1. Introduction
Diabetes mellitus is one of the most serious and complex chronic illnesses causing deaths all over the world. It has been reported that the disorder affects about 4% of the world population and the prevalence is projected to surpass 5.4% by 2025 [
1]. Diabetes is caused by a chain of complex reactions characterized by hyperglycemia and alterations in the metabolism of carbohydrates, protein, or lipids [
2]. The chronic condition is associated with various acute metabolic side effects, such as ketoacidosis and hyperosmolar coma leading to chronic disorders, including retinopathy, renal failure, neuropathy, and other cardiovascular-related complications [
3]. Moreover, elevated blood sugar may trigger non-enzymatic interactions between glucose and proteins, leading to the formation of glycated products known as advanced glycation end products (AGE) [
4]. In diabetic patients, the accumulation of AGEs is accelerated, leading to further complications, including inflammation of tissues and formation of permanent crosslinks with body macromolecules [
5]. Diabetes can be classified into insulin-dependent (IDDM) and non-insulin-dependent diabetes mellitus (NIDDM), with the latter being the most prevalent among diabetic patients [
1]. The high prevalence rate of diabetes has led to various interventions, including developments of natural remedies and dietary management strategies steered toward finding safer alternatives to synthetic drugs in efforts to treat the disorder. From the perspective of dietary management, the development of antihyperglycemic foods that possess the potential to reduce the negative side effects of diabetes is a priority to researchers. Such foods can act as antioxidants, reducing inflammatory responses due to free radicles; as enzyme inhibitors blocking the hydrolyzing activities of α-glucosidase, amylase, and pancreatic lipase; and as AGE formation inhibitors preventing the glycation of fats or protein complexes in diabetic patients [
1,
4,
5].
Numerous epidemiological studies have demonstrated that the consumption of brassica vegetables such as broccoli, red cabbage, and radish have health-promoting effects in consumers [
6,
7,
8]. Growing evidence reveals that consuming young seedlings from these vegetables offers potential therapeutic benefits in diabetic management by eliminating predisposing factors [
8,
9]. For example, broccoli sprouts are known to rectify insulin resistance in type 2 diabetes, in addition to reducing side effects associated with the chronic condition [
8]. Broccoli sprouts are rich in flavonoids and other polyphenolic compounds that can exert both anti-inflammatory and antioxidant activities, thereby protecting against diabetes [
9]. Similarly, red cabbage offers therapeutic potential due to its high content of bioactive metabolites, including anthocyanins, flavonols, and glucosinolate [
7]. Thus, among the target food products, broccoli and red cabbage sprouts are becoming major candidates in the search for diabetes remedies. Germination is a simple but effective technique to produce edible seeds with improved biological functions. Gan et al. asserted that germination can lead to accumulation of various bioactive components of edible seeds including vitamins, polyphenolic, and non-polyphenolic compounds through de novo synthesis of secondary metabolites [
10]. It has been widely recognized that germinated seeds comprise a wide range of bioactive components that require an ongoing assessment to fully reveal their potential role in attenuating chronic illnesses.
Metabolomics is a new discipline in the area of “omics”. The technique allows simultaneous detection and characterization of metabolites synthesized in different metabolic processes. Untargeted metabolomics approaches screen a whole spectrum of metabolites in sample extracts and can be used to compare metabolite profiles of different samples. Owing to the diversity of plant metabolites and the underlying metabolic pathways involved in their synthesis, multiple metabolites can be produced during the germination process of edible seeds. Therefore, the first aim of this study was to investigate the alterations caused by germination on metabolite profiles of broccoli and red cabbage seeds using a UHPLC-ESI-QTOF-MS/MS-based untargeted metabolite profiling approach. The second objective was to assess the impact of germination on the biological activities of these seeds related to their antioxidant and antidiabetic potentials. To the best of our knowledge, this is the first time the untargeted metabolomics method has been used to compare the entire metabolite profile of broccoli and red cabbage seeds and sprouts. We also report for the first time the presence of diverse amino acids in broccoli and red cabbage seeds after germination. The study will contribute to the ongoing assessment of the potential therapeutic effects of germinated seeds in managing chronic conditions.
