
Discovery of Novel Pterostilbene Derivatives That Might Treat Sepsis by Attenuating Oxidative Stress and Inflammation through Modulation of MAPKs/NF-κB Signaling Pathways


 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemistry
2.1.1. General
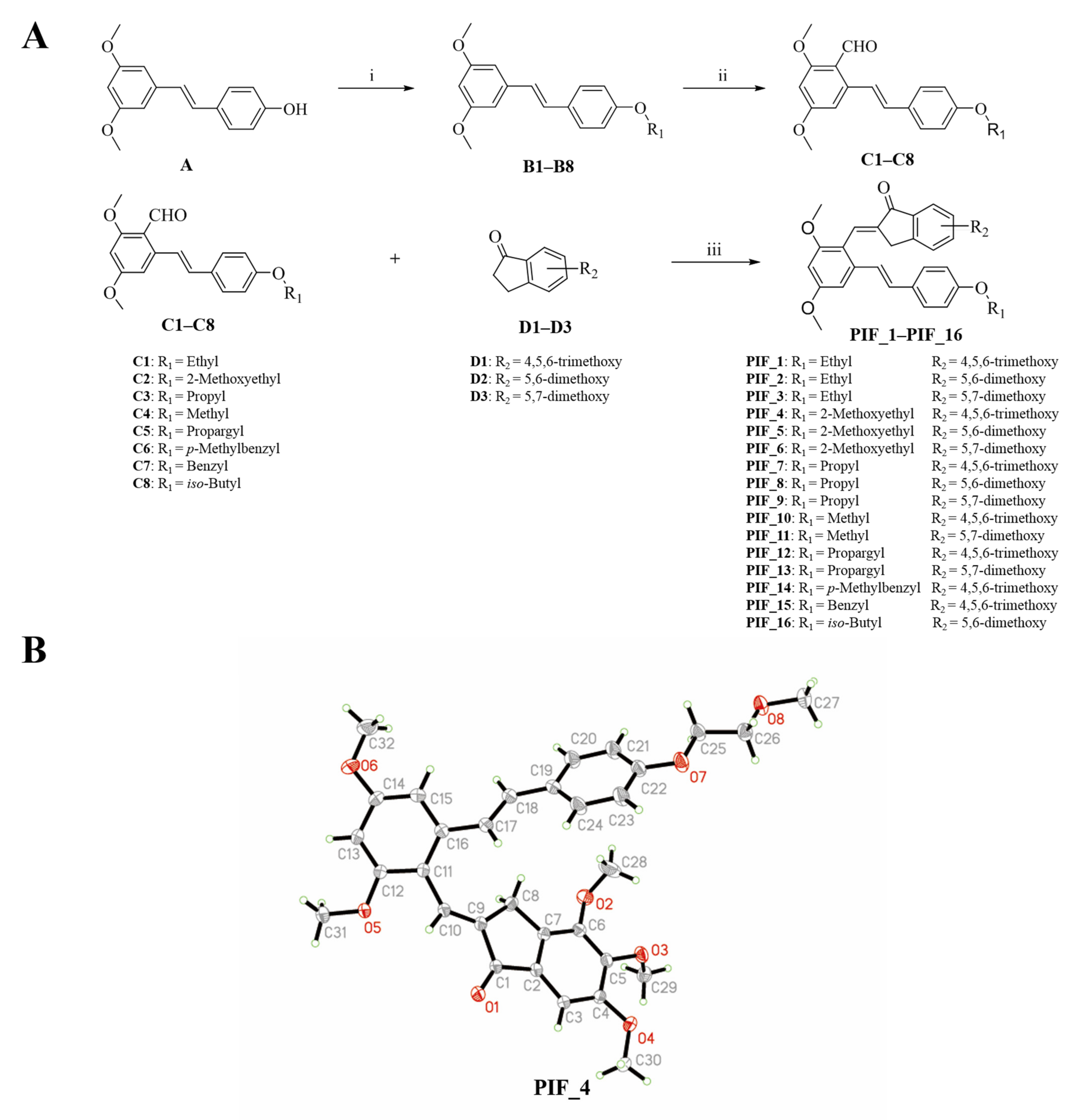
2.1.2. General Procedure for the Preparation of B1–B8
2.1.3. General Procedure for the Preparation of C1–C8
2.1.4. Synthesis of Pterostilbene-Based Indanone Derivatives PIF_1–PIF_16
2.2. Reagents and Cell Culture
2.3. Determination of Cell Viability and Medium NO Levels
2.4. Determination Protein and mRNA Expression
2.5. Determination of ROS Levels in RAW264.7 Cells
2.6. In Vivo Study
2.7. Determination of ROS Levels in the Liver
2.8. TUNEL Assay
2.9. Determination of Serum Biochemical Indicators
2.10. Hematoxylin and Eosin (HE) Staining
2.11. Data and Statistical Analysis
3. Results and Discussion
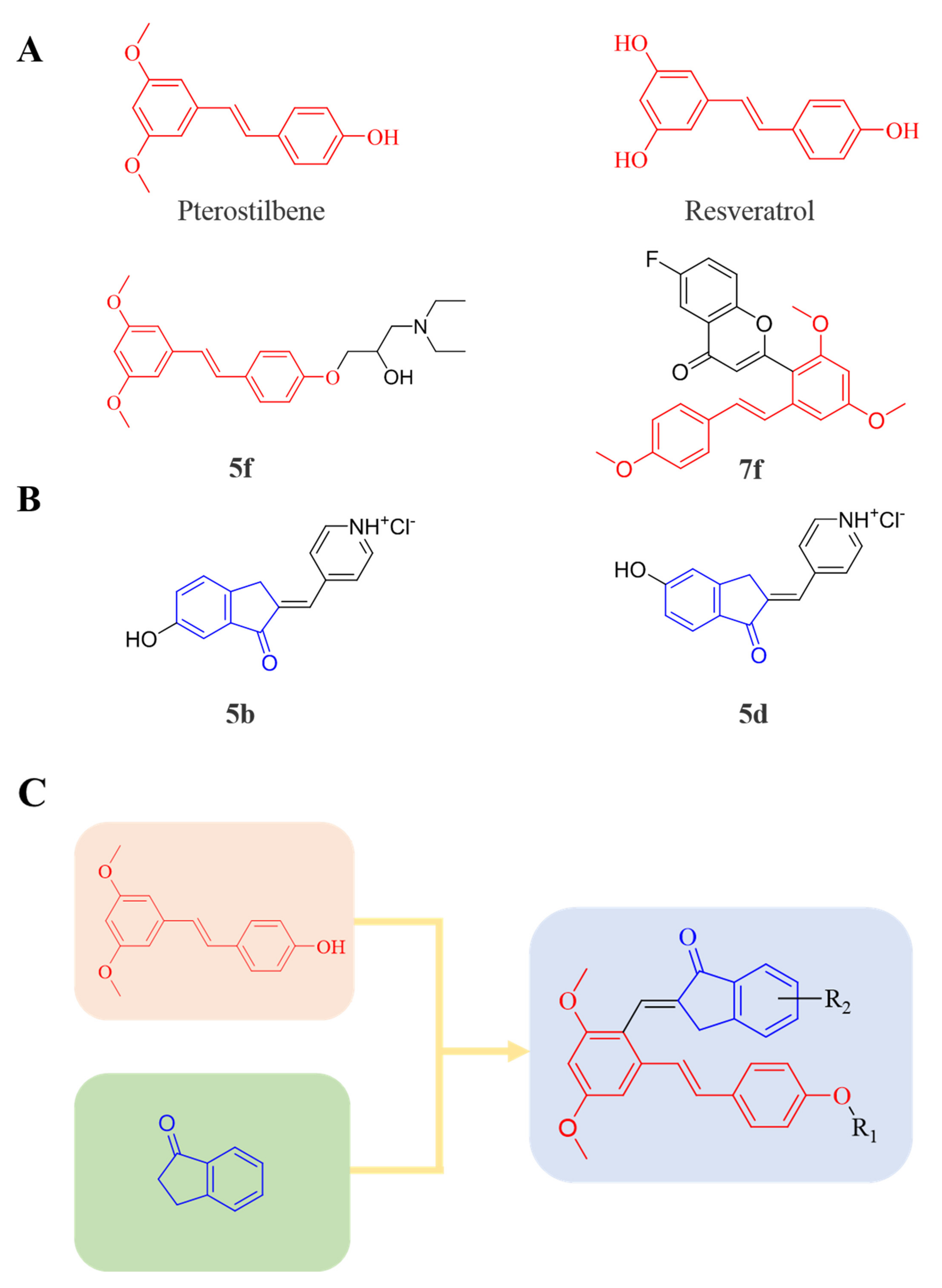
3.1. Chemistry
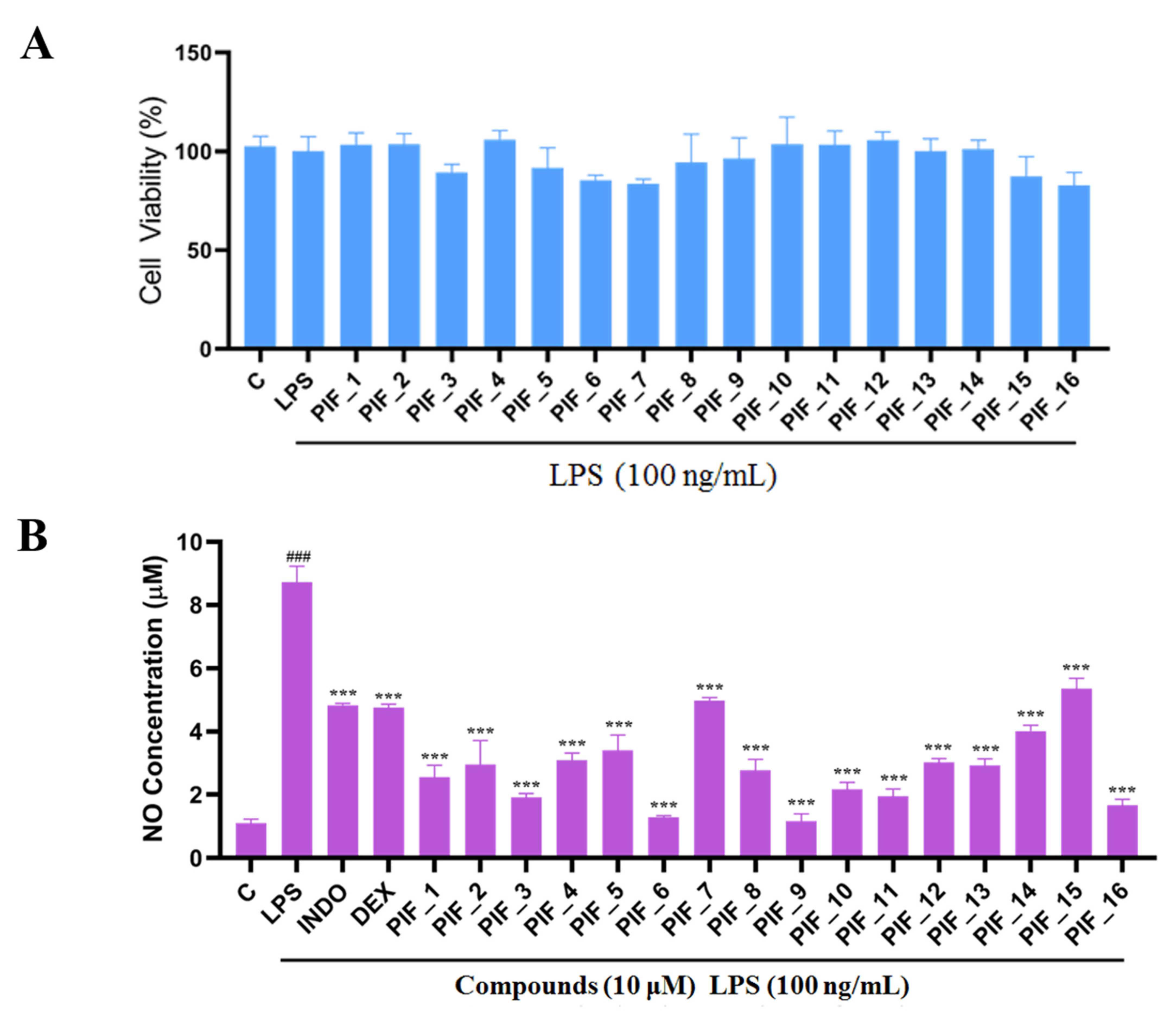
3.2. Intrinsic Cytotoxicity of the Title Compounds against RAW264.7 Cells
3.3. Inhibitory SAR of PIF_1–PIF_16 against LPS-Induced NO Release
3.4. Dose-Dependent Inhibition of NO and Cytokine Production by the Active Compound PIF_9
3.5. Assessment of the Ability of Compound PIF_9 to Inhibit LPS-Induced iNOS and COX-2 Upregulation
3.6. Effect of Compound PIF_9 on LPS-Induced Activation of NF-κB and MAPKs
3.7. Anti-Oxidantive Activities of the Compound PIF_9
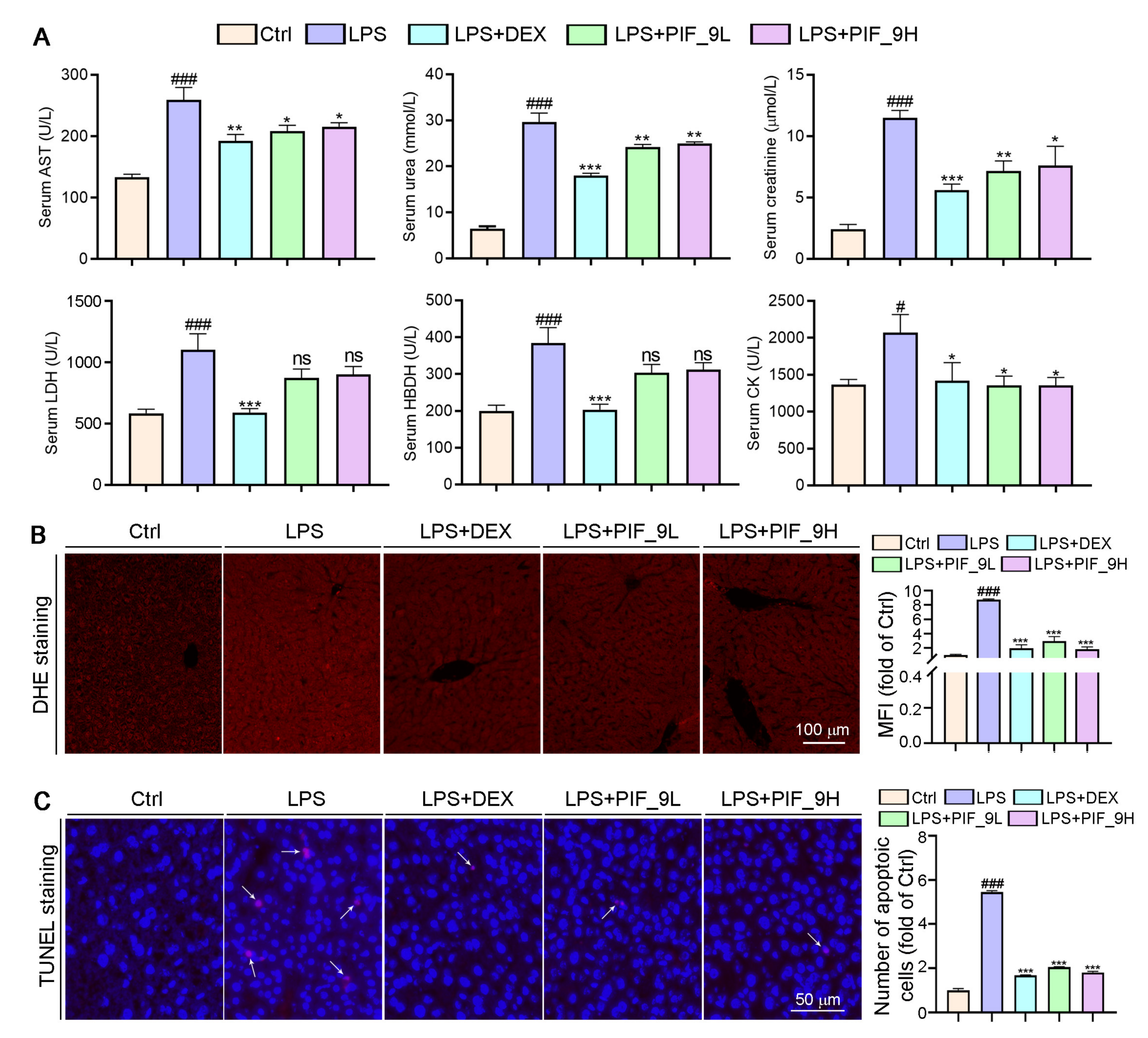
3.8. Compound PIF_9 Improves LPS-Induced Sepsis in C57BL/6J Mice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Z.; Tang, L.; Zhu, H.; Xu, T.; Qiu, C.; Zheng, S.; Gu, Y.; Feng, J.; Zhang, Y.; Liang, G. Design, synthesis, and structure–Activity relationship study of novel indole-2-carboxamide derivatives as anti-inflammatory agents for the treatment of sepsis. J. Med. Chem. 2016, 59, 4637–4650. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Y.; Liu, X.; Fang, Q.; Wang, Z.; Fu, L.; Liu, Z.; Wang, Y.; Zhao, Y.; Li, X.; et al. Discovery of a new inhibitor of myeloid differentiation 2 from cinnamamide derivatives with anti-inflammatory activity in sepsis and acute lung injury. J. Med. Chem. 2016, 59, 2436–2451. