,303,304,305,306,307,308,309,310,311,312,313,314,315,316,317,318,319,320,321,322,323,324,325,326,327]. Nanocurcumin has been evaluated as an add-on therapy to Riluzole in a pilot randomized clinical trial for safety and efficacy in ALS [272] and AD patients as dietary supplements [274]. In another study, solid–lipid curcumin showed significantly improved cognition and mood in healthy older population [273].
Edaravone-loaded ceria nanoparticles have demonstrated to cross the BBB via receptor-mediated transcytosis and protect the BBB [328]. In addition to the antioxidant property of ceria nanoparticles, edaravone provided its effect against oxidative stress in a stroke model [328]. Jin et al. demonstrated that the treatment with edaravone-encapsulated agonistic micelles caused rapid infarct volume reduction, prolonged survival, improved axonal remodeling, and reduced behavioral deficits than free edaravone-treated animals [329]. Wang et al. reviewed nanotechnology-based strategies for the treatment of ALS, including antioxidant agents [330]. Nanoparticle-loaded edaravone has been tested on the postoperative effects in patients with cerebral hemorrhage. The nanoparticle-loaded edaravone showed reduced edema as compared to free edaravone treated group, significantly improved neurological function, and reduced the production and release of interleukin and tumor necrosis factor, which was considered beneficial to protect healthy brain tissue and other organs, and conducive to the recovery and healing [275].
Antioxidant Enzymes
When they interact with free radicals, natural or synthetic antioxidants become inactive [331]. To continue to neutralize free radicals formed over a prolonged period, in chronic disease conditions, therapeutic levels of these antioxidants need to be maintained, which could be challenging, as repeated and high dosing cause dose-limiting toxicity in humans [331]. The main advantage of antioxidant enzymes is their catalytic mode of action [6]; hence, they can effectively neutralize free radicals at low doses. However, due to their short half-lives (5–11 min) [332], exogenously delivered antioxidant enzymes are ineffective in combating oxidative stress. Modifications such as PEGlylation and lecithinization improve their stability in the circulation [333] and fusion with cell membrane-penetrating peptides like a transactivator of transcription peptide or tetanus toxin fragment increases their ability to cross the BBB [334]. However, there are limitations to these modifications. Although PEGylated SOD (PEG-SOD) increases the enzyme’s stability in the circulation from 6 min to 36 h, PEG limits the permeability of the conjugated SOD across cerebral cell membranes [335]. Similarly, a chemical reaction involved in the fusion of different cell-penetrating or cell-specific peptides could cause denaturation and loss of enzyme activity [336]. In addition, the newly formulated hybrid enzyme could trigger immune-mediated anaphylactic responses to patients [337]. Intravenous delivery of SOD loaded into liposomes has shown to partially inhibit the infarct volume, but instability of liposomes in vivo (half-life ~4.2 h) limits the duration of SOD activity and, hence, its efficacy [338,339].
The recent effort includes formulations of antioxidant enzymes, SOD1, and catalase by electrostatic coupling of enzymes with cationic block copolymers called nanozymes [340]. In mice, this formulation demonstrated increased stability of enzymes in both blood and the brain and showed increased accumulation in the brain tissues than enzyme alone treated animals [340]. In a rat MCAO model, nanozymes reduced I/R-induced tissue injury and improved the sensorimotor functions [341]. In a moderate SCI rat model, treatment with nanozymes showed a recovery of locomotor functions, reduction of swelling, and post-traumatic cysts in the spinal cords of the treated animals [342]. Muzykantov’s group reviewed different nanocarriers to deliver antioxidant enzymes for vascular targeting in oxidative stress conditions associated with cardiovascular, pulmonary, and nervous systems [343].
Our research group has been investigating the efficacy of antioxidant enzymes encapsulated in PLGA-based sustained release nanoparticles. The neuroprotective efficacy of the SOD-encapsulated nanoparticles (nano-SOD) was initially demonstrated in the H2O2-induced oxidative stress model in human neuronal cells and, subsequently, with the CAT-encapsulated nanoparticles (nano-CAT) in human astrocytes [344,345]. In the MCAO model in rats, intracarotid administration of nano-SOD following 1 h of ischemia inhibited reperfusion injury. The treatment demonstrated improved neurological recovery and survival compared to controls (saline or SOD solution). There was evidence of neuronal recovery and regeneration with time in the above study [346]. The follow-up study in a thromboembolic rat stroke model, where tissue plasminogen activator (t-PA) was administered first for clot lysis followed by nano-SOD/CAT, both via the carotid artery, demonstrated the protective effect of the treatment. Significantly, the t-PA + nano-SOD combination treatment stimulated the migration of stem/progenitor cells from the subventricular zone and circulation, promoting neurogenesis. In contrast, this process was inhibited in the animals which received t-PA only treatment [347]. The above sequential treatment also inhibited edema formation, suggesting protection of the BBB from reperfusion injury [347]. In a separate study, we demonstrated aggravation of the BBB permeability when t-PA alone was administered via the carotid artery in the same thromboembolic rat stroke model [348]. Thus, the delivery of antioxidant enzyme nanoparticles in the above sequential treatment study protected the BBB from reperfusion injury and, also, from the effect of t-PA.
In our recent study, we demonstrated that intravenous administration of nano-SOD/CAT, 6-hr following injury in a rat severe contusion model of SCI, partially attenuated mitochondrial dysfunction, reduced ROS levels, and the expression of apoptotic factors (Figure 6C). Further, the isolated mitochondria from the spinal cords of the treated animals demonstrated reduced ROS activity and higher ATP production capacity than those isolated from untreated control animals (Figure 6D). The overall effect of the treatment was found to be the protection of the injured spinal cord from cell apoptosis and further degeneration [349]. The protective effect of the treatment was seen across the entire spinal cord as there was reduced expression of apoptotic factors, suggesting that the treatment significantly attenuated the progression of secondary injury. Neuroprotective efficacy of the nano-SOD/CAT in the above SCI model study is attributed to the protection of the encapsulated enzymes and sustained antioxidant effect at the lesion site [350] (Figure 6). Since nano-SOD/CAT is formulated with PLGA, an FDA-approved polymer, its translational potential is high.
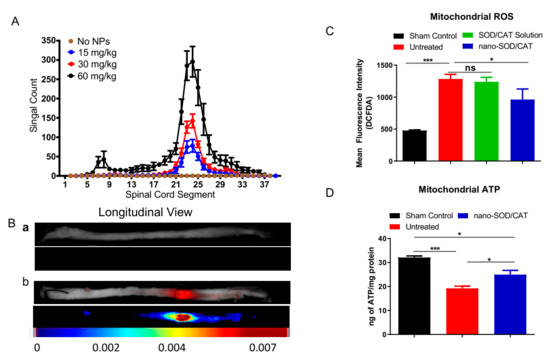
Figure 6.
Antioxidant enzyme-based nanotherapy for spinal cord injury: Localization of nanoparticles at the lesion site following intravenous administration. Nanoparticles were injected 6 h post-injury, and spinal cords were analyzed for localization of the nanoparticles. Nanoparticles contained a near-infrared dye, and the spinal cords were analyzed 24 h after the injury using the Maestro Optical Imaging System site. Reproduced with permission from [350], copyright 2019 Elsevier. (A) Dose-dependent localization of nanoparticles at the lesion site. (B) Images of the spinal cord taken with Maestro Optical Imaging (Ba) Normal spinal cord without injury and nanoparticles. (Bb) Injured spinal cord from the animals that received dye-loaded nanoparticles intravenously. Efficacy of nano-SOD/CAT treatment (C)-treated animals show reduced mitochondrial ROS levels. (D) Mitochondrial isolated from the spinal cord of the treated animals show more ATP production capacity than those isolated from the spinal cords of untreated animals. * p < 0.05; *** p < 0.001 Reproduced with permission from [350], copyright 2019 Elsevier.
6. Concluding Remarks/Future Perspective
Effective treatment for neurodegenerative diseases is a clinically unmet need. Substantial evidence supports the hypothesis that oxidative stress plays a key role in disease progression; hence, antioxidant treatment could provide a potential solution. However, several challenges, including inadequate dosing, low bioavailability, limited transport to the CNS and transient retention, and low antioxidant capacity to completely detoxify the effect of free radicals, could have limited their translation to clinical practice. In this regard, nanoparticle-based drug delivery systems could address some of the above issues. Antioxidant enzymes hold promise due to their high catalytic activities; therefore, much work has been done in recent years to develop the nanotherapy-based approach to delivering these antioxidant enzymes. Since oxidative stress is a common pathophysiological process in multiple diseases, an effective antioxidant system could have broad therapeutic applicability in many clinical settings.
Author Contributions
A.A., S.S.A. and S.M. contributed to writing, with A.A. taking the lead in compiling information, preparing the draft and figures, and editing. S.S.A. and Y.K. were involved in the studies with antioxidant enzyme formulations described in the review. B.K.K. reviewed and edited the manuscript. V.L. provided guidance and contributed to writing and editing. All authors have read and agreed to the published version of the manuscript.
Funding
The studies described from our laboratory were supported by the National Institute of Neurological Disorders and Stroke of the National Institutes of Health under grants R01NS092033, 1R01NS113680, and 1R01NS070896 and the Department of Defense through the Spinal Cord Injury Research Program under award no. W81XWH-16-1-0786. The opinions, interpretations, conclusions, and recommendations are those of the authors and not necessarily endorsed by the Department of Defense.
Conflicts of Interest
V.L. is a co-inventor on US and EU patents/patent applications related to antioxidant nanoparticles for treating spinal cord injury and stroke. AxoNeural Therapeutics, Inc. is a spinout company of Cleveland Clinic Venture, developing a treatment for neuronal diseases. VL is the founder and scientific advisor of AxoNeural Therapeutics. This conflict of interest is managed by the Conflict of Interest Committee of Cleveland Clinic according to its conflict of interest policies.
Abbreviations
| ALA | α-Lipoic acid |
| ALSFRSr | Amyotrophic Lateral Sclerosis Functional Rating Scale Revised |
| APP | Amyloid Precursor Protein |
| ARE | Antioxidant Response Element |
| BBB | Blood-Brain Barrier |
| CAT | Catalase |
| CNS | Central Nervous System |
| CoQ10 | Coenzyme Q10 |
| EGCG | Epigallocatechin Gallates |
| ETC | Electron Transport Chain |
| GSH | Glutathione |
| GST | Glutathione S-Transferase |
| GPx | Glutathione Peroxidases |
| GR | Glutathione Reductase |
| HO-1 | Heme Oxygenase 1 |
| 4-HNE | 4-Hydroxynonenal |
| KEAP1 | Kelch-like ECH-Associated Protein 1 |
| LDL | Low-Density Lipoproteins |
| LPO | Lipid Peroxidation |
| MCAO | Middle Cerebral Artery Occlusion |
| MDA | Malondialdehyde |
| MMP | Matrix Metalloproteinases |
| NF-κB | Nuclear Factor Kappa B |
| NFTs | Neurofibrillary Tangles |
| Nrf2 | Nuclear Factor Erythroid 2-Related Factor 2 |
| NQO1 | NADPH Quinine Oxidoreductase 1 |
| PLGA | Poly(Lactic-co-Glycolic Acid) |
| PNS | Peripheral Nervous System |
| RA | Retinoic Acid |
| ROS | Reactive Oxygen Species |
| RNS | Reactive Nitrogen Species |
| SIRT-1 | Sirtuin 1 |
| SOD | Superoxide Dismutase |
| TAC | Total Antioxidant Capacity |
| UPDRS | Unified Parkinson’s Disease Rating Scale |
References
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Lu, Y.; Saredy, J.; Wang, X.; Drummer Iv, C.; Shao, Y.; Saaoud, F.; Xu, K.; Liu, M.; Yang, W.Y.; et al. ROS systems are a new integrated network for sensing homeostasis and alarming stresses in organelle metabolic processes. Redox Biol. 2020, 37, 101696. [Google Scholar] [CrossRef] [PubMed]
- Collin, F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Man, A.W.C.; Li, H.; Xia, N. Impact of Lifestyles (Diet and Exercise) on Vascular Health: Oxidative Stress and Endothelial Function. Oxid. Med. Cell Longev. 2020, 2020, 1496462. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha-Oliveira, T.; Montezinho, L.; Mendes, C.; Firuzi, O.; Saso, L.; Oliveira, P.J.; Silva, F.S.G. Oxidative Stress in Amyotrophic Lateral Sclerosis: Pathophysiology and Opportunities for Pharmacological Intervention. Oxid. Med. Cell Longev. 2020, 2020, 5021694. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Journey through the Diagnosis of Dementia—World Alzheimer Report 2021. Available online: https://www.alzint.org/resource/world-alzheimer-report-2021/ (accessed on 14 January 2022).