4. Discussion
Phenolic compounds are important secondary metabolites that demonstrate potential health benefits when consumed. These compounds may act as natural antioxidants attenuating the side effects of free radicles in chronic conditions in humans [
18]. It was, therefore, essential to determine their levels in this research. Past studies have reported on changes of the TPC content when seeds were germinated [
19,
20,
21]. Previous studies have indicated that germination can gradually lead to TPC accumulation in germinated edible seeds by stimulating enzymes such as phenylalanine ammonia lyase (PAL), which plays key roles in the de novo pathways [
10,
19,
22]. However, the current study showed contrasting results, revealing that germination either had no effect or significantly decreased TPC levels in sprouts. For instance, germination did not significantly affect TPC levels in red cabbage seeds but significantly reduced their levels in broccoli seeds from 480.4 ± 19.13 to 385.4 ± 13.66 mg FAE/100 g DW. The variation between these findings and those reported in previous studies may be due to factors such as the type of seed variety and germination conditions used [
23]. Nevertheless, our results were consistent with those reported for red cabbage [
23] and mung bean [
24,
25], indicating that germination can actually lead to either increased or decreased levels or may not significantly affect TPC levels in the germinated seeds.
Flavonoids are the major types of phenolic compounds, which are useful in protecting against diabetes in humans, in addition to playing various key roles, such as protecting the plant against environmental stress [
22]. Earlier, it was reported that germination improved TFC levels in broccoli [
20] and red cabbage [
21]. In contrast to these reports, our study revealed that germination significantly decreased the levels of TFC in red cabbage from 215.7 ± 0.16 to 209.9 ± 0.03 mg CE/100 g. Similarly, TFC levels in broccoli seeds significantly decreased upon germination from 216.9 ± 0.38 mg CE/100 g in raw seeds to 206.9 ± 0.02 mg CE/100 g in sprouts. There were no significant differences in the levels of TFC in broccoli seed (216.9 ± 0.38 mg CE/100 g) and red cabbage seed (215.6 ± 10.2 mg CE/100 g) extracts. Likewise, no significant difference was observed in the TFC levels of broccoli sprout extract (206.9 ± 10.2 mg CE/100 g) and red cabbage sprout extract (209.9 ± 10.2 mg CE/100 g). Factors that may lead to decreased TPC and TFC levels in seeds upon germination may include leaching of soluble phenolic compounds into the soaking water and the breakdown of bound phenolic compounds to form other compounds during germination.
Saponins are naturally occurring surface-active glycosides in plants. Saponin have been shown to exert potent glycemic control and prevent disorders associated with hyperglycemia. According to Elekofehinti, the antidiabetic activity of saponin is mainly due to its ability to induce insulin production from the pancreas [
26]. Saponins could also be involved in various therapeutic pharmacological activities in diabetic conditions, including inhibition of AGE formation and oxidative stress, as well as transformation of growth factor β1 (TGFβ1), thereby preventing the development of diabetic nephropathy [
26]. Our study revealed the presence of saponin in both broccoli and red cabbage. However, while germination did not significantly impact their levels in red cabbage (
p < 0.05), the process sharply enhanced the TSC levels in broccoli sprouts, as described in
Table 1. The literature also suggests that there is a notable impact on saponin content upon germination of seeds [
27,
28]. The decreases in the saponin levels observed for red cabbage sprouts in this work is in agreement with reports that germination process reduces these compounds in seeds [
28]. Additionally, the observation for broccoli sprouts that germination actually improved saponin levels in seeds were in agreement with the results reported by Ayet, who noted lower levels of saponin and phytic acid during germination of seeds [
27]. It is known that germination reduces antinutritive factors in plants, however this has little effect on the compounds involved in the plants’ defense mechanisms [
27]. Therefore, an increase or decrease in saponin content in seeds during germination could be explained by the implication of these compounds in the plant defense system.