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.; Koo, J.Y.; Park, Y.; Oh, K.; Lee, S.; Song, J.-S.; Bae, M.A.; Lim, D.; Lee, D.-S.; Park, S.B. Treatment of sepsis pathogenesis with high mobility group box protein 1-regulating anti-inflammatory agents. J. Med. Chem. 2017, 60, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, L.; Liu, J.; Yuan, S.; Liao, W.; Slominski, A.T.; Li, W.; Żmijewski, M.A.; Chen, J. Discovery of novel 3-hydroxyandrosta-5,7-diene-17-carboxylic acid derivatives as anti-inflammatory bowel diseases (IBD) agents. Eur. J. Med. Chem. 2021, 220, 113468. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Xiao, B.; Chen, L.; Bai, B.; Zhang, Y.; Xu, Z.; Fu, L.; Liu, Z.; Li, X.; Zhao, Y.; et al. Discovery of new MD2-targeted anti-inflammatory compounds for the treatment of sepsis and acute lung injury. Eur. J. Med. Chem. 2017, 139, 726–740. [Google Scholar] [CrossRef]
- Kamisoglu, K.; Haimovich, B.; Calvano, S.E.; Coyle, S.M.; Corbett, S.A.; Langley, R.J.; Kingsmore, S.F.; Androulakis, I.P. Human metabolic response to systemic inflammation: Assessment of the concordance between experimental endotoxemia and clinical cases of sepsis/SIRS. Crit. Care 2015, 19, 71. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.; Huang, M.; Yao, Y. Recent advances in the biology of IL-1 Family cytokines and their potential roles in development of sepsis. Cytokine Growth Factor Rev. 2019, 45, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Pohl, R.; Aslanidis, C. Pro-resolving molecules—New approaches to treat sepsis? IJMS 2017, 18, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Lai, Z.; Xu, X.; Ji, C.; Hu, M.; Zhang, G. Effects of early continuous venovenous hemofiltration on e-selectin, hemodynamic stability, and ventilatory function in patients with septic-shock-induced acute respiratory distress syndrome. BioMed Res. Int. 2016, 2016, 7463130. [Google Scholar] [CrossRef] [PubMed]