- Anjum, A.; Yazid, M.D.; Fauzi Daud, M.; Idris, J.; Ng, A.M.H.; Selvi Naicker, A.; Ismail, O.H.R.; Athi Kumar, R.K.; Lokanathan, Y. Spinal Cord Injury: Pathophysiology, Multimolecular Interactions, and Underlying Recovery Mechanisms. Int. J. Mol. Sci. 2020, 21, 7533. [Google Scholar] [CrossRef]
- von Arnim, C.A.; Herbolsheimer, F.; Nikolaus, T.; Peter, R.; Biesalski, H.K.; Ludolph, A.C.; Riepe, M.; Nagel, G.; Acti, F.E.U.S.G. Dietary antioxidants and dementia in a population-based case-control study among older people in South Germany. J. Alzheimers Dis. 2012, 31, 717–724. [Google Scholar] [CrossRef] [Green Version]
- Urano, S.; Asai, Y.; Makabe, S.; Matsuo, M.; Izumiyama, N.; Ohtsubo, K.; Endo, T. Oxidative injury of synapse and alteration of antioxidative defense systems in rats, and its prevention by vitamin E. Eur. J. Biochem. 1997, 245, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Magistretti, P.J.; Allaman, I. A cellular perspective on brain energy metabolism and functional imaging. Neuron 2015, 86, 883–901. [Google Scholar] [CrossRef] [Green Version]
- Siraki, A.G.; O’Brien, P.J. Prooxidant activity of free radicals derived from phenol-containing neurotransmitters. Toxicology 2002, 177, 81–90. [Google Scholar] [CrossRef]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Popa-Wagner, A.; Mitran, S.; Sivanesan, S.; Chang, E.; Buga, A.M. ROS and brain diseases: The good, the bad, and the ugly. Oxid. Med. Cell Longev. 2013, 2013, 963520. [Google Scholar] [CrossRef]
- Selivanov, V.A.; Votyakova, T.V.; Pivtoraiko, V.N.; Zeak, J.; Sukhomlin, T.; Trucco, M.; Roca, J.; Cascante, M. Reactive oxygen species production by forward and reverse electron fluxes in the mitochondrial respiratory chain. PLoS Comput. Biol. 2011, 7, e1001115. [Google Scholar] [CrossRef] [Green Version]
- Mailloux, R.J.; McBride, S.L.; Harper, M.E. Unearthing the secrets of mitochondrial ROS and glutathione in bioenergetics. Trends Biochem. Sci. 2013, 38, 592–602. [Google Scholar] [CrossRef]
- Walker, C.L.; Pomatto, L.C.D.; Tripathi, D.N.; Davies, K.J.A. Redox Regulation of Homeostasis and Proteostasis in Peroxisomes. Physiol. Rev. 2018, 98, 89–115. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Hawkins, C.L. The Role of Myeloperoxidase in Biomolecule Modification, Chronic Inflammation, and Disease. Antioxid. Redox Signal. 2020, 32, 957–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballance, W.C.; Qin, E.C.; Chung, H.J.; Gillette, M.U.; Kong, H. Reactive oxygen species-responsive drug delivery systems for the treatment of neurodegenerative diseases. Biomaterials 2019, 217, 119292. [Google Scholar] [CrossRef] [PubMed]
- Veith, A.; Moorthy, B. Role of Cytochrome P450s in the Generation and Metabolism of Reactive Oxygen Species. Curr. Opin. Toxicol. 2018, 7, 44–51. [Google Scholar] [CrossRef]
- Araujo, J.E.; Jorge, S.; Santos, H.M.; Chiechi, A.; Galstyan, A.; Lodeiro, C.; Diniz, M.; Kleinman, M.T.; Ljubimova, J.Y.; Capelo, J.L. Proteomic changes driven by urban pollution suggest particulate matter as a deregulator of energy metabolism, mitochondrial activity, and oxidative pathways in the rat brain. Sci. Total Environ. 2019, 687, 839–848. [Google Scholar] [CrossRef]
- Calderon-Garciduenas, L.; Leray, E.; Heydarpour, P.; Torres-Jardon, R.; Reis, J. Air pollution, a rising environmental risk factor for cognition, neuroinflammation and neurodegeneration: The clinical impact on children and beyond. Rev. Neurol. 2016, 172, 69–80. [Google Scholar] [CrossRef]
- Rai, Y.; Anita; Kumari, N.; Singh, S.; Kalra, N.; Soni, R.; Bhatt, A.N. Mild mitochondrial uncoupling protects from ionizing radiation induced cell death by attenuating oxidative stress and mitochondrial damage. Biochim. Biophys. Acta Bioenerg. 2021, 1862, 148325. [Google Scholar] [CrossRef]
- Mullenders, L.H.F. Solar UV damage to cellular DNA: From mechanisms to biological effects. Photochem. Photobiol. Sci. 2018, 17, 1842–1852. [Google Scholar] [CrossRef]
- Akakin, D.; Tok, O.E.; Anil, D.; Akakin, A.; Sirvanci, S.; Sener, G.; Ercan, F. Electromagnetic Waves from Mobile Phones may Affect Rat Brain During Development. Turk. Neurosurg. 2021, 31, 412–421. [Google Scholar] [CrossRef]
- Choi, S.; Krishnan, J.; Ruckmani, K. Cigarette smoke and related risk factors in neurological disorders: An update. Biomed. Pharmacother. 2017, 85, 79–86. [Google Scholar] [CrossRef]
- Naha, N.; Gandhi, D.N.; Gautam, A.K.; Prakash, J.R. Nicotine and cigarette smoke modulate Nrf2-BDNF-dopaminergic signal and neurobehavioral disorders in adult rat cerebral cortex. Hum. Exp. Toxicol. 2018, 37, 540–556. [Google Scholar] [CrossRef]
- Rodriguez-Martinez, E.; Nava-Ruiz, C.; Escamilla-Chimal, E.; Borgonio-Perez, G.; Rivas-Arancibia, S. The Effect of Chronic Ozone Exposure on the Activation of Endoplasmic Reticulum Stress and Apoptosis in Rat Hippocampus. Front. Aging Neurosci. 2016, 8, 245. [Google Scholar] [CrossRef]
- Gargouri, B.; Bhatia, H.S.; Bouchard, M.; Fiebich, B.L.; Fetoui, H. Inflammatory and oxidative mechanisms potentiate bifenthrin-induced neurological alterations and anxiety-like behavior in adult rats. Toxicol. Lett. 2018, 294, 73–86. [Google Scholar] [CrossRef]
- Lahouel, A.; Kebieche, M.; Lakroun, Z.; Rouabhi, R.; Fetoui, H.; Chtourou, Y.; Djamila, Z.; Soulimani, R. Neurobehavioral deficits and brain oxidative stress induced by chronic low dose exposure of persistent organic pollutants mixture in adult female rat. Environ. Sci. Pollut. Res. Int. 2016, 23, 19030–19040. [Google Scholar] [CrossRef]
- Cruces-Sande, A.; Rodriguez-Perez, A.I.; Herbello-Hermelo, P.; Bermejo-Barrera, P.; Mendez-Alvarez, E.; Labandeira-Garcia, J.L.; Soto-Otero, R. Copper Increases Brain Oxidative Stress and Enhances the Ability of 6-Hydroxydopamine to Cause Dopaminergic Degeneration in a Rat Model of Parkinson’s Disease. Mol. Neurobiol. 2019, 56, 2845–2854. [Google Scholar] [CrossRef]
- Yauger, Y.J.; Bermudez, S.; Moritz, K.E.; Glaser, E.; Stoica, B.; Byrnes, K.R. Iron accentuated reactive oxygen species release by NADPH oxidase in activated microglia contributes to oxidative stress in vitro. J. Neuroinflamm. 2019, 16, 41. [Google Scholar] [CrossRef]
- Langley, M.R.; Ghaisas, S.; Ay, M.; Luo, J.; Palanisamy, B.N.; Jin, H.; Anantharam, V.; Kanthasamy, A.; Kanthasamy, A.G. Manganese exposure exacerbates progressive motor deficits and neurodegeneration in the MitoPark mouse model of Parkinson’s disease: Relevance to gene and environment interactions in metal neurotoxicity. Neurotoxicology 2018, 64, 240–255. [Google Scholar] [CrossRef]
- Ashok, A.; Rai, N.K.; Tripathi, S.; Bandyopadhyay, S. Exposure to As-, Cd-, and Pb-mixture induces Abeta, amyloidogenic APP processing and cognitive impairments via oxidative stress-dependent neuroinflammation in young rats. Toxicol. Sci 2015, 143, 64–80. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Perez de la Lastra, J.M.; Plou, F.J.; Perez-Lebena, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Damiano, M.; Starkov, A.A.; Petri, S.; Kipiani, K.; Kiaei, M.; Mattiazzi, M.; Flint Beal, M.; Manfredi, G. Neural mitochondrial Ca2+ capacity impairment precedes the onset of motor symptoms in G93A Cu/Zn-superoxide dismutase mutant mice. J. Neurochem. 2006, 96, 1349–1361. [Google Scholar] [CrossRef]
- Singh, B.K.; Tripathi, M.; Pandey, P.K.; Kakkar, P. Alteration in mitochondrial thiol enhances calcium ion dependent membrane permeability transition and dysfunction in vitro: A cross-talk between mtThiol, Ca(2+), and ROS. Mol. Cell Biochem. 2011, 357, 373–385. [Google Scholar] [CrossRef]
- Chiang, S.C.; Meagher, M.; Kassouf, N.; Hafezparast, M.; McKinnon, P.J.; Haywood, R.; El-Khamisy, S.F. Mitochondrial protein-linked DNA breaks perturb mitochondrial gene transcription and trigger free radical-induced DNA damage. Sci. Adv. 2017, 3, e1602506. [Google Scholar] [CrossRef] [Green Version]
- Eratne, D.; Loi, S.M.; Farrand, S.; Kelso, W.; Velakoulis, D.; Looi, J.C. Alzheimer’s disease: Clinical update on epidemiology, pathophysiology and diagnosis. Australas. Psychiatry 2018, 26, 347–357. [Google Scholar] [CrossRef]
- DeTure, M.A.; Dickson, D.W. The neuropathological diagnosis of Alzheimer’s disease. Mol. Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Markesbery, W.R. Oxidative stress hypothesis in Alzheimer’s disease. Free Radic. Biol. Med. 1997, 23, 134–147. [Google Scholar] [CrossRef]
- Sharma, C.; Kim, S.R. Linking Oxidative Stress and Proteinopathy in Alzheimer’s Disease. Antioxidants 2021, 10, 1231. [Google Scholar] [CrossRef]
- Harman, D. Free radical theory of aging. Mutat. Res. 1992, 275, 257–266. [Google Scholar] [CrossRef]
- Adav, S.S.; Park, J.E.; Sze, S.K. Quantitative profiling brain proteomes revealed mitochondrial dysfunction in Alzheimer’s disease. Mol. Brain 2019, 12, 8. [Google Scholar] [CrossRef]
- Kim, S.H.; Vlkolinsky, R.; Cairns, N.; Lubec, G. Decreased levels of complex III core protein 1 and complex V beta chain in brains from patients with Alzheimer’s disease and Down syndrome. Cell Mol. Life Sci. 2000, 57, 1810–1816. [Google Scholar] [CrossRef]
- Kim, S.H.; Vlkolinsky, R.; Cairns, N.; Fountoulakis, M.; Lubec, G. The reduction of NADH ubiquinone oxidoreductase 24- and 75-kDa subunits in brains of patients with Down syndrome and Alzheimer’s disease. Life Sci. 2001, 68, 2741–2750. [Google Scholar] [CrossRef]
- Liang, W.S.; Reiman, E.M.; Valla, J.; Dunckley, T.; Beach, T.G.; Grover, A.; Niedzielko, T.L.; Schneider, L.E.; Mastroeni, D.; Caselli, R.; et al. Alzheimer’s disease is associated with reduced expression of energy metabolism genes in posterior cingulate neurons. Proc. Natl. Acad. Sci. USA 2008, 105, 4441–4446. [Google Scholar] [CrossRef] [Green Version]
- Gauba, E.; Chen, H.; Guo, L.; Du, H. Cyclophilin D deficiency attenuates mitochondrial F1Fo ATP synthase dysfunction via OSCP in Alzheimer’s disease. Neurobiol. Dis. 2019, 121, 138–147. [Google Scholar] [CrossRef]
- Beck, S.J.; Guo, L.; Phensy, A.; Tian, J.; Wang, L.; Tandon, N.; Gauba, E.; Lu, L.; Pascual, J.M.; Kroener, S.; et al. Deregulation of mitochondrial F1FO-ATP synthase via OSCP in Alzheimer’s disease. Nat. Commun. 2016, 7, 11483. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.; Pan, Y.; Kao, S.Y.; Li, C.; Kohane, I.; Chan, J.; Yankner, B.A. Gene regulation and DNA damage in the ageing human brain. Nature 2004, 429, 883–891. [Google Scholar] [CrossRef]
- Reed, T.; Perluigi, M.; Sultana, R.; Pierce, W.M.; Klein, J.B.; Turner, D.M.; Coccia, R.; Markesbery, W.R.; Butterfield, D.A. Redox proteomic identification of 4-hydroxy-2-nonenal-modified brain proteins in amnestic mild cognitive impairment: Insight into the role of lipid peroxidation in the progression and pathogenesis of Alzheimer’s disease. Neurobiol. Dis. 2008, 30, 107–120. [Google Scholar] [CrossRef]
- Esposito, L.; Raber, J.; Kekonius, L.; Yan, F.; Yu, G.Q.; Bien-Ly, N.; Puolivali, J.; Scearce-Levie, K.; Masliah, E.; Mucke, L. Reduction in mitochondrial superoxide dismutase modulates Alzheimer’s disease-like pathology and accelerates the onset of behavioral changes in human amyloid precursor protein transgenic mice. J. Neurosci. 2006, 26, 5167–5179. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P.; Fu, W.; Waeg, G.; Uchida, K. 4-Hydroxynonenal, a product of lipid peroxidation, inhibits dephosphorylation of the microtubule-associated protein tau. Neuroreport 1997, 8, 2275–2281. [Google Scholar] [CrossRef] [PubMed]
- Misrani, A.; Tabassum, S.; Yang, L. Mitochondrial Dysfunction and Oxidative Stress in Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 617588. [Google Scholar] [CrossRef] [PubMed]
- Patten, D.A.; Germain, M.; Kelly, M.A.; Slack, R.S. Reactive oxygen species: Stuck in the middle of neurodegeneration. J. Alzheimers Dis. 2010, 20 (Suppl. 2), S357–S367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, D.L.; Thomas, C.; Rodriguez, C.; Simberkoff, K.; Tsai, J.S.; Strafaci, J.A.; Freedman, M.L. Increased peroxidation and reduced antioxidant enzyme activity in Alzheimer’s disease. Exp. Neurol. 1998, 150, 40–44. [Google Scholar] [CrossRef]
