
MnTnHex-2-PyP5+ Displays Anticancer Properties and Enhances Cisplatin Effects in Non-Small Cell Lung Cancer Cells
 ,
,  ,
,  , ,
, ,  , , ,
, , ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. Gene Expression
2.4. Crystal Violet (CV) Staining Assay
2.5. MTS Reduction Assay
2.6. Combinatory Assays with MnTnHex and Cisplatin
2.7. Cell Cycle Analysis
2.8. Migration
2.8.1. Selection of MnTnHex and Cisplatin Concentrations for the Migration Assay
2.8.2. In Vitro Wound-Healing Assay
2.9. Metabolomics
2.9.1. Cell Culture and Collection of the Extracellular Medium
2.9.2. GC-MS Analysis: Sample Preparation and Analytical Conditions
2.9.3. Compound Identification and GC-MS Data Pre-Processing
2.9.4. Statistical Analyses of the GC-MS Data
3. Results
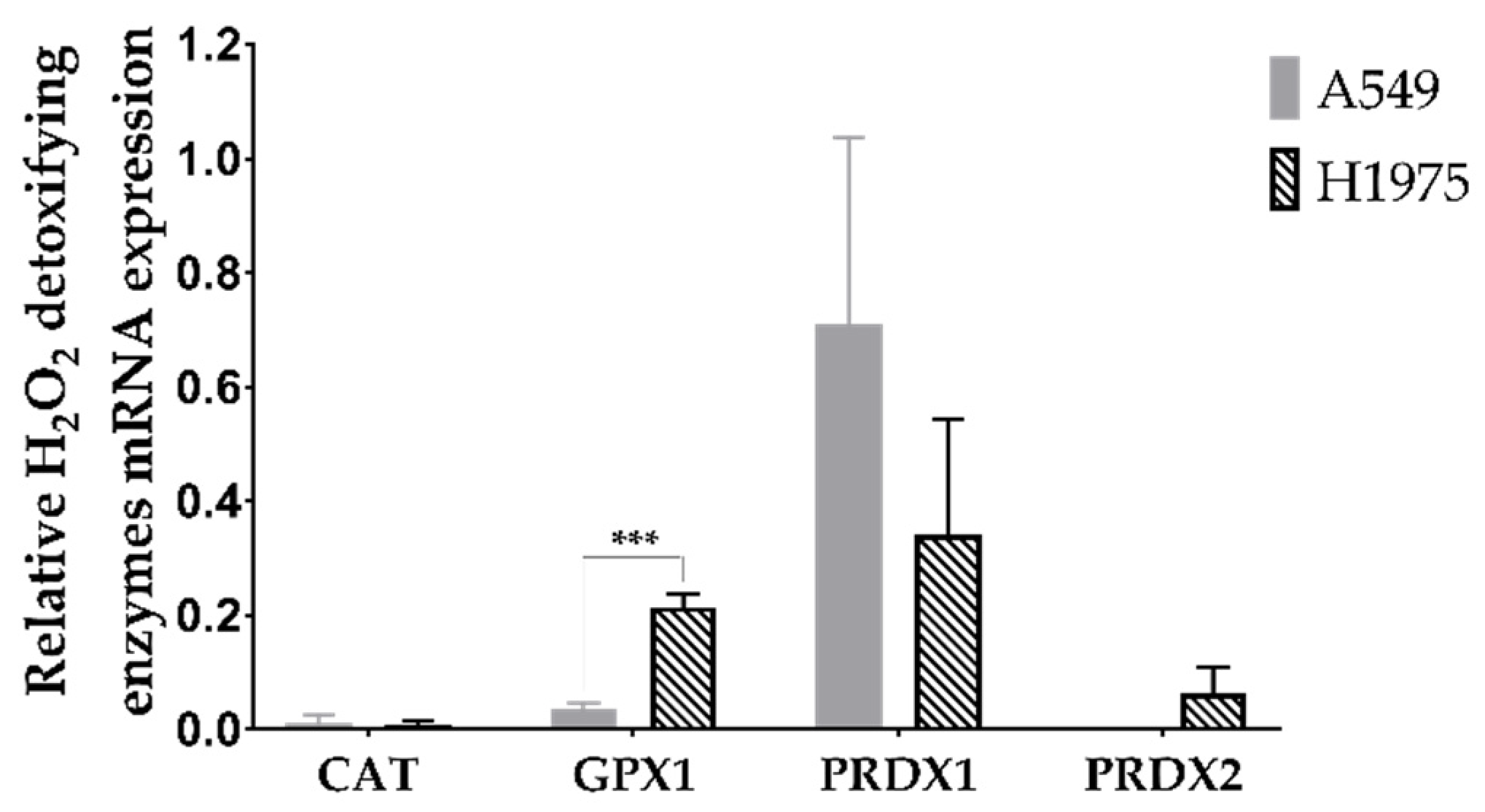
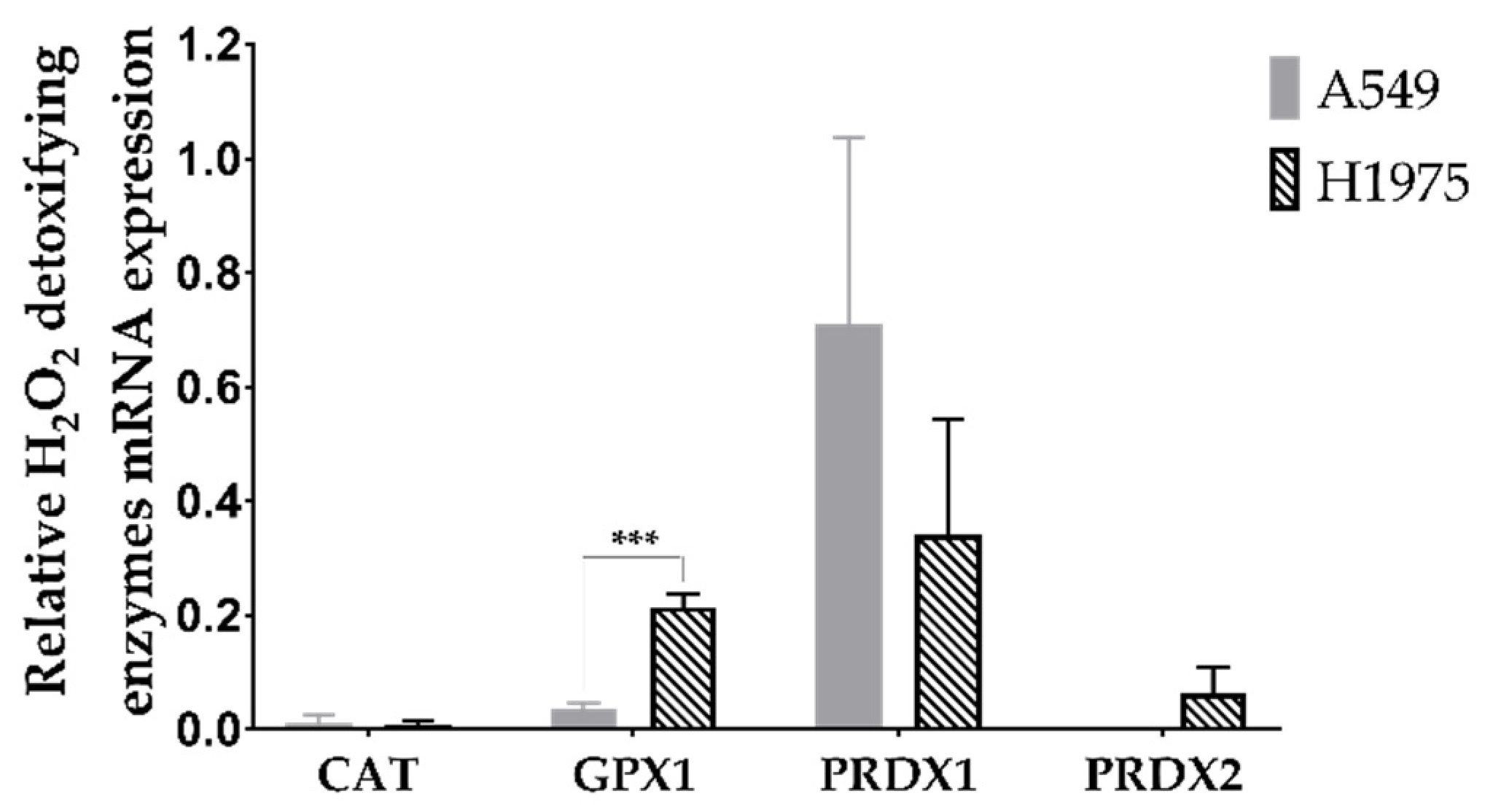
3.1. A549 and H1975 Cell Lines Express Low Catalase Levels
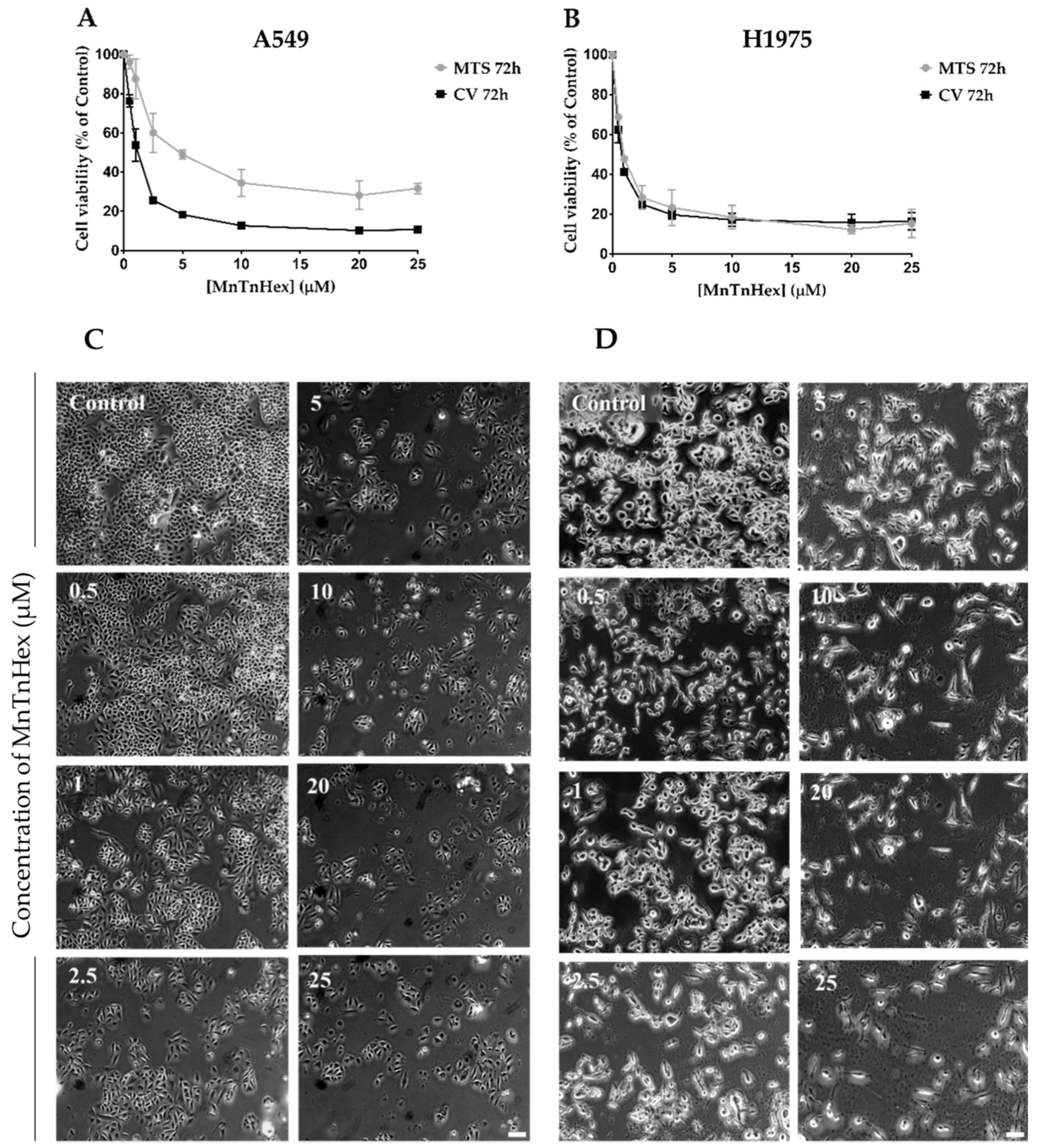
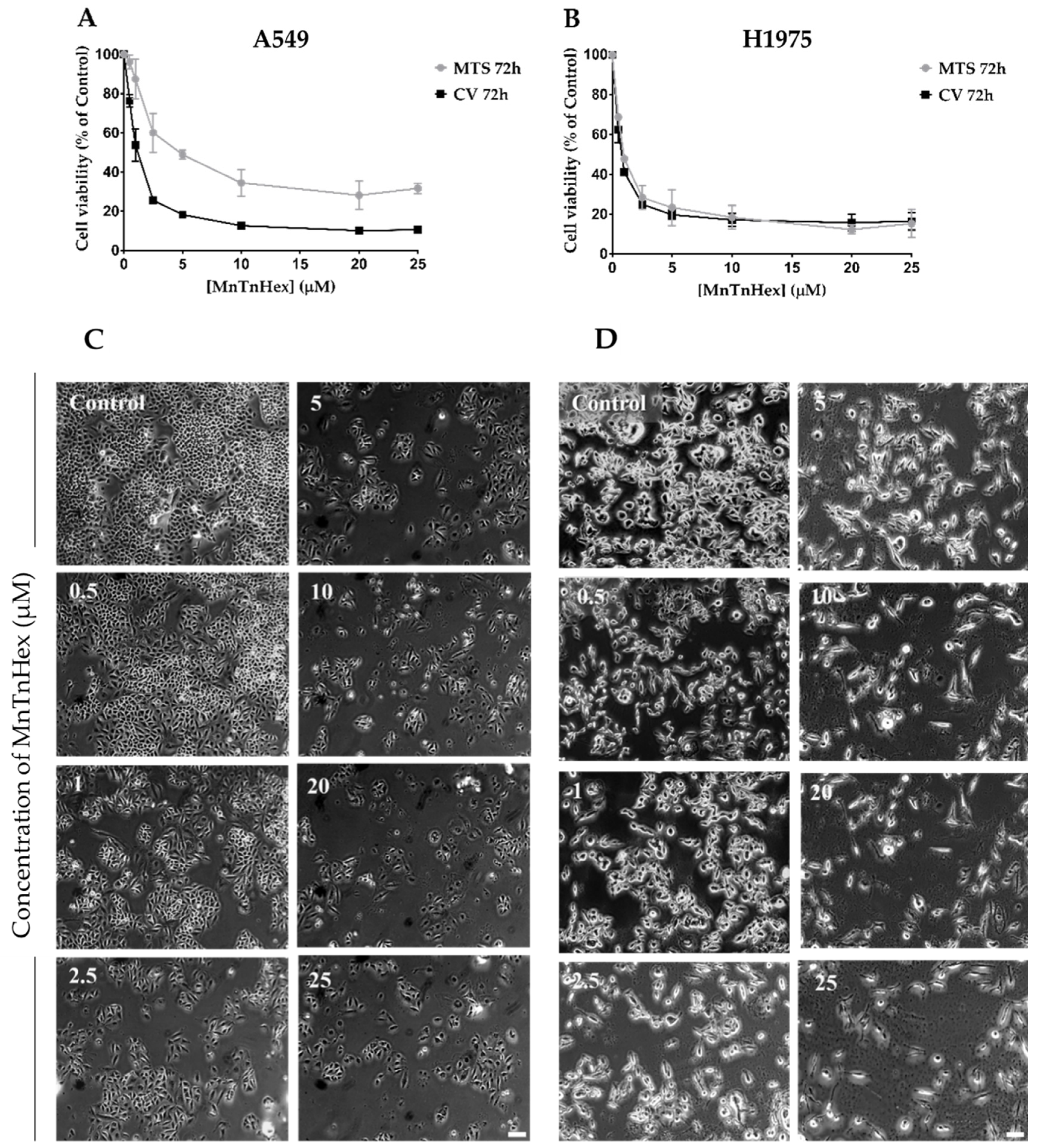
3.2. MnTnHex as a Single Drug Displays a Marked Cytotoxic Effect in NSCLC Cells
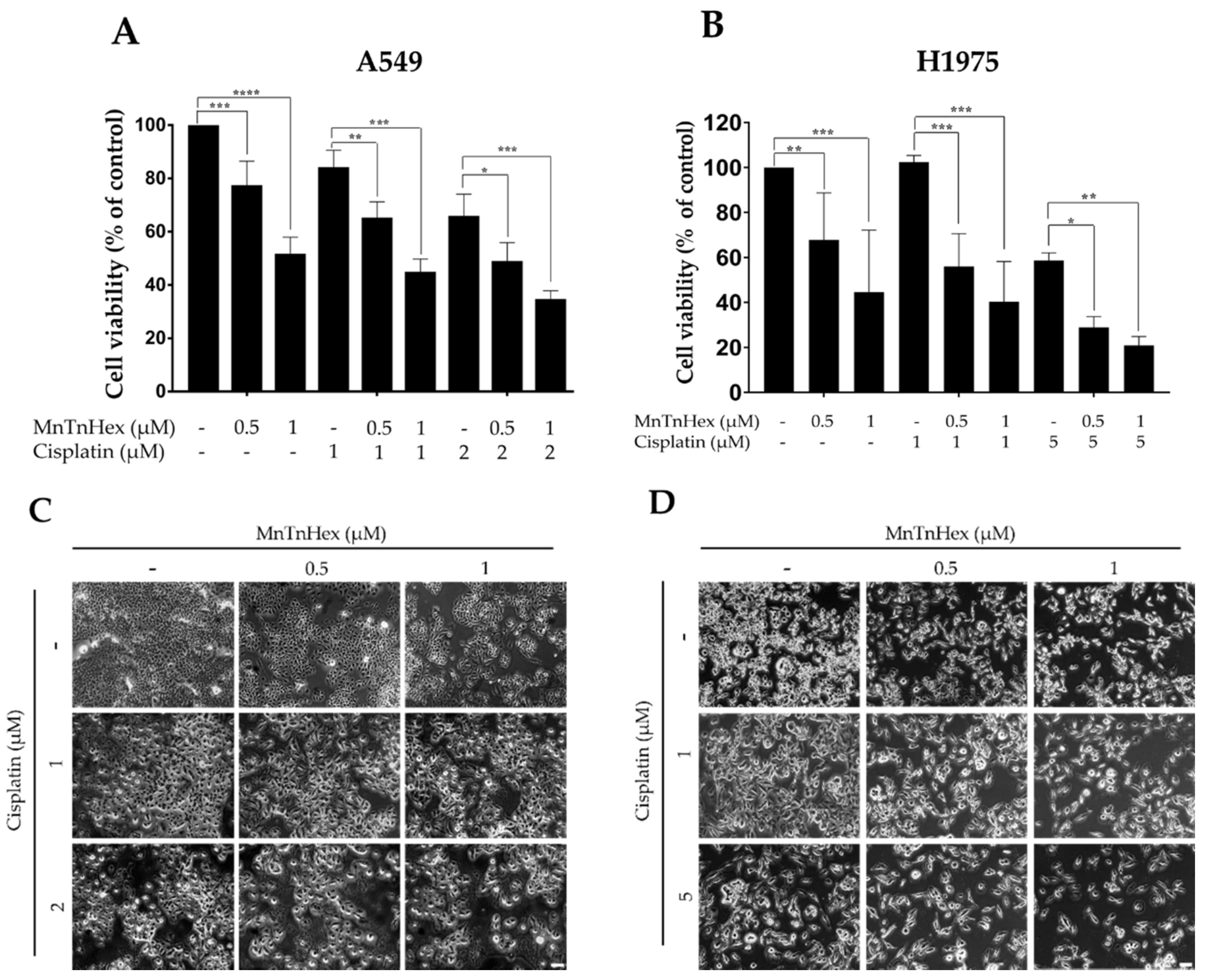
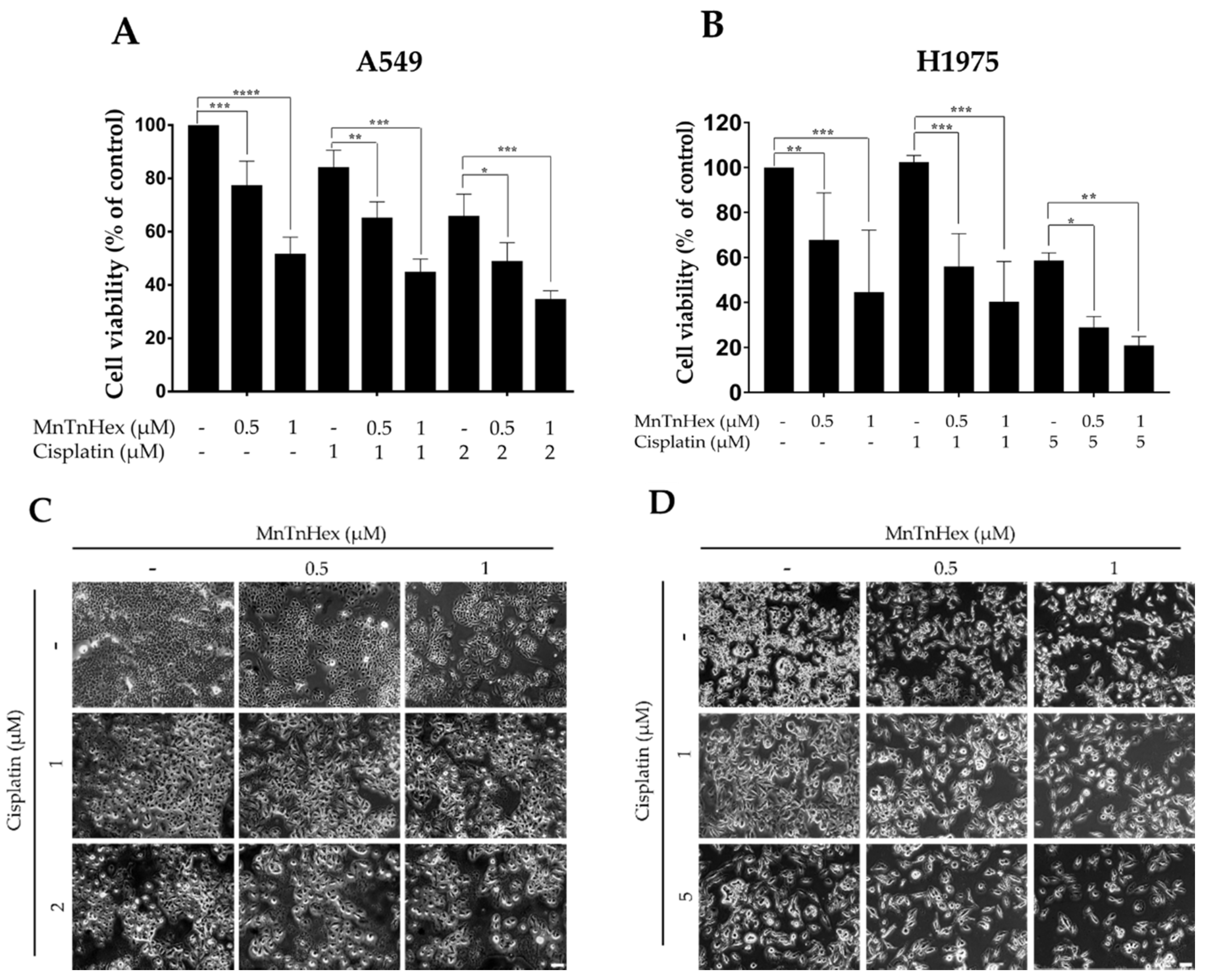
3.3. MnTnHex Enhances the Cytotoxicity of Cisplatin
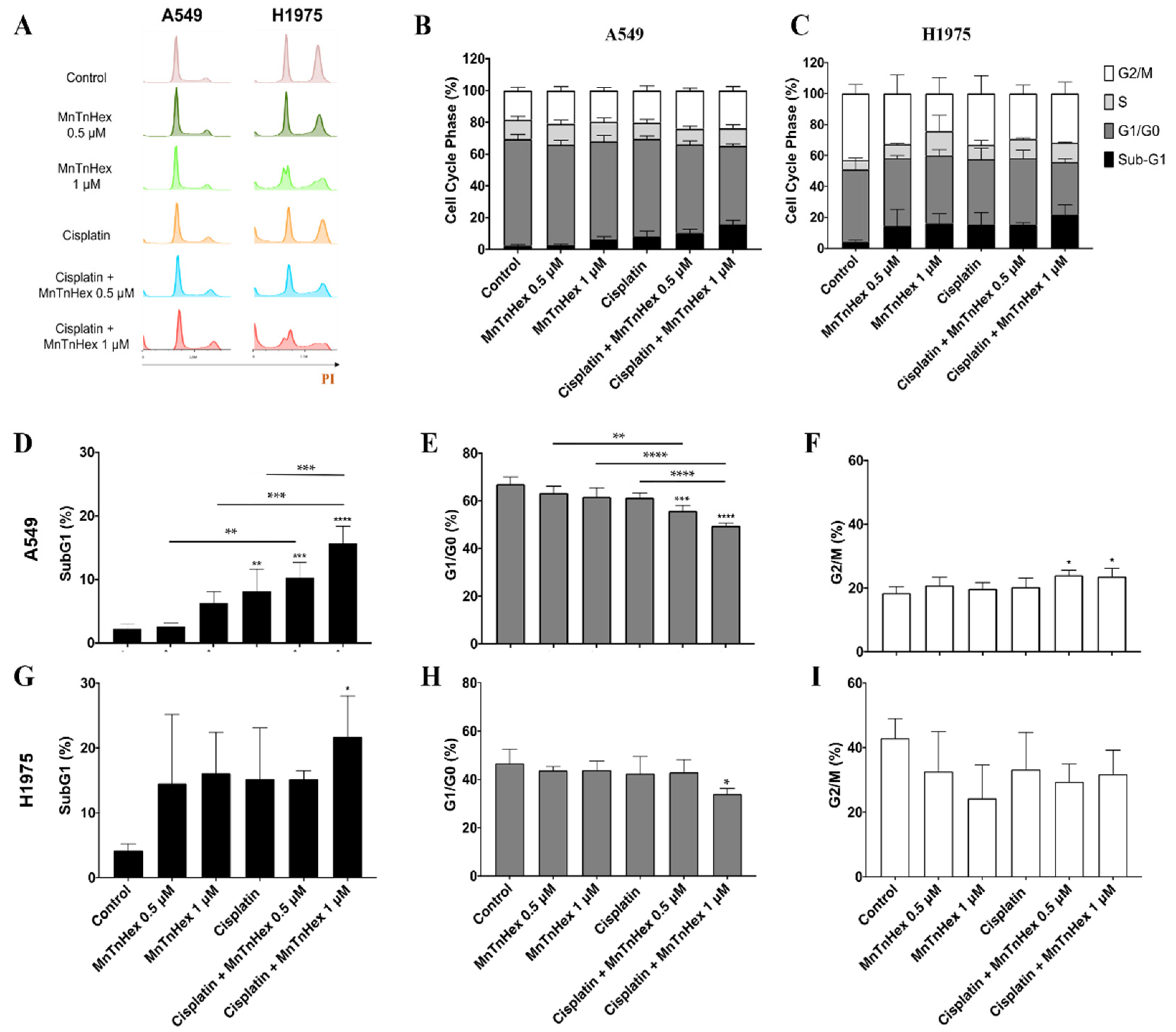
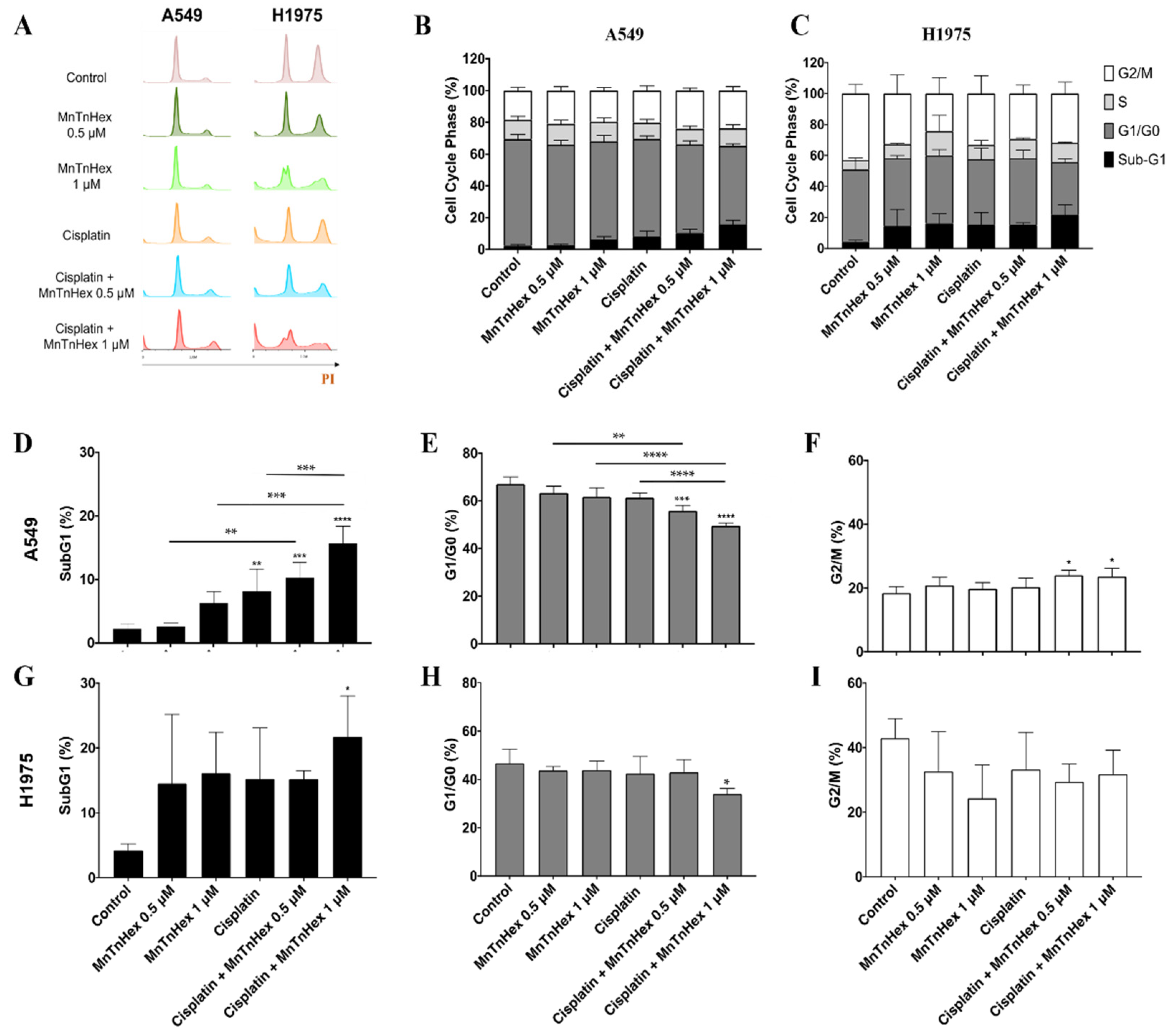
3.4. MnTnHex Enhances Cisplatin-Induced Cell Death
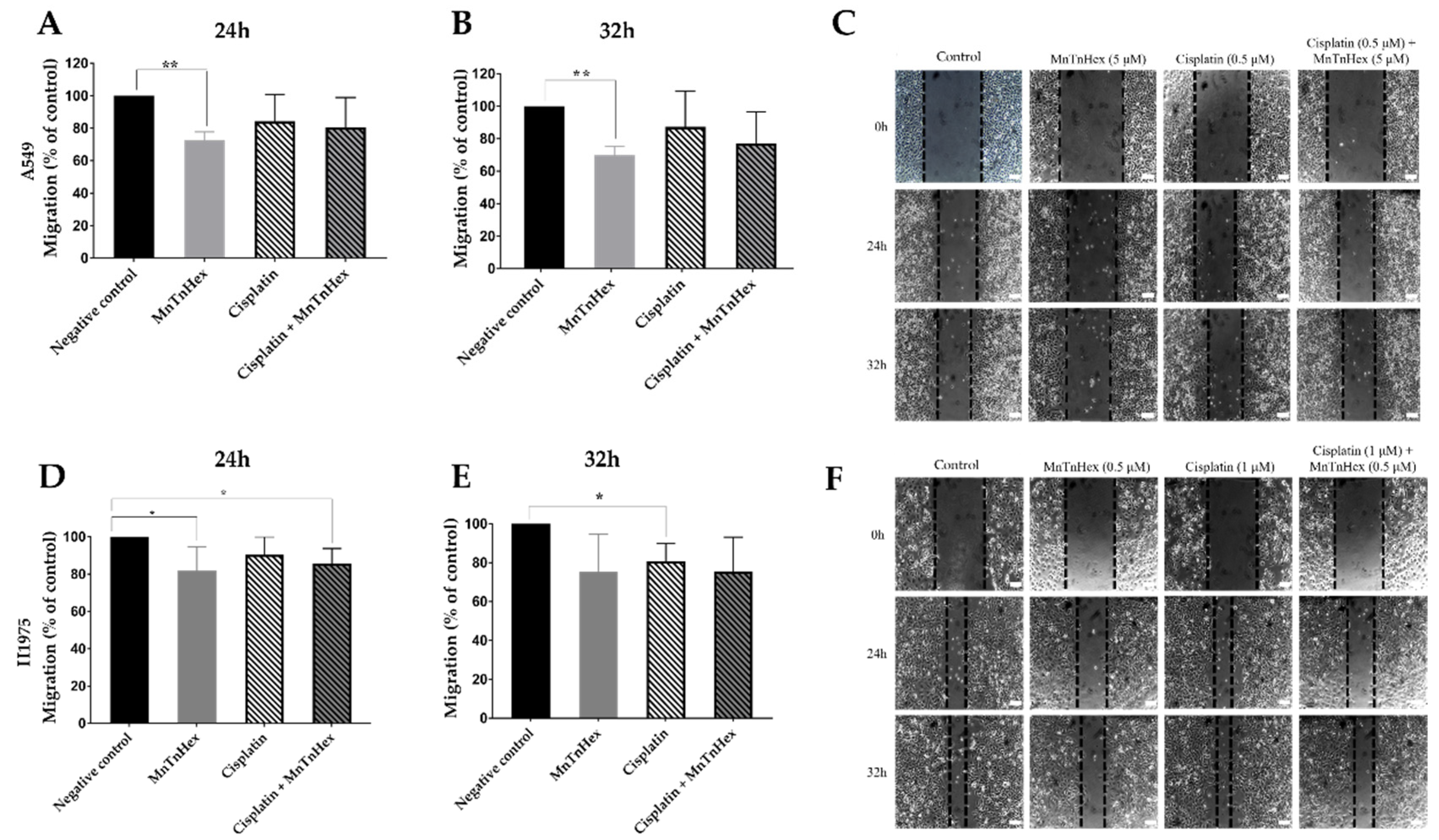
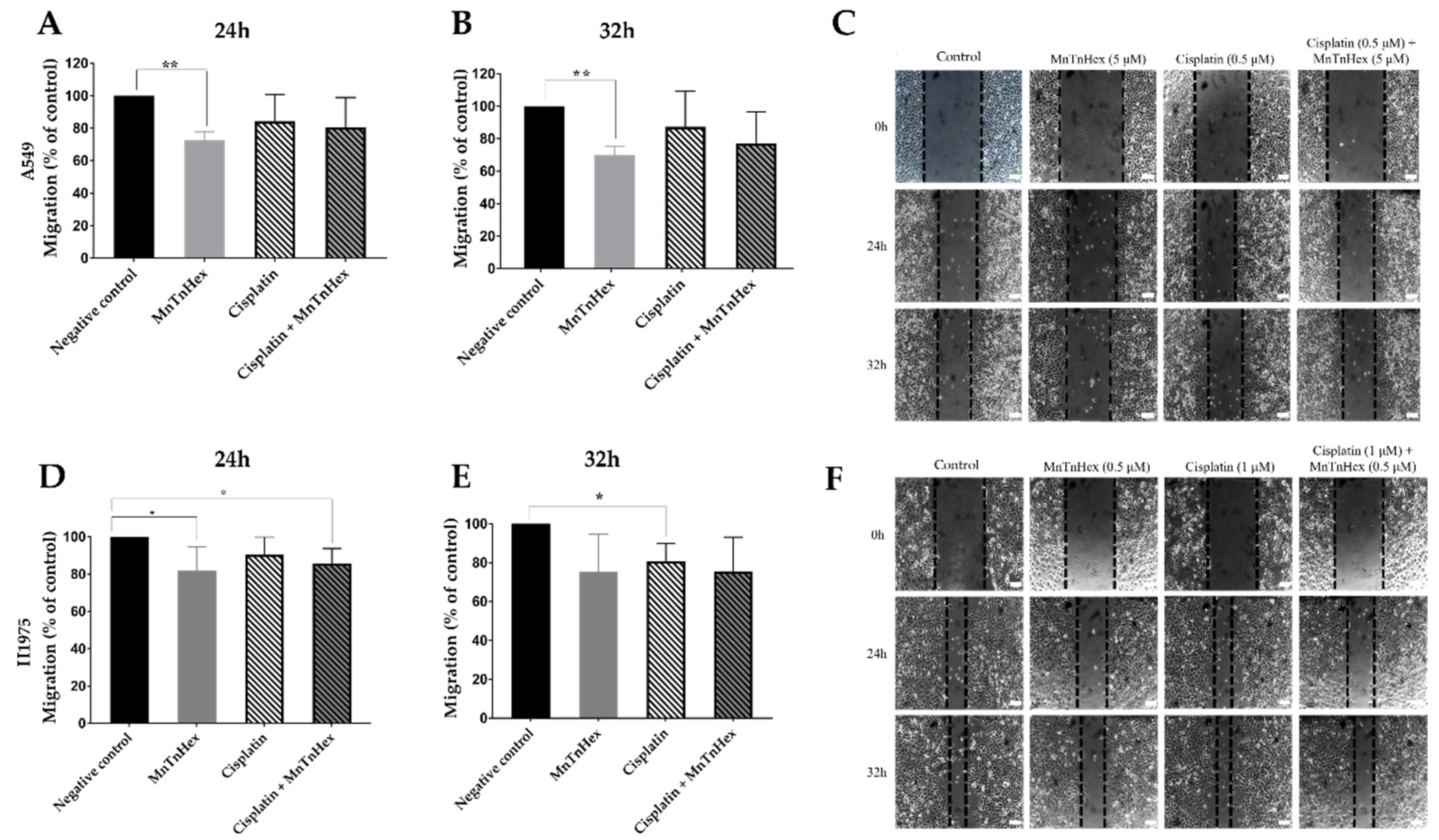
3.5. MnTnHex Reduces Collective Cell Migration
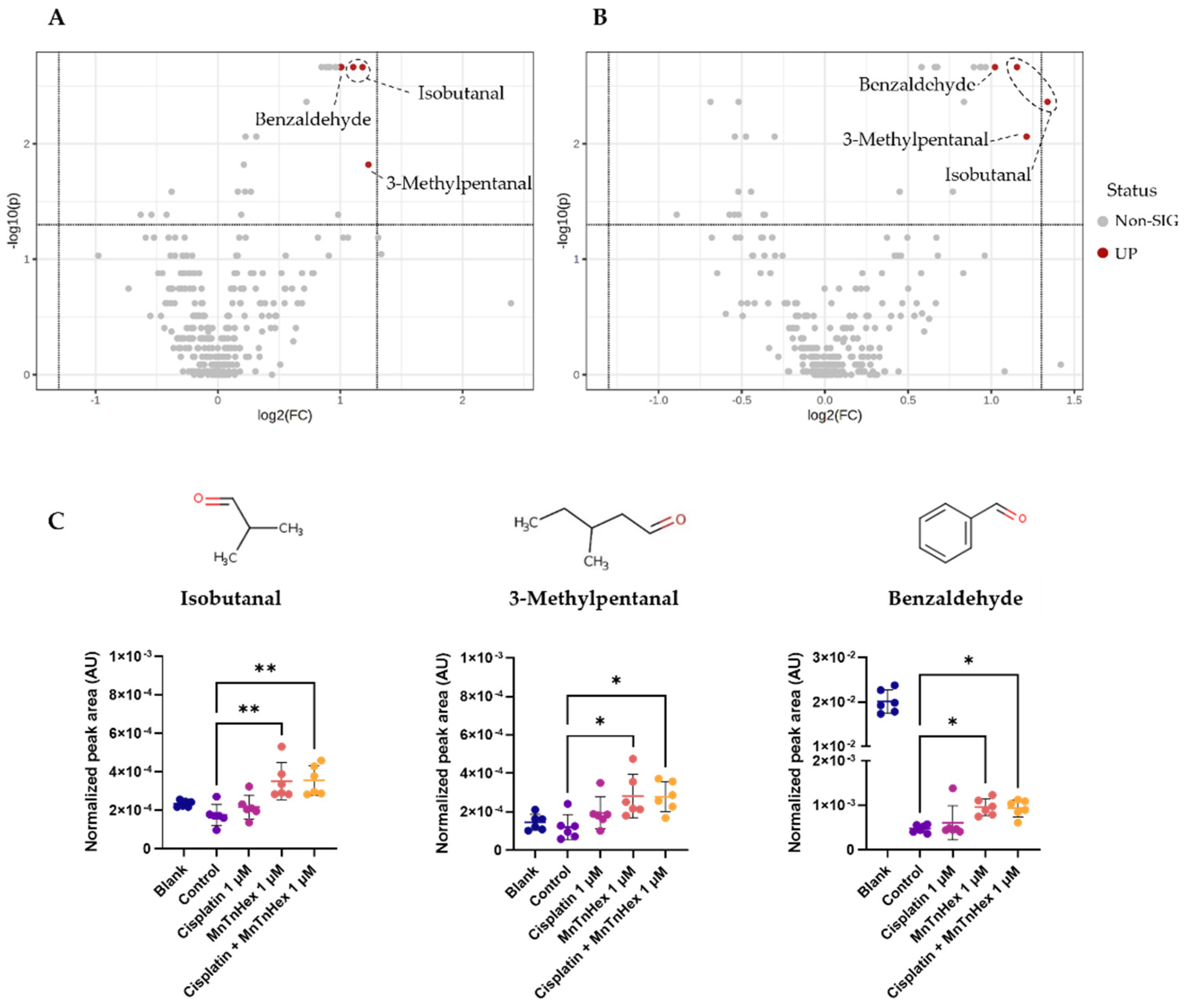
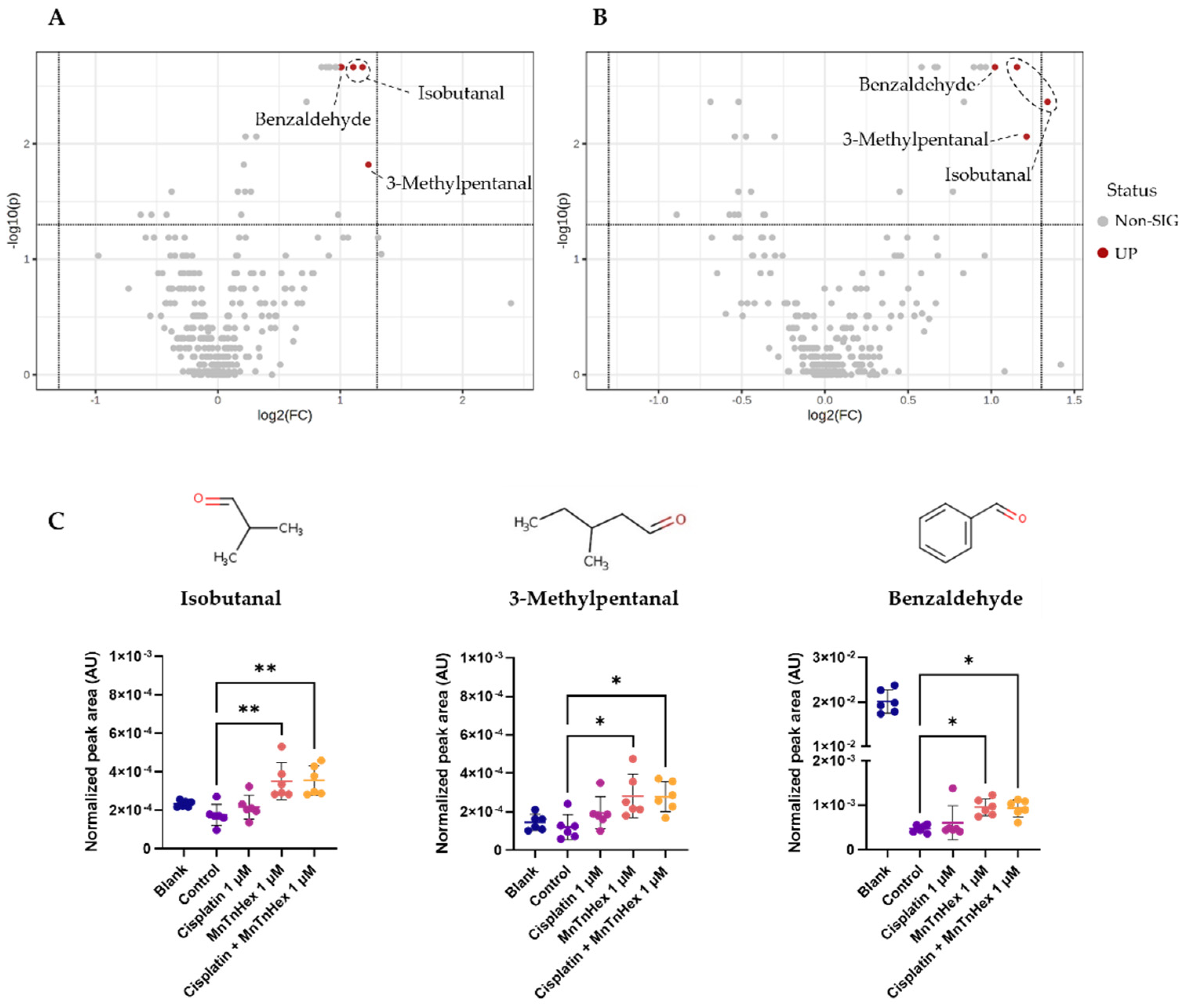
3.6. MnTnHex Alone and/or Combined with Cisplatin Induced Alterations in the Metabolic Response of NSCLC Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The Biology and Management of Non-Small Cell Lung Cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Schabath, M.B.; Cote, M.L. Cancer Progress and Priorities: Lung Cancer. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Song, J.; Feng, P.; Wang, Y.; Long, W.; Liu, M.; Li, L. Elevated Serum Apolipoprotein E Is Associated with Metastasis and Poor Prognosis of Non-Small Cell Lung Cancer. Tumor Biol. 2016, 37, 10715–10721. [Google Scholar] [CrossRef]
