Anti-Osteoporotic Mechanisms of Polyphenols Elucidated Based on In Vivo Studies Using Ovariectomized Animals
 ,
, 
Abstract
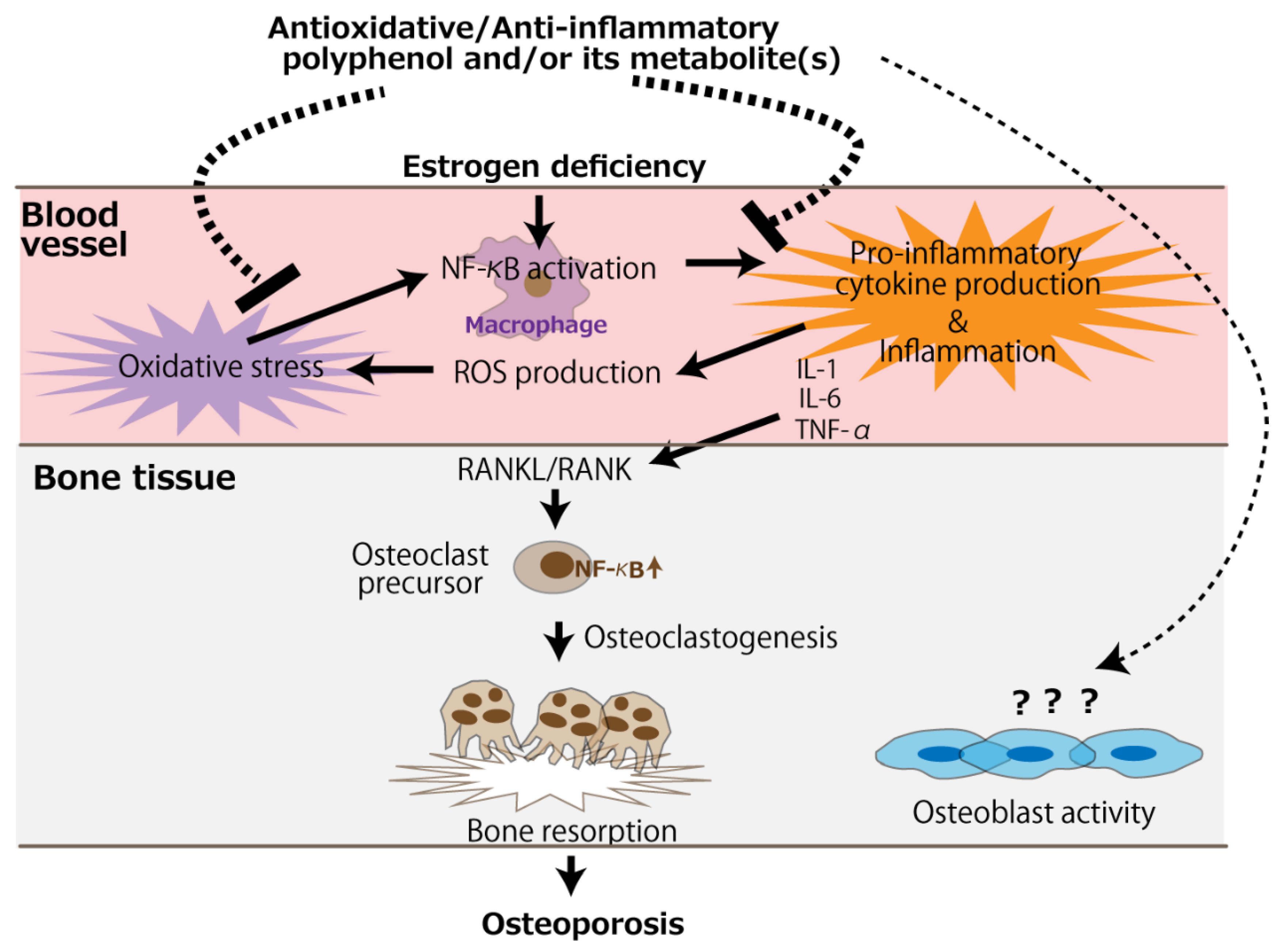
:1. Introduction
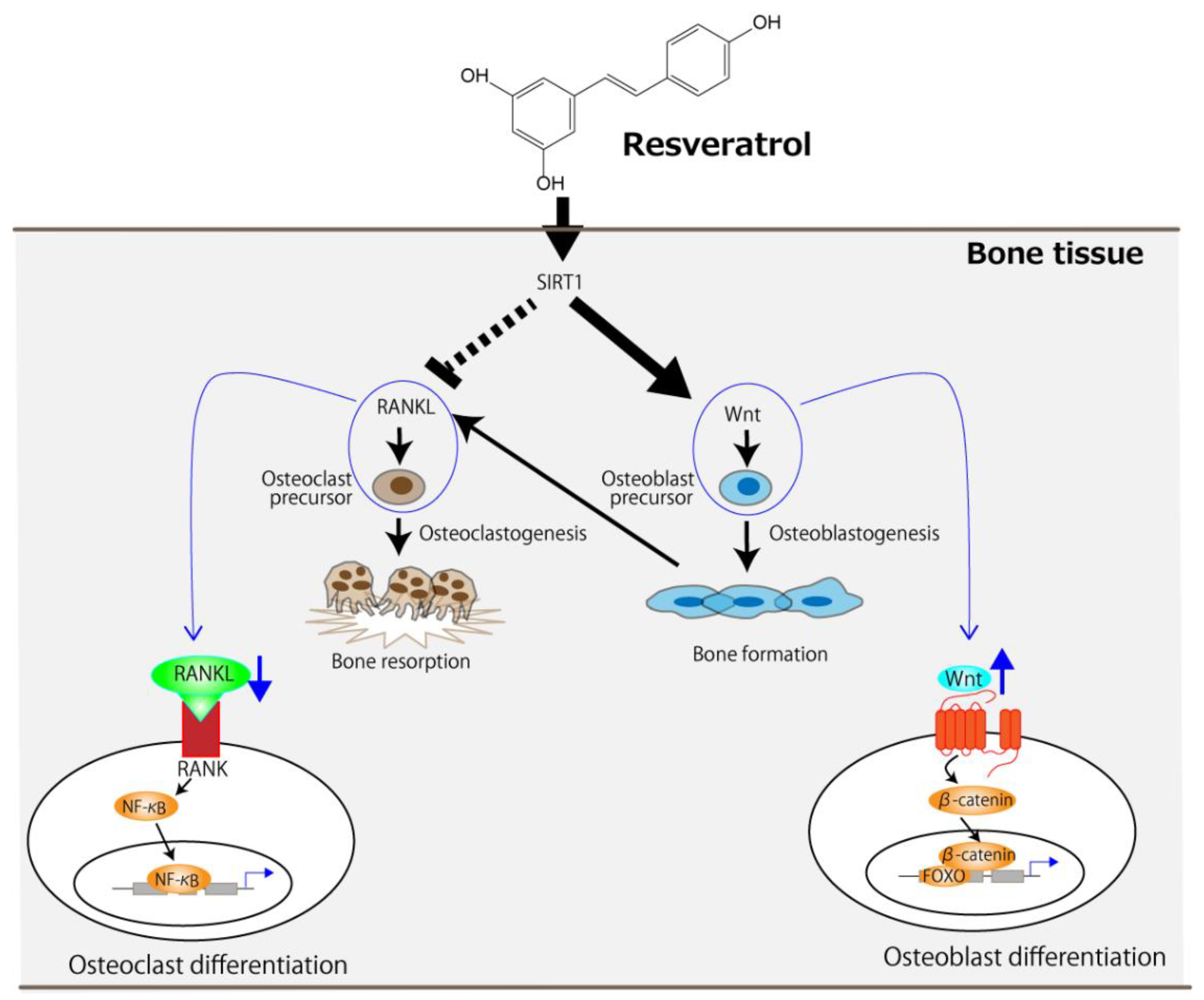
2. Resveratrol
3. Purified Polyphenolic Compounds Other Than Resveratrol
4. Polyphenol-Rich Substances or Extracts
5. Studies in Which Polyphenols Did Not Show Any Beneficial or Positive Effects on Bone Health in Ovariectomized Animals
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Christiansen, C. Consensus development conference: Prophylaxis and treatment of osteoporosis. Am. J. Med. 1991, 90, 107–110. [Google Scholar]
- Manolagas, S.C. From estrogen-centric to aging and oxidative stress: A revised perspective of the pathogenesis of osteoporosis. Endocr. Rev. 2010, 31, 266–300. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Zhu, L.; Zhang, D.; Li, N.; Li, Q.; Dai, P.; Mao, Y.; Li, X.; Ma, J.; Huang, S. Oxidative stress-related biomarkers in postmenopausal osteoporosis: A systematic review and meta-analyses. Dis. Markers 2016, 2016, 7067984. [Google Scholar] [CrossRef]
- Callaway, D.A.; Jiang, J.X. Reactive oxygen species and oxidative stress in osteoclastogenesis, skeletal aging and bone diseases. J. Bone Miner. Metab. 2015, 33, 359–370. [Google Scholar] [CrossRef]
- Vrtačnik, P.; Zupan, J.; Mlakar, V.; Kranjc, T.; Marc, J.; Kern, B.; Ostanek, B. Epigenetic enzymes influenced by oxidative stress and hypoxia mimetic in osteoblasts are differentially expressed in patients with osteoporosis and osteoarthritis. Sci. Rep. 2018, 8, 16215. [Google Scholar] [CrossRef]
- Mohamad, N.V.; Ima-Nirwana, S.; Chin, K.Y. Are Oxidative Stress and Inflammation Mediators of Bone Loss Due to Estrogen Deficiency? A Review of Current Evidence. Endocr. Metab. Immune Disord. Drug. Targets 2020, 20, 1478–1487. [Google Scholar] [CrossRef]
- Abdul Rahim, R.; Jayusman, P.A.; Muhammad, N.; Mohamed, N.; Lim, V.; Ahmad, N.H.; Mohamad, S.; Abdul Hamid, Z.A.; Ahmad, F.; Mokhtar, N.; et al. Potential antioxidant and anti-inflammatory efects of Spilanthes acmella and its health beneficial effects: A review. Int. J. Environ. Res. Public Health 2021, 18, 3532. [Google Scholar] [CrossRef]
- Nani, A.; Murtaza, B.; Sayed Khan, A.; Khan, N.A.; Hichami, A. Antioxidant and anti-Inflammatory potential of polyphenols contained in Mediterranean diet in obesity: Molecular mechanisms. Molecules 2021, 26, 985. [Google Scholar] [CrossRef]
- Hardcastle, A.C.; Aucott, L.; Reid, D.M.; Macdonald, H.M. Associations between dietary flavonoid intakes and bone health in a Scottish population. J. Bone Miner. Res. 2011, 26, 941–947. [Google Scholar] [CrossRef]
- Welch, A.; MacGregor, A.; Jennings, A.; Fairweather-Tait, S.; Spector, T.; Cassidy, A. Habitual flavonoid intakes are positively associated with bone mineral density in women. J. Bone Miner. Res. 2012, 27, 1872–1878. [Google Scholar] [CrossRef]