- Chen, L.; Na, R.; Gu, M.; Richardson, A.; Ran, Q. Lipid peroxidation up-regulates BACE1 expression in vivo: A possible early event of amyloidogenesis in Alzheimer’s disease. J. Neurochem. 2008, 107, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef]
- Simon, D.K.; Tanner, C.M.; Brundin, P. Parkinson Disease Epidemiology, Pathology, Genetics, and Pathophysiology. Clin. Geriatr. Med. 2020, 36, 1–12. [Google Scholar] [CrossRef]
- Gomez-Benito, M.; Granado, N.; Garcia-Sanz, P.; Michel, A.; Dumoulin, M.; Moratalla, R. Modeling Parkinson’s Disease With the Alpha-Synuclein Protein. Front. Pharmacol. 2020, 11, 356. [Google Scholar] [CrossRef]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in Parkinson’s disease. J. Parkinsons Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [Green Version]
- Puspita, L.; Chung, S.Y.; Shim, J.W. Oxidative stress and cellular pathologies in Parkinson’s disease. Mol. Brain 2017, 10, 53. [Google Scholar] [CrossRef] [Green Version]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative stress in the aging substantia nigra and the etiology of Parkinson’s disease. Aging Cell 2019, 18, e13031. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.; Zhang, X.; Meng, X.; Lu, L.; Mai, D.; Qu, S. Simvastatin Inhibits Activation of NADPH Oxidase/p38 MAPK Pathway and Enhances Expression of Antioxidant Protein in Parkinson Disease Models. Front. Mol. Neurosci. 2018, 11, 165. [Google Scholar] [CrossRef] [Green Version]
- Park, H.R.; Yang, E.J. Oxidative Stress as a Therapeutic Target in Amyotrophic Lateral Sclerosis: Opportunities and Limitations. Diagnostics 2021, 11, 1546. [Google Scholar] [CrossRef]
- Petrov, D.; Daura, X.; Zagrovic, B. Effect of Oxidative Damage on the Stability and Dimerization of Superoxide Dismutase 1. Biophys. J. 2016, 110, 1499–1509. [Google Scholar] [CrossRef] [Green Version]
- Richardson, K.; Allen, S.P.; Mortiboys, H.; Grierson, A.J.; Wharton, S.B.; Ince, P.G.; Shaw, P.J.; Heath, P.R. The effect of SOD1 mutation on cellular bioenergetic profile and viability in response to oxidative stress and influence of mutation-type. PLoS ONE 2013, 8, e68256. [Google Scholar] [CrossRef]
- Pansarasa, O.; Bordoni, M.; Diamanti, L.; Sproviero, D.; Gagliardi, S.; Cereda, C. SOD1 in Amyotrophic Lateral Sclerosis: “Ambivalent” Behavior Connected to the Disease. Int. J. Mol. Sci. 2018, 19, 1345. [Google Scholar] [CrossRef] [Green Version]
- Huai, J.; Zhang, Z. Structural Properties and Interaction Partners of Familial ALS-Associated SOD1 Mutants. Front. Neurol. 2019, 10, 527. [Google Scholar] [CrossRef] [Green Version]
- Obrador, E.; Salvador, R.; Lopez-Blanch, R.; Jihad-Jebbar, A.; Valles, S.L.; Estrela, J.M. Oxidative Stress, Neuroinflammation and Mitochondria in the Pathophysiology of Amyotrophic Lateral Sclerosis. Antioxidants 2020, 9, 901. [Google Scholar] [CrossRef]
- Blasco, H.; Garcon, G.; Patin, F.; Veyrat-Durebex, C.; Boyer, J.; Devos, D.; Vourc’h, P.; Andres, C.R.; Corcia, P. Panel of Oxidative Stress and Inflammatory Biomarkers in ALS: A Pilot Study. Can. J. Neurol. Sci. 2017, 44, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Bellezza, I.; Grottelli, S.; Costanzi, E.; Scarpelli, P.; Pigna, E.; Morozzi, G.; Mezzasoma, L.; Peirce, M.J.; Moresi, V.; Adamo, S.; et al. Peroxynitrite Activates the NLRP3 Inflammasome Cascade in SOD1(G93A) Mouse Model of Amyotrophic Lateral Sclerosis. Mol. Neurobiol. 2018, 55, 2350–2361. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Bai, Z.; Qin, X.; Cheng, Y. Aberrations in Oxidative Stress Markers in Amyotrophic Lateral Sclerosis: A Systematic Review and Meta-Analysis. Oxid. Med. Cell Longev. 2019, 2019, 1712323. [Google Scholar] [CrossRef] [PubMed]
- Kazama, M.; Kato, Y.; Kakita, A.; Noguchi, N.; Urano, Y.; Masui, K.; Niida-Kawaguchi, M.; Yamamoto, T.; Watabe, K.; Kitagawa, K.; et al. Astrocytes release glutamate via cystine/glutamate antiporter upregulated in response to increased oxidative stress related to sporadic amyotrophic lateral sclerosis. Neuropathology 2020, 40, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Kolarcik, C.L.; Bowser, R. Retinoid signaling alterations in amyotrophic lateral sclerosis. Am. J. Neurodegener. Dis. 2012, 1, 130–145. [Google Scholar] [CrossRef]
- Sun, M.S.; Jin, H.; Sun, X.; Huang, S.; Zhang, F.L.; Guo, Z.N.; Yang, Y. Free Radical Damage in Ischemia-Reperfusion Injury: An Obstacle in Acute Ischemic Stroke after Revascularization Therapy. Oxid. Med. Cell Longev. 2018, 2018, 3804979. [Google Scholar] [CrossRef]
- Kalogeris, T.; Baines, C.P.; Krenz, M.; Korthuis, R.J. Cell biology of ischemia/reperfusion injury. Int. Rev. Cell Mol. Biol. 2012, 298, 229–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Xiong, X.; Wu, X.; Ye, Y.; Jian, Z.; Zhi, Z.; Gu, L. Targeting Oxidative Stress and Inflammation to Prevent Ischemia-Reperfusion Injury. Front. Mol. Neurosci. 2020, 13, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Qazzaz, N.K.; Ali, S.H.; Ahmad, S.A.; Islam, S.; Mohamad, K. Cognitive impairment and memory dysfunction after a stroke diagnosis: A post-stroke memory assessment. Neuropsychiatr. Dis. Treat. 2014, 10, 1677–1691. [Google Scholar] [CrossRef] [Green Version]
- Ahad, M.A.; Kumaran, K.R.; Ning, T.; Mansor, N.I.; Effendy, M.A.; Damodaran, T.; Lingam, K.; Wahab, H.A.; Nordin, N.; Liao, P.; et al. Insights into the neuropathology of cerebral ischemia and its mechanisms. Rev. Neurosci. 2020, 31, 521–538. [Google Scholar] [CrossRef]
- Lorenzano, S.; Rost, N.S.; Khan, M.; Li, H.; Lima, F.O.; Maas, M.B.; Green, R.E.; Thankachan, T.K.; Dipietro, A.J.; Arai, K.; et al. Oxidative Stress Biomarkers of Brain Damage: Hyperacute Plasma F2-Isoprostane Predicts Infarct Growth in Stroke. Stroke 2018, 49, 630–637. [Google Scholar] [CrossRef]
- Nakano, Y.; Yamashita, T.; Li, Q.; Sato, K.; Ohta, Y.; Morihara, R.; Hishikawa, N.; Abe, K. Time-dependent change of in vivo optical imaging of oxidative stress in a mouse stroke model. J. Neurosci. Res. 2017, 95, 2030–2039. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, M.; Suenaga, J.; Takase, H.; Araki, K.; Yao, T.; Fujimura, T.; Murayama, K.; Okumura, K.; Ueno, R.; Shimizu, N.; et al. Oxidative stress-responsive apoptosis inducing protein (ORAIP) plays a critical role in cerebral ischemia/reperfusion injury. Sci. Rep. 2019, 9, 13512. [Google Scholar] [CrossRef]
- Sacco, R.L.; Kasner, S.E.; Broderick, J.P.; Caplan, L.R.; Connors, J.J.; Culebras, A.; Elkind, M.S.; George, M.G.; Hamdan, A.D.; Higashida, R.T.; et al. An updated definition of stroke for the 21st century: A statement for healthcare professionals from the American Heart Association/American Stroke Association. Stroke 2013, 44, 2064–2089. [Google Scholar] [CrossRef] [Green Version]
- Lou, Z.; Wang, A.P.; Duan, X.M.; Hu, G.H.; Song, G.L.; Zuo, M.L.; Yang, Z.B. Upregulation of NOX2 and NOX4 Mediated by TGF-beta Signaling Pathway Exacerbates Cerebral Ischemia/Reperfusion Oxidative Stress Injury. Cell Physiol. Biochem. 2018, 46, 2103–2113. [Google Scholar] [CrossRef]
- Xu, N.; Meng, H.; Liu, T.; Feng, Y.; Qi, Y.; Wang, H. TRPC1 Deficiency Exacerbates Cerebral Ischemia/Reperfusion-Induced Neurological Injury by Potentiating Nox4-Derived Reactive Oxygen Species Generation. Cell Physiol. Biochem. 2018, 51, 1723–1738. [Google Scholar] [CrossRef]
- Staiculescu, M.C.; Foote, C.; Meininger, G.A.; Martinez-Lemus, L.A. The role of reactive oxygen species in microvascular remodeling. Int. J. Mol. Sci. 2014, 15, 23792–23835. [Google Scholar] [CrossRef] [Green Version]
- Rempe, R.G.; Hartz, A.M.S.; Bauer, B. Matrix metalloproteinases in the brain and blood-brain barrier: Versatile breakers and makers. J. Cereb. Blood Flow Metab. 2016, 36, 1481–1507. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Shao, L.; Ma, L. Cerebral Edema Formation After Stroke: Emphasis on Blood-Brain Barrier and the Lymphatic Drainage System of the Brain. Front. Cell Neurosci. 2021, 15, 716825. [Google Scholar] [CrossRef]
- van Den Hauwe, L.; Sundgren, P.C.; Flanders, A.E. Spinal Trauma and Spinal Cord Injury (SCI). In Diseases of the Brain, Head and Neck, Spine 2020–2023: Diagnostic Imaging; Hodler, J., Kubik-Huch, R.A., von Schulthess, G.K., Eds.; IDKD Springer Series: Cham, Switzerland, 2020. [Google Scholar] [CrossRef] [Green Version]
- Eckert, M.J.; Martin, M.J. Trauma: Spinal Cord Injury. Surg. Clin. N. Am. 2017, 97, 1031–1045. [Google Scholar] [CrossRef]
- Ahuja, C.S.; Nori, S.; Tetreault, L.; Wilson, J.; Kwon, B.; Harrop, J.; Choi, D.; Fehlings, M.G. Traumatic Spinal Cord Injury-Repair and Regeneration. Neurosurgery 2017, 80, S9–S22. [Google Scholar] [CrossRef]
- Lin, J.; Xiong, Z.; Gu, J.; Sun, Z.; Jiang, S.; Fan, D.; Li, W. Sirtuins: Potential Therapeutic Targets for Defense against Oxidative Stress in Spinal Cord Injury. Oxid. Med. Cell Longev. 2021, 2021, 7207692. [Google Scholar] [CrossRef]
- Bock, F.J.; Tait, S.W.G. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar] [CrossRef]
- Rabchevsky, A.G.; Michael, F.M.; Patel, S.P. Mitochondria focused neurotherapeutics for spinal cord injury. Exp. Neurol. 2020, 330, 113332. [Google Scholar] [CrossRef]
- Popa, C.; Popa, F.; Grigorean, V.T.; Onose, G.; Sandu, A.M.; Popescu, M.; Burnei, G.; Strambu, V.; Sinescu, C. Vascular dysfunctions following spinal cord injury. J. Med. Life 2010, 3, 275–285. [Google Scholar]
- Hussain, G.; Wang, J.; Rasul, A.; Anwar, H.; Qasim, M.; Zafar, S.; Aziz, N.; Razzaq, A.; Hussain, R.; de Aguilar, J.G.; et al. Current Status of Therapeutic Approaches against Peripheral Nerve Injuries: A Detailed Story from Injury to Recovery. Int. J. Biol. Sci. 2020, 16, 116–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walco, G.A.; Dworkin, R.H.; Krane, E.J.; LeBel, A.A.; Treede, R.D. Neuropathic pain in children: Special considerations. Mayo Clin. Proc. 2010, 85, S33–S41. [Google Scholar] [CrossRef] [Green Version]
- Agbaje, J.O.; Van de Casteele, E.; Hiel, M.; Verbaanderd, C.; Lambrichts, I.; Politis, C. Neuropathy of Trigeminal Nerve Branches After Oral and Maxillofacial Treatment. J. Maxillofac. Oral Surg. 2016, 15, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menorca, R.M.; Fussell, T.S.; Elfar, J.C. Nerve physiology: Mechanisms of injury and recovery. Hand Clin. 2013, 29, 317–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khezri, M.K.; Turkkan, A.; Koc, C.; Salman, B.; Levent, P.; Cakir, A.; Kafa, I.M.; Cansev, M.; Bekar, A. Anti-Apoptotic and Anti-Oxidant Effects of Systemic Uridine Treatment in an Experimental Model of Sciatic Nerve Injury. Turk. Neurosurg. 2021, 31, 373–378. [Google Scholar] [CrossRef]
- Lattimore, M.R., Jr.; Varr, W.F., 3rd. Disposable soft lens ulcerative keratitis in an Army aviator: A case report. Aviat. Space Environ. Med. 1991, 62, 888–889. [Google Scholar]
- Costa, L.S.; Aidar, F.J.; Matos, D.G.; Oliveira, J.U.; Santos, J.L.D.; Almeida-Neto, P.F.; Souza, R.F.; Pereira, D.D.; Garrido, N.D.; Nunes-Silva, A.; et al. Effects of Resistance Training and Bowdichia virgilioides Hydroethanolic Extract on Oxidative Stress Markers in Rats Submitted to Peripheral Nerve Injury. Antioxidants 2020, 9, 941. [Google Scholar] [CrossRef]
- Zafar, S.; Anwar, H.; Qasim, M.; Irfan, S.; Maqbool, J.; Sajid, F.; Naqvi, S.A.R.; Hussain, G. Calotropis procera (root) escalates functions rehabilitation and attenuates oxidative stress in a mouse model of peripheral nerve injury. Pak. J. Pharm. Sci. 2020, 33, 2801–2807. [Google Scholar]
- Tang, C.; Han, R.; Wu, J.; Fang, F. Effects of baicalin capsules combined with alpha-lipoic acid on nerve conduction velocity, oxidative stress and inflammatory injury in patients wi.ith diabetic peripheral neuropathy. Am. J. Transl. Res. 2021, 13, 2774–2783. [Google Scholar]
- Luca, M.; Luca, A.; Calandra, C. The Role of Oxidative Damage in the Pathogenesis and Progression of Alzheimer’s Disease and Vascular Dementia. Oxid. Med. Cell Longev. 2015, 2015, 504678. [Google Scholar] [CrossRef] [Green Version]
