
Imbalance of Systemic Redox Biomarkers in Children with Epilepsy: Role of Ferroptosis

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Participants’ Enrollment
2.2. Blood Sample Collection
2.3. Glutathione Assay
2.4. Determination of Plasma 4-HNE and 15(S)-HETE Content
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Western Blot Analysis
2.7. GPX4 Enzyme Activity Assay
2.8. GST Enzyme Activity Assay
2.9. Statistical Analysis
3. Results
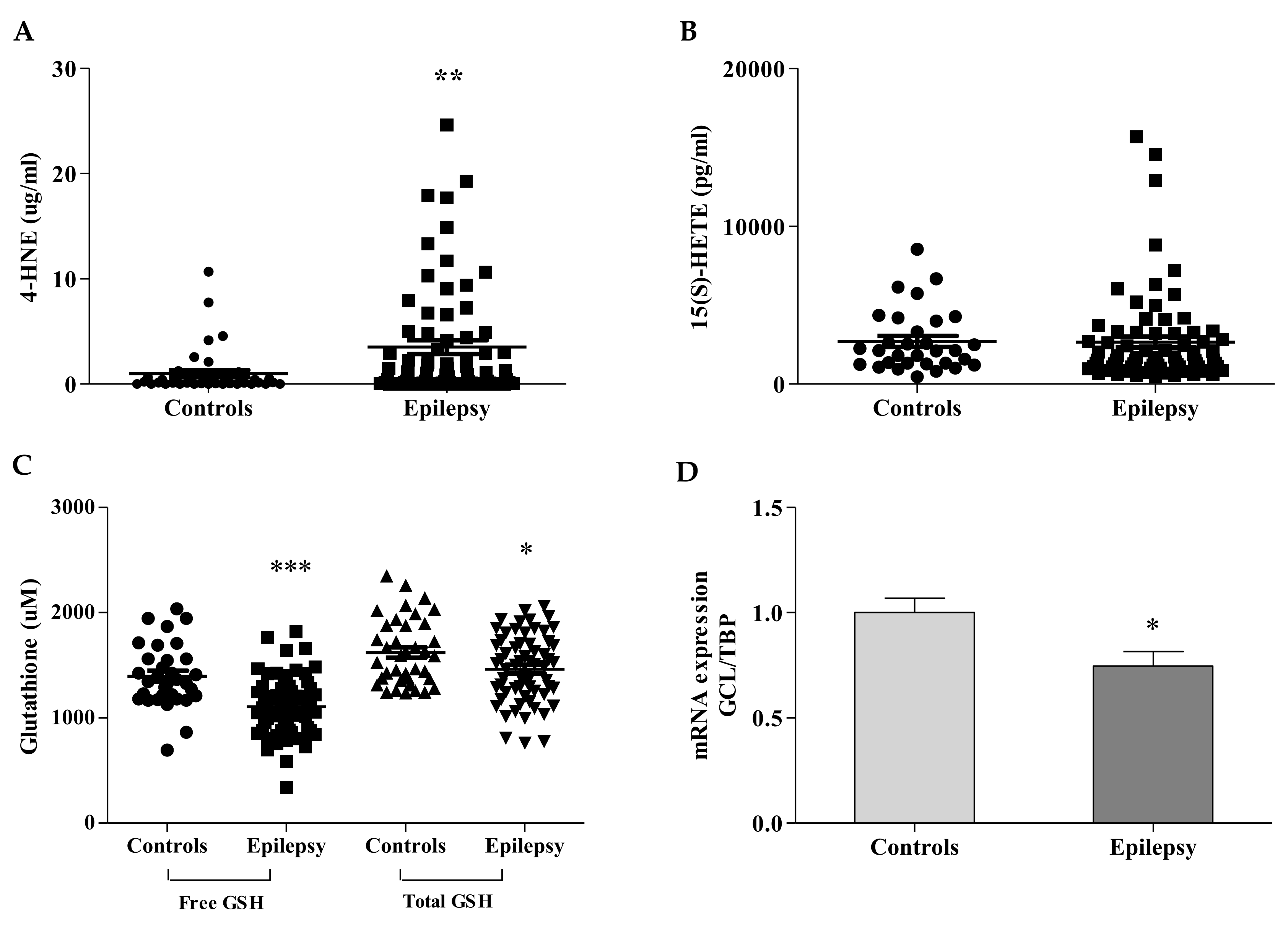
3.1. Ferroptosis Bio-Markers Are Imbalanced in Children with Epilepsy
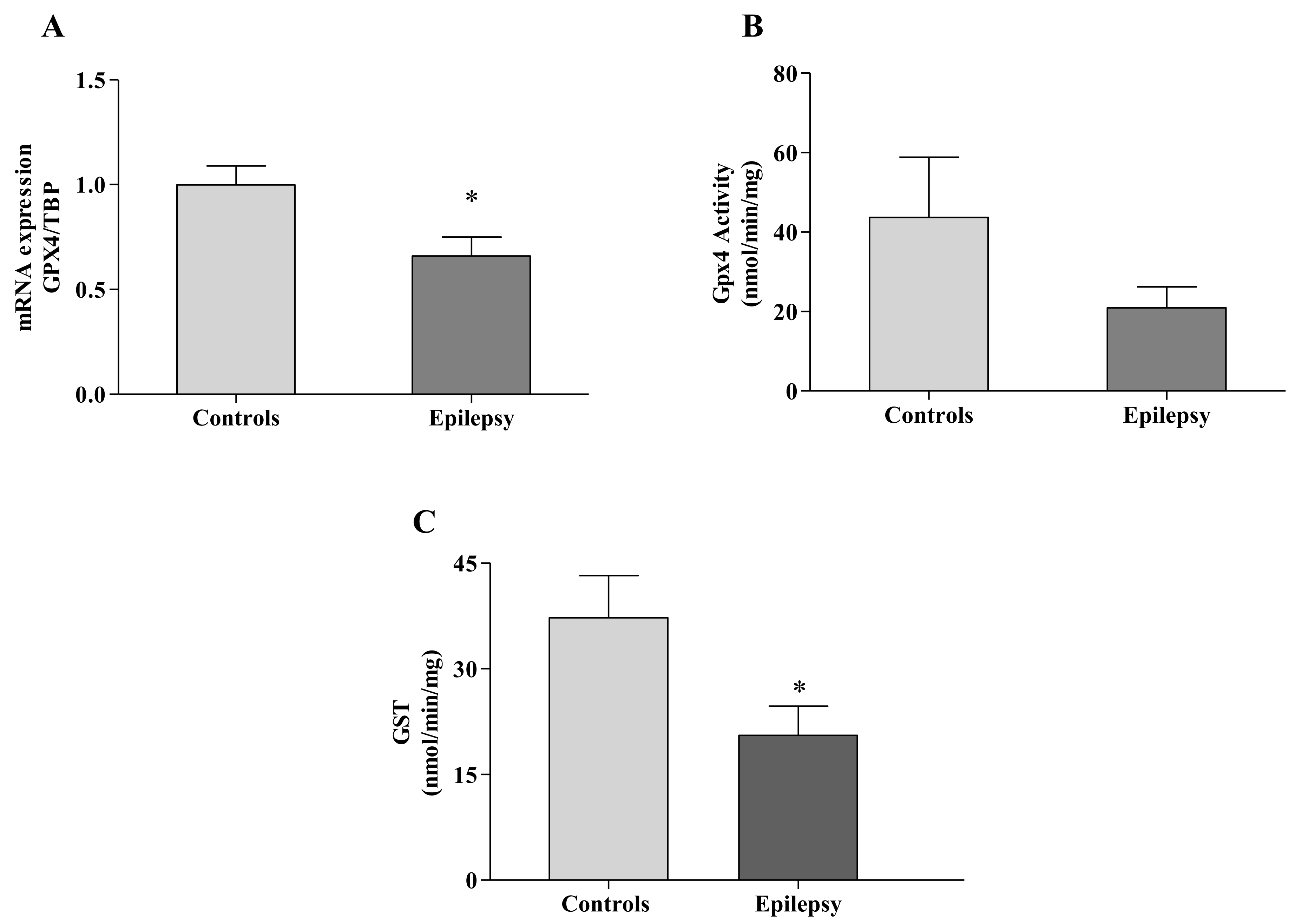
3.2. GPX4 Reflects the 4-HNE/GSH Blood Imbalance
3.3. GST Fails to Detoxify Ferroptosis By-Products in Epilepsy
3.4. NRF2 at the Crossroad of Redox Balance and Ferroptosis in Epilepsy
3.5. NOX2: A Primer of Ferroptosis in Epilepsy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shin, E.J.; Jeong, J.H.; Chung, Y.H.; Kim, W.K.; Ko, K.H.; Bach, J.H.; Hong, J.S.; Yoneda, Y.; Kim, H.C. Role of oxidative stress in epileptic seizures. Neurochem. Int. 2011, 59, 122–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shekh-Ahmad, T.; Kovac, S.; Abramov, A.Y.; Walker, M.C. Reactive oxygen species in status epilepticus. Epilepsy Behav. 2019, 101, 106410. [Google Scholar] [CrossRef]
- Ambrogini, P.; Torquato, P.; Bartolini, D. Excitotoxicity, neuroinflammation and oxidant stress as molecular bases of epileptogenesis and epilepsy-derived neurodegeneration: The role of vitamin E. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1098–1112. [Google Scholar] [CrossRef] [PubMed]
- Olowe, R.; Sandouka, S.; Saadi, A.; Shekh-Ahmad, T. Approaches for Reactive Oxygen Species and Oxidative Stress Quantification in Epilepsy. Antioxidants 2020, 9, 990. [Google Scholar] [CrossRef]
- Kalita, J.; Misra, U.K.; Singh, L.S.; Tiwari, A. Oxidative stress in status epilepticus: A clinical-radiological correlation. Brain Res. 2019, 1704, 85–93. [Google Scholar] [CrossRef]
- Mao, X.Y.; Zhou, H.H.; Jin, W.L. Redox-Related Neuronal Death and Crosstalk as Drug Targets: Focus on Epilepsy. Front. Neurosci. 2019, 13, 512. [Google Scholar] [CrossRef] [Green Version]