- Arakawa, H.; Maeda, M.; Okubo, S.; Shimamura, T. Role of hydrogen peroxide in bactericidal action of catechin. Biol. Pharm. Bull. 2004, 27, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Khan, H.Y.; Zubair, H.; Faisal, M.; Ullah, M.F.; Farhan, M.; Sarkar, F.H.; Ahmad, A.; Hadi, S.M. Plant polyphenol induced cell death in human cancer cells involves mobilization of intracellular copper ions and reactive oxygen species generation: A mechanism for cancer chemopreventive action. Mol. Nutr. Food Res. 2014, 58, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.O.; Kieran, N.E.; Howell, K.; Burne, M.J.; Varadarajan, R.; Dhakshinamoorthy, S.; Porter, A.G.; O’Farrelly, C.; Rabb, H.; Taylor, C.T. Reoxygenation-specific activation of the antioxidant transcription factor Nrf2 mediates cytoprotective gene expression in ischemia-reperfusion injury. FASEB J. 2006, 20, 2624–2626. [Google Scholar] [CrossRef]
- Shelton, L.M.; Park, B.K.; Copple, I.M. Role of Nrf2 in protection against acute kidney injury. Kidney Int. 2013, 84, 1090–1095. [Google Scholar] [CrossRef]
- Smith, R.E.; Tran, K.; Smith, C.C.; McDonald, M.; Shejwalkar, P.; Hara, K. The role of the Nrf2/ARE antioxidant system in preventing cardiovascular diseases. Diseases 2016, 4, 34. [Google Scholar] [CrossRef] [PubMed]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.L.; et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, M.; Kim, S.K.; Berdichevsky, A.; Guarente, L. A role for SIR-2.1 regulation of ER stress response genes in determining C. elegans life span. Dev. Cell 2005, 9, 605–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, J.H.; Goupil, S.; Garber, G.B.; Helfand, S.L. An accelerated assay for the identification of lifespan-extending interventions in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2004, 101, 12980–12985. [Google Scholar] [CrossRef] [Green Version]
- Valenzano, D.R.; Terzibasi, E.; Genade, T.; Cattaneo, A.; Domenici, L.; Cellerino, A. Resveratrol prolongs lifespan and retards the onset of age-related markers in a short-lived vertebrate. Curr. Biol. 2006, 16, 296–300. [Google Scholar] [CrossRef] [Green Version]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef]
- Guerrero, R.F.; García-Parrilla, M.C.; Puertas, B.; Cantos-Villar, E. Wine, resveratrol and health: A review. Nat. Prod. Commun. 2009, 4, 635–658. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Liu, S.; Ma, S.; Zhao, J.; Zhang, W.; Qi, W.; Cao, P.; Wang, Z.; Lei, W. Protective effects of resveratrol on postmenopausal osteoporosis: Regulation of SIRT1-NF-κB signaling pathway. Acta Biochim. Biophys. Sin. 2014, 46, 1024–1033. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Li, X.; Li, N.; Liu, T.; Liu, J.; Li, Z.; Xiao, H.; Li, J. Long-term resveratrol treatment prevents ovariectomy-induced osteopenia in rats without hyperplastic effects on the uterus. Br. J. Nutr. 2014, 111, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhang, L.M.; Guo, C.; Han, J.F. Resveratrol promotes osteoblastic differentiation in a rat model of postmenopausal osteoporosis by regulating autophagy. Nutr. Metab. 