- Galley, H.F. Oxidative stress and mitochondrial dysfunction in sepsis. Br. J. Anaesth. 2011, 107, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, L.; Qu, L.; Wang, C.; Peng, W.; Li, S.; Yang, H.; Luo, H.; Yin, F.; Lu, D.; Liu, X.; et al. Identification and optimization of piperlongumine analogues as potential antioxidant and anti-inflammatory agents via activation of Nrf2. Eur. J. Med. Chem. 2021, 210, 112965. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative stress mitigation by antioxidants—An overview on their chemistry and influences on health status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar] [CrossRef]
- Gu, X.; Chen, J.; Zhang, Y.; Guan, M.; Li, X.; Zhou, Q.; Song, Q.; Qiu, J. Synthesis and assessment of phenylacrylamide derivatives as potential anti-oxidant and anti-inflammatory agents. Eur. J. Med. Chem. 2019, 180, 62–71. [Google Scholar] [CrossRef]
- Kellum, J.A.; Kong, L.; Fink, M.P.; Weissfeld, L.A.; Yealy, D.M.; Pinsky, M.R.; Fine, J.; Krichevsky, A.; Delude, R.L.; Angus, D.C. Understanding the inflammatory cytokine response in pneumonia and sepsis. Arch. Intern. Med. 2007, 167, 1655–1663. [Google Scholar] [CrossRef] [Green Version]
- Victor, V.; Rocha, M.; Esplugues, J.; Fuente, M. Role of free radicals in sepsis: Antioxidant therapy. CPD 2005, 11, 3141–3158. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Wei, R.; Deng, A.; Lei, T. Protective effects of ethanolic extracts from artichoke, an edible herbal medicine, against acute alcohol-induced liver injury in mice. Nutrients 2017, 9, 1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, R.L.; Galley, H.F.; Dhillon, J.K.; Webster, N.R. Increased nuclear factor ΚB activation in critically Ill patients who die. Crit. Care Med. 2000, 28, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.L.; Lowes, D.A.; Webster, N.R.; Sheth, C.C.; Gow, N.A.R.; Galley, H.F. Regulation of pentraxin-3 by antioxidants. Br. J. Anaesth. 2009, 103, 833–839. [Google Scholar] [CrossRef] [Green Version]