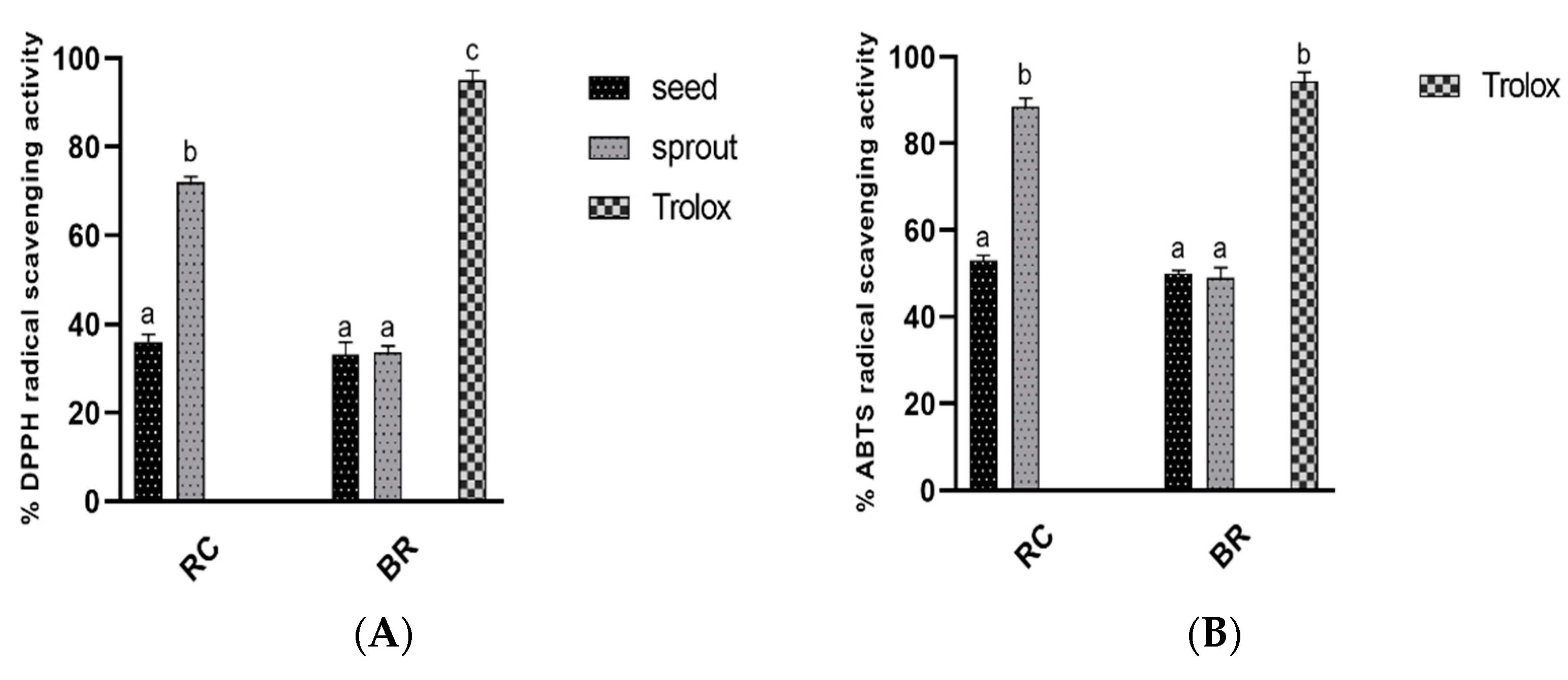
Over decades, studies have supported the role of oxygen free radicals as mediators in the development of diabetic complications [
29]. Developing a natural antioxidant that can inhibit the effects of the free radicle is considered a milestone in diabetic management [
29]. The antioxidant activity levels of raw seeds and sprouts were analyzed in this study. Even though germination did not significantly affect the antioxidant ability of broccoli seeds, it led to a sharp increase in the antioxidant capacity of red cabbage (shown in
Figure 2A,B and
Table 2). The current findings were consistent with those reported by Vale et al., who described increased antioxidant activities for red cabbage samples germinated under light [
30]. Therefore, germination can act as a simple and effective method to improve the antioxidant activities of edible seeds.