- Albuquerque, C.; Manguinhas, R.; Costa, J.G.; Gil, N.; Codony-Servat, J.; Castro, M.; Miranda, J.P.; Fernandes, A.S.; Rosell, R.; Oliveira, N.G. A narrative review of the migration and invasion features of non-small cell lung cancer cells upon xenobiotic exposure: Insights from in vitro studies. Transl. Lung Cancer Res. 2021, 10, 2698–2714. [Google Scholar] [CrossRef]
- Duma, N.; Santana-Davila, R.; Molina, J.R. Non–Small Cell Lung Cancer: Epidemiology, Screening, Diagnosis, and Treatment. Mayo Clin. Proc. 2019, 94, 1623–1640. [Google Scholar] [CrossRef]
- Antonia, S.J.; Villegas, A.; Daniel, D.; Vicente, D.; Murakami, S.; Hui, R.; Kurata, T.; Chiappori, A.; Lee, K.H.; de Wit, M.; et al. Overall Survival with Durvalumab after Chemoradiotherapy in Stage III NSCLC. N. Engl. J. Med. 2018, 379, 2342–2350. [Google Scholar] [CrossRef]
- Rocha, C.R.R.; Silva, M.M.; Quinet, A.; Cabral-Neto, J.B.; Menck, C.F.M. DNA Repair Pathways and Cisplatin Resistance: An Intimate Relationship. Clinics 2018, 73, e478s. [Google Scholar] [CrossRef]
- Ghosh, S. Cisplatin: The First Metal Based Anticancer Drug. Bioorg. Chem. 2019, 88, 102925. [Google Scholar] [CrossRef]
- Griesinger, F.; Korol, E.E.; Kayaniyil, S.; Varol, N.; Ebner, T.; Goring, S.M. Efficacy and Safety of First-Line Carboplatin-versus Cisplatin-Based Chemotherapy for Non-Small Cell Lung Cancer: A Meta-Analysis. Lung Cancer 2019, 135, 196–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, A.; Di Maio, M. Platinum-Based Chemotherapy in Advanced Non-Small-Cell Lung Cancer: Optimal Number of Treatment Cycles. Expert Rev. Anticancer Ther. 2016, 16, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Patel, A.K.; Shah, N.; Chaudhary, A.K.; Jha, U.K.; Yadav, U.C.; Gupta, P.K.; Pakuwal, U. Oxidative Stress and Antioxidants in Disease and Cancer: A Review. Asian Pac. J. Cancer Prev. 2014, 15, 4405–4409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egea, J.; Fabregat, I.; Frapart, Y.M.; Ghezzi, P.; Görlach, A.; Kietzmann, T.; Kubaichuk, K.; Knaus, U.G.; Lopez, M.G.; Olaso-Gonzalez, G.; et al. European Contribution to the Study of ROS: A Summary of the Findings and Prospects for the Future from the COST Action BM1203 (EU-ROS). Redox Biol. 2017, 13, 94–162. [Google Scholar] [CrossRef] [Green Version]
- Pisoschi, A.M.; Pop, A. The Role of Antioxidants in the Chemistry of Oxidative Stress: A Review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Oberley, T.D. Antioxidant Enzyme Levels in Cancer. Histol. Histopathol. 1997, 12, 525–535. [Google Scholar] [PubMed]
