
Isoprene: An Antioxidant Itself or a Molecule with Multiple Regulatory Functions in Plants?


Abstract
1. Isoprene: Some Generalities
2. Isoprene Protection from Abiotic Stress
3. Isoprene Modulation of Genomic, Proteomic and Metabolomic Profiles
4. Isoprene as Hormone Regulating Plant Antioxidant System and Growth
5. Isoprene as Defense Priming Molecule: Emerging Clues
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sharkey, T.D.; Yeh, S. Isoprene emission from plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 407–436. [Google Scholar] [CrossRef] [PubMed]
- Silver, G.M.; Fall, R. Enzymatic synthesis of isoprene from dimethylallyl diphosphate in aspen leaf extracts. Plant Physiol. 1991, 97, 1588–1591. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F.; Ciccioli, P.; Brancaleoni, E.; Cecinato, A.; Frattoni, M.; Sharkey, T.D. Different sources of reduced carbon contribute to form three classes of terpenoid emitted by Quercus ilex L leaves. Proc. Natl. Acad. Sci. USA 1996, 93, 9966–9969. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Schwender, J.; Disch, A.; Rohmer, M. Biosynthesis of isoprenoids in higher plant chloroplasts proceeds via a mevalonate-independent pathway. FEBS Lett. 1997, 400, 271–274. [Google Scholar] [CrossRef]
- Dani, K.G.S.; Fineschi, S.; Michelozzi, M.; Loreto, F. Do cytokinins, volatile isoprenoids and carotenoids synergically delay leaf senescence? Plant Cell Environ. 2016, 39, 1103–1111. [Google Scholar] [CrossRef]
- Monson, R.K.; Fall, R. Isoprene emission from aspen leaves: Influence of environment and relation to photosynthesis and photorespiration. Plant Physiol. 1989, 90, 267–274. [Google Scholar] [CrossRef]
- Sanadze, G.A. Biogenic Isoprene (A Review). Russ. J. Plant Physiol. 2004, 51, 729–741. [Google Scholar] [CrossRef]
- Dani, K.G.S.; Torzillo, G.; Michelozzi, M.; Baraldi, R.; Loreto, F. Isoprene Emission in Darkness by a Facultative Heterotrophic Green Alga. Front. Plant Sci. 2020, 11, 598786. [Google Scholar] [CrossRef]
- Wagner, W.P.; Helmig, D.; Fall, R. Isoprene biosynthesis in Bacillus subtilis via the methylerythritol phosphate pathway. J. Nat. Prod. 2000, 63, 37–40. [Google Scholar] [CrossRef]
- Sharkey, T.D. Isoprene synthesis by plants and animals. Endeavor 1996, 20, 74–78. [Google Scholar] [CrossRef]
- Vickers, C.E.; Possell, M.; Cojocariu, C.I.; Velikova, V.B.; Laothawornkitkul, J.; Ryan, A.; Mullineaux, P.M.; Nicholas Hewitt, C. Isoprene synthesis protects transgenic tobacco plants from oxidative stress. Plant Cell Environ. 2009, 32, 520–531. [Google Scholar] [CrossRef]
- Karl, T.; Harley, P.; Emmons, L.; Thornton, B.; Guenther, A.; Basu, C.; Turnipseed, A.; Jardine, K. Efficient atmospheric cleansing of oxidized organic trace gases by vegetation. Science 2010, 330, 816–819. [Google Scholar] [CrossRef]
- Muramoto, S.; Matsubara, Y.; Mwenda, C.M.; Koeduka, T.; Sakami, T.; Tani, A.; Matsui, K. Glutathionylation and Reduction of Methacrolein in Tomato Plants Account for Its Absorption from the Vapor Phase. Plant Physiol. 2015, 169, 1744–1754. [Google Scholar] [CrossRef]