The hydrolyzing enzymes such as α-glucosidase and pancreatic lipase are important in diabetes control. While α-glucosidase catalyzes the cleavage of the absorbable sugars from disaccharides and oligosaccharides, increasing the blood glucose levels, pancreatic lipase hydrolyzes lipids, leading to fat accumulation in the pancreas [
31,
32,
33]. Hence, the primary target to control postprandial blood glucose levels and lipid accumulation is to develop compounds that can competitively inhibit digestive enzymes, thereby reducing the hydrolysis of carbohydrates and lipids in the body. In the past, commercially prepared α-glucosidase and lipase inhibitors such as acarbose and orlistat have been used to control the activity of these enzymes in the body. Nevertheless, these compounds are associated with gastrointestinal side effects, limiting their use [
34]. The production of natural inhibitors of α-glucosidase and pancreatic lipase from plant-based sources may provide an alternative means to treat diabetes or manage it without involving synthetic inhibitors. Within the current collection of foods, germinated seeds have demonstrated an effective source for an attractive strategy to control postprandial hyperglycemia and excessive accumulation of lipids in the body [
31]. The inhibition of α-glucosidase, pancreatic lipase activities, and AGE formation by ethanol extracts of broccoli and red cabbage were investigated in this study (
Figure 3A–C). Germinating red cabbage and broccoli seeds led to increased α-glucosidase inhibition by 33% and 16%, respectively. Similar observations were made for pancreatic lipase inhibition activities, for which increases of about 7% and 10% were observed for red cabbage and broccoli, respectively, upon germination.
Simple sugars such as glucose can non-enzymatically undergo a reactive process with the amino groups of proteins, nucleic acids, and fats, leading to the formation of glycated senescent macromolecules known as AGEs. The formation of AGEs progresses faster under inflammatory and hyperglycemic conditions, leading to loss of the integrity of the macromolecules [
5]. Earlier reports indicated that plant extracts were able to block the expression of RAGE pathways, thereby reducing the effects of AGEs in vivo [
35]. Germination significantly improved the AGE activities of broccoli sprouts but had little impact on the activities of red cabbage (
Figure 3C). Maeda et al. also reported similar results for the broccoli sprouts with higher sulforaphane contents [