- Perluigi, M.; Butterfield, D.A. Oxidative Stress and Down Syndrome: A Route toward Alzheimer-Like Dementia. Curr. Gerontol. Geriatr. Res. 2012, 2012, 724904. [Google Scholar] [CrossRef] [Green Version]
- Pangrazzi, L.; Balasco, L.; Bozzi, Y. Oxidative Stress and Immune System Dysfunction in Autism Spectrum Disorders. Int. J. Mol. Sci. 2020, 21, 3293. [Google Scholar] [CrossRef]
- Oztop, D.; Altun, H.; Baskol, G.; Ozsoy, S. Oxidative stress in children with attention deficit hyperactivity disorder. Clin. Biochem 2012, 45, 745–748. [Google Scholar] [CrossRef]
- Paul, B.D.; Snyder, S.H. Impaired Redox Signaling in Huntington’s Disease: Therapeutic Implications. Front. Mol. Neurosci. 2019, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Ohl, K.; Tenbrock, K.; Kipp, M. Oxidative stress in multiple sclerosis: Central and peripheral mode of action. Exp. Neurol. 2016, 277, 58–67. [Google Scholar] [CrossRef]
- Bhatt, S.; Nagappa, A.N.; Patil, C.R. Role of oxidative stress in depression. Drug Discov. Today 2020, 25, 1270–1276. [Google Scholar] [CrossRef]
- Geronzi, U.; Lotti, F.; Grosso, S. Oxidative stress in epilepsy. Expert Rev. Neurother. 2018, 18, 427–434. [Google Scholar] [CrossRef]
- Khatri, N.; Thakur, M.; Pareek, V.; Kumar, S.; Sharma, S.; Datusalia, A.K. Oxidative Stress: Major Threat in Traumatic Brain Injury. CNS Neurol. Disord. Drug Targets 2018, 17, 689–695. [Google Scholar] [CrossRef]
- Elfawy, H.A.; Das, B. Crosstalk between mitochondrial dysfunction, oxidative stress, and age related neurodegenerative disease: Etiologies and therapeutic strategies. Life Sci. 2019, 218, 165–184. [Google Scholar] [CrossRef]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, A. A review on mitochondrial restorative mechanism of antioxidants in Alzheimer’s disease and other neurological conditions. Front. Pharmacol. 2015, 6, 206. [Google Scholar] [CrossRef] [Green Version]
- Gerhke, S.A.; Shibli, J.A.; Salles, M.B. Potential of the use of an antioxidant compound to promote peripheral nerve regeneration after injury. Neural Regen. Res. 2015, 10, 1063–1064. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.Y.; Wang, N.; Li, S.; Hong, M.; Wang, X.; Feng, Y. The Reactive Oxygen Species in Macrophage Polarization: Reflecting Its Dual Role in Progression and Treatment of Human Diseases. Oxid. Med. Cell Longev. 2016, 2016, 2795090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, A.; Terzo, S.; Mule, F. Natural Compounds as Beneficial Antioxidant Agents in Neurodegenerative Disorders: A Focus on Alzheimer’s Disease. Antioxidants 2019, 8, 608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.R.; Dos Santos, N.B.; Scavone, C.; Munhoz, C.D. Nrf2/ARE Pathway Modulation by Dietary Energy Regulation in Neurological Disorders. Front. Pharmacol. 2019, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Villavicencio Tejo, F.; Quintanilla, R.A. Contribution o.of the Nrf2 Pathway on Oxidative Damage and Mitochondrial Failure in Parkinson and Alzheimer’s Disease. Antioxidants 2021, 10, 1069. [Google Scholar] [CrossRef]
- Lee, K.H.; Cha, M.; Lee, B.H. Neuroprotective Effect of Antioxidants in the Brain. Int. J. Mol. Sci. 2020, 21, 7152. [Google Scholar] [CrossRef]
- Poljsak, B. Strategies for reducing or preventing the generation of oxidative stress. Oxid. Med. Cell Longev. 2011, 2011, 194586. [Google Scholar] [CrossRef] [Green Version]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative stress mitigation by antioxidants—An overview on their chemistry and influences on health status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid. Med. Cell Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef] [Green Version]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [Green Version]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New roles in redox signaling for an old antioxidant. Front. Pharmacol. 2014, 5, 196. [Google Scholar] [CrossRef] [Green Version]
- Lushchak, V.I. Glutathione homeostasis and functions: Potential targets for medical interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef] [Green Version]
- Bjorklund, G.; Aaseth, J.; Crisponi, G.; Rahman, M.M.; Chirumbolo, S. Insights on alpha lipoic and dihydrolipoic acids as promising scavengers of oxidative stress and possible chelators in mercury toxicology. J. Inorg. Biochem. 2019, 195, 111–119. [Google Scholar] [CrossRef]
- Fabbrini, E.; Serafini, M.; Colic Baric, I.; Hazen, S.L.; Klein, S. Effect of plasma uric acid on antioxidant capacity, oxidative stress, and insulin sensitivity in obese subjects. Diabetes 2014, 63, 976–981. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhang, Y.; Xu, H.; Luo, X.; Yu, J.; Liu, J.; Chang, R.C. Neuroprotection of Coenzyme Q10 in Neurodegenerative Diseases. Curr. Top. Med. Chem 2016, 16, 858–866. [Google Scholar] [CrossRef]
- Conti, V.; Izzo, V.; Corbi, G.; Russomanno, G.; Manzo, V.; De Lise, F.; Di Donato, A.; Filippelli, A. Antioxidant Supplementation in the Treatment of Aging-Associated Diseases. Front. Pharmacol. 2016, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Bouayed, J.; Bohn, T. Exogenous antioxidants—Double-edged swords in cellular redox state: Health beneficial effects at physiologic doses versus deleterious effects at high doses. Oxid. Med. Cell Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Jakubczyk, K.; Kaldunska, J.; Dec, K.; Kawczuga, D.; Janda, K. Antioxidant properties of small-molecule non-enzymatic compounds. Pol. Merkur. Lekarski 2020, 48, 128–132. [Google Scholar]
- Kiokias, S.; Proestos, C.; Oreopoulou, V. Effect of Natural Food Antioxidants against LDL and DNA Oxidative Changes. Antioxidants 2018, 7, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janciauskiene, S. The Beneficial Effects of Antioxidants in Health and Diseases. Chronic Obstr. Pulm. Dis. 2020, 7, 182–202. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, S.; Raza, S.T.; Ahmed, F.; Ahmad, A.; Abbas, S.; Mahdi, F. The role of vitamin e in human health and some diseases. Sultan Qaboos Univ. Med. J. 2014, 14, e157–e165. [Google Scholar]
- Lee, K.H.; Kim, U.J.; Cha, M.; Lee, B.H. Chronic Treatment of Ascorbic Acid Leads to Age-Dependent Neuroprotection against Oxidative Injury in Hippocampal Slice Cultures. Int. J. Mol. Sci. 2021, 22, 1608. [Google Scholar] [CrossRef]
- Ramani, M.; van Groen, T.; Kadish, I.; Ambalavanan, N.; McMahon, L.L. Vitamin A and retinoic acid combination attenuates neonatal hyperoxia-induced neurobehavioral impairment in adult mice. Neurobiol. Learn. Mem. 2017, 141, 209–216. [Google Scholar] [CrossRef]
- Chen, P.; Li, L.; Gao, Y.; Xie, Z.; Zhang, Y.; Pan, Z.; Tu, Y.; Wang, H.; Han, Q.; Hu, X.; et al. beta-carotene provides neuro protection after experimental traumatic brain injury via the Nrf2-ARE pathway. J. Integr. Neurosci. 2019, 18, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Hwang, S.; Yu, J.H.; Lim, J.W.; Kim, H. Lycopene inhibits regulator of calcineurin 1-mediated apoptosis by reducing oxidative stress and down-regulating Nucling in neuronal cells. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Huang, C.; Wen, C.; Yang, M.; Gan, D.; Fan, C.; Li, A.; Li, Q.; Zhao, J.; Zhu, L.; Lu, D. Lycopene protects against t-BHP-induced neuronal oxidative damage and apoptosis via activation of the PI3K/Akt pathway. Mol. Biol. Rep. 2019, 46, 3387–3397. [Google Scholar] [CrossRef]
- Manochkumar, J.; Doss, C.G.P.; El-Seedi, H.R.; Efferth, T.; Ramamoorthy, S. The neuroprotective potential of carotenoids in vitro and in vivo. Phytomedicine 2021, 91, 153676. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Bodoira, R.; Maestri, D. Phenolic Compounds from Nuts: Extraction, Chemical Profiles, and Bioactivity. J. Agric. Food Chem. 2020, 68, 927–942. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Syarifah-Noratiqah, S.B.; Naina-Mohamed, I.; Zulfarina, M.S.; Qodriyah, H.M.S. Natural Polyphenols in the Treatment of Alzheimer’s Disease. Curr. Drug Targets 2018, 19, 927–937. [Google Scholar] [CrossRef]
- Ma, H.; Johnson, S.L.; Liu, W.; DaSilva, N.A.; Meschwitz, S.; Dain, J.A.; Seeram, N.P. Evaluation of Polyphenol Anthocyanin-Enriched Extracts of Blackberry, Black Raspberry, Blueberry, Cranberry, Red Raspberry, and Strawberry for Free Radical Scavenging, Reactive Carbonyl Species Trapping, Anti-Glycation, Anti-beta-Amyloid Aggregation, and Microglial Neuroprotective Effects. Int. J. Mol. Sci. 2018, 19, 461. [Google Scholar] [CrossRef] [Green Version]
- Lourenco, S.C.; Moldao-Martins, M.; Alves, V.D. Antioxidants of Natural Plant Origins: From Sources to Food Industry Applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef] [Green Version]
- Taghvaei, M.; Jafari, S.M. Application and stability of natural antioxidants in edible oils in order to substitute synthetic additives. J. Food Sci. Technol. 2015, 52, 1272–1282. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Tang, X.; Cao, X.; Yu, L.; Chen, J.; Zhao, H.; Chen, Y.; Han, L.; Bao, X.; Li, F.; et al. 4-((5-(Tert-butyl)-3-chloro-2-hydroxybenzyl) amino)-2-hydroxybenzoic acid protects against oxygen-glucose deprivation/reperfusion injury. Life Sci. 2018, 204, 46–54. [Google Scholar] [CrossRef]
- Mohsin Alvi, A.; Tariq Al Kury, L.; Umar Ijaz, M.; Ali Shah, F.; Tariq Khan, M.; Sadiq Sheikh, A.; Nadeem, H.; Khan, A.U.; Zeb, A.; Li, S. Post-Treatment of Synthetic Polyphenolic 1,3,4 Oxadiazole Compound A3, Attenuated Ischemic Stroke-Induced Neuroinflammation and Neurodegeneration. Biomolecules 2020, 10, 816. [Google Scholar] [CrossRef] [PubMed]
- Che, H.; Li, Q.; Zhang, T.; Wang, D.; Yang, L.; Xu, J.; Yanagita, T.; Xue, C.; Chang, Y.; Wang, Y. Effects of Astaxanthin and Docosahexaenoic-Acid-Acylated Astaxanthin on Alzheimer’s Disease in APP/PS1 Double-Transgenic Mice. J. Agric. Food Chem. 2018, 66, 4948–4957. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Shi, H.H.; Xu, J.; Yanagita, T.; Xue, C.H.; Zhang, T.T.; Wang, Y.M. Docosahexaenoic acid-acylated astaxanthin ester exhibits superior performance over non-esterified astaxanthin in preventing behavioral deficits coupled with apoptosis in MPTP-induced mice with Parkinson’s disease. Food Funct. 2020, 11, 8038–8050. [Google Scholar] [CrossRef] [PubMed]
- Akaishi, T.; Abe, K. CNB-001, a synthetic pyrazole derivative of curcumin, suppresses lipopolysaccharide-induced nitric oxide production through the inhibition of NF-kappaB and p38 MAPK pathways in microglia. Eur. J. Pharmacol. 2018, 819, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ma, Z.; Ning, X.; Chen, Y.; Tian, C.; Wang, X.; Zhang, Z.; Liu, J. A Novel Synthetic Precursor of Styryl Sulfone Neuroprotective Agents Inhibits Neuroinflammatory Responses and Oxidative Stress Damage through the P38 Signaling Pathway in the Cell and Animal Model of Parkinson’s Disease. Molecules 2021, 26, 5371. [Google Scholar] [CrossRef] [PubMed]
- Song, X.Y.; Hu, J.F.; Wu, D.H.; Ji, H.J.; Chen, N.H. IMM-H004, a Novel Coumarin Derivative Compound, Inhibits H2O2-Induced Neurotoxicity via Antioxidant and Antiapoptosis in PC12 Cells. J. Stroke Cerebrovasc. Dis. 2018, 27, 3396–3403. [Google Scholar] [CrossRef]
- Saleh, T.M.; Saleh, M.C.; Connell, B.J.; Kucukkaya, I.; Abd-El-Aziz, A.S. A novel synthetic chemical entity (UPEI-800) is neuroprotective in vitro and in an in vivo rat model of oxidative stress. Clin. Exp. Pharmacol. Physiol. 2017, 44, 993–1000. [Google Scholar] [CrossRef]
- Lee, S.Y.; Chiu, Y.J.; Yang, S.M.; Chen, C.M.; Huang, C.C.; Lee-Chen, G.J.; Lin, W.; Chang, K.H. Novel synthetic chalcone-coumarin hybrid for Abeta aggregation reduction, antioxidation, and neuroprotection. CNS Neurosci. Ther. 2018, 24, 1286–1298. [Google Scholar] [CrossRef]
- Uppakara, K.; Jamornwan, S.; Duan, L.X.; Yue, K.R.; Sunrat, C.; Dent, E.W.; Wan, S.B.; Saengsawang, W. Novel alpha-Lipoic Acid/3-n-Butylphthalide Conjugate Enhances Protective Effects against Oxidative Stress and 6-OHDA Induced Neuronal Damage. ACS Chem. Neurosci. 2020, 11, 1634–1642. [Google Scholar] [CrossRef]