- Doskey, C.M.; Buranasudja, V.; Wagner, B.A.; Wilkes, J.G.; Du, J.; Cullen, J.J.; Buettner, G.R. Tumor Cells Have Decreased Ability to Metabolize H2O2: Implications for Pharmacological Ascorbate in Cancer Therapy. Redox Biol. 2016, 10, 274–284. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.G.; Yang, K.S.; Kang, S.W.; Woo, H.A.; Chang, T.S. Controlled Elimination of Intracellular H2O2: Regulation of Peroxiredoxin, Catalase, and Glutathione Peroxidase via Post-Translational Modification. Antioxid. Redox Signal. 2005, 7, 619–626. [Google Scholar] [CrossRef]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of Oxidative Stress as an Anticancer Strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef]
- Wondrak, G.T. Redox-Directed Cancer Therapeutics: Molecular Mechanisms and Opportunities. Antioxid. Redox Signal. 2009, 11, 3013–3069. [Google Scholar] [CrossRef]
- Larosa, V.; Remacle, C. Insights into the Respiratory Chain and Oxidative Stress. Biosci. Rep. 2018, 38, BSR20171492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukai, T.; Ushio-Fukai, M. Superoxide Dismutases: Role in Redox Signaling, Vascular Function, and Diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonetta, R. Potential Therapeutic Applications of MnSODs and SOD-Mimetics. Chem. A Eur. J. 2018, 24, 5032–5041. [Google Scholar] [CrossRef] [Green Version]
- Flórido, A.; Saraiva, N.; Cerqueira, S.; Almeida, N.; Parsons, M.; Batinic-Haberle, I.; Miranda, J.P.; Costa, J.G.; Carrara, G.; Castro, M.; et al. The Manganese(III) Porphyrin MnTnHex-2-PyP5+ Modulates Intracellular ROS and Breast Cancer Cell Migration: Impact on Doxorubicin-Treated Cells. Redox Biol. 2019, 20, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Batinic-Haberle, I.; Tovmasyan, A.; Huang, Z.; Duan, W.; Du, L.; Siamakpour-Reihani, S.; Cao, Z.; Sheng, H.; Spasojevic, I.; Alvarez Secord, A. H2O2-Driven Anticancer Activity of Mn Porphyrins and the Underlying Molecular Pathways. Oxid. Med. Cell Longev. 2021, 2021, 6653790. [Google Scholar] [CrossRef]
- Fernandes, A.S.; Flórido, A.; Saraiva, N.; Cerqueira, S.; Ramalhete, S.; Cipriano, M.; Cabral, M.F.; Miranda, J.P.; Castro, M.; Costa, J.; et al. Role of the Copper(II) Complex Cu[15]pyN5 in Intracellular ROS and Breast Cancer Cell Motility and Invasion. Chem. Biol. Drug Des. 2015, 86, 578–588. [Google Scholar] [CrossRef]