- Kajarabille, N.; Latunde-Dada, G.O. Programmed Cell-Death by Ferroptosis: Antioxidants as Mitigators. Int. J. Mol. Sci. 2019, 20, 4968. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.Y.; Zhou, H.H.; Jin, W.L. Ferroptosis Induction in Pentylenetetrazole Kindling and Pilocarpine-Induced Epileptic Seizures in Mice. Front. Neurosci. 2019, 13, 721. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Friedmann Angeli, J.P.; Schneider, M.; Proneth, B.; Tyurina, Y.Y.; Tyurin, V.A.; Hammond, V.J.; Herbach, N.; Aichler, M.; Walch, A.; Eggenhofer, E.; et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat. Cell Biol. 2014, 16, 1180–1191. [Google Scholar] [CrossRef] [Green Version]
- Ingold, I.; Berndt, C.; Schmitt, S.; Doll, S.; Poschmann, G.; Buday, K.; Roveri, A.; Peng, X.; Porto Freitas, F.; Seibt, T.; et al. Selenium Utilization by GPX4 Is Required to Prevent Hydroperoxide-Induced Ferroptosis. Cell 2017, 172, 409–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, G.; Riahi, Y.; Sasson, S. Lipid peroxidation of poly-unsaturated fatty acids in normal and obese adipose tissues. Arch. Physiol. Biochem. 2011, 117, 131–139. [Google Scholar] [CrossRef]
- Pecorelli, A.; Natrella, F.; Belmonte, G.; Miracco, C.; Cervellati, F.; Ciccoli, L.; Mariottini, A.; Rocchi, R.; Vatti, G.; Bua, A.; et al. NADPH oxidase activation and 4-hydroxy-2-nonenal/aquaporin-4 adducts as possible new players in oxidative neuronal damage presents in drug resistant epilepsy. Biochim. Biophys. Acta 2015, 1852, 507–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Q.; Zeng, C.; Dong, L.; Wu, Y.; Huang, Q.; Wu, Y. Inhibition of ferroptosis processes ameliorates cognitive impairment in kainic acid-induced temporal lobe epilepsy in rats. Am. J. Transl. Res. 2019, 11, 875–884. [Google Scholar]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Niki, E. Biomarkers of lipid peroxidation in clinical material. Biochim. Biophys. Acta 2014, 1840, 809–817. [Google Scholar] [CrossRef]
- Forcina, G.C.; Dixon, S.J. GPX4 at the Crossroads of Lipid Homeostasis and Ferroptosis. Proteomics 2019, 19, e1800311. [Google Scholar] [CrossRef]
- Huang, Y.; Li, W.; Kong, A.N. Anti-oxidative stress regulator NF-E2-related factor 2 mediates the adaptive induction of antioxidant and detoxifying enzymes by lipid peroxidation metabolite 4-hydroxynonenal. Cell Biosci. 2012, 2, 40. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wu, Y.; Zhang, G.; Fang, H.; Wang, H.; Zang, H.; Xie, T.; Wang, W. Activation of Nrf2-ARE signal pathway protects the brain from damage induced by epileptic seizure. Brain Res. 2014, 28, 54–61. [Google Scholar] [CrossRef]