- Villamena, F.A.; Das, A.; Nash, K.M. Potential implication of the chemical properties and bioactivity of nitrone spin traps for therapeutics. Future Med. Chem. 2012, 4, 1171–1207. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.J. Detection and characterisation of radicals using electron paramagnetic resonance (EPR) spin trapping and related methods. Methods 2016, 109, 21–30. [Google Scholar] [CrossRef]
- Floyd, R.A.; Castro Faria Neto, H.C.; Zimmermaan, G.A.; Hensley, K.; Towner, R.A. Nitrone-based therapeutics for neurodegenerative diseases: Their use alone or in combination with lanthionines. Free Radic. Biol. Med. 2013, 62, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska, D.G.; Mediavilla, L.; Cuarental, L.; Glowacka, I.E.; Marco-Contelles, J.; Hadjipavlou-Litina, D.; Lopez-Munoz, F.; Oset-Gasque, M.J. Synthesis and Neuroprotective Properties of N-Substituted C-Dialkoxyphosphorylated Nitrones. ACS Omega 2019, 4, 8581–8587. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, H. Edaravone for the treatment of amyotrophic lateral sclerosis. Expert. Rev. Neurother. 2019, 19, 185–193. [Google Scholar] [CrossRef]
- Watanabe, T.; Tahara, M.; Todo, S. The novel antioxidant edaravone: From bench to bedside. Cardiovasc. Ther. 2008, 26, 101–114. [Google Scholar] [CrossRef]
- Kang, L.; Liu, S.; Li, J.; Tian, Y.; Xue, Y.; Liu, X. The mitochondria-targeted anti-oxidant MitoQ protects against intervertebral disc degeneration by ameliorating mitochondrial dysfunction and redox imbalance. Cell Prolif. 2020, 53, 1–19. [Google Scholar] [CrossRef]
- Young, M.L.; Franklin, J.L. The mitochondria-targeted antioxidant MitoQ inhibits memory loss, neuropathology, and extends lifespan in aged 3xTg-AD mice. Mol. Cell Neurosci. 2019, 101, 103409. [Google Scholar] [CrossRef]
- Suarez-Rivero, J.M.; Pastor-Maldonado, C.J.; Povea-Cabello, S.; Alvarez-Cordoba, M.; Villalon-Garcia, I.; Munuera-Cabeza, M.; Suarez-Carrillo, A.; Talaveron-Rey, M.; Sanchez-Alcazar, J.A. Coenzyme Q10 Analogues: Benefits and Challenges for Therapeutics. Antioxidants 2021, 10, 236. [Google Scholar] [CrossRef]
- Shinn, L.J.; Lagalwar, S. Treating Neurodegenerative Disease with Antioxidants: Efficacy of the Bioactive Phenol Resveratrol and Mitochondrial-Targeted MitoQ and SkQ. Antioxidants 2021, 10, 573. [Google Scholar] [CrossRef]
- Fonseca-Fonseca, L.A.; Nunez-Figueredo, Y.; Sanchez, J.R.; Guerra, M.W.; Ochoa-Rodriguez, E.; Verdecia-Reyes, Y.; Hernadez, R.D.; Menezes-Filho, N.J.; Costa, T.C.S.; de Santana, W.A.; et al. KM-34, a Novel Antioxidant Compound, Protects against 6-Hydroxydopamine-Induced Mitochondrial Damage and Neurotoxicity. Neurotox Res. 2019, 36, 279–291. [Google Scholar] [CrossRef]
- Federico, A.; Cardaioli, E.; Da Pozzo, P.; Formichi, P.; Gallus, G.N.; Radi, E. Mitochondria, oxidative stress and neurodegeneration. J. Neurol. Sci. 2012, 322, 254–262. [Google Scholar] [CrossRef]
- Yang, M.; Lian, N.; Yu, Y.; Wang, Y.; Xie, K.; Yu, Y. Coenzyme Q10 alleviates sevofluraneinduced neuroinflammation by regulating the levels of apolipoprotein E and phosphorylated tau protein in mouse hippocampal neurons. Mol. Med. Rep. 2020, 22, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Komaki, H.; Faraji, N.; Komaki, A.; Shahidi, S.; Etaee, F.; Raoufi, S.; Mirzaei, F. Investigation of protective effects of coenzyme Q10 on impaired synaptic plasticity in a male rat model of Alzheimer’s disease. Brain Res. Bull. 2019, 147, 14–21. [Google Scholar] [CrossRef]
- Wang, J.; Li, L.; Wang, Z.; Cui, Y.; Tan, X.; Yuan, T.; Liu, Q.; Liu, Z.; Liu, X. Supplementation of lycopene attenuates lipopolysaccharide-induced amyloidogenesis and cognitive impairments via mediating neuroinflammation and oxidative stress. J. Nutr. Biochem. 2018, 56, 16–25. [Google Scholar] [CrossRef]
- Han, J.H.; Lee, Y.S.; Im, J.H.; Ham, Y.W.; Lee, H.P.; Han, S.B.; Hong, J.T. Astaxanthin Ameliorates Lipopolysaccharide-Induced Neuroinflammation, Oxidative Stress and Memory Dysfunction through Inactivation of the Signal Transducer and Activator of Transcription 3 Pathway. Mar. Drugs 2019, 17, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hira, S.; Saleem, U.; Anwar, F.; Sohail, M.F.; Raza, Z.; Ahmad, B. beta-Carotene: A Natural Compound Improves Cognitive Impairment and Oxidative Stress in a Mouse Model of Streptozotocin-Induced Alzheimer’s Disease. Biomolecules 2019, 9, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhami, M.; Raj, K.; Singh, S. Neuroprotective Effect of Fucoxanthin against Intracerebroventricular Streptozotocin (ICV-STZ) Induced Cognitive Impairment in Experimental Rats. Curr. Alzheimer Res. 2021, 18, 623–637. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, T.; Li, W.; Gao, N.; Zhang, T. Resveratrol attenuates oxidative damage through activating mitophagy in an in vitro model of Alzheimer’s disease. Toxicol. Lett. 2018, 282, 100–108. [Google Scholar] [CrossRef]
- Khan, M.S.; Muhammad, T.; Ikram, M.; Kim, M.O. Dietary Supplementation of the Antioxidant Curcumin Halts Systemic LPS-Induced Neuroinflammation-Associated Neurodegeneration and Memory/Synaptic Impairment via the JNK/NF-kappaB/Akt Signaling Pathway in Adult Rats. Oxid. Med. Cell Longev. 2019, 2019, 7860650. [Google Scholar] [CrossRef] [Green Version]
- Doghri, R.; Ellefi, A.; Degrach, I.; Srairi-Abid, N.; Gati, A. Curcumin Attenuated Neurotoxicity in Sporadic Animal Model of Alzheimer’s Disease. Molecules 2021, 26, 3011. [Google Scholar] [CrossRef]
- Kong, D.; Yan, Y.; He, X.Y.; Yang, H.; Liang, B.; Wang, J.; He, Y.; Ding, Y.; Yu, H. Effects of Resveratrol on the Mechanisms of Antioxidants and Estrogen in Alzheimer’s Disease. Biomed. Res. Int 2019, 2019, 8983752. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Ali, T.; Kim, M.W.; Jo, M.H.; Jo, M.G.; Badshah, H.; Kim, M.O. Anthocyanins protect against LPS-induced oxidative stress-mediated neuroinflammation and neurodegeneration in the adult mouse cortex. Neurochem. Int. 2016, 100, 1–10. [Google Scholar] [CrossRef]
- Ali, T.; Kim, T.; Rehman, S.U.; Khan, M.S.; Amin, F.U.; Khan, M.; Ikram, M.; Kim, M.O. Natural Dietary Supplementation of Anthocyanins via PI3K/Akt/Nrf2/HO-1 Pathways Mitigate Oxidative Stress, Neurodegeneration, and Memory Impairment in a Mouse Model of Alzheimer’s Disease. Mol. Neurobiol. 2018, 55, 6076–6093. [Google Scholar] [CrossRef]
- Khan, M.S.; Ali, T.; Kim, M.W.; Jo, M.H.; Chung, J.I.; Kim, M.O. Anthocyanins Improve Hippocampus-Dependent Memory Function and Prevent Neurodegeneration via JNK/Akt/GSK3beta Signaling in LPS-Treated Adult Mice. Mol. Neurobiol. 2019, 56, 671–687. [Google Scholar] [CrossRef]
- Kushairi, N.; Phan, C.W.; Sabaratnam, V.; David, P.; Naidu, M. Lion’s Mane Mushroom, Hericium erinaceus (Bull.: Fr.) Pers. Suppresses H2O2-Induced Oxidative Damage and LPS-Induced Inflammation in HT22 Hippocampal Neurons and BV2 Microglia. Antioxidants 2019, 8, 261. [Google Scholar] [CrossRef] [Green Version]
- Cordaro, M.; Salinaro, A.T.; Siracusa, R.; D’Amico, R.; Impellizzeri, D.; Scuto, M.; Ontario, M.L.; Cuzzocrea, S.; Di Paola, R.; Fusco, R.; et al. Key Mechanisms and Potential Implications of Hericium erinaceus in NLRP3 Inflammasome Activation by Reactive Oxygen Species during Alzheimer’s Disease. Antioxidants 2021, 10, 1664. [Google Scholar] [CrossRef]
- Frontinan-Rubio, J.; Sancho-Bielsa, F.J.; Peinado, J.R.; LaFerla, F.M.; Gimenez-Llort, L.; Duran-Prado, M.; Alcain, F.J. Sex-dependent co-occurrence of hypoxia and beta-amyloid plaques in hippocampus and entorhinal cortex is reversed by long-term treatment with ubiquinol and ascorbic acid in the 3xTg-AD mouse model of Alzheimer’s disease. Mol. Cell Neurosci. 2018, 92, 67–81. [Google Scholar] [CrossRef]
- Yu, L.; Wang, W.; Pang, W.; Xiao, Z.; Jiang, Y.; Hong, Y. Dietary Lycopene Supplementation Improves Cognitive Performances in Tau Transgenic Mice Expressing P301L Mutation via Inhibiting Oxidative Stress and Tau Hyperphosphorylation. J. Alzheimers Dis. 2017, 57, 475–482. [Google Scholar] [CrossRef]
- Ibrahim Fouad, G. Combination of Omega 3 and Coenzyme Q10 Exerts Neuroprotective Potential against Hypercholesterolemia-Induced Alzheimer’s-Like Disease in Rats. Neurochem. Res. 2020, 45, 1142–1155. [Google Scholar] [CrossRef]
- Zaky, A.; Bassiouny, A.; Farghaly, M.; El-Sabaa, B.M. A Combination of Resveratrol and Curcumin is Effective Against Aluminum Chloride-Induced Neuroinflammation in Rats. J. Alzheimers Dis. 2017, 60, S221–S235. [Google Scholar] [CrossRef] [PubMed]
- Wan, T.; Wang, Z.; Luo, Y.; Zhang, Y.; He, W.; Mei, Y.; Xue, J.; Li, M.; Pan, H.; Li, W.; et al. FA-97, a New Synthetic Caffeic Acid Phenethyl Ester Derivative, Protects against Oxidative Stress-Mediated Neuronal Cell Apoptosis and Scopolamine-Induced Cognitive Impairment by Activating Nrf2/HO-1 Signaling. Oxid. Med. Cell Longev. 2019, 2019, 8239642. [Google Scholar] [CrossRef] [PubMed]
- Schirinzi, T.; Martella, G.; Imbriani, P.; Di Lazzaro, G.; Franco, D.; Colona, V.L.; Alwardat, M.; Sinibaldi Salimei, P.; Mercuri, N.B.; Pierantozzi, M.; et al. Dietary Vitamin E as a Protective Factor for Parkinson’s Disease: Clinical and Experimental Evidence. Front. Neurol. 2019, 10, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Nuccio, F.; Cianciulli, A.; Porro, C.; Kashyrina, M.; Ruggiero, M.; Calvello, R.; Miraglia, A.; Nicolardi, G.; Lofrumento, D.D.; Panaro, M.A. Inflammatory Response Modulation by Vitamin C in an MPTP Mouse Model of Parkinson’s Disease. Biology 2021, 10, 1155. [Google Scholar] [CrossRef]
- Zeng, X.; Xu, K.; Wang, J.; Xu, Y.; Qu, S. Pretreatment of Ascorbic Acid Inhibits MPTP-Induced Astrocytic Oxidative Stress through Suppressing NF-kappaB Signaling. Neural Plast. 2020, 2020, 8872296. [Google Scholar] [CrossRef]
- Attia, H.N.; Maklad, Y.A. Neuroprotective effects of coenzyme Q10 on paraquat-induced Parkinson’s disease in experimental animals. Behav. Pharmacol. 2018, 29, 79–86. [Google Scholar] [CrossRef]
- Park, E.; Gim, J.; Kim, D.K.; Kim, C.S.; Chun, H.S. Protective Effects of Alpha-Lipoic Acid on Glutamate-Induced Cytotoxicity in C6 Glioma Cells. Biol. Pharm. Bull. 2019, 42, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Cheng, Y. Alpha-lipoic acid alleviated 6-OHDA-induced cell damage by inhibiting AMPK/mTOR mediated autophagy. Neuropharmacology 2019, 155, 98–103. [Google Scholar] [CrossRef]
- Rao, S.V.; Hemalatha, P.; Yetish, S.; Muralidhara, M.; Rajini, P.S. Prophylactic neuroprotective propensity of Crocin, a carotenoid against rotenone induced neurotoxicity in mice: Behavioural and biochemical evidence. Metab. Brain Dis. 2019, 34, 1341–1353. [Google Scholar] [CrossRef]
- Wu, W.; Han, H.; Liu, J.; Tang, M.; Wu, X.; Cao, X.; Zhao, T.; Lu, Y.; Niu, T.; Chen, J.; et al. Fucoxanthin Prevents 6-OHDA-Induced Neurotoxicity by Targeting Keap1. Oxid. Med. Cell Longev. 2021, 2021, 6688708. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Adedara, A.O.; Adie, M.A.; Vicente-Crespo, M.; Farombi, E.O. Resveratrol prolongs lifespan and improves 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced oxidative damage and behavioural deficits in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2018, 503, 1042–1048. [Google Scholar] [CrossRef]
- Zhang, L.F.; Yu, X.L.; Ji, M.; Liu, S.Y.; Wu, X.L.; Wang, Y.J.; Liu, R.T. Resveratrol alleviates motor and cognitive deficits and neuropathology in the A53T alpha-synuclein mouse model of Parkinson’s disease. Food Funct. 2018, 9, 6414–6426. [Google Scholar] [CrossRef]
- Singh, S.; Jamwal, S.; Kumar, P. Neuroprotective potential of Quercetin in combination with piperine against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurotoxicity. Neural Regen. Res. 2017, 12, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, M.S.U.; Tripathi, M.K.; Patel, D.K.; Singh, M.P. Resveratrol Regulates Nrf2-Mediated Expression of Antioxidant and Xenobiotic Metabolizing Enzymes in Pesticides-Induced Parkinsonism. Protein Pept. Lett. 2020, 27, 1038–1045. [Google Scholar] [CrossRef]