2020, 17, 29. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Deng, L.; Fan, J.; Zhang, Y. Effects of resveratrol on bone metabolism and bone turnover related indexes in ovariectomized osteoporosis rats. Cell Mol. Biol. 2020, 66, 92–97. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, M.W.; He, Y.; Deng, N.; Chen, Y.; Huang, J.; Xie, W. Protective effect of resveratrol on estrogen deficiency-induced osteoporosis though attenuating NADPH oxidase 4/nuclear factor kappa B pathway by increasing miR-92b-3p expression. Int. J. Immunopathol. Pharmacol. 2020, 34. [Google Scholar] [CrossRef]
- Molez, A.M.; do Nascimento, E.H.L.; Haiter Neto, F.; Cirano, F.R.; Pimentel, S.P.; Ribeiro, F.V.; Casati, M.Z.; Corrêa, M.G. Effect of resveratrol on the progression of experimental periodontitis in an ovariectomized rat model of osteoporosis: Morphometric, immune-enzymatic, and gene expression analysis. J. Periodontal. Res. 2020, 55, 840–849. [Google Scholar] [CrossRef]
- Elseweidy, M.M.; El-Swefy, S.E.; Shaheen, M.A.; Baraka, N.M.; Hammad, S.K. Effect of resveratrol and mesenchymal stem cell monotherapy and combined treatment in management of osteoporosis in ovariectomized rats: Role of SIRT1/FOXO3a and Wnt/β-catenin pathways. Arch. Biochem. Biophys. 2021, 703, 108856. [Google Scholar] [CrossRef]
- Guo, D.W.; Han, Y.X.; Cong, L.; Liang, D.; Tu, G.J. Resveratrol prevents osteoporosis in ovariectomized rats by regulating microRNA-338-3p. Mol. Med. Rep. 2015, 12, 2098–2106. [Google Scholar] [CrossRef] [Green Version]
- Franzoso, G.; Carlson, L.; Xing, L.; Poljak, L.; Shores, E.W.; Brown, K.D.; Leonardi, A.; Tran, T.; Boyce, B.F.; Siebenlist, U. Requirement for NF-kappaB in osteoclast and B-cell development. Genes Dev. 1997, 11, 3482–3496. [Google Scholar] [CrossRef] [Green Version]
- Iotsova, V.; Caamaño, J.; Loy, J.; Yang, Y.; Lewin, A.; Bravo, R. Osteopetrosis in mice lacking NF-kappaB1 and NF-kappaB2. Nat. Med. 1997, 3, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Jimi, E.; Aoki, K.; Saito, H.; D’Acquisto, F.; May, M.J.; Nakamura, I.; Sudo, T.; Kojima, T.; Okamoto, F.; Fukushima, H.; et al. Selective inhibition of NF-kappa B blocks osteoclastogenesis and prevents inflammatory bone destruction in vivo. Nat. Med. 2004, 10, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, H.; Mani, D.; Singh, D.; Gupta, A. The underlying pathophysiology and therapeutic approaches for osteoporosis. Med. Res. Rev. 2018, 38, 2024–2057. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.; Han, L.; Bartell, S.M.; Kim, H.N.; Gubrij, I.; de Cabo, R.; O’Brien, C.A.; Manolagas, S.C.; Almeida, M. Sirtuin1 (Sirt1) promotes cortical bone formation by preventing β-catenin sequestration by FoxO transcription factors in osteoblast progenitors. J. Biol. Chem. 2014, 289, 24069–24078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, Z.; Zhang, W.; Hu, X.; Wei, H.; Peng, J.; Jiang, S. SIRT1 inhibits adipogenesis and promotes myogenic differentiation in C3H10T1/2 pluripotent cells by regulating Wnt signaling. Cell Biosci. 2015, 5, 61. [Google Scholar] [CrossRef] [Green Version]