- Mapuskar, K.A.; Anderson, C.M.; Spitz, D.R.; Batinic-Haberle, I.; Allen, B.G.; Oberley-Deegan, R.E. Utilizing Superoxide Dismutase Mimetics to Enhance Radiation Therapy Response While Protecting Normal Tissues. Semin. Radiat. Oncol. 2019, 29, 72–80. [Google Scholar] [CrossRef]
- Tovmasyan, A.; Bueno-Janice, J.C.; Jaramillo, M.C.; Sampaio, R.S.; Reboucas, J.S.; Kyui, N.; Benov, L.; Deng, B.; Huang, T.T.; Tome, M.E.; et al. Radiation-Mediated Tumor Growth Inhibition Is Significantly Enhanced with Redox-Active Compounds That Cycle with Ascorbate. Antioxid. Redox Signal. 2018, 29, 1196–1214. [Google Scholar] [CrossRef]
- Costa, J.G.; Saraiva, N.; Batinic-Haberle, I.; Castro, M.; Oliveira, N.G.; Fernandes, A.S. The SOD Mimic MnTnHex-2-Pyp5+ Reduces the Viability and Migration of 786-O Human Renal Cancer Cells. Antioxidants 2019, 8, 490. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.W.; Choi, C.; Kim, H.; Kim, Y.; Park, S.; Kim, S.Y.; Batinic-Haberle, I.; Park, W. MnTnHex-2-PyP5+, Coupled to Radiation, Suppresses Metastasis of 4T1 and MDA-MB-231 Breast Cancer via AKT/Snail/EMT Pathways. Antioxidants 2021, 10, 1769. [Google Scholar] [CrossRef]
- Keir, S.T.; Dewhirst, M.W.; Kirkpatrick, J.P.; Bigner, D.D.; Batinic-Haberle, I. Cellular Redox Modulator, Ortho Mn(III) Meso-Tetrakis(N-n-Hexylpyridinium-2-Yl)Porphyrin, MnTnHex-2-PyP5+ in the Treatment of Brain Tumors. Anticancer Agents Med. Chem. 2011, 11, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Pollard, J.M.; Reboucas, J.S.; Durazo, A.; Kos, I.; Fike, F.; Panni, M.; Gralla, E.B.; Valentine, J.S.; Batinic-Haberle, I.; Gatti, R.A. Radioprotective Effects of Manganese-Containing Superoxide Dismutase Mimics on Ataxia-Telangiectasia Cells. Free Radic. Biol. Med. 2009, 47, 250–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, A.S.; Gaspar, J.; Cabral, M.F.; Rueff, J.; Castro, M.; Batinic-Haberle, I.; Costa, J.; Oliveira, N.G. Protective Role of Ortho-Substituted Mn(III) N-Alkylpyridylporphyrins against the Oxidative Injury Induced by Tert-Butylhydroperoxide. Free Radic. Res. 2010, 44, 430–440. [Google Scholar] [CrossRef]