- Chen, Y.F.; Wu, S.N.; Gao, J.M.; Liao, Z.Y.; Tseng, Y.T.; Fulop, F.; Chang, F.R.; Lo, Y.C. The Antioxidant, Anti-Inflammatory, and Neuroprotective Properties of the Synthetic Chalcone Derivative AN07. Molecules 2020, 25, 2907. [Google Scholar] [CrossRef]
- Lee, J.A.; Son, H.J.; Choi, J.W.; Kim, J.; Han, S.H.; Shin, N.; Kim, J.H.; Kim, S.J.; Heo, J.Y.; Kim, D.J.; et al. Activation of the Nrf2 signaling pathway and neuroprotection of nigral dopaminergic neurons by a novel synthetic compound KMS99220. Neurochem. Int. 2018, 112, 96–107. [Google Scholar] [CrossRef]
- Drummond, N.J.; Davies, N.O.; Lovett, J.E.; Miller, M.R.; Cook, G.; Becker, T.; Becker, C.G.; McPhail, D.B.; Kunath, T. A synthetic cell permeable antioxidant protects neurons against acute oxidative stress. Sci. Rep. 2017, 7, 11857. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Cheng, J.; Wang, S.; Wang, X.; Jiang, H.; Yang, Y.; Wang, Y.; Zhang, C.; Liang, W.; Feng, H. alpha-Lipoic acid attenuates oxidative stress and neurotoxicity via the ERK/Akt-dependent pathway in the mutant hSOD1 related Drosophila model and the NSC34 cell line of amyotrophic lateral sclerosis. Brain Res. Bull. 2018, 140, 299–310. [Google Scholar] [CrossRef]
- Bhatia, N.K.; Srivastava, A.; Katyal, N.; Jain, N.; Khan, M.A.; Kundu, B.; Deep, S. Curcumin binds to the pre-fibrillar aggregates of Cu/Zn superoxide dismutase (SOD1) and alters its amyloidogenic pathway resulting in reduced cytotoxicity. Biochim. Biophys. Acta 2015, 1854, 426–436. [Google Scholar] [CrossRef]
- Winter, A.N.; Ross, E.K.; Wilkins, H.M.; Stankiewicz, T.R.; Wallace, T.; Miller, K.; Linseman, D.A. An anthocyanin-enriched extract from strawberries delays disease onset and extends survival in the hSOD1(G93A) mouse model of amyotrophic lateral sclerosis. Nutr. Neurosci. 2018, 21, 414–426. [Google Scholar] [CrossRef]
- Zhao, B.; Zhuang, X.; Pi, Z.; Liu, S.; Liu, Z.; Song, F. Determining the Effect of Catechins on SOD1 Conformation and Aggregation by Ion Mobility Mass Spectrometry Combined with Optical Spectroscopy. J. Am. Soc. Mass. Spectrom. 2018, 29, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, E.; Rajasekaran, R. Probing the inhibitory activity of epigallocatechin-gallate on toxic aggregates of mutant (L84F) SOD1 protein through geometry based sampling and steered molecular dynamics. J. Mol. Graph. Model. 2017, 74, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Ip, P.; Sharda, P.R.; Cunningham, A.; Chakrabartty, S.; Pande, V.; Chakrabartty, A. Quercitrin and quercetin 3-beta-d-glucoside as chemical chaperones for the A4V SOD1 ALS-causing mutant. Protein Eng. Des. Sel. 2017, 30, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Inden, M.; Shirai, K.; Sekine, S.I.; Masaki, Y.; Kurita, H.; Ichihara, K.; Inuzuka, T.; Hozumi, I. The effects of Brazilian green propolis that contains flavonols against mutant copper-zinc superoxide dismutase-mediated toxicity. Sci. Rep. 2017, 7, 2882. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.H.; Wang, S.Y.; Wang, X.D.; Jiang, H.Q.; Yang, Y.Q.; Wang, Y.; Cheng, J.L.; Zhang, C.T.; Liang, W.W.; Feng, H.L. Fisetin Exerts Antioxidant and Neuroprotective Effects in Multiple Mutant hSOD1 Models of Amyotrophic Lateral Sclerosis by Activating ERK. Neuroscience 2018, 379, 152–166. [Google Scholar] [CrossRef]
- Koza, L.A.; Winter, A.N.; Holsopple, J.; Baybayon-Grandgeorge, A.N.; Pena, C.; Olson, J.R.; Mazzarino, R.C.; Patterson, D.; Linseman, D.A. Protocatechuic Acid Extends Survival, Improves Motor Function, Diminishes Gliosis, and Sustains Neuromuscular Junctions in the hSOD1(G93A) Mouse Model of Amyotrophic Lateral Sclerosis. Nutrients 2020, 12, 1824. [Google Scholar] [CrossRef]
- Lv, C.; Maharjan, S.; Wang, Q.; Sun, Y.; Han, X.; Wang, S.; Mao, Z.; Xin, Y.; Zhang, B. alpha-Lipoic Acid Promotes Neurological Recovery after Ischemic Stroke by Activating the Nrf2/HO-1 Pathway to Attenuate Oxidative Damage. Cell Physiol. Biochem. 2017, 43, 1273–1287. [Google Scholar] [CrossRef]
- Haghnejad Azar, A.; Oryan, S.; Bohlooli, S.; Panahpour, H. Alpha-Tocopherol Reduces Brain Edema and Protects Blood-Brain Barrier Integrity following Focal Cerebral Ischemia in Rats. Med. Princ. Pract. 2017, 26, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y.; Chen, J.Y.; Wu, M.H.; Hu, M.L. Therapeutic treatment with vitamin C reduces focal cerebral ischemia-induced brain infarction in rats by attenuating disruptions of blood brain barrier and cerebral neuronal apoptosis. Free Radic. Biol. Med. 2020, 155, 29–36. [Google Scholar] [CrossRef]
- Zhang, X.; Fan, Z.; Jin, T. Crocin protects against cerebral- ischemia-induced damage in aged rats through maintaining the integrity of blood-brain barrier. Restor. Neurol. Neurosci. 2017, 35, 65–75. [Google Scholar] [CrossRef]
- Yang, J.; Huang, J.; Shen, C.; Cheng, W.; Yu, P.; Wang, L.; Tang, F.; Guo, S.; Yang, Q.; Zhang, J. Resveratrol Treatment in Different Time-Attenuated Neuronal Apoptosis After Oxygen and Glucose Deprivation/Reoxygenation via Enhancing the Activation of Nrf-2 Signaling Pathway In Vitro. Cell Transplant. 2018, 27, 1789–1797. [Google Scholar] [CrossRef]
- Zhou, X.; Qi, Y.; Chen, T. Long-term pre-treatment of antioxidant Ginkgo biloba extract EGb-761 attenuates cerebral-ischemia-induced neuronal damage in aged mice. Biomed. Pharmacother. 2017, 85, 256–263. [Google Scholar] [CrossRef]
- Nai, Y.; Liu, H.; Bi, X.; Gao, H.; Ren, C. Protective effect of astaxanthin on acute cerebral infarction in rats. Hum. Exp. Toxicol. 2018, 37, 929–936. [Google Scholar] [CrossRef]
- Li, X.; Zhan, J.; Hou, Y.; Chen, S.; Hou, Y.; Xiao, Z.; Luo, D.; Lin, D. Coenzyme Q10 suppresses oxidative stress and apoptosis via activating the Nrf-2/NQO-1 and NF-kappaB signaling pathway after spinal cord injury in rats. Am. J. Transl. Res. 2019, 11, 6544–6552. [Google Scholar]
- Cordero, K.; Coronel, G.G.; Serrano-Illan, M.; Cruz-Bracero, J.; Figueroa, J.D.; De Leon, M. Effects of Dietary Vitamin E Supplementation in Bladder Function and Spasticity during Spinal Cord Injury. Brain Sci 2018, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Wang, H.; Liu, Z.; Liu, Y.; Wang, R.; Luo, X.; Huang, Y. Neuroprotective effects of lycopene in spinal cord injury in rats via antioxidative and anti-apoptotic pathway. Neurosci. Lett. 2017, 642, 107–112. [Google Scholar] [CrossRef]
- Zhou, L.; Ouyang, L.; Lin, S.; Chen, S.; Liu, Y.; Zhou, W.; Wang, X. Protective role of beta-carotene against oxidative stress and neuroinflammation in a rat model of spinal cord injury. Int. Immunopharmacol. 2018, 61, 92–99. [Google Scholar] [CrossRef]
- Xi, J.; Luo, X.; Wang, Y.; Li, J.; Guo, L.; Wu, G.; Li, Q. Tetrahydrocurcumin protects against spinal cord injury and inhibits the oxidative stress response by regulating FOXO4 in model rats. Exp. Ther. Med. 2019, 18, 3681–3687. [Google Scholar] [CrossRef] [Green Version]
- Machova Urdzikova, L.; Ruzicka, J.; Karova, K.; Kloudova, A.; Svobodova, B.; Amin, A.; Dubisova, J.; Schmidt, M.; Kubinova, S.; Jhanwar-Uniyal, M.; et al. A green tea polyphenol epigallocatechin-3-gallate enhances neuroregeneration after spinal cord injury by altering levels of inflammatory cytokines. Neuropharmacology 2017, 126, 213–223. [Google Scholar] [CrossRef]
- Masoudi, A.; Jorjani, M.; Alizadeh, M.; Mirzamohammadi, S.; Mohammadi, M. Anti-inflammatory and antioxidant effects of astaxanthin following spinal cord injury in a rat animal model. Brain Res. Bull. 2021, 177, 324–331. [Google Scholar] [CrossRef]
- Senturk, S.; Yaman, M.E.; Aydin, H.E.; Guney, G.; Bozkurt, I.; Paksoy, K.; Abdioglu, A.A. Effects of Resveratrol on Inflammation and Apoptosis After Experimental Spinal Cord Injury. Turk. Neurosurg. 2018, 28, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Botchway, B.O.A.; Tan, X.; Zhang, Y.; Fang, M. Resveratrol treatment of spinal cord injury in rat model. Microsc. Res. Tech. 2019, 82, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Mei, X.; Yang, D.; Tu, G. Resveratrol inhibits inflammation after spinal cord injury via SIRT-1/NF-kappaB signaling pathway. Neurosci. Lett. 2021, 762, 136151. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.Y.; Shao, D.C.; Li, H.; Huang, X.D.; Yang, G.; Xu, B.; Niu, H.Y. Resveratrol improves neurological outcome and neuroinflammation following spinal cord injury through enhancing autophagy involving the AMPK/mTOR pathway. Mol. Med. Rep. 2018, 18, 2237–2244. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Chen, S.; Gao, K.; Zhou, Z.; Wang, C.; Shen, Z.; Guo, Y.; Li, Z.; Wan, Z.; Liu, C.; et al. Resveratrol protects against spinal cord injury by activating autophagy and inhibiting apoptosis mediated by the SIRT1/AMPK signaling pathway. Neuroscience 2017, 348, 241–251. [Google Scholar] [CrossRef]
- Wang, Y.; Li, W.; Wang, M.; Lin, C.; Li, G.; Zhou, X.; Luo, J.; Jin, D. Quercetin reduces neural tissue damage and promotes astrocyte activation after spinal cord injury in rats. J. Cell Biochem. 2018, 119, 2298–2306. [Google Scholar] [CrossRef]
- Fan, H.; Tang, H.B.; Shan, L.Q.; Liu, S.C.; Huang, D.G.; Chen, X.; Chen, Z.; Yang, M.; Yin, X.H.; Yang, H.; et al. Quercetin prevents necroptosis of oligodendrocytes by inhibiting macrophages/microglia polarization to M1 phenotype after spinal cord injury in rats. J. Neuroinflamm. 2019, 16, 206. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Yang, Q.; Ma, X. Synergistic effect of ascorbic acid and taurine in the treatment of a spinal cord injury-induced model in rats. 3Biotech 2020, 10, 50. [Google Scholar] [CrossRef]
- Baum, L.; Lam, C.W.; Cheung, S.K.; Kwok, T.; Lui, V.; Tsoh, J.; Lam, L.; Leung, V.; Hui, E.; Ng, C.; et al. Six-month randomized, placebo-controlled, double-blind, pilot clinical trial of curcumin in patients with Alzheimer disease. J. Clin. Psychopharmacol. 2008, 28, 110–113. [Google Scholar] [CrossRef] [Green Version]
- Chico, L.; Ienco, E.C.; Bisordi, C.; Lo Gerfo, A.; Petrozzi, L.; Petrucci, A.; Mancuso, M.; Siciliano, G. Amyotrophic Lateral Sclerosis and Oxidative Stress: A Double-Blind Therapeutic Trial after Curcumin Supplementation. CNS Neurol. Disord. Drug Targets 2018, 17, 767–779. [Google Scholar] [CrossRef]
- Moussa, C.; Hebron, M.; Huang, X.; Ahn, J.; Rissman, R.A.; Aisen, P.S.; Turner, R.S. Resveratrol regulates neuro-inflammation and induces adaptive immunity in Alzheimer’s disease. J. Neuroinflamm. 2017, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Bai, Q.; Zhao, Z.; Sui, H.; Xie, X. Resveratrol improves delayed r-tPA treatment outcome by reducing MMPs. Acta Neurol. Scand. 2016, 134, 54–60. [Google Scholar] [CrossRef]
- Wang, X.H.; You, Y.P. Epigallocatechin Gallate Extends Therapeutic Window of Recombinant Tissue Plasminogen Activator Treatment for Brain Ischemic Stroke: A Randomized Double-Blind and Placebo-Controlled Trial. Clin. Neuropharmacol. 2017, 40, 24–28. [Google Scholar] [CrossRef]
- Mischley, L.K.; Leverenz, J.B.; Lau, R.C.; Polissar, N.L.; Neradilek, M.B.; Samii, A.; Standish, L.J. A randomized, double-blind phase I/IIa study of intranasal glutathione in Parkinson’s disease. Mov. Disord. 2015, 30, 1696–1701. [Google Scholar] [CrossRef]
- Mischley, L.K.; Lau, R.C.; Shankland, E.G.; Wilbur, T.K.; Padowski, J.M. Phase IIb Study of Intranasal Glutathione in Parkinson’s Disease. J. Parkinsons Dis. 2017, 7, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Seet, R.C.; Lim, E.C.; Tan, J.J.; Quek, A.M.; Chow, A.W.; Chong, W.L.; Ng, M.P.; Ong, C.N.; Halliwell, B. Does high-dose coenzyme Q10 improve oxidative damage and clinical outcomes in Parkinson’s disease? Antioxid. Redox. Signal. 2014, 21, 211–217. [Google Scholar] [CrossRef]
- Parkinson Study Group, Q.E.I.; Beal, M.F.; Oakes, D.; Shoulson, I.; Henchcliffe, C.; Galpern, W.R.; Haas, R.; Juncos, J.L.; Nutt, J.G.; Voss, T.S.; et al. A randomized clinical trial of high-dosage coenzyme Q10 in early Parkinson disease: No evidence of benefit. JAMA Neurol. 2014, 71, 543–552. [Google Scholar] [CrossRef]