- Puel, C.; Quintin, A.; Mathey, J.; Obled, C.; Davicco, M.J.; Lebecque, P.; Kati-Coulibaly, S.; Horcajada, M.N.; Coxam, V. Prevention of bone loss by phloridzin, an apple polyphenol, in ovariectomized rats under inflammation conditions. Calcif. Tissue Int. 2005, 77, 311–318. [Google Scholar] [CrossRef]
- Puel, C.; Mathey, J.; Agalias, A.; Kati-Coulibaly, S.; Mardon, J.; Obled, C.; Davicco, M.J.; Lebecque, P.; Horcajada, M.N.; Skaltsounis, A.L.; et al. Dose-response study of effect of oleuropein, an olive oil polyphenol, in an ovariectomy/inflammation experimental model of bone loss in the rat. Clin. Nutr. 2006, 25, 859–868. [Google Scholar] [CrossRef]
- Puel, C.; Mardon, J.; Agalias, A.; Davicco, M.J.; Lebecque, P.; Mazur, A.; Horcajada, M.N.; Skaltsounis, A.L.; Coxam, V. Major phenolic compounds in olive oil modulate bone loss in an ovariectomy/inflammation experimental model. J. Agric. Food Chem. 2008, 56, 9417–9422. [Google Scholar] [CrossRef]
- Hagiwara, K.; Goto, T.; Araki, M.; Miyazaki, H.; Hagiwara, H. Olive polyphenol hydroxytyrosol prevents bone loss. Eur. J. Pharmacol. 2011, 662, 78–84. [Google Scholar] [CrossRef]
- Sehmisch, S.; Hammer, F.; Christoffel, J.; Seidlova-Wuttke, D.; Tezval, M.; Wuttke, W.; Stuermer, K.M.; Stuermer, E.K. Comparison of the phytohormones genistein, resveratrol and 8-prenylnaringenin as agents for preventing osteoporosis. Planta Med. 2008, 74, 794–801. [Google Scholar] [CrossRef]
- Kim, T.H.; Jung, J.W.; Ha, B.G.; Hong, J.M.; Park, E.K.; Kim, H.J.; Kim, S.Y. The effects of luteolin on osteoclast differentiation, function in vitro and ovariectomy-induced bone loss. J. Nutr. Biochem. 2011, 22, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Mundy, G.R. Osteoporosis and inflammation. Nutr. Rev. 2007, 65, S147–S151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.L.; Wang, P.; Guerrieri, J.; Yeh, J.K.; Wang, J.S. Protective effect of green tea polyphenols on bone loss in middle-aged female rats. Osteoporos Int. 2008, 19, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.L.; Yeh, J.K.; Stoecker, B.J.; Chyu, M.C.; Wang, J.S. Green tea polyphenols mitigate deterioration of bone microarchitecture in middle-aged female rats. Bone 2009, 44, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.L.; Smith, B.J.; Li, J.; Cao, J.J.; Song, X.; Newhardt, M.F.; Corry, K.A.; Tomison, M.D.; Tang, L.; Wang, J.S.; et al. Effect of long-term green tea polyphenol supplementation on bone architecture, turnover, and mechanical properties in middle-aged ovariectomized rats. Calcif. Tissue Int. 2019, 104, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Dreher, I.; Schütze, N.; Baur, A.; Hesse, K.; Schneider, D.; Köhrle, J.; Jakob, F. Selenoproteins are expressed in fetal human osteoblast-like cells. Biochem. Biophys. Res. Commun. 1998, 245, 101–107. [Google Scholar] [CrossRef]