- Cappellin, L.; Loreto, F.; Biasioli, F.; Pastore, P.; McKinney, K. A mechanism for biogenic production and emission of MEK from MVK decoupled from isoprene biosynthesis. Atmos. Chem. Phys. 2019, 19, 3125–3135. [Google Scholar] [CrossRef]
- Singsaas, E.L.; Lerdau, M.; Winter, K.; Sharkey, T.D. Isoprene increases thermotolerance of isoprene-emitting species. Plant Physiol. 1997, 115, 1413–1420. [Google Scholar] [CrossRef]
- Loreto, F.; Ciccioli, P.; Brancaleoni, E.; Cecinato, A.; Frattoni, M. Simultaneous measurements of emission and content of terpenes in leaves of oak species. Plant Physiol. 1997, 114, 1100. [Google Scholar]
- Harvey, C.M.; Li, Z.; Tjellström, H.; Blanchard, G.J.; Sharkey, T.D. Concentration of isoprene in artificial and thylakoid membranes. J. Bioenerg. Biomembr. 2015, 47, 419–429. [Google Scholar] [CrossRef]
- Lantz, A.T.; Allman, J.; Weraduwage, S.M.; Sharkey, T.D. Isoprene: New insights into the control of emission and mediation of stress tolerance by gene expression. Plant Cell Environ. 2019, 42, 2808–2826. [Google Scholar] [CrossRef]
- Keenan, T.F.; Grote, R.; Sabaté, S. Overlooking the canopy: The importance of canopy structure in scaling isoprenoid emissions from the leaf to the landscape. Ecol. Model. 2011, 222, 737–747. [Google Scholar] [CrossRef]
- Guenther, A.; Karl, T.; Harley, P.; Wiedinmyer, C.; Palmer, P.I.; Geron, C. Atmospheric Chemistry and Physics Estimates of global terrestrial isoprene emissions using MEGAN (Model of Emissions of Gases and Aerosols from Nature). Atmos. Chem. Phys. 2006, 6, 3181–3210. [Google Scholar] [CrossRef]
- Sindelarova, K.; Granier, C.; Bouarar, I.; Guenther, A.; Tilmes, S.; Stavrakou, T.; Müller, J.-F.; Kuhn, U.; Stefani, P.; Knorr, W. Global data set of biogenic VOC emissions calculated by the MEGAN model over the last 30 years. Atmos. Chem. Phys. 2014, 14, 9317–9341. [Google Scholar] [CrossRef]
- Loreto, F.; Fineschi, S. Reconciling functions and evolution of isoprene emission in higher plants. New Phytol. 2015, 206, 578–582. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Gray, D.W.; Pell, H.K.; Breneman, S.R.; Topper, L. Isoprene synthase genes form a monophyletic clade of acyclic terpene synthases in the Tps-b terpene synthase family. Evolution 2013, 67, 1026–1040. [Google Scholar] [CrossRef]
- Li, M.; Xu, J.; Algarra Alarcon, A.; Carlin, S.; Barbaro, E.; Cappellin, L.; Velikova, V.; Vrhovsek, U.; Loreto, F.; Varotto, C. In Planta Recapitulation of Isoprene Synthase Evolution from Ocimene Synthases. Mol. Biol. Evol. 2017, 34, 2583–2599. [Google Scholar] [CrossRef]
- Sanadze, G.A. Emission of Gaseous Organic Substances from Plants. Rep. Akad. Nauk GruzSSR 1956, 17, 429–433. [Google Scholar]
- Sanadze, G.A.; Kursanov, A.L. On certain conditions of the evolution of the diene C5H8 from poplar leaves. Sov. Plant Physiol. 1996, 13, 184–189. [Google Scholar]
- Paré, P.W.; Tumlinson, J.H. Plant volatiles as a defense against insect herbivores. Plant Physiol. 1999, 121, 325–332. [Google Scholar] [CrossRef]