- Cline, J.M.; Dugan, G.; Bourland, J.D.; Perry, D.L.; Stitzel, J.D.; Weaver, A.A.; Jiang, C.; Tovmasyan, A.; Owzar, K.; Spasojevic, I.; et al. Post-Irradiation Treatment with a Superoxide Dismutase Mimic, MnTnHex-2-PyP5+, Mitigates Radiation Injury in the Lungs of Non-Human Primates after Whole-Thorax Exposure to Ionizing Radiation. Antioxidants 2018, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Batinić-Haberle, I.; Spasojević, I.; Stevens, R.D.; Hambright, P.; Fridovich, I. Manganese(III) Meso-Tetrakis(Ortho-N-Alkylpyridyl)Porphyrins. Synthesis, Characterization, and Catalysis of O2·− Dismutation. J. Chem. Soc. Dalt. Trans. 2002, 13, 2689–2696. [Google Scholar] [CrossRef]
- Cipriano, M.; Pinheiro, P.F.; Sequeira, C.O.; Rodrigues, J.S.; Oliveira, N.G.; Antunes, A.M.M.; Castro, M.; Marques, M.M.; Pereira, S.A.; Miranda, J.P. Nevirapine Biotransformation Insights: An Integrated in Vitro Approach Unveils the Biocompetence and Glutathiolomic Profile of a Human Hepatocyte-like Cell 3d Model. Int. J. Mol. Sci. 2020, 21, 3998. [Google Scholar] [CrossRef]
- Takemoto, S.; Nakamura, Y.; Gyoutoku, H.; Senju, H.; Ogawara, D.; Ikeda, T.; Yamaguchi, H.; Kitazaki, T.; Nakano, H.; Nakatomi, K.; et al. Phase II Trial of a Non-Platinum Triplet for Patients with Advanced Non-Small Cell Lung Carcinoma (NSCLC) Overexpressing ERCC1 Messenger RNA. Thorac. Cancer 2019, 10, 452–458. [Google Scholar] [CrossRef]
- Singh, B.; Patwardhan, R.S.; Jayakumar, S.; Sharma, D.; Sandur, S.K. Oxidative Stress Associated Metabolic Adaptations Regulate Radioresistance in Human Lung Cancer Cells. J. Photochem. Photobiol. B Biol. 2020, 213, 112080. [Google Scholar] [CrossRef]
- Romanowska, M.; Kikawa, K.D.; Fields, J.R.; Maciag, A.; North, S.L.; Shiao, Y.H.; Kasprzak, K.S.; Anderson, L.M. Effects of Selenium Supplementation on Expression of Glutathione Peroxidase Isoforms in Cultured Human Lung Adenocarcinoma Cell Lines. Lung Cancer 2007, 55, 35–42. [Google Scholar] [CrossRef]
- Cai, A.L.; Zeng, W.; Cai, W.L.; Liu, J.L.; Zheng, X.W.; Liu, Y.; Yang, X.C.; Long, Y.; Li, J. Peroxiredoxin-1 Promotes Cell Proliferation and Metastasis through Enhancing Akt/MTOR in Human Osteosarcoma Cells. Oncotarget 2018, 9, 8290–8302. [Google Scholar] [CrossRef]