36]. The phytochemicals, including alkaloids, flavonoids, saponins, and steroids, are responsible for the antidiabetic activities in brassica vegetables [
37].
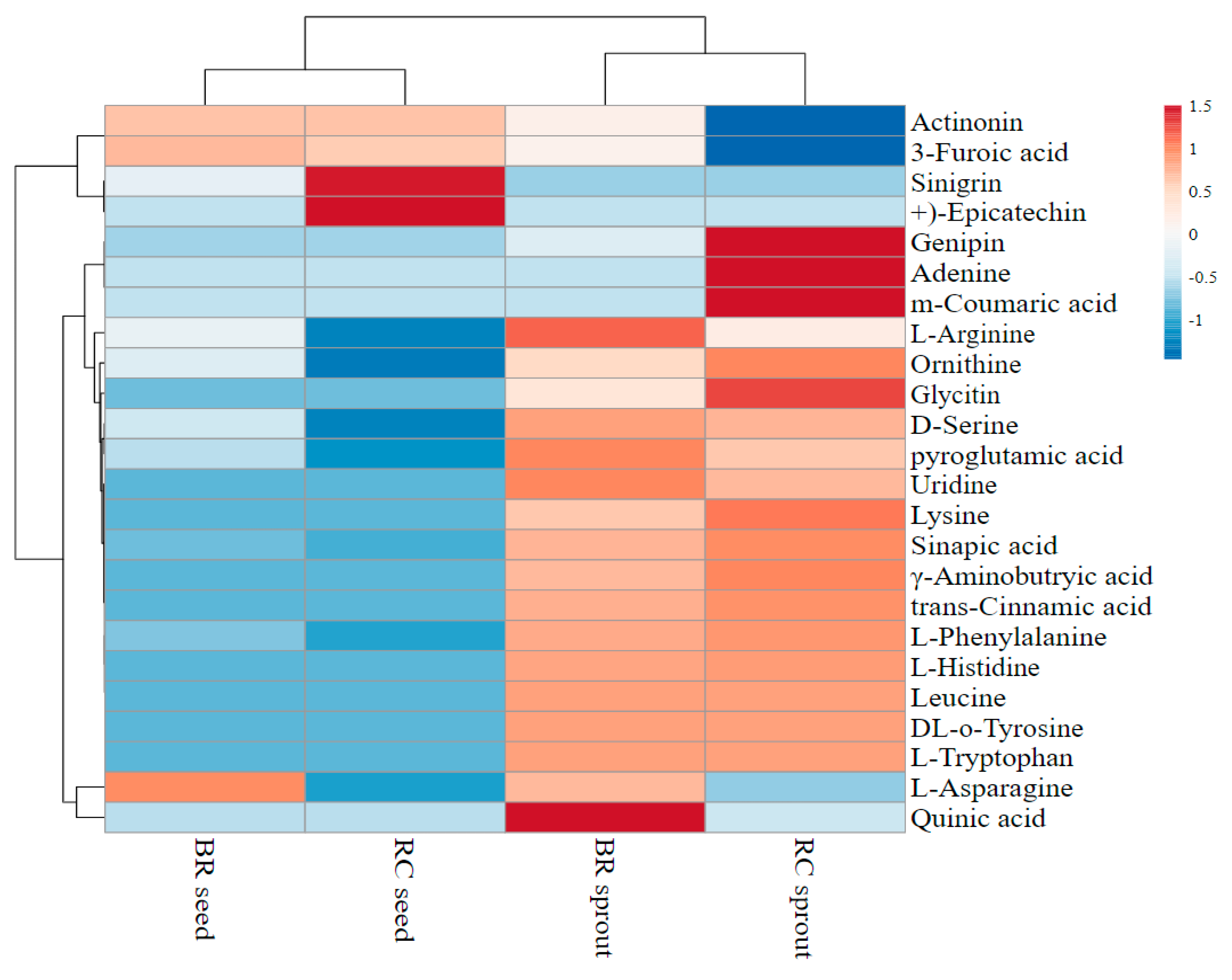
The rich bio-composition of key secondary metabolites within the Brassicaceae vegetables covers many compounds that can be extracted in alcohol solutions. Most of these metabolites are recognized as phytochemicals, which are known for their roles in the prevention of chronic diseases, as well as other health-promoting benefits. The current study used UHPLC-Q-TOF-MS2 to analyze the metabolites in the broccoli and red cabbage extracts. In the sprout–raw seed comparison, 16 metabolites were upregulated and 4 were downregulated in broccoli seeds after germination (
Figure 4). Similarly, the contents of 17 metabolites were enhanced while the levels of 4 metabolites were reduced by the germination process in red cabbage seeds. Correspondingly, the UHPLC-Q-TOF-MS2 analysis showed that the change trends for metabolites in seed triggered by the germination process varied. For example, germination caused 0.75-, 0.64-, 1.52-, and 5.53-fold increases in the levels of ornithine,
l-arginine,
d-serine, and
l-phenylalanine, respectively, in broccoli sprouts; while
l-asparagine was decreased by 5.53-fold. Similarly,
l-arginine and
l-asparagine were increased by 1.50- and 0.24-fold in red cabbage sprouts, respectively. Moreover, some of the metabolites that were previously not present in seeds were synthesized in the germination process.
Amino acids, other protein-derived compounds, and phenolic compounds were the prominent metabolites observed in this study. Nonetheless, amino acids were undisputedly the most detected and identified compounds in the extracts. By comparing the spectral data with those available in standard databases and other literature reports, the metabolites were tentatively identified (
Table 3,
Table 4,
Table 5 and
Table 6). For instance, amino acids with deprotonated [M−H]
- molecules at
m/
z values of 145.098, 154.062, 104.035, 131.046, 180.067, 130.087, 164.072, 203.083, and 445.115 were tentatively identified as lysine,
l-histidine,
d-serine,
l-asparagine,
dl-o-tyrosine, leucine,
l-phenylalanine,
l-tryptophan, and glycitin, respectively, by comparing their masses (
m/
z) with those in the mass spectral libraries XCMS Online (Metlin) and Metabolomics Workbench and further crosschecked with the
m/
z data reported by Hanhineva et al. [
38]. Fourteen (14) and fifteen (15) amino acids were tentatively identified in broccoli and red cabbage sprout extracts, respectively, (
Table 3;
Table 5).