- Shekh-Ahmad, T.; Eckel, R.; Dayalan Naidu, S.; Higgins, M.; Yamamoto, M.; Dinkova-Kostova, A.T.; Kovac, S.; Abramov, A.Y.; Walker, M.C. KEAP1 inhibition is neuroprotective and suppresses the development of epilepsy. Brain 2018, 141, 1390–1403. [Google Scholar] [CrossRef] [PubMed]
- Shekh-Ahmad, T.; Lieb, A.; Kovac, S.; Gola, L.; Christian Wigley, W.; Abramov, A.Y.; Walker, M.C. Combination antioxidant therapy prevents epileptogenesis and modifies chronic epilepsy. Redox Biol. 2019, 26, 101278. [Google Scholar] [CrossRef]
- Yang, N.; Guan, Q.W.; Chen, F.H.; Xia, Q.X.; Yin, X.X.; Zhou, H.H.; Mao, X.Y. Antioxidants Targeting Mitochondrial Oxidative Stress: Promising Neuroprotectants for Epilepsy. Oxid. Mes. Cell Longev. 2020, 25, 6687185. [Google Scholar] [CrossRef]
- Scheffer, I.E.; Berkovic, S.; Capovilla, G.; Connolly, M.B.; French, J.; Guilhoto, L.; Hirsch, E.; Jain, S.; Mathern, G.W.; Moshé, S.L.; et al. ILAE classification of the epilepsies: Position paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 512–521. [Google Scholar] [CrossRef] [Green Version]
- Kwan, P.; Arzimanoglou, A.; Berg, A.T.; Brodie, M.J.; Allen Hauser, W.; Mathern, G.; Moshé, S.L.; Perucca, E.; Wiebe, S.; French, J. Definition of drug resistant epilepsy: Consensus proposal by the ad hoc Task Force of the ILAE Commission on Therapeutic Strategies. Epilepsia 2010, 51, 1069–1077. [Google Scholar] [CrossRef]
- Mayr, L.; Grabherr, F.; Schwärzler, J.; Reitmeier, I.; Sommer, F.; Gehmacher, T.; Niederreiter, L.; He, G.W.; Ruder, B.; Kunz, K.T.R.; et al. Dietary lipids fuel GPX4-restricted enteritis resembling Crohn’s disease. Nat. Comm. 2020, 11, 1775. [Google Scholar] [CrossRef] [Green Version]
- Balogh, L.M.; Atkins, W.M. Interactions of glutathione transferases with 4-hydroxynonenal. Drug Metab. Rev. 2011, 43, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Jang, B.G.; Choi, B.Y.; Kim, H.S.; Sohn, M.; Chung, T.N.; Choi, H.C.; Song, H.K.; Suh, S.W. Post-treatment of an NADPH oxidase inhibitor prevents seizure induced neuronal death. Brain Res. 2013, 1499, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Pestana, R.R.F.; Kinjo, E.R.; Hernandes, M.S.; Britto, L.R.G. Reactive oxygen species generated by NADPH oxidase are involved in neurodegeneration in the pilocarpine model of temporal lobe epilepsy. Neurosci. Lett. 2010, 484, 187–191. [Google Scholar] [CrossRef]