- Kim, H.J.; Bae, Y.C.; Park, R.W.; Choi, S.W.; Cho, S.H.; Choi, Y.S.; Lee, W.J. Bone-protecting effect of safflower seeds in ovariectomized rats. Calcif. Tissue Int. 2002, 71, 88–94. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, Z.G.; Li, C.; Hu, S.J.; Liu, L.; Wang, J.P.; Mei, Q.B. Du-Zhong (Eucommia ulmoides Oliv.) cortex extract prevent OVX-induced osteoporosis in rats. Bone 2009, 45, 553–559. [Google Scholar] [CrossRef]
- Cladis, D.P.; Debelo, H.; Lachcik, P.J.; Ferruzzi, M.G.; Weaver, C.M. Increasing doses of bueberry polyphenols alters colonic metabolism and calcium absorption in ovariectomized rats. Mol. Nutr. Food Res. 2020, 64, e2000031. [Google Scholar] [CrossRef]
- Mei, F.; Meng, K.; Gu, Z.; Yun, Y.; Zhang, W.; Zhang, C.; Zhong, Q.; Pan, F.; Shen, X.; Xia, G.; et al. Arecanut (Areca catechu L.) seed polyphenol-ameliorated osteoporosis by altering gut microbiome via LYZ and the immune system in estrogen-deficient rats. J. Agric. Food Chem. 2021, 69, 246–258. [Google Scholar] [CrossRef]
- Kim, B.; Lee, S.H.; Song, S.J.; Kim, W.H.; Song, E.S.; Lee, J.C.; Lee, S.J.; Han, D.W.; Lee, J.H. Protective efects of mlon extracts on bone strength, mneralization, and metabolism in rats with ovariectomy-induced osteoporosis. Antioxidants 2019, 8, 306. [Google Scholar] [CrossRef] [Green Version]
- Turner, R.T.; Iwaniec, U.T.; Andrade, J.E.; Branscum, A.J.; Neese, S.L.; Olson, D.A.; Wagner, L.; Wang, V.C.; Schantz, S.L.; Helferich, W.G. Genistein administered as a once-daily oral supplement had no beneficial effect on the tibia in rat models for postmenopausal bone loss. Menopause 2013, 20, 677–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambati, S.; Miller, C.N.; Bass, E.F.; Hohos, N.M.; Hartzell, D.L.; Kelso, E.W.; Trunnell, E.R.; Yang, J.Y.; Della-Fera, M.A.; Baile, C.A.; et al. Synergistic phytochemicals fail to protect against ovariectomy induced bone loss in rats. J. Med. Food 2018, 21, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Cladis, D.P.; Swallow, E.A.; Allen, M.R.; Hill Gallant, K.M.; Weaver, C.M. Blueberry polyphenols do not improve bone mineral density or mechanical properties in ovariectomized rats. Calcif. Tissue Int. 2021; epub ahead of print. [Google Scholar] [CrossRef]
- Keiler, A.M.; Zierau, O.; Bernhardt, R.; Scharnweber, D.; Lemonakis, N.; Termetzi, A.; Skaltsounis, L.; Vollmer, G.; Halabalaki, M. Impact of a functionalized olive oil extract on the uterus and the bone in a model of postmenopausal osteoporosis. Eur. J. Nutr. 2014, 53, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Faughnan, M.; Hoey, L.; Wähälä, K.; Williamson, G.; Cassidy, A. Bioavailability of phyto-oestrogens. Br. J. Nutr. 2003, 89 (Suppl. 1), S45–S58. [Google Scholar] [CrossRef]