- Nikhil, K.; Sharan, S.; Palla, S.R.; Sondhi, S.M.; Peddinti, R.K.; Roy, P. Understanding the mode of action of a pterostilbene derivative as anti-inflammatory agent. Int. Immunopharmacol. 2015, 28, 10–21. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Peñalver, P.; Zodio, S.; Lucas, R.; de-Paz, M.V.; Morales, J.C. Neuroprotective and Anti-inflammatory effects of pterostilbene metabolites in human neuroblastoma SH-SY5Y and RAW 264.7 macrophage cells. J. Agric. Food Chem. 2020, 68, 1609–1620. [Google Scholar] [CrossRef]
- Chen, L.Z.; Yao, L.; Jiao, M.M.; Shi, J.B.; Tan, Y.; Ruan, B.F.; Liu, X.H. Novel resveratrol-based flavonol derivatives: Synthesis and anti-inflammatory activity in vitro and in vivo. Eur. J. Med. Chem. 2019, 175, 114–128. [Google Scholar] [CrossRef]
- Zheng, Y.; Qiang, X.; Xu, R.; Song, Q.; Tian, C.; Liu, H.; Li, W.; Tan, Z.; Deng, Y. Design, synthesis and evaluation of pterostilbene β-amino alcohol derivatives as multifunctional agents for Alzheimer’s disease treatment. Bioorganic Chem. 2018, 78, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Abolhasani, H.; Zarghi, A.; Komeili Movahhed, T.; Abolhasani, A.; Daraei, B.; Dastmalchi, S. Design, synthesis and biological evaluation of novel indanone containing spiroisoxazoline derivatives with selective COX-2 inhibition as anticancer agents. Bioorganic Med. Chem. 2021, 32, 115960. [Google Scholar] [CrossRef] [PubMed]
- Kadayat, T.M.; Banskota, S.; Bist, G.; Gurung, P.; Magar, T.B.T.; Shrestha, A.; Kim, J.-A.; Lee, E.-S. Synthesis and biological evaluation of pyridine-linked indanone derivatives: Potential agents for inflammatory bowel disease. Bioorganic Med. Chem. Lett. 2018, 28, 2436–2441. [Google Scholar] [CrossRef]
- Zheng, S.; Cao, P.; Yin, Z.; Wang, X.; Chen, Y.; Yu, M.; Xu, B.; Liao, C.; Duan, Y.; Zhang, S.; et al. Apigenin protects mice against 3,5-diethoxycarbonyl-1,4-dihydrocollidine-induced cholestasis. Food Funct. 2021, 12, 2323–2334. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Li, J.; Xue, C.; Feng, K.; Liu, L.; Zeng, P.; Wang, X.; Chen, Y.; Li, L.; Zhang, Z.; et al. TL1A inhibits atherosclerosis in ApoE-deficient mice by regulating the phenotype of vascular smooth muscle cells. J. Biol. Chem. 2020, 295, 16314–16327. [Google Scholar] [CrossRef]