- Dicke, M.; Baldwin, I.T. The evolutionary context for herbivore induced plant volatiles: Beyond the ‘cry for help’. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef]
- Loreto, F.; Dicke, M.; Schnitzler, J.-P.; Turlings, T.C.J. Plant volatiles and the environment. Plant Cell Environ. 2014, 37, 1905–1908. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Chen, X.; Yeh, S. Isoprene increases thermotolerance of fosmidomycin-fed leaves. Plant Physiol. 2001, 125, 2001–2006. [Google Scholar] [CrossRef]
- Loivamäki, M.; Gilmer, F.; Fischbach, R.J.; Sörgel, C.; Bachl, A.; Walter, A.; Schnitzler, J.P. Arabidopsis, a model to study biological functions of isoprene emission? Plant Physiol. 2007, 144, 1066–1078. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Saito, T.; Lamsa, M.; Oksman-Caldentey, K.M.; Suzuki, M.; Ohyama, K.; Yazaki, K. Plants utilize isoprene emission as a thermotolerance mechanism. Plant Cell Physiol. 2007, 48, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Velikova, V.; Loreto, F. On the relationship between isoprene emission and thermotolerance in Phragmites australis leaves exposed to high temperatures and during the recovery from a heat stress. Plant Cell Environ. 2005, 28, 318–327. [Google Scholar] [CrossRef]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus × acerifolia plants during Mediterranean summers. New Phytol. 2015, 207, 613–626. [Google Scholar] [CrossRef]
- Loreto, F.; Mannozzi, M.; Maris, C.; Nascetti, P.; Ferranti, F.; Pasqualini, S. Ozone quenching properties of isoprene and its antioxidant role in plants. Plant Physiol. 2001, 126, 1–8. [Google Scholar] [CrossRef]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Velikova, V.; Edreva, A.; Loreto, F. Endogenous isoprene protects Phragmites australis leaves against singlet oxygen. Physiol. Plant. 2004, 122, 219–225. [Google Scholar] [CrossRef]
- Affek, H.; Yakir, D. Protection by Isoprene against Singlet Oxygen in Leaves. Plant Physiol. 2002, 129, 269–277. [Google Scholar] [CrossRef]
- Velikova, V.; Pinelli, P.; Pasqualini, S.; Reale, L.; Ferranti, F.; Loreto, F. Isoprene decreases the concentration of nitric oxide in leaves exposed to elevated ozone. New Phytol. 2005, 166, 419–426. [Google Scholar] [CrossRef]
- Jardine, K.; Chambers, J.; Alves, E.G.; Teixeira, A.; Garcia, S.; Holm, J.; Higuchi, N.; Manzi, A.; Abrell, L.; Fuentes, J.D.; et al. Dynamic Balancing of Isoprene Carbon Sources Reflects Photosynthetic and Photorespiratory Responses to Temperature Stress. Plant Physiol. 2014, 166, 2051–2064. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Singsaas, E.L. Why plants emit isoprene. Nature 1995, 374, 769. [Google Scholar] [CrossRef]
- Velikova, V.; Varkonyi, Z.; Szabo, M.; Maslenkova, L.; Nogues, I.; Kovacs, L.; Peeva, V.; Busheva, M.; Garab, G.; Sharkey, T.D.; et al. Increased thermostability of thylakoid membranes in isoprene-emitting leaves probed with three biophysical techniques. Plant Physiol. 2011, 157, 905–916. [Google Scholar] [CrossRef]