l-Arginine, pyroglutamic acid, and
l-asparagine were the only amino acids present in the red cabbage seed extract (
Table 6), while ornithine,
l-asparagine, pyroglutamic acid, lysine,
d-serine, and
l-phenylalanine were the prominent amino acids in broccoli seed extract (
Table 4). Amino acids are important in plants as well as in humans. Some of these amino acids have already been shown to have positive physiological importance in the body, such as participating in the synthesis of macromolecules. However, plasma-free amino acid profiles have been associated with increased risks of diabetes type 2 [
39]. Therefore, amino acids have the potential to be used as biomarkers for assessing diabetes risk, as well as in monitoring strategies designed to reduce such risks [
39]. In addition to enhancing their levels, germination processes also resulted in the synthesis of new amino acids, including
l-histidine,
l-tryptophan, leucine, and lysine, which were previously not present in raw seed samples. Consequently, germination was an essential process needed to improve the concentrations of various amino acids in edible seeds. Furthermore, γ-aminobutryic acid, the major non-protein amino acid identified in the germinated broccoli and red cabbage sprout extracts deprotonated at the [M−H]
-m/
z value of 102.056 was confirmed by the product ions of
m/z 102 (100%), 101 (5%), using the Metlin database (
https://isometlin.scripps.edu). GABA is a known neurotransmitter in the body, as well as an antioxidant, antidiabetic, and antiobesity compound, among other biological functions [
40]. A similar identification method was applied to tentatively identify other protein-derived compounds that were deprotonated at
m/
z 243.062 and 134.047, namely uridine and adenine, respectively. The uridine and adenine are nucleic acid components that were only found in the sprouted seed extracts.
Previous studies have shown that glucosinolates are prominent groups of compounds exerting beneficial health effects in brassica vegetables [
41]. Glucosinolates are key metabolites involved in controlling obesity, diabetes, and many other cardiovascular diseases [
38]. In the present study, sinigrin was identified as the only glucosinolate at
m/
z 358.027 present in broccoli and red cabbage seed extracts after comparison with MS spectra data from the literature [
42]. In terms of phenolic profile subclasses, flavanol and phenolic acids were found in the extracts. Compound C
15H
14O
6 exhibited a deprotonated [M−H]
- molecule at
m/
z 289.0721, with fragmented ions at
m/
z values of 245 and 151, with the most intense value of 289 being detected in raw broccoli seeds. The compound C
15H
14O
6 was tentatively identified as (+)-epicatechin via comparison with MS spectral data in the literature [
43]. (+)-Epicatechin and other catechin derivatives have been reported to exert powerful antioxidant and anti-inflammatory activities in diabetic conditions [
44].
Phenolic acids and other organic acids were widely present in broccoli and red cabbage extracts. Compounds with deprotonated [M−H]
- molecules at
m/
z 191.056, 223.061, 163.040, and 111.009 were identified as quinic acid, sinapic acid, m-coumaric acid, and 3-furoic acid, respectively, using the method described above and via crosschecking with figures reported in the literature by Hong et al. [
45]. Phenolic acids play key roles in various biological processes in humans, in addition to their roles in the synthesis of other phenolic compounds in plants. For instance, quinic acid derivatives are polyphenol-rich compounds consisting of a large group of esters synthesized from quinic acid and one or more phenylpropanoic acid, such as sinapic acid, coumaric acid, and cinnamic acid [
46]. Quinic acid and its derivatives have been reported to exert antidiabetic properties by alleviating structural degeneration in the pancreas, liver, and kidneys [
46]. Moreover, sinapic acid was also reported as a strong antioxidant compound in vitro [
47]. Interestingly, sinapic acid and quinic acid were found at lower levels in seed extracts compared to sprout extracts. The absence of certain antioxidant and antidiabetic phytochemicals or their lower concentrations in raw seed explains why germinated samples exhibited higher activities compared to raw samples. All other remaining metabolites were tentatively identified using the methods described above.
We reported for the first time the synthesis of diverse amino acids, including l-arginine, d-serine, and l-phenylalanine, as well as lysin, in broccoli and red cabbage seeds upon germination. These findings are unique in revealing the role of germination in enhancing not only phenolic compounds but also amino acid profiles in edible seeds. The current findings confirm the results of previous studies showing that germination can significantly alter the metabolite compositions of edible seeds, in addition to enhancing their functional properties.











{kind=link}
{kind=link}
{kind=link}
{kind=link}