- Jacobs, A.T.; Ignarro, L.J. Lipopolysaccharide-induced expression of interferon-β mediates the timing of inducible nitric-oxide synthase induction in RAW 264.7 macrophages. J. Biol. Chem. 2001, 276, 47950–47957. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.-H.; Hong, H.-M.; Lin, C.-L.; Jhang, A.-Z.; Tsai, J.-H.; Badmaev, V.; Nagabhushanam, K.; Ho, C.-T.; Chen, W.-J. Se-methylselenocysteine inhibits lipopolysaccharide-induced NF-ΚB activation and INOS induction in RAW 264.7 murine macrophages. Mol. Nutr. Food Res. 2011, 55, 723–732. [Google Scholar] [CrossRef]
- Celes, M.R.N.; Torres-Dueñas, D.; Prado, C.M.; Campos, E.C.; Moreira, J.E.; Cunha, F.Q.; Rossi, M.A. Increased sarcolemmal permeability as an early event in experimental septic cardiomyopathy: A potential role for oxidative damage to lipids and proteins. Shock 2010, 33, 322–331. [Google Scholar] [CrossRef]
- Chen, L.Z.; Wu, J.; Li, K.; Wu, Q.Q.; Chen, R.; Liu, X.H.; Ruan, B.F. Novel phthalide derivatives: Synthesis and anti-inflammatory activity in vitro and in vivo. Eur. J. Med. Chem. 2020, 206, 112722. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol. Cell Physiol. 1996, 271, C1424–C1437. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Hsu, A.; Moore, P.K. Actions and interactions of nitric oxide, carbon monoxide and hydrogen sulphide in the cardiovascular system and in inflammation—A tale of three gases! Pharmacol. Ther. 2009, 123, 386–400. [Google Scholar] [CrossRef]
- Surh, Y.-J.; Chun, K.-S.; Cha, H.-H.; Han, S.S.; Keum, Y.-S.; Park, K.-K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-Inflammatory phytochemicals: Down-regulation of COX-2 and INOS through suppression of NF-kappaB activation. Mutat. Res. 2001, 480–481, 243–268. [Google Scholar] [CrossRef]
- Borish, L.C.; Steinke, J.W. 2. Cytokines and chemokines. J. Allergy Clin. Immunol. 2003, 111, S460–S475. [Google Scholar] [CrossRef]