- Jing, X.; Du, L.; Niu, A.; Wang, Y.; Wang, Y.; Wang, C. Silencing of PRDX2 Inhibits the Proliferation and Invasion of Non-Small Cell Lung Cancer Cells. BioMed Res. Int. 2020, 2020, 1276328. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.G.; Keser, V.; Jackson, C.; Saraiva, N.; Guerreiro, Í.; Almeida, N.; Camões, S.P.; Manguinhas, R.; Castro, M.; Miranda, J.P.; et al. A Multiple Endpoint Approach Reveals Potential in Vitro Anticancer Properties of Thymoquinone in Human Renal Carcinoma Cells. Food Chem. Toxicol. 2020, 136, 111076. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, P.S.; Corvacho, E.; Costa, J.G.; Saraiva, N.; Fernandes, A.S.; Castro, M.; Miranda, J.P.; Oliveira, N.G. The APE1 Redox Inhibitor E3330 Reduces Collective Cell Migration of Human Breast Cancer Cells and Decreases Chemoinvasion and Colony Formation When Combined with Docetaxel. Chem. Biol. Drug Des. 2017, 90, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Manguinhas, R.; Fernandes, A.S.; Costa, J.G.; Saraiva, N.; Camões, S.P.; Gil, N.; Rosell, R.; Castro, M.; Miranda, J.P.; Oliveira, N.G. Impact of the Ape1 Redox Function Inhibitor E3330 in Non-Small Cell Lung Cancer Cells Exposed to Cisplatin: Increased Cytotoxicity and Impairment of Cell Migration and Invasion. Antioxidants 2020, 9, 550. [Google Scholar] [CrossRef] [PubMed]
- Amaro, F.; Pinto, J.; Rocha, S.; Margarida, A.; Miranda-gonçalves, V.; Jer, C.; Henrique, R.; Bastos, M.D.L.; Carvalho, M.; De Pinho, P.G. Volatilomics Reveals Potential Biomarkers for Identification of Renal Cell Carcinoma: An In Vitro Approach. Metabolites 2020, 10, 174. [Google Scholar] [CrossRef]
- Lima, A.R.; Pinto, J.; Azevedo, A.I.; Barros-Silva, D.; Jerónimo, C.; Henrique, R.; de Lourdes Bastos, M.; Guedes de Pinho, P.; Carvalho, M. Identification of a Biomarker Panel for Improvement of Prostate Cancer Diagnosis by Volatile Metabolic Profiling of Urine. Br. J. Cancer 2019, 121, 857–868. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Neumann, C.A.; Cao, J.; Manevich, Y. Peroxiredoxin 1 and Its Role in Cell Signaling. Cell Cycle 2009, 8, 4072–4078. [Google Scholar] [CrossRef] [Green Version]
- Sharapov, M.G.; Goncharov, R.G.; Filkov, G.I.; Trofimenko, A.V.; Boyarintsev, V.V.; Novoselov, V.I. Comparative Study of Protective Action of Exogenous 2-Cys Peroxiredoxins (Prx1 and Prx2) Under Renal Ischemia-Reperfusion Injury. Antioxidants 2020, 9, 680. [Google Scholar] [CrossRef]