- Malkov, A.; Ivanov, A.I.; Latyshkova, A.; Bregestovski, P.; Zilberter, M.; Zilberter, Y. Activation of nicotinamide adenine dinucleotide phosphate oxidase is the primary trigger of epileptic seizures in rodent models. Ann. Neurol. 2019, 85, 907–920. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.P.; Henry, Y.K.; Henkel, J.S.; Smith, R.G.; Appel, S.H. Increased lipid peroxidation in sera of ALS patients: A potential biomarker of disease burden. Neurology 2004, 62, 1758–1765. [Google Scholar] [CrossRef] [PubMed]
- Brodovitch, A.; Boucraut, J.; Delmont, E.; Parlanti, A.; Grapperon, A.M.; Attarian, S.; Verschueren, A. Combination of serum and CSF neurofilament-light and neuroinflammatory biomarkers to evaluate ALS. Sci. Rep. 2021, 11, 703. [Google Scholar] [CrossRef]
- Walker, L.E.; Janigro, D.; Heinemann, U.; Riikonen, R.; Bernard, C.; Patel, M. WONOEP appraisal: Molecular and cellular biomarkers for epilepsy. Epilepsia 2016, 57, 1354–1362. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.G.; Mu, R.Z.; Liu, Y.; Jiang, D.; Jia, T.T.; Huang, Y.J. Increased Serum S100B Levels in Patients With Epilepsy: A Systematic Review and Meta-Analysis Study. Front. Neurosci. 2019, 13, 456. [Google Scholar] [CrossRef] [PubMed]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef]
- Abdallah, D.M. Anticonvulsant potential of the peroxisome proliferator activated receptor gamma agonist pioglitazone in pentylenetetrazole-induced acute seizures and kindling in mice. Brain Res. 2010, 1351, 246–253. [Google Scholar] [CrossRef]
- Mao, X.; Cao, Y.; Li, X.; Yin, J.; Wang, Z.; Zhang, Y.; Mao, C.; Fan, K.; Zhou, H.; Cai, J.; et al. Baicalein ameliorates cognitive deficits in epilepsy-like tremor rat. Neurol. Sci. 2014, 35, 1261–1268. [Google Scholar] [CrossRef]
- Zou, X.; Jiang, S.; Wu, Z.; Shi, Y.; Cai, S.; Zhu, R.; Chen, L. Effectiveness of deferoxamine on ferric chloride-induced epilepsy in rats. Brain Res. 2017, 1658, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Simeone, T.A.; Matthews, S.A.; Simeone, K.A. Synergistic protection against acute flurothyl-induced seizures by adjuvant treatment of the ketogenic diet with the type 2 diabetes drug pioglitazone. Epilepsia 2017, 58, 1440–1450. [Google Scholar] [CrossRef]