- Singh, G.; Singh, G.; Bhatti, R.; Gupta, M.; Kumar, A.; Sharma, A.; Singh Ishar, M.P. Indolyl-isoxazolidines attenuate LPS-stimulated pro-inflammatory cytokines and increase survival in a mouse model of sepsis: Identification of potent lead. Eur. J. Med. Chem. 2018, 153, 56–64. [Google Scholar] [CrossRef]
- Krakauer, T. Molecular therapeutic targets in inflammation: Cyclooxygenase and NF-kappaB. CDTIA 2004, 3, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2020, 40, 158–189. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Rahman, I.; MacNee, W. Role of transcription factors in inflammatory lung diseases. Thorax 1998, 53, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Aderem, A.; Ulevitch, R.J. Toll-like receptors in the induction of the innate immune response. Nature 2000, 406, 782–787. [Google Scholar] [CrossRef]
- Patel, B.V.; Wilson, M.R.; O’Dea, K.P.; Takata, M. TNF-induced death signaling triggers alveolar epithelial dysfunction in acute lung injury. J.Immunol. 2013, 190, 4274–4282. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Xu, T.; Xu, F.; Zhang, Y.; Liu, Z.; Chen, W.; Fu, W.; Dai, Y.; Zhao, Y.; Feng, J.; et al. Development of resveratrol-curcumin hybrids as potential therapeutic agents for inflammatory lung diseases. Eur. J. Med. Chem. 2017, 125, 478–491. [Google Scholar] [CrossRef]
- Deng, S.; Gu, B.; Yu, Z.; Shen, Z.; Ren, H. MIR210HG aggravates sepsis-induced inflammatory response of proximal tubular epithelial cell via the NF-ΚB signaling pathway. Yonsei Med. J. 2021, 62, 461. [Google Scholar] [CrossRef]
- Yang, H.-Z.; Wang, J.-P.; Mi, S.; Liu, H.-Z.; Cui, B.; Yan, H.-M.; Yan, J.; Li, Z.; Liu, H.; Hua, F.; et al. TLR4 activity is required in the resolution of pulmonary inflammation and fibrosis after acute and chronic lung injury. Am. J. Pathol. 2012, 180, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Ve, T.; Vajjhala, P.R.; Hedger, A.; Croll, T.; DiMaio, F.; Horsefield, S.; Yu, X.; Lavrencic, P.; Hassan, Z.; Morgan, G.P.; et al. Structural basis of TIR-domain-assembly formation in MAL- and MyD88-dependent TLR4 signaling. Nat. Struct. Mol. Biol. 2017, 24, 743–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, K.; Nappo, F.; Marfella, R.; Giugliano, G.; Giugliano, F.; Ciotola, M.; Quagliaro, L.; Ceriello, A.; Giugliano, D. Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: Role of oxidative stress. Circulation 2002, 106, 2067–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gongora, M.C.; Lob, H.E.; Landmesser, U.; Guzik, T.J.; Martin, W.D.; Ozumi, K.; Wall, S.M.; Wilson, D.S.; Murthy, N.; Gravanis, M.; et al. Loss of extracellular superoxide dismutase leads to acute lung damage in the presence of ambient air. Am. J. Pathol. 2008, 173, 915–926. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Chen, J.; Xing, C.; Huang, A.; Zhuang, Y.; Yang, F.; Zhang, C.; Hu, G.; Mao, Y.; Cao, H. Molybdenum induces mitochondrial oxidative damage in kidney of goats. Biol. Trace Elem. Res. 2020, 197, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Li, S.; Li, S. The role of the liver in sepsis. Int. Rev. Immunol. 2014, 33, 498–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doggrell, S.A. Suramin: Potential in acute liver failure. Expert Opin. Investig. Drugs 2004, 13, 1361–1363. [Google Scholar] [CrossRef] [PubMed]
- Doğanyiğit, Z.; Okan, A.; Kaymak, E.; Pandır, D.; Silici, S. Investigation of Protective effects of apilarnil against lipopolysaccharide induced liver injury in rats via TLR 4/ HMGB-1/ NF-ΚB pathway. Biomed. Pharmacother. 2020, 125, 109967. [Google Scholar] [CrossRef]
- Kirkinezos, I.G.; Moraes, C.T. Reactive oxygen species and mitochondrial diseases. Semin. Cell Dev. Biol. 2001, 12, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.-Q.; Li, Z.; Xie, W.-R.; Liu, C.-M.; Liu, S.-S. Quercetin protects mouse liver against CCl4-induced inflammation by the TLR2/4 and MAPK/NF-ΚB pathway. Int. Immunopharmacol. 2015, 28, 531–539. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
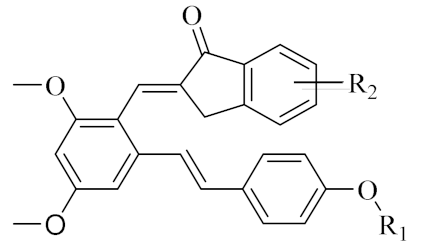 PIF_1–PIF_16 | |||||
|---|---|---|---|---|---|
| Compd. | R1 | R2 | Compd. | R1 | R2 |
| PIF_1 | 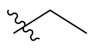 | 4,5,6-OCH3 | PIF_9 | 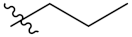 | 5,7-OCH3 |