- Kaufmann, P.; Thompson, J.L.; Levy, G.; Buchsbaum, R.; Shefner, J.; Krivickas, L.S.; Katz, J.; Rollins, Y.; Barohn, R.J.; Jackson, C.E.; et al. Phase II trial of CoQ10 for ALS finds insufficient evidence to justify phase III. Ann. Neurol. 2009, 66, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Snitz, B.E.; O’Meara, E.S.; Carlson, M.C.; Arnold, A.M.; Ives, D.G.; Rapp, S.R.; Saxton, J.; Lopez, O.L.; Dunn, L.O.; Sink, K.M.; et al. Ginkgo biloba for preventing cognitive decline in older adults: A randomized trial. JAMA 2009, 302, 2663–2670. [Google Scholar] [CrossRef] [Green Version]
- Writing, G.; Edaravone, A.L.S.S.G. Safety and efficacy of edaravone in well defined patients with amyotrophic lateral sclerosis: A randomised, double-blind, placebo-controlled trial. Lancet Neurol. 2017, 16, 505–512. [Google Scholar] [CrossRef]
- Takei, K.; Takahashi, F.; Liu, S.; Tsuda, K.; Palumbo, J. Post-hoc analysis of randomised, placebo-controlled, double-blind study (MCI186-19) of edaravone (MCI-186) in amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Frontotemporal Degener. 2017, 18, 49–54. [Google Scholar] [CrossRef]
- Shinohara, Y.; Saito, I.; Kobayashi, S.; Uchiyama, S. Edaravone (radical scavenger) versus sodium ozagrel (antiplatelet agent) in acute noncardioembolic ischemic stroke (EDO trial). Cerebrovasc. Dis. 2009, 27, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Isahaya, K.; Yamada, K.; Yamatoku, M.; Sakurai, K.; Takaishi, S.; Kato, B.; Hirayama, T.; Hasegawa, Y. Effects of edaravone, a free radical scavenger, on serum levels of inflammatory biomarkers in acute brain infarction. J. Stroke Cerebrovasc. Dis. 2012, 21, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, Y.; Wang, A.; Gao, Z.; Gao, X.; Chen, H.; Zhou, J.; Zhao, X.; Wang, Y. Safety and efficacy of Edaravone Dexborneol versus edaravone for patients with acute ischaemic stroke: A phase II, multicentre, randomised, double-blind, multiple-dose, active-controlled clinical trial. Stroke Vasc. Neurol. 2019, 4, 109–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NCT04714177; Edaravone Dexborneol for Treatment of Hypertensive Intracerebral Hemorrhage (ED-ICH). US National Library of Medicine: Bethesda, MD, USA, 2021.
- Shinto, L.; Quinn, J.; Montine, T.; Dodge, H.H.; Woodward, W.; Baldauf-Wagner, S.; Waichunas, D.; Bumgarner, L.; Bourdette, D.; Silbert, L.; et al. A randomized placebo-controlled pilot trial of omega-3 fatty acids and alpha lipoic acid in Alzheimer’s disease. J. Alzheimers Dis. 2014, 38, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dysken, M.W.; Sano, M.; Asthana, S.; Vertrees, J.E.; Pallaki, M.; Llorente, M.; Love, S.; Schellenberg, G.D.; McCarten, J.R.; Malphurs, J.; et al. Effect of vitamin E and memantine on functional decline in Alzheimer disease: The TEAM-AD VA cooperative randomized trial. JAMA 2014, 311, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Galasko, D.R.; Peskind, E.; Clark, C.M.; Quinn, J.F.; Ringman, J.M.; Jicha, G.A.; Cotman, C.; Cottrell, B.; Montine, T.J.; Thomas, R.G.; et al. Antioxidants for Alzheimer disease: A randomized clinical trial with cerebrospinal fluid biomarker measures. Arch. Neurol. 2012, 69, 836–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taghizadeh, M.; Tamtaji, O.R.; Dadgostar, E.; Daneshvar Kakhaki, R.; Bahmani, F.; Abolhassani, J.; Aarabi, M.H.; Kouchaki, E.; Memarzadeh, M.R.; Asemi, Z. The effects of omega-3 fatty acids and vitamin E co-supplementation on clinical and metabolic status in patients with Parkinson’s disease: A randomized, double-blind, placebo-controlled trial. Neurochem. Int. 2017, 108, 183–189. [Google Scholar] [CrossRef]
- Ahmadi, M.; Agah, E.; Nafissi, S.; Jaafari, M.R.; Harirchian, M.H.; Sarraf, P.; Faghihi-Kashani, S.; Hosseini, S.J.; Ghoreishi, A.; Aghamollaii, V.; et al. Safety and Efficacy of Nanocurcumin as Add-On Therapy to Riluzole in Patients With Amyotrophic Lateral Sclerosis: A Pilot Randomized Clinical Trial. Neurotherapeutics 2018, 15, 430–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, K.H.; Pipingas, A.; Scholey, A.B. Investigation of the effects of solid lipid curcumin on cognition and mood in a healthy older population. J. Psychopharmacol. 2015, 29, 642–651. [Google Scholar] [CrossRef]
- NCT01001637; Efficacy and Safety of Curcumin Formulation in Alzheimer’s Disease. US National Library of Medicine: Bethesda, MD, USA, 2009.
- Dang, L.; Dong, X.; Yang, J. Influence of Nanoparticle-Loaded Edaravone on Postoperative Effects in Patients with Cerebral Hemorrhage. J. Nanosci. Nanotechnol. 2021, 21, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, X.; Fang, Q.; Zhou, J.; Zhang, M.; Wang, H.; Chen, Y.; Xu, B.; Wu, Y.; Qian, L.; et al. Ginkgo biloba extract improved cognitive and neurological functions of acute ischaemic stroke: A randomised controlled trial. Stroke Vasc. Neurol. 2017, 2, 189–197. [Google Scholar] [CrossRef]
- Allison, D.J.; Ditor, D.S. Targeting inflammation to influence mood following spinal cord injury: A randomized clinical trial. J. Neuroinflamm. 2015, 12, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breiner, A.; Zinman, L.; Bourque, P.R. Edaravone for amyotrophic lateral sclerosis: Barriers to access and lifeboat ethics. CMAJ 2020, 192, E319–E320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Medicine Agency. Radicava: Withdrawal of the Marketing Authorisation Application; European Medicine Agency: Amsterdam, The Netherlands, 2019; Available online: https://www.ema.europa.eu/en/medicines/human/withdrawn-applications/radicava (accessed on 14 January 2022).
- Panzarini, E.; Mariano, S.; Tacconi, S.; Carata, E.; Tata, A.M.; Dini, L. Novel Therapeutic Delivery of Nanocurcumin in Central Nervous System Related Disorders. Nanomaterials 2020, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Dong, X. Current Strategies for Brain Drug Delivery. Theranostics 2018, 8, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Ringman, J.M.; Frautschy, S.A.; Teng, E.; Begum, A.N.; Bardens, J.; Beigi, M.; Gylys, K.H.; Badmaev, V.; Heath, D.D.; Apostolova, L.G.; et al. Oral curcumin for Alzheimer’s disease: Tolerability and efficacy in a 24-week randomized, double blind, placebo-controlled study. Alzheimers Res. Ther. 2012, 4, 43. [Google Scholar] [CrossRef] [Green Version]
- Kokjohn, T.A.; Roher, A.E. Amyloid precursor protein transgenic mouse models and Alzheimer’s disease: Understanding the paradigms, limitations, and contributions. Alzheimers Dement. 2009, 5, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Schwedhelm, E.; Maas, R.; Troost, R.; Boger, R.H. Clinical pharmacokinetics of antioxidants and their impact on systemic oxidative stress. Clin. Pharmacokinet. 2003, 42, 437–459. [Google Scholar] [CrossRef]
- Bast, A.; Haenen, G.R. The toxicity of antioxidants and their metabolites. Environ. Toxicol. Pharmacol. 2002, 11, 251–258. [Google Scholar] [CrossRef]
- Nimse, D.P. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Van Norman, G.A. Limitations of Animal Studies for Predicting Toxicity in Clinical Trials: Is it Time to Rethink Our Current Approach? JACC Basic Transl. Sci. 2019, 4, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Jurcau, A. The Role of Natural Antioxidants in the Prevention of Dementia-Where Do We Stand and Future Perspectives. Nutrients 2021, 13, 282. [Google Scholar] [CrossRef] [PubMed]
- van der Worp, H.B.; Howells, D.W.; Sena, E.S.; Porritt, M.J.; Rewell, S.; O’Collins, V.; Macleod, M.R. Can animal models of disease reliably inform human studies? PLoS Med. 2010, 7, e1000245. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Al-Jamal, K.T.; Kostarelos, K.; Reineke, J. Physiologically based pharmacokinetic modeling of nanoparticles. ACS Nano 2010, 4, 6303–6317. [Google Scholar] [CrossRef]
- De Matteis, V.; Rinaldi, R. Toxicity Assessment in the Nanoparticle Era. Adv. Exp. Med. Biol. 2018, 1048, 1–19. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Senthil, N.; Min, T. Nanocurcumin: A Promising Candidate for Therapeutic Applications. Front. Pharmacol. 2020, 11, 487. [Google Scholar] [CrossRef]
- Fan, S.; Zheng, Y.; Liu, X.; Fang, W.; Chen, X.; Liao, W.; Jing, X.; Lei, M.; Tao, E.; Ma, Q.; et al. Curcumin-loaded PLGA-PEG nanoparticles conjugated with B6 peptide for potential use in Alzheimer’s disease. Drug Deliv. 2018, 25, 1091–1102. [Google Scholar] [CrossRef] [Green Version]
- Fidelis, E.M.; Savall, A.S.P.; da Luz Abreu, E.; Carvalho, F.; Teixeira, F.E.G.; Haas, S.E.; Bazanella Sampaio, T.; Pinton, S. Curcumin-Loaded Nanocapsules Reverses the Depressant-Like Behavior and Oxidative Stress Induced by beta-Amyloid in Mice. Neuroscience 2019, 423, 122–130. [Google Scholar] [CrossRef]
- Huo, X.; Zhang, Y.; Jin, X.; Li, Y.; Zhang, L. A novel synthesis of selenium nanoparticles encapsulated PLGA nanospheres with curcumin molecules for the inhibition of amyloid beta aggregation in Alzheimer’s disease. J. Photochem. Photobiol. B 2019, 190, 98–102. [Google Scholar] [CrossRef]
- Ashafaq, M.; Intakhab Alam, M.; Khan, A.; Islam, F.; Khuwaja, G.; Hussain, S.; Ali, R.; Alshahrani, S.; Antar Makeen, H.; Alhazmi, H.A.; et al. Nanoparticles of resveratrol attenuates oxidative stress and inflammation after ischemic stroke in rats. Int. Immunopharmacol. 2021, 94, 107494. [Google Scholar] [CrossRef]
- Lu, X.; Dong, J.; Zheng, D.; Li, X.; Ding, D.; Xu, H. Reperfusion combined with intraarterial administration of resveratrol-loaded nanoparticles improved cerebral ischemia-reperfusion injury in rats. Nanomedicine 2020, 28, 102208. [Google Scholar] [CrossRef]
- Kannan, S.; Dai, H.; Navath, R.S.; Balakrishnan, B.; Jyoti, A.; Janisse, J.; Romero, R.; Kannan, R.M. Dendrimer-based postnatal therapy for neuroinflammation and cerebral palsy in a rabbit model. Sci. Transl. Med. 2012, 4, 130ra46. [Google Scholar] [CrossRef] [Green Version]
- Tardiolo, G.; Bramanti, P.; Mazzon, E. Overview on the Effects of N-Acetylcysteine in Neurodegenerative Diseases. Molecules 2018, 23, 3305. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Jin, L.; Wu, Z.; Xie, Y.; Zhang, P.; Wang, Q.; Yan, S.; Chen, B.; Liang, H.; Naman, C.B.; et al. PLGA-PEG Nanoparticles Facilitate In Vivo Anti-Alzheimer’s Effects of Fucoxanthin, a Marine Carotenoid Derived from Edible Brown Algae. J. Agric. Food Chem. 2021, 69, 9764–9777. [Google Scholar] [CrossRef]
- Dhas, N.; Mehta, T. Cationic biopolymer functionalized nanoparticles encapsulating lutein to attenuate oxidative stress in effective treatment of Alzheimer’s disease: A non-invasive approach. Int. J. Pharm. 2020, 586, 119553. [Google Scholar] [CrossRef]
- Singh, N.A.; Mandal, A.K.A.; Khan, Z.A. Inhibition of Al(III)-Induced Abeta42 Fibrillation and Reduction of Neurotoxicity by Epigallocatechin-3-Gallate Nanoparticles. J. Biomed. Nanotechnol. 2018, 14, 1147–1158. [Google Scholar] [CrossRef]
- Liu, Z.; Li, X.; Wu, X.; Zhu, C. A dual-inhibitor system for the effective antifibrillation of Abeta40 peptides by biodegradable EGCG-Fe(iii)/PVP nanoparticles. J. Mater. Chem. B 2019, 7, 1292–1299. [Google Scholar] [CrossRef]
- Liu, H.; Yu, L.; Dong, X.; Sun, Y. Synergistic effects of negatively charged hydrophobic nanoparticles and (-)-epigallocatechin-3-gallate on inhibiting amyloid beta-protein aggregation. J. Colloid Interface Sci. 2017, 491, 305–312. [Google Scholar] [CrossRef]
- Cano, A.; Ettcheto, M.; Chang, J.H.; Barroso, E.; Espina, M.; Kuhne, B.A.; Barenys, M.; Auladell, C.; Folch, J.; Souto, E.B.; et al. Dual-drug loaded nanoparticles of Epigallocatechin-3-gallate (EGCG)/Ascorbic acid enhance therapeutic efficacy of EGCG in a APPswe/PS1dE9 Alzheimer’s disease mice model. J. Control. Release 2019, 301, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Amin, F.U.; Shah, S.A.; Badshah, H.; Khan, M.; Kim, M.O. Anthocyanins encapsulated by PLGA@PEG nanoparticles potentially improved its free radical scavenging capabilities via p38/JNK pathway against Abeta1-42-induced oxidative stress. J. Nanobiotechnology 2017, 15, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loureiro, J.A.; Andrade, S.; Duarte, A.; Neves, A.R.; Queiroz, J.F.; Nunes, C.; Sevin, E.; Fenart, L.; Gosselet, F.; Coelho, M.A.; et al. Resveratrol and Grape Extract-loaded Solid Lipid Nanoparticles for the Treatment of Alzheimer’s Disease. Molecules 2017, 22, 277. [Google Scholar] [CrossRef] [PubMed]