- Dal-Pizzol, F.; Klamt, F.; Vianna, M.M.; Schröder, N.; Quevedo, J.; Benfato, M.S.; Moreira, J.C.; Walz, R. Lipid peroxidation in hippocampus early and late after status epilepticus induced by pilocarpine or kainic acid in Wistar rats. Neurosci. Lett. 2000, 291, 179–182. [Google Scholar] [CrossRef]
- Liang, L.P.; Ho, Y.S.; Patel, M. Mitochondrial superoxide production in kainate-induced hippocampal damage. Neuroscience 2000, 101, 563–570. [Google Scholar] [CrossRef]
- Cárdenas-Rodríguez, N.; Coballase-Urrutia, E.; Rivera-Espinosa, L.; Romero-Toledo, A.; Sampieri, A., III; Ortega-Cuellar, D.; Montesinos-Correa, H.; Floriano-Sánchez, E.; Carmona-Aparicio, L. Modulation of antioxidant enzymatic activities by certain antiepileptic drugs (valproic acid, oxcarbazepine, and topiramate): Evidence in humans and experimental models. Oxid. Med. Cell Longev. 2013, 2013, 598493. [Google Scholar] [CrossRef] [PubMed]
- Turkdogan, D.; Toplan, S.; Karakoc, Y. Lipid peroxidation and antioxidative enzyme activities in childhood epilepsy. J. Child Neurol. 2002, 17, 673–676. [Google Scholar] [CrossRef]
- Martinc, B.; Grabnar, I.; Vovk, T. Antioxidants as a preventive treatment for epileptic process: A review of the current status. Curr. Neuropharmacol. 2014, 12, 527–550. [Google Scholar] [CrossRef] [Green Version]
- Yüksel, A.; Cengiz, M.; Seven, M.; Ulutin, T. Erythrocyte glutathione, glutathione peroxidase, superoxide dismutase and serum lipid peroxidation in epileptic children with valproate and carbamazepine monotherapy. J. Basic Clin. Physiol. Pharmacol. 2000, 11, 73–81. [Google Scholar] [CrossRef]
- Yiş, U.; Seçkin, E.; Kurul, S.H.; Kuralay, F.; Dirik, E. Effects of epilepsy and valproic acid on oxidant status in children with idiopathic epilepsy. Epilepsy Res. 2009, 84, 232–237. [Google Scholar] [CrossRef]
- Chen, L.; Hambright, W.S.; Na, R.; Ran, Q. Ablation of the ferroptosis inhibitor glutathione peroxidase 4 in neurons results in rapid motor neuron degeneration and paralysis. J. Biol. Chem. 2015, 290, 28097–28106. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.C.; Mears, A.J.; Bunker, R.; Ahmed, A.; MacKenzie, M.; Schwartzentruber, J.A.; Beaulieu, C.L.; Ferretti, E. Mutations in the enzyme glutathione peroxidase 4 cause Sedaghatian-type spondylometaphyseal dysplasia. J. Med. Genet. 