| PIF_2 | 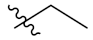 | 5,6-OCH3 | PIF_10 |  | 4,5,6-OCH3 |
| PIF_3 | 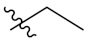 | 5,7-OCH3 | PIF_11 |  | 5,7-OCH3 |
| PIF_4 |  | 4,5,6-OCH3 | PIF_12 |  | 4,5,6-OCH3 |
| PIF_5 |  | 5,6-OCH3 | PIF_13 |  | 5,7-OCH3 |
| PIF_6 | 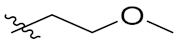 | 5,7-OCH3 | PIF_14 | 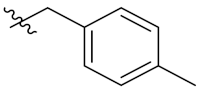 | 4,5,6-OCH3 |
| PIF_7 |  | 4,5,6-OCH3 | PIF_15 | 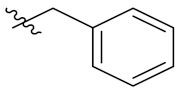 | 4,5,6-OCH3 |
| PIF_8 | 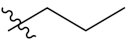 | 5,6-OCH3 | PIF_16 | 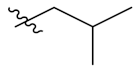 | 5,6-OCH3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, M.; Zou, T.; Yang, X.; Zhang, Z.; Cao, P.; Han, J.; Duan, Y.; Ruan, B.-F.; Li, Q.-S. Discovery of Novel Pterostilbene Derivatives That Might Treat Sepsis by Attenuating Oxidative Stress and Inflammation through Modulation of MAPKs/NF-κB Signaling Pathways. Antioxidants 2021, 10, 1333. https://doi.org/10.3390/antiox10091333
Fang M, Zou T, Yang X, Zhang Z, Cao P, Han J, Duan Y, Ruan B-F, Li Q-S. Discovery of Novel Pterostilbene Derivatives That Might Treat Sepsis by Attenuating Oxidative Stress and Inflammation through Modulation of MAPKs/NF-κB Signaling Pathways. Antioxidants. 2021; 10(9):1333. https://doi.org/10.3390/antiox10091333
Chicago/Turabian StyleFang, Mengyuan, Tingfeng Zou, Xiaoxiao Yang, Zhen Zhang, Peichang Cao, Jihong Han, Yajun Duan, Ban-Feng Ruan, and Qing-Shan Li. 2021. "Discovery of Novel Pterostilbene Derivatives That Might Treat Sepsis by Attenuating Oxidative Stress and Inflammation through Modulation of MAPKs/NF-κB Signaling Pathways" Antioxidants 10, no. 9: 1333. https://doi.org/10.3390/antiox10091333
APA StyleFang, M., Zou, T., Yang, X., Zhang, Z., Cao, P., Han, J., Duan, Y., Ruan, B.-F., & Li, Q.-S. (2021). Discovery of Novel Pterostilbene Derivatives That Might Treat Sepsis by Attenuating Oxidative Stress and Inflammation through Modulation of MAPKs/NF-κB Signaling Pathways. Antioxidants, 10(9), 1333. https://doi.org/10.3390/antiox10091333