- Amanzadeh Jajin, E.; Esmaeili, A.; Rahgozar, S.; Noorbakhshnia, M. Quercetin-Conjugated Superparamagnetic Iron Oxide Nanoparticles Protect AlCl3-Induced Neurotoxicity in a Rat Model of Alzheimer’s Disease via Antioxidant Genes, APP Gene, and miRNA-101. Front. Neurosci. 2020, 14, 598617. [Google Scholar] [CrossRef]
- Halevas, E.; Mavroidi, B.; Nday, C.M.; Tang, J.; Smith, G.C.; Boukos, N.; Litsardakis, G.; Pelecanou, M.; Salifoglou, A. Modified magnetic core-shell mesoporous silica nano-formulations with encapsulated quercetin exhibit anti-amyloid and antioxidant activity. J. Inorg. Biochem. 2020, 213, 111271. [Google Scholar] [CrossRef]
- Pinheiro, R.G.R.; Granja, A.; Loureiro, J.A.; Pereira, M.C.; Pinheiro, M.; Neves, A.R.; Reis, S. Quercetin lipid nanoparticles functionalized with transferrin for Alzheimer’s disease. Eur. J. Pharm. Sci. 2020, 148, 105314. [Google Scholar] [CrossRef]
- Sun, D.; Li, N.; Zhang, W.; Zhao, Z.; Mou, Z.; Huang, D.; Liu, J.; Wang, W. Design of PLGA-functionalized quercetin nanoparticles for potential use in Alzheimer’s disease. Colloids Surf. B Biointerfaces 2016, 148, 116–129. [Google Scholar] [CrossRef]
- Rifaai, R.A.; Mokhemer, S.A.; Saber, E.A.; El-Aleem, S.A.A.; El-Tahawy, N.F.G. Neuroprotective effect of quercetin nanoparticles: A possible prophylactic and therapeutic role in alzheimer’s disease. J. Chem. Neuroanat. 2020, 107, 101795. [Google Scholar] [CrossRef]
- Moreno, L.; Puerta, E.; Suarez-Santiago, J.E.; Santos-Magalhaes, N.S.; Ramirez, M.J.; Irache, J.M. Effect of the oral administration of nanoencapsulated quercetin on a mouse model of Alzheimer’s disease. Int. J. Pharm. 2017, 517, 50–57. [Google Scholar] [CrossRef]
- Ramires Junior, O.V.; Alves, B.D.S.; Barros, P.A.B.; Rodrigues, J.L.; Ferreira, S.P.; Monteiro, L.K.S.; Araujo, G.M.S.; Fernandes, S.S.; Vaz, G.R.; Dora, C.L.; et al. Nanoemulsion Improves the Neuroprotective Effects of Curcumin in an Experimental Model of Parkinson’s Disease. Neurotox. Res. 2021, 39, 787–799. [Google Scholar] [CrossRef]
- Kundu, P.; Das, M.; Tripathy, K.; Sahoo, S.K. Delivery of Dual Drug Loaded Lipid Based Nanoparticles across the Blood-Brain Barrier Impart Enhanced Neuroprotection in a Rotenone Induced Mouse Model of Parkinson’s Disease. ACS Chem. Neurosci. 2016, 7, 1658–1670. [Google Scholar] [CrossRef]
- Fernandes, E.J.; Poetini, M.R.; Barrientos, M.S.; Bortolotto, V.C.; Araujo, S.M.; Santos Musachio, E.A.; De Carvalho, A.S.; Leimann, F.V.; Goncalves, O.H.; Ramborger, B.P.; et al. Exposure to lutein-loaded nanoparticles attenuates Parkinson’s model-induced damage in Drosophila melanogaster: Restoration of dopaminergic and cholinergic system and oxidative stress indicators. Chem. Biol. Interact. 2021, 340, 109431. [Google Scholar] [CrossRef]
- Palle, S.; Neerati, P. Improved neuroprotective effect of resveratrol nanoparticles as evinced by abrogation of rotenone-induced behavioral deficits and oxidative and mitochondrial dysfunctions in rat model of Parkinson’s disease. Naunyn. Schmiedebergs Arch. Pharmacol. 2018, 391, 445–453. [Google Scholar] [CrossRef]
- Gaba, B.; Khan, T.; Haider, M.F.; Alam, T.; Baboota, S.; Parvez, S.; Ali, J. Vitamin E Loaded Naringenin Nanoemulsion via Intranasal Delivery for the Management of Oxidative Stress in a 6-OHDA Parkinson’s Disease Model. Biomed. Res. Int. 2019, 2019, 2382563. [Google Scholar] [CrossRef]
- Trapani, A.; Guerra, L.; Corbo, F.; Castellani, S.; Sanna, E.; Capobianco, L.; Monteduro, A.G.; Manno, D.E.; Mandracchia, D.; Di Gioia, S.; et al. Cyto/Biocompatibility of Dopamine Combined with the Antioxidant Grape Seed-Derived Polyphenol Compounds in Solid Lipid Nanoparticles. Molecules 2021, 26, 916. [Google Scholar] [CrossRef]
- Chen, L.; Watson, C.; Morsch, M.; Cole, N.J.; Chung, R.S.; Saunders, D.N.; Yerbury, J.J.; Vine, K.L. Improving the Delivery of SOD1 Antisense Oligonucleotides to Motor Neurons Using Calcium Phosphate-Lipid Nanoparticles. Front. Neurosci. 2017, 11, 476. [Google Scholar] [CrossRef]
- Medina, D.X.; Chung, E.P.; Teague, C.D.; Bowser, R.; Sirianni, R.W. Intravenously Administered, Retinoid Activating Nanoparticles Increase Lifespan and Reduce Neurodegeneration in the SOD1(G93A) Mouse Model of ALS. Front. Bioeng. Biotechnol. 2020, 8, 224. [Google Scholar] [CrossRef]
- Mauricio, M.D.; Guerra-Ojeda, S.; Marchio, P.; Valles, S.L.; Aldasoro, M.; Escribano-Lopez, I.; Herance, J.R.; Rocha, M.; Vila, J.M.; Victor, V.M. Nanoparticles in Medicine: A Focus on Vascular Oxidative Stress. Oxid. Med. Cell Longev. 2018, 2018, 6231482. [Google Scholar] [CrossRef] [Green Version]
- Mei, T.; Kim, A.; Vong, L.B.; Marushima, A.; Puentes, S.; Matsumaru, Y.; Matsumura, A.; Nagasaki, Y. Encapsulation of tissue plasminogen activator in pH-sensitive self-assembled antioxidant nanoparticles for ischemic stroke treatment—Synergistic effect of thrombolysis and antioxidant. Biomaterials 2019, 215, 119209. [Google Scholar] [CrossRef]
- Marques, M.S.; Cordeiro, M.F.; Marinho, M.A.G.; Vian, C.O.; Vaz, G.R.; Alves, B.S.; Jardim, R.D.; Hort, M.A.; Dora, C.L.; Horn, A.P. Curcumin-loaded nanoemulsion improves haemorrhagic stroke recovery in wistar rats. Brain Res. 2020, 1746, 147007. [Google Scholar] [CrossRef]
- Chen, W.; Zhao, Z.; Zhao, S.; Zhang, L.; Song, Q. Resveratrol and Puerarin loaded polymeric nanoparticles to enhance the chemotherapeutic efficacy in spinal cord injury. Biomed. Microdevices 2020, 22, 69. [Google Scholar] [CrossRef] [PubMed]
- Luan, X.; Sansanaphongpricha, K.; Myers, I.; Chen, H.; Yuan, H.; Sun, D. Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacol. Sin. 2017, 38, 754–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, Q.; Hu, P.; Xu, Y.; Cheng, T.; Wei, C.; Pan, L.; Shi, J. Simultaneous Blood-Brain Barrier Crossing and Protection for Stroke Treatment Based on Edaravone-Loaded Ceria Nanoparticles. ACS Nano 2018, 12, 6794–6805. [Google Scholar] [CrossRef]
- Jin, Q.; Cai, Y.; Li, S.; Liu, H.; Zhou, X.; Lu, C.; Gao, X.; Qian, J.; Zhang, J.; Ju, S.; et al. Edaravone-Encapsulated Agonistic Micelles Rescue Ischemic Brain Tissue by Tuning Blood-Brain Barrier Permeability. Theranostics 2017, 7, 884–898. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.Y.; Rayner, S.L.; Chung, R.; Shi, B.Y.; Liang, X.J. Advances in nanotechnology-based strategies for the treatments of amyotrophic lateral sclerosis. Mater. Today Bio 2020, 6, 100055. [Google Scholar] [CrossRef] [PubMed]
- Alkadi, H. A Review on Free Radicals and Antioxidants. Infect. Disord. Drug Targets 2020, 20, 16–26. [Google Scholar] [CrossRef]
- Beckman, J.S.; Minor, R.L., Jr.; White, C.W.; Repine, J.E.; Rosen, G.M.; Freeman, B.A. Superoxide dismutase and catalase conjugated to polyethylene glycol increases endothelial enzyme activity and oxidant resistance. J. Biol. Chem. 1988, 263, 6884–6892. [Google Scholar] [CrossRef]
- Tsubokawa, T.; Jadhav, V.; Solaroglu, I.; Shiokawa, Y.; Konishi, Y.; Zhang, J.H. Lecithinized superoxide dismutase improves outcomes and attenuates focal cerebral ischemic injury via antiapoptotic mechanisms in rats. Stroke 2007, 38, 1057–1062. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.W.; Eum, W.S.; Jang, S.H.; Kim, S.Y.; Choi, H.S.; Choi, S.H.; An, J.J.; Lee, S.H.; Lee, K.S.; Han, K.; et al. Transduced Tat-SOD fusion protein protects against ischemic brain injury. Mol. Cells 2005, 19, 88–96. [Google Scholar]
- Veronese, F.M.; Caliceti, P.; Schiavon, O.; Sergi, M. Polyethylene glycol-superoxide dismutase, a conjugate in search of exploitation. Adv. Drug Deliv. Rev. 2002, 54, 587–606. [Google Scholar] [CrossRef]
- Morris, M.C.; Depollier, J.; Mery, J.; Heitz, F.; Divita, G. A peptide carrier for the delivery of biologically active proteins into mammalian cells. Nat. Biotech. 2001, 19, 1173–1176. [Google Scholar] [CrossRef]
- Francis, J.W.; Ren, J.; Warren, L.; Brown, R.H., Jr.; Finklestein, S.P. Postischemic infusion of Cu/Zn superoxide dismutase or SOD:Tet451 reduces cerebral infarction following focal ischemia/reperfusion in rats. Exp. Neurol. 1997, 146, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, S.; Woolworth, V.; Fishman, R.A.; Chan, P.H. Liposome-entrapped superoxide dismutase reduces cerebral infarction in cerebral ischemia in rats. Stroke 1990, 21, 1312–1317. [Google Scholar] [CrossRef] [Green Version]
- Sinha, J.; Das, N.; Basu, M.K. Liposomal antioxidants in combating ischemia-reperfusion injury in rat brain. Biomed. Pharmacother. 2001, 55, 264–271. [Google Scholar] [CrossRef]
- Klyachko, N.L.; Manickam, D.S.; Brynskikh, A.M.; Uglanova, S.V.; Li, S.; Higginbotham, S.M.; Bronich, T.K.; Batrakova, E.V.; Kabanov, A.V. Cross-linked antioxidant nanozymes for improved delivery to CNS. Nanomedicine 2012, 8, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Manickam, D.S.; Brynskikh, A.M.; Kopanic, J.L.; Sorgen, P.L.; Klyachko, N.L.; Batrakova, E.V.; Bronich, T.K.; Kabanov, A.V. Well-defined cross-linked antioxidant nanozymes for treatment of ischemic brain injury. J. Control. Release 2012, 162, 636–645. [Google Scholar] [CrossRef] [Green Version]
- Nukolova, N.V.; Aleksashkin, A.D.; Abakumova, T.O.; Morozova, A.Y.; Gubskiy, I.L.; Kirzhanova capital Ie, C.A.C.; Abakumov, M.A.; Chekhonin, V.P.; Klyachko, N.L.; Kabanov, A.V. Multilayer polyion complex nanoformulations of superoxide dismutase 1 for acute spinal cord injury. J. Control. Release 2018, 270, 226–236. [Google Scholar] [CrossRef]
- Hood, E.; Simone, E.; Wattamwar, P.; Dziubla, T.; Muzykantov, V. Nanocarriers for vascular delivery of antioxidants. Nanomedicine 2011, 6, 1257–1272. [Google Scholar] [CrossRef] [Green Version]
- Reddy, M.K.; Wu, L.; Kou, W.; Ghorpade, A.; Labhasetwar, V. Superoxide dismutase-loaded PLGA nanoparticles protect cultured human neurons under oxidative stress. Appl. Biochem. Biotechnol. 2008, 151, 565–577. [Google Scholar] [CrossRef] [Green Version]
- Singhal, A.; Morris, V.B.; Labhasetwar, V.; Ghorpade, A. Nanoparticle-mediated catalase delivery protects human neurons from oxidative stress. Cell Death Dis. 2013, 4, e903. [Google Scholar] [CrossRef] [Green Version]
- Reddy, M.K.; Labhasetwar, V. Nanoparticle-mediated delivery of superoxide dismutase to the brain: An effective strategy to reduce ischemia-reperfusion injury. FASEB J. 2009, 23, 1384–1395. [Google Scholar] [CrossRef] [PubMed]
- Petro, M.; Jaffer, H.; Yang, J.; Kabu, S.; Morris, V.B.; Labhasetwar, V. Tissue plasminogen activator followed by antioxidant-loaded nanoparticle delivery promotes activation/mobilization of progenitor cells in infarcted rat brain. Biomaterials 2016, 81, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffer, H.; Adjei, I.M.; Labhasetwar, V. Optical imaging to map blood-brain barrier leakage. Sci. Rep. 2013, 3, 3117. [Google Scholar] [CrossRef] [Green Version]
- Andrabi, S.S.; Yang, J.; Gao, Y.; Kuang, Y.; Labhasetwar, V. Nanoparticles with antioxidant enzymes protect injured spinal cord from neuronal cell apoptosis by attenuating mitochondrial dysfunction. J. Control. Release 2020, 317, 300–311. [Google Scholar] [CrossRef]
- Gao, Y.; Vijayaraghavalu, S.; Stees, M.; Kwon, B.K.; Labhasetwar, V. Evaluating accessibility of intravenously administered nanoparticles at the lesion site in rat and pig contusion models of spinal cord injury. J. Control. Release 2019, 302, 160–168. [Google Scholar] [CrossRef]
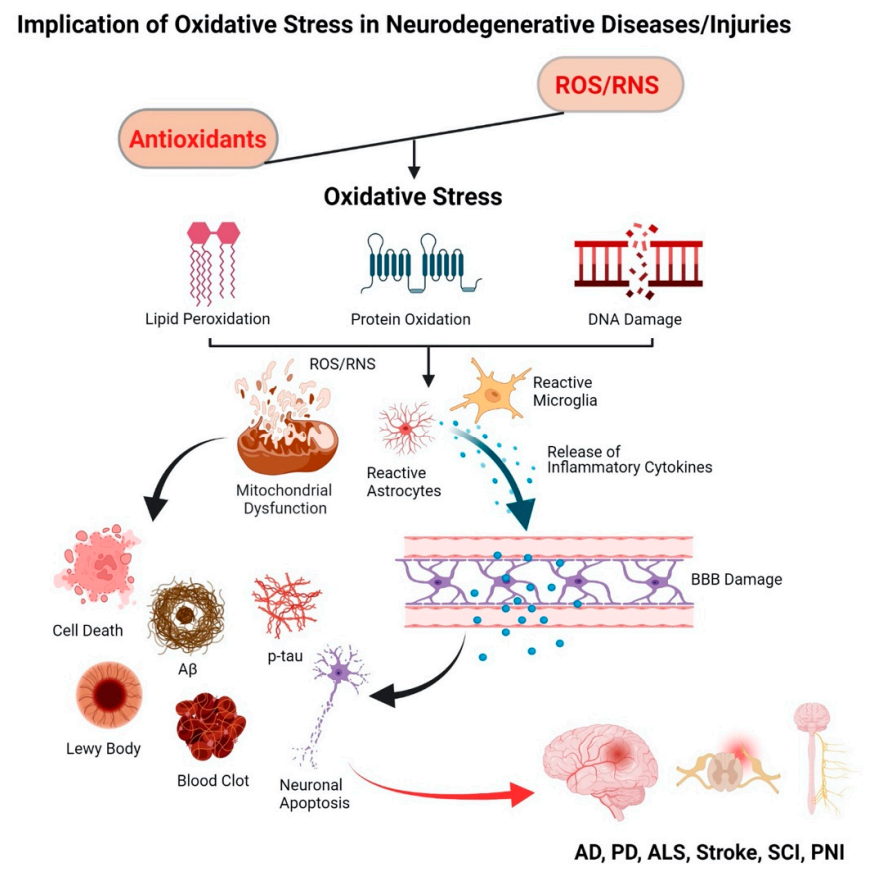
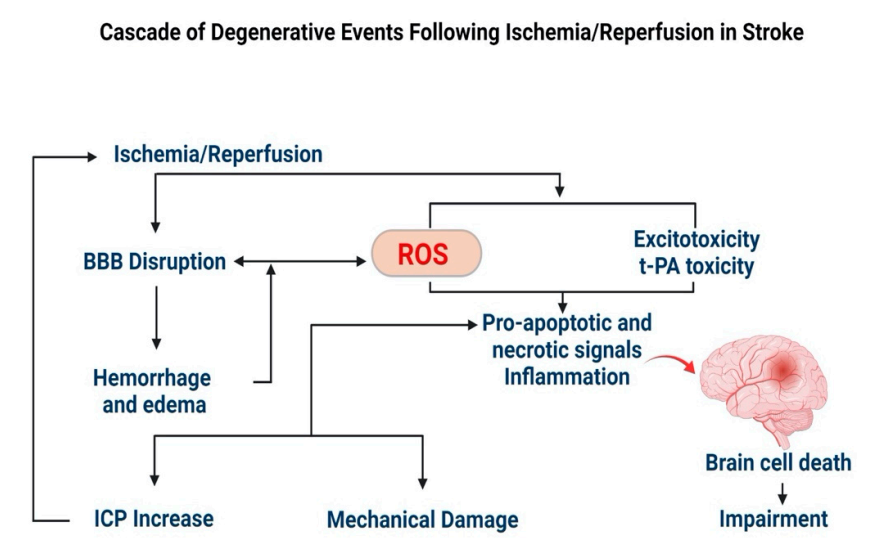
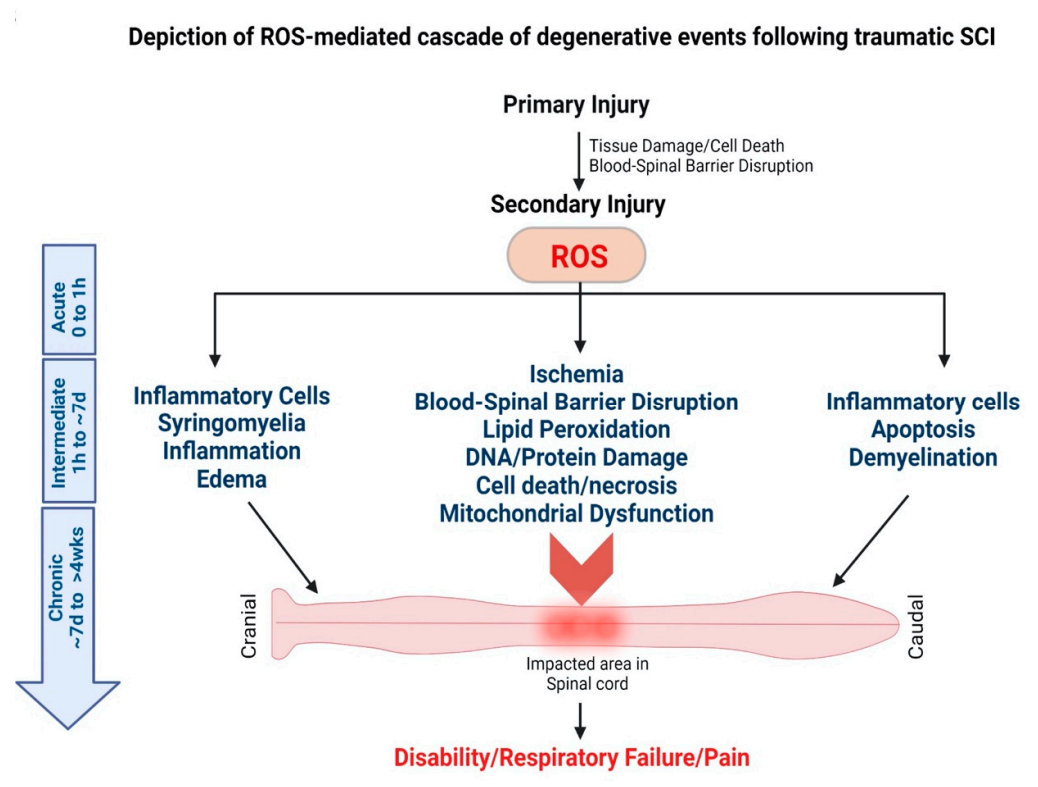
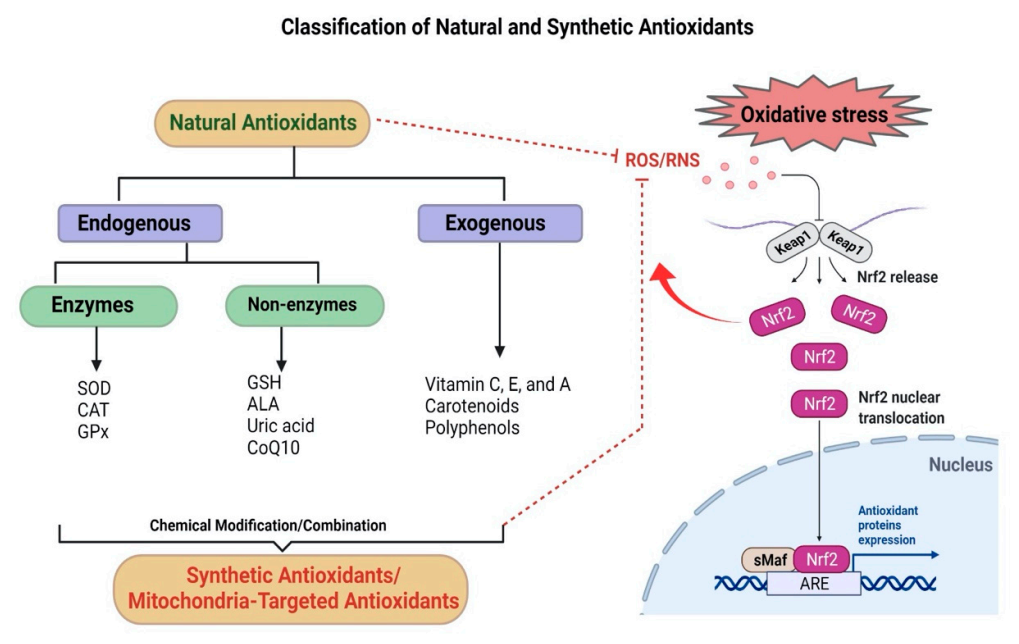
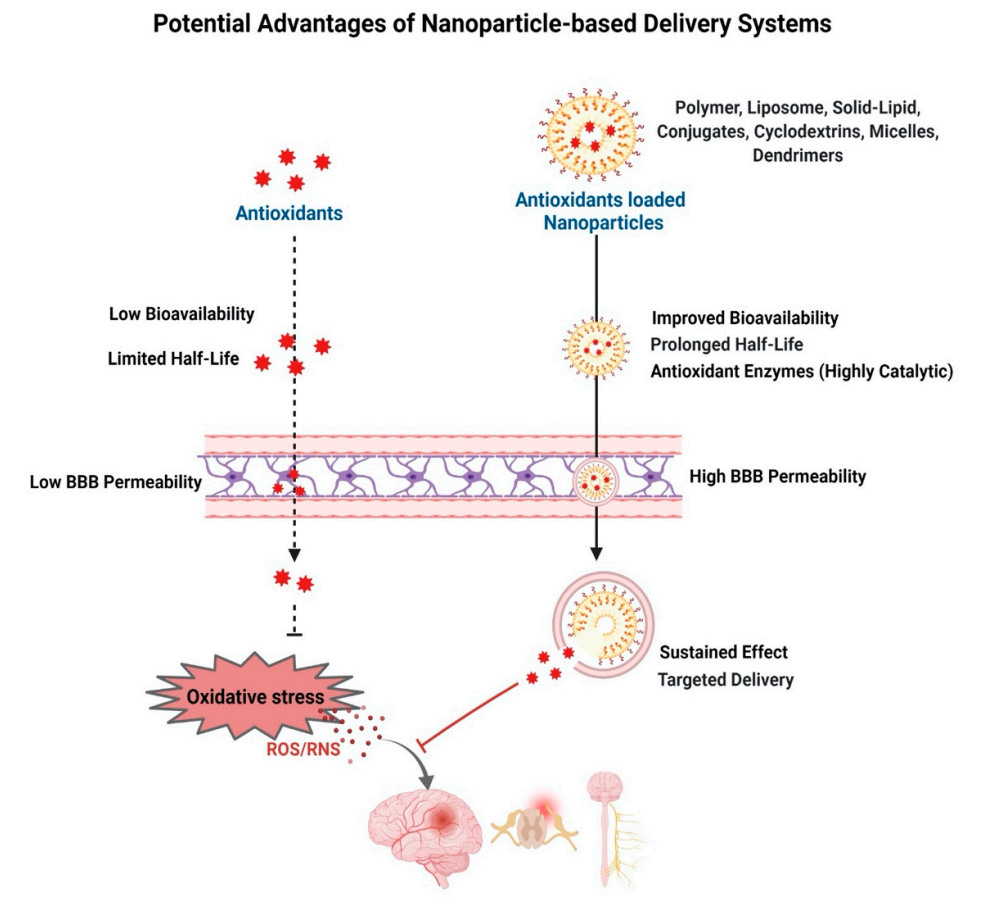
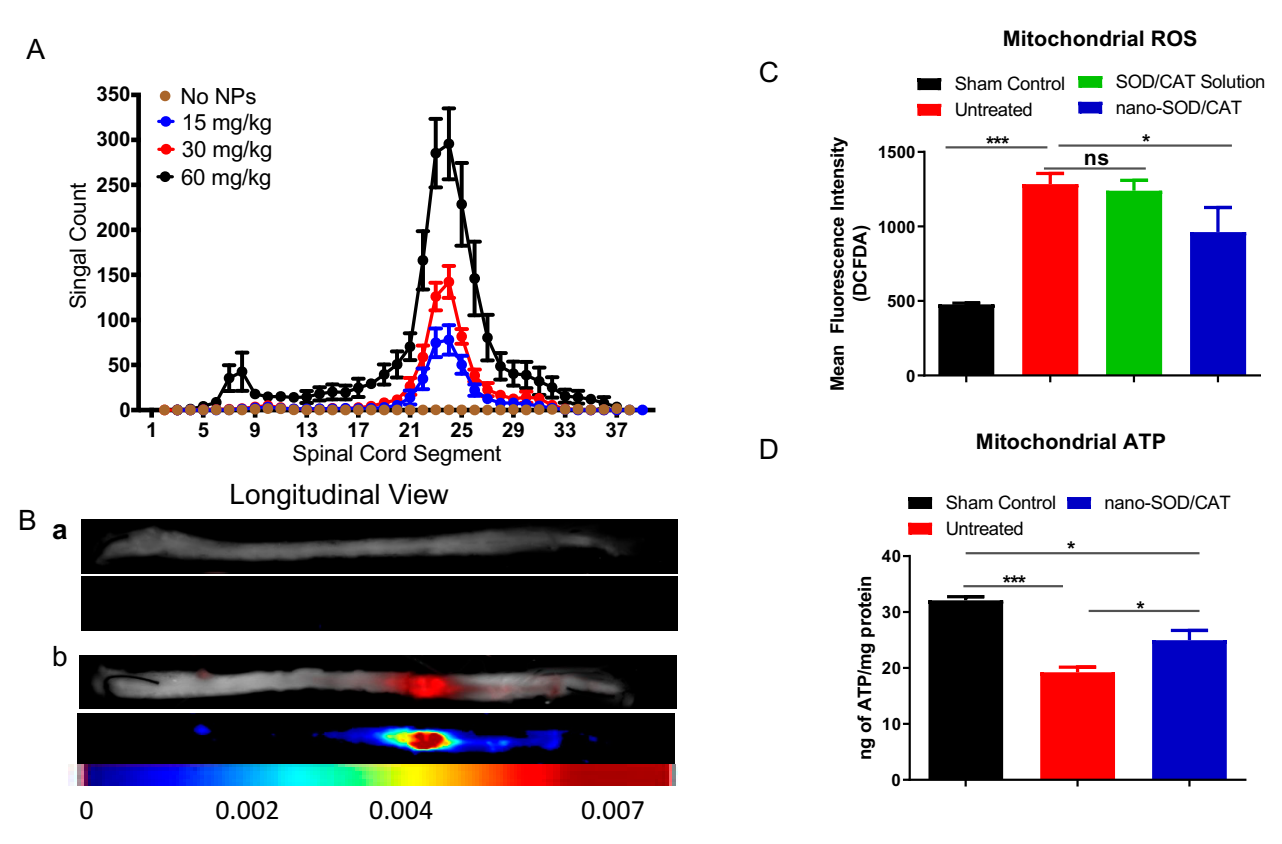
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).