2014, 51, 470–474. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Li, Q.Y.; Chang, L.Y.; Crapo, J.; Liang, L.P. Activation of NADPH oxidase and extracellular superoxide production in seizure-induced hippocampal damage. J. Neurochem. 2005, 92, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, M.; Ko, D.G.; Choi, B.Y.; Suh, S.W. The Role of NADPH Oxidase in Neuronal Death and Neurogenesis after Acute Neurological Disorders. Antioxidants 2021, 10, 739. [Google Scholar] [CrossRef]
- Shang, W.; Liu, W.H.; Zhao, X.H.; Sun, Q.J.; Bi, J.Z.; Chi, Z.F. Expressions of glutathione S-transferase alpha, mu, and pi in brains of medically intractable epileptic patients. BMC Neurosci. 2008, 18, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persike, D.S.; Marques-Carneiro, J.E.; Stein, M.L.L. Altered Proteins in the Hippocampus of Patients with Mesial Temporal Lobe Epilepsy. Pharmaceuticals 2018, 11, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, I.M.; Tomé, A.R.; Saldanha, G.B.; Ferreira, P.M.; Militão, G.C.; Freitas, R.M. Oxidative stress in the hippocampus during experimental seizures can be ameliorated with the antioxidant ascorbic acid. Oxid. Med. Cell Longev. 2009, 2, 214–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diniz, T.C.; Silva, J.C.; de Lima-Saraiva, S.R.; Ribeiro, F.P.; Pacheco, A.G.; de Freitas, R.M.; Quintans-Júnior, L.J.; Quintans Jde, S.; Mendes, R.L.; Almeida, J.R. The role of flavonoids on oxidative stress in epilepsy. Oxid. Med. Cell Longev. 2015, 2015, 171756. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Controls | Patients |
|---|---|---|
| Age at enrollment, mean (range) | 6 years (0.6–15) | 4 years (0.42–12) |
| Age at epilepsy onset, mean (range) | 1.5 years (0–7) | |
| Sex, n (%) | ||
| Male | 23 (52%) | 42 (51%) |
| Female | 21 (48%) | 41 (49%) |
| Drug-resistant, n (%) | 29 (35%) | |
| Drug-responsive, n (%) | 54 (65%) | |
| Focal epilepsy, n (%) | 52 (63%) | |
| Generalized epilepsy, n (%) | 17 (20%) | |
| DEE, n (%) | 14 (17%) |
| Genes | Sequence (5′ → 3′) | |
|---|---|---|
| GCL *601176 | Fw-TTGCCTCCTGCTGTGTGATG | Rv-ATCATTGTGAGTCAACAGCTGTATGTC |
| GPX4 *138322 | Fw-GCTCCATGCACGAGTTTTCC | Rv-ACTTCGGTCTTGCCTCACTG |
| NRF2 *600492 | Fw-ACACGGTCCACAGCTCATC | Rv-TGTCAATCAAATCCATGTCCTG |