- Raisz, L.G. Pathogenesis of osteoporosis: Concepts, conflicts, and prospects. J. Clin. Investig. 2005, 115, 3318–3325. [Google Scholar] [CrossRef] [Green Version]
- Richardson, S.J.; Bai, A.; Kulkarni, A.A.; Moghaddam, M.F. Efficiency in drug discovery: Liver S9 fraction assay as a sreen for metabolic stability. Drug Metab. Lett. 2016, 10, 83–90. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Reference, Year of Publication, and Animal Species with Age at OVX | Dosage | Major Improved Parameter(s) | Mode of Action | |
|---|---|---|---|---|
| Overview | Molecular Level | |||
| [22] 2014 Rats aged 3 months | 5, 25, and 45 mg/kg/day for 8 weeks starting at 1 week after OVX | BMD of lumbar vertebrae (L3) and femur | Increased osteoblast differentiation | Activation of SIRT1 and subsequent suppression of NF-κB activity |
| [23] 2014 Rats aged 3–4 months | 20, 40, and 80 mg/kg/day for 12 weeks starting at 2 weeks after OVX | BMD of femur | ― | ― |
| [24] 2020 Rats aged 10–12 weeks | 10, 20, and 40 mg/kg/day for 8 weeks starting at 1 week after OVX | BMD of the lumbar vertebrae (L3) and the right distal femur–tibia bone region | Promotion of osteoblast differentiation and suppression of osteoclast differentiation via autophagy regulation | ― |
| [25] 2020 Rats aged 3 months | 5, 15, and 45 mg/kg/day for 12 weeks starting at 3 days after OVX | BMD of femur | Estrogen-like activity | ― |
| [26] 2020 Rats aged 6 months | 50, 100, and 200 mg/kg/day for 12 weeks after OVX | BMD of femur | ― | Suppression of cathepsin K expression and the Nox4/NF-κB signaling pathway through the elevated expression of miR-92b-3p |
| [27] 2020 Adult rats | 10 mg/kg/day for 140 days after OVX (sacrificed 28 days after experimental periodontitis initiation) | Alveolar bone mass | ― | Downregulation of NADPH oxidase levels |
| [28] 2021 Rats aged 3 months | 80 mg/kg/day for 8 weeks starting at 8 weeks after OVX | BMC and BMD of femur | Stimulation of osteogenesis and inhibition of osteoclastogenesis | Activation of sirtuin 1 (SIRT1) and wingless-related MMTV integration site (Wnt) pathways |
| Reference, Year of Publication, and Animal Species with Age at OVX | Dosage | Major Improved Parameter(s) | Mode of Action |
|---|---|---|---|
| Phloridzin (an apple polyphenol) | |||
| [36] 2005 Rats aged 6 months | Oral administration with a diet supplemented with 0.25% phloridzin for 80 days after OVX. Inflammation was induced by a subcutaneous injection of magnesium silicate 3 weeks before necropsy. | BMD of femur | Improving inflammatory marker levels and decreasing bone resorption |
| Oleuropein (an olive oil polyphenol) | |||
| [37] 2006 Rats aged 6 months | 2.5, 5, 10, and 15 mg/kg/day for 100 days after OVX | BMD of femur | Reducing inflammatory state |
| Tyrosol and hydroxytyrosol (olive oil polyphenols) | |||