- Hanke, N.T.; Imler, E.; Marron, M.T.; Seligmann, B.E.; Garland, L.L.; Baker, A.F. Characterization of Carfilzomib-Resistant Non-Small Cell Lung Cancer Cell Lines. J. Cancer Res. Clin. Oncol. 2018, 144, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lei, L.; Liu, Y. Propofol Improves Sensitivity of Lung Cancer Cells to Cisplatin and Its Mechanism. Med. Sci. Monit. 2020, 26, e919786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umelo, I.A.; De Wever, O.; Kronenberger, P.; Noor, A.; Teugels, E.; Chen, G.; Bracke, M.; De Grève, J. Combined Inhibition of Rho-Associated Protein Kinase and EGFR Suppresses the Invasive Phenotype in EGFR-Dependent Lung Cancer Cells. Lung Cancer 2015, 90, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.X.; Wang, J.; Song, B.; Wei, H.; Lv, W.P.; Tian, L.M.; Li, M.; Lv, S. Establishment and Biological Characteristics of Acquired Gefitinib Resistance in Cell Line NCI-H1975/Gefinitib-Resistant with Epidermal Growth Factor Receptor T790M Mutation. Mol. Med. Rep. 2015, 11, 2767–2774. [Google Scholar] [CrossRef] [Green Version]
- Chung-man Ho, J.; Zheng, S.; A Comhair, S.A.; Farver, C.; Erzurum, S.C. Differential Expression of Manganese Superoxide Dismutase and Catalase in Lung Cancer. Cancer Res. 2001, 61, 8578–8585. [Google Scholar]
- Jiang, H.; Wu, L.; Mishra, M.; Chawsheen, H.A.; Wei, Q. Expression of Peroxiredoxin 1 and 4 Promotes Human Lung Cancer Malignancy. Am. J. Cancer Res. 2014, 4, 445–460. [Google Scholar]
- Costa, J.G.; Saraiva, N.; Guerreiro, P.S.; Louro, H.; Silva, M.J.; Miranda, J.P.; Castro, M.; Batinic-Haberle, I.; Fernandes, A.S.; Oliveira, N.G. Ochratoxin A-Induced Cytotoxicity, Genotoxicity and Reactive Oxygen Species in Kidney Cells: An Integrative Approach of Complementary Endpoints. Food Chem. Toxicol. 2016, 87, 65–76. [Google Scholar] [CrossRef]
- Chaiswing, L.; Yarana, C.; St. Clair, W.; Tovmasyan, A.; Batinic-Haberle, I.; Spasojevic, I.; St. Clair, D. A Redox-Active Mn Porphyrin, MnTnBuOE-2-PyP5+, Synergizes with Carboplatin in Treatment of Chemoresistant Ovarian Cell Line. Oxid. Med. Cell Longev. 2022, 2022, 9664636. [Google Scholar] [CrossRef]
- Batinic-Haberle, I.; Duan, W.; Du, L.; Huang, Z.; Tovmasyan, A.; Cao, Z.; Siamakpour-Reihani, S.; Spasojevic, I.; Sheng, H.; Secord, A.A. MnTnBuOE-2-PyP5+ Ascorbate in the Treatment of Ovarian Cancer. Free. Radic. Biol. Med. 2020, 159, S51. [Google Scholar] [CrossRef]