- Jardine, K.J.; Monson, R.K.; Abrell, L.; Saleska, S.R.; Arneth, A.; Jardine, A.; Ishida, F.Y.; Serrano, A.M.Y.; Artaxo, P.; Karl, T.; et al. Within-plant isoprene oxidation confirmed by direct emissions of oxidation products methyl vinyl ketone and methacrolein. Glob. Chang. Biol. 2012, 18, 973–984. [Google Scholar] [CrossRef]
- Loreto, F.; Fares, S. Is ozone flux inside leaves only a damage indicator? Clues from volatile isoprenoid studies. Plant Physiol. 2007, 143, 1096–1100. [Google Scholar] [CrossRef]
- Pollastri, S.; Tsonev, T.; Loreto, F. Isoprene improves photochemical efficiency and enhances heat dissipation in plants at 30 physiological temperatures. J. Exp. Bot. 2014, 65, 1565–1570. [Google Scholar] [CrossRef]
- Loreto, F.; Schnitzler, J.-P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Pollastri, S.; Jorba, I.; Hawkins, T.J.; Llusià, J.; Michelozzi, M.; Navajas, D.; Peñuelas, J.; Hussey, P.J.; Knight, M.R.; Loreto, F. Leaves of isoprene-emitting tobacco plants maintain PSII stability at high temperatures. New Phytol. 2019, 223, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Siwko, M.E.; Marrink, S.J.; de Vries, A.H.; Kozubek, A.; Schoot Uiterkamp, A.J.M.; Mark, A.E. Does isoprene protect plant membranes from thermal shock? A molecular dynamics study. Biochim. Biophys. Acta 2007, 1768, 198–206. [Google Scholar] [CrossRef]
- Kirchoff, H.; Hall, C.; Wood, M.; Herbstová, M.; Tsabari, O.; Nevo, R.; Charuvi, D.; Shimoni, E.; Reich, Z. Dynamic control of protein diffusion within the granal thylakoid lumen. Proc. Natl. Acad. Sci. USA 2011, 108, 20248–20253. [Google Scholar] [CrossRef]
- Behnke, K.; Loivamäki, M.; Zimmer, I.; Rennenberg, H.; Schnitzler, J.-P.; Louis, S. Isoprene emission protects photosynthesis in sunfleck exposed Grey poplar. Photosynth. Res. 2010, 104, 5–17. [Google Scholar] [CrossRef]
- Zuo, Z.; Weraduwage, S.M.; Lantz, A.T.; Sanchez, L.M.; Weise, S.E.; Wang, J.; Childs, K.L.; Sharkey, T.D. Isoprene Acts as a Signaling Molecule in Gene Networks Important for Stress Responses and Plant Growth. Plant Physiol. 2019, 180, 124–152. [Google Scholar] [CrossRef] [PubMed]
- Harvey, C.M.; Sharkey, T.D. Exogenous isoprene modulates gene expression in unstressed Arabidopsis thaliana plants. Plant Cell Environ. 2016, 39, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Trainotti, L.; Li, M.; Varotto, C. Overexpression of Isoprene Synthase Affects ABA- and Drought-Related Gene Expression and Enhances Tolerance to Abiotic Stress. Int. J. Mol. Sci. 2020, 21, 4276. [Google Scholar] [CrossRef] [PubMed]
- Monson, R.K.; Weraduwage, S.M.; Rosenkranz, M.; Schnitzler, J.-P.; Sharkey, T.D. Leaf isoprene emission as a trait that mediates the growth-defense tradeoff in the face of climate stress. Oecologia 2021, 1–18. [Google Scholar] [CrossRef]
- Velikova, V.; Ghirardo, A.; Vanzo, E.; Merl, J.; Hauck, S.M.; Schnitzler, J.P. Genetic manipulation of isoprene emissions in poplar plants remodels the chloroplast proteome. J. Proteome Res. 2014, 13, 2005–2018. [Google Scholar] [CrossRef]
- Monson, R.K.; Winkler, B.; Rosenstiel, T.N.; Block, K.; Merl-Pham, J.; Strauss, S.H.; Ault, K.; Maxfield, J.; Moore, D.J.P.; Trahan, N.A.; et al. High productivity in hybrid-poplar plantations without isoprene emission to the atmosphere. Proc. Natl. Acad. Sci. USA 2020, 117, 1596–1605. [Google Scholar] [CrossRef]