| [38] 2008 Rats aged 6 months | Oral administration with a diet supplemented with either 0.017% tyrosol or 0.017% hydroxytyrosol for 84 days after OVX; Three weeks before the end of the investigation (d 63), inflammation was provoked by a subcutaneous injection of magnesium silicate. | BMD of femur | Possibly lowering the risk of inflammation-induced osteopenia via their antioxidant activity |
| Oleuropein and hydroxytyrosol (olive oil polyphenols) | |||
| [39] 2011 Mice aged 6 weeks | 10 mg/kg at 3-day intervals for 28 days after OVX | BMD of femur | Regulating oxidative stress via their antioxidant effects |
| Genistein (GEN) and 8-prenylnaringenin (8PN) | |||
| [40] 2008 Rats aged 3 months | 6 and 60 mg GEN/kg/day or 6.8 and 68 mg 8PN/kg/day for 3 months after OVX | BMD of tibia | Acting as phytoestrogens |
| Luteolin | |||
| [41] 2011 Mice aged 9 weeks | 5 and 20 mg/kg/day for 30 days starting at 1 week after OVX | BMD and BMC of femur | Reducing both osteoclast differentiation and function |
| Reference, Year of Publication, and Animal Species with Age at OVX | Dosage | Major Improved Parameter(s) | Mode of Action |
|---|---|---|---|
| Green tea (Camellia sinensis) polyphenols (GTPs) containing epigallocatechin-3-gallate | |||
| [43] 2008 Rats aged 14 months | 0.1% and 0.5% (w/v) GTP aqueous solution for 16 weeks after OVX | BMD of femur | An increase in antioxidant capacity and/or a decrease in oxidative stress damage |
| [44] 2009 Rats aged 14 months | 0.1% and 0.5% (w/v) GTP aqueous solution for 16 weeks after OVX | BMD of femur; Bone microarchitecture of tibia | An increase in antioxidant capacity and/or a decrease in oxidative stress damage |
| [45] 2019 Rats aged 6 months | 0.15%, 0.5%, 1.0%, and 1.5% (w/v) GTP aqueous solution for 3 and 6 months after OVX | Bone microarchitecture and mechanical properties of tibia, femur, and lumbar vertebrae (L3) | GTP’s antioxidative and anti-inflammatory actions |
| Defatted safflower (Carthamus tinctorius L.) seed powder containing lignans and flavones | |||
| [47] 2002 Rats aged 12 weeks | 290 g/kg diet for 4 weeks starting at 1 week after OVX | Bone mass of proximal tibia | Possibly stimulating osteoblast proliferation |
| Polyphenol-rich Du-Zhong (Eucommia ulmoides Oliv.) cortex extract containing lignans, phenolic acid, and flavonoids | |||
| [48] 2009 Rats aged 3 months | 100, 300, and 500 mg/kg/day for 16 weeks starting at 4 weeks after OVX | BMD of femur | Possibly stimulating osteoblast activity and inhibiting osteoclast resorption through Erβ |
| Polyphenol-rich blueberry (Vaccinium corymbosum) extract containing anthocyanins, phenolic acids, plavan-3-ols, and flavonols | |||
| [49] 2020 Rats aged 5 months | 75, 350, and 1000 mg total polyphenols/kg/day for 8 days starting at 12 days after OVX | Calcium absorption | ― |
| Arecanut (Areca catechu L.) seed polyphenol containing proanthocyanidin b2, procyanidin b1, catechin, etc. | |||
| [50] 2021 Rats weighing 190 ± 10 g | 400 and 800 mg/kg/day for 90 days starting at 30 days after OVX | Trabecular microstructure of femur | Promoting bone formation by altering gut microbiota along with controlling inflammatory reaction |