- Wang, G.; Reed, E.; Li, Q.Q. Molecular Basis of Cellular Response to Cisplatin Chemotherapy in Non-Small Cell Lung Cancer (Review). Oncol. Rep. 2004, 12, 955–965. [Google Scholar] [CrossRef]
- Janfaza, S.; Khorsand, B.; Nikkhah, M.; Zahiri, J. Digging Deeper into Volatile Organic Compounds Associated with Cancer. Biol. Methods Protoc. 2019, 4, bpz014. [Google Scholar] [CrossRef] [PubMed]
- Mochalski, P.; Al-Zoairy, R.; Niederwanger, A.; Unterkofler, K.; Amann, A. Quantitative Analysis of Volatile Organic Compounds Released and Consumed by Rat L6 Skeletal Muscle Cells in Vitro. J. Breath Res. 2014, 8, 046003. [Google Scholar] [CrossRef] [PubMed]
- Klyosov, A.A. Kinetics and Specificity of Human Liver Aldehyde Dehydrogenases toward Aliphatic, Aromatic, and Fused Polycyclic Aldehydes. Biochemistry 1996, 35, 4457–4467. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Sequence | Reference | Accession Number | ||
|---|---|---|---|---|
| CAT | F: TGGAGCTGGTAACCCAGTAGG | [38] | NM_001752.4 | |
| R: CCTTTGCCTTGGAGTATTTGGTA | ||||
| GPX1 | F: CGCTTCCAGACCATTGACATC | [39] | NM_000581.4 | |
| R: CGAGGTGGTATTTTCTGTAAGATCA | ||||
| PRDX1 | F: CCACGGAGATCATTGCTTTCA | [40] | NM_001202431.2 | |
| R: AGGTGTATTGACCCATGCTAGAT | ||||
| PRDX2 | F: CCTTCAAAGAGGTGAAGCTG | [41] | NM_005809.6 | |
| R: GTTGCTGAACGCGATGAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, R.B.; Manguinhas, R.; Costa, J.G.; Saraiva, N.; Gil, N.; Rosell, R.; Camões, S.P.; Batinic-Haberle, I.; Spasojevic, I.; Castro, M.; et al. MnTnHex-2-PyP5+ Displays Anticancer Properties and Enhances Cisplatin Effects in Non-Small Cell Lung Cancer Cells. Antioxidants 2022, 11, 2198. https://doi.org/10.3390/antiox11112198
Soares RB, Manguinhas R, Costa JG, Saraiva N, Gil N, Rosell R, Camões SP, Batinic-Haberle I, Spasojevic I, Castro M, et al. MnTnHex-2-PyP5+ Displays Anticancer Properties and Enhances Cisplatin Effects in Non-Small Cell Lung Cancer Cells. Antioxidants. 2022; 11(11):2198. https://doi.org/10.3390/antiox11112198
Chicago/Turabian StyleSoares, Rita B., Rita Manguinhas, João G. Costa, Nuno Saraiva, Nuno Gil, Rafael Rosell, Sérgio P. Camões, Ines Batinic-Haberle, Ivan Spasojevic, Matilde Castro, and et al. 2022. "MnTnHex-2-PyP5+ Displays Anticancer Properties and Enhances Cisplatin Effects in Non-Small Cell Lung Cancer Cells" Antioxidants 11, no. 11: 2198. https://doi.org/10.3390/antiox11112198