| TBP *605521 | Fw-CCGAAACGCCGAATATAATCC | Rv-AAATCAGTGCCGTGGTTCGT |
| Biomarkers | 4-HNE, Mean ± SEM | Free GSH, Mean ± SEM | Total GSH, Mean ± SEM |
|---|---|---|---|
| Controls | 0.98 ± 0.32 | 1396 ± 52 | 1621 ± 50 |
| Focal epilepsy | 3.44 ± 0.80 ** | 1088 ± 51 *** | 1438 ± 55 * |
| Generalized epilepsy | 2.97 ± 1.51 * | 1235 ± 65 | 1492 ± 122 |
| DEE | 2.42 ± 1.00 | 1143 ± 84 * | 1514 ± 135 |
| Drug-resistant epilepsy | 2.45 ± 0.68 * | 1087 ± 75 ** | 1427 ± 87 * |
| Drug-responsive Epilepsy | 3.52 ± 0.84 ** | 1135 ± 45 *** | 1470 ± 54 * |
| Age at enrollment (months) | |||
| 0–12 | 5.64 ± 1.81 *** | 1062 ± 74 ** | 1485 ± 107 |
| 12–24 | 3.235 ± 1.57 * | 1161 ± 47 * | 1472 ± 118 |
| 24–48 | 4.15 ± 1.40 ** | 1121 ± 71 ** | 1514 ± 81 |
| 48–144 | 2.52 ± 0.69 | 1117 ± 69 ** | 1436 ± 72 * |
| Epilepsy onset (months) | |||
| 0–12 | 3.66 ± 1.15 ** | 1034 ± 47 *** | 1389 ± 89 * |
| 12–24 | 4.15 ± 1.44 ** | 1188 ± 46 * | 1545 ± 71 |
| 24–48 | 2.10 ± 0.73 | 1185 ± 99 * | 1543 ± 92 |
| 48–144 | 2.58 ± 1.12 | 980 ± 113 *** | 1273 ± 109 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrillo, S.; Pietrafusa, N.; Trivisano, M.; Calabrese, C.; Saura, F.; Gallo, M.G.; Bertini, E.S.; Vigevano, F.; Specchio, N.; Piemonte, F. Imbalance of Systemic Redox Biomarkers in Children with Epilepsy: Role of Ferroptosis. Antioxidants 2021, 10, 1267. https://doi.org/10.3390/antiox10081267
Petrillo S, Pietrafusa N, Trivisano M, Calabrese C, Saura F, Gallo MG, Bertini ES, Vigevano F, Specchio N, Piemonte F. Imbalance of Systemic Redox Biomarkers in Children with Epilepsy: Role of Ferroptosis. Antioxidants. 2021; 10(8):1267. https://doi.org/10.3390/antiox10081267
Chicago/Turabian StylePetrillo, Sara, Nicola Pietrafusa, Marina Trivisano, Costanza Calabrese, Francesca Saura, Maria Giovanna Gallo, Enrico Silvio Bertini, Federico Vigevano, Nicola Specchio, and Fiorella Piemonte. 2021. "Imbalance of Systemic Redox Biomarkers in Children with Epilepsy: Role of Ferroptosis" Antioxidants 10, no. 8: 1267. https://doi.org/10.3390/antiox10081267
APA StylePetrillo, S., Pietrafusa, N., Trivisano, M., Calabrese, C., Saura, F., Gallo, M. G., Bertini, E. S., Vigevano, F., Specchio, N., & Piemonte, F. (2021). Imbalance of Systemic Redox Biomarkers in Children with Epilepsy: Role of Ferroptosis. Antioxidants, 10(8), 1267. https://doi.org/10.3390/antiox10081267