| Polyphenol-rich heat-treated melon (Cucumis melo L.) extract | |||
| [51] 2019 Rats aged 7 weeks | 1 mL of the extract 3 times/day for 4 weeks starting at 8 weeks after OVX | Bone strengths of femur; BMC and BMD of whole body, femur, and lumbar vertebrae 4–6 | Potent antioxidant activity leading to protection from the decline in bone strength, mineralization, and metabolism |
| Reference, Year of Publication, and Animal Species with Age at OVX | Dosage | Observations |
|---|---|---|
| Genistein | ||
| [52] 2013 Rats aged 7, 16, and 22 months | A pellet/day (485 and 970 μg genistein/pellet) for 5 months after OVX | No significant effects on cancellous or cortical bone mass or architecture of tibia |
| Mixure of quercetin (QUE), genistein (GEN), resveratrol (RES), and vitamin D3 (VD) | ||
| [53] 2018 Aged rats (retired breeder) | 1000 mg QUE/kg diet, 500 mg GEN/kg diet, 200 mg RES/kg diet, and 2400 IU VD/kg diet for 4 weeks after OVX; 2000 mg QUE/kg diet, 1000 mg GEN/kg diet, 400 mg RES/kg diet, and 2400 IU VD/kg diet, for 4 weeks after OVX | No significant effects on BMD of whole femur or L4 or L5 |
| Purified extract of blueberry (Vaccinium corymbosum) polyphenols or lyophilized blueberries | ||
| [54] 2021 Rats aged 5 months | 50, 250, or 1000 mg total polyphenols/kg/day for purified blueberry polyphenols or 50 mg total polyphenols/kg/day for lyophilized whole blueberries, for 90 days starting 1 month after OVX | Insignificant effects on BMD and bone mechanical properties |
| Extra virgin olive (Olea europaea var. Koroneiki) oil total polyphenolic fraction (TPF) containing oleocanthal, oleacein, and ligstroside as major polyphenols | ||
| [55] 2014 Rats aged ~12 months | 800 mg TPF/kg diet for 12 weeks after OVX | No significant effects on the bone loss of tibia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niwano, Y.; Kohzaki, H.; Shirato, M.; Shishido, S.; Nakamura, K. Anti-Osteoporotic Mechanisms of Polyphenols Elucidated Based on In Vivo Studies Using Ovariectomized Animals. Antioxidants 2022, 11, 217. https://doi.org/10.3390/antiox11020217
Niwano Y, Kohzaki H, Shirato M, Shishido S, Nakamura K. Anti-Osteoporotic Mechanisms of Polyphenols Elucidated Based on In Vivo Studies Using Ovariectomized Animals. Antioxidants. 2022; 11(2):217. https://doi.org/10.3390/antiox11020217
Chicago/Turabian StyleNiwano, Yoshimi, Hidetsugu Kohzaki, Midori Shirato, Shunichi Shishido, and Keisuke Nakamura. 2022. "Anti-Osteoporotic Mechanisms of Polyphenols Elucidated Based on In Vivo Studies Using Ovariectomized Animals" Antioxidants 11, no. 2: 217. https://doi.org/10.3390/antiox11020217
APA StyleNiwano, Y., Kohzaki, H., Shirato, M., Shishido, S., & Nakamura, K. (2022). Anti-Osteoporotic Mechanisms of Polyphenols Elucidated Based on In Vivo Studies Using Ovariectomized Animals. Antioxidants, 11(2), 217. https://doi.org/10.3390/antiox11020217