- Ghirardo, A.; Wright, L.P.; Bi, Z.; Rosenkranz, M.; Pulido, P.; Rodríguez-Concepción, M.; Niinemets, Ü.; Brüggemann, N.; Gershenzon, J.; Schnitzler, J.-P. Metabolic flux analysis of plastidic isoprenoid biosynthesis in poplar leaves emitting and nonemitting isoprene. Plant Physiol. 2014, 165, 37–51. [Google Scholar] [CrossRef]
- Behnke, K.; Kaiser, A.; Zimmer, I.; Brüggemann, N.; Janz, D.; Polle, A.; Hampp, R.; Hänsch, R.; Popko, J.; Schmitt-Kopplin, P.; et al. RNAi-mediated suppression of isoprene emission in poplar transiently impacts phenolic metabolism under high temperature and high light intensities: A transcriptomic and metabolomic analysis. Plant Mol. Biol. 2010, 74, 61–75. [Google Scholar] [CrossRef]
- Behnke, K.; Ehlting, B.; Teuber, M.; Bauerfeind, M.; Louis, S.; Hänsch, R.; Polle, A.; Bohlmann, J.; Schnitzler, J.-P. Transgenic, non-isoprene emitting poplars don’t like it hot. Plant J. 2007, 51, 485–499. [Google Scholar] [CrossRef]
- Tattini, M.; Velikova, V.; Vickers, C.; Brunetti, C.; Di Ferdinando, M.; Trivellini, A.; Fineschi, S.; Agati, G.; Ferrini, F.; Loreto, F. Isoprene production in transgenic tobacco alters isoprenoid, non-structural carbohydrate and phenylpropanoid metabolism, and protects photosynthesis from drought stress. Plant Cell Environ. 2014, 37, 1950–1964. [Google Scholar] [CrossRef]
- Velikova, V.; Muller, C.; Ghirardo, A.; Rock, T.M.; Aichler, M.; Walch, A.; Schmitt-Kopplin, P.; Schnitzler, J.-P. Knocking down of isoprene emission modifies the lipid matrix of thylakoid membranes and influences the chloroplast ultrastructure in poplar. Plant Physiol. 2015, 168, 859–870. [Google Scholar] [CrossRef]
- Tarkowská, D.; Strnad, M. Isoprenoid-derived plant signaling molecules: Biosynthesis and biological importance. Planta 2018, 247, 1051–1066. [Google Scholar] [CrossRef]
- Riedlmeier, M.; Ghirardo, A.; Wenig, M.; Knappe, C.; Koch, K.; Georgii, E.; Dey, S.; Parker, J.E.; Schnitzler, J.P.; Vlot, A.C. Monoterpenes Support Systemic Acquired Resistance within and between Plants. Plant Cell 2017, 29, 1440–1459. [Google Scholar] [CrossRef]
- Wenig, M.; Ghirardo, A.; Sales, J.H.; Pabst, E.S.; Breitenbach, H.H.; Antritter, F.; Weber, B.; Lange, B.; Lenk, M.; Cameron, R.K.; et al. Systemic acquired resistance networks amplify airborne defense cues. Nat. Commun. 2019, 10, 3813. [Google Scholar] [CrossRef]
- Miloradovic van Doorn, M.; Merl-Pham, J.; Ghirardo, A.; Fink, S.; Polle, A.; Schnitzler, J.P.; Rosenkranz, M. Root isoprene formation alters lateral root development. Plant Cell Environ. 2020, 43, 2207–2223. [Google Scholar] [CrossRef]
- Vanzo, E.; Merl-Pham, J.; Velikova, V.; Ghirardo, A.; Lindermayr, C.; Hauck, S.M.; Bernhardt, J.; Riedel, K.; Durner, J.; Schnitzler, J.-P. Modulation of Protein S-Nitrosylation by Isoprene Emission in Poplar. Plant Physiol. 2016, 170, 1945–1961. [Google Scholar] [CrossRef]
- Velikova, V.; Fares, S.; Loreto, F. Isoprene and nitric oxide reduce damages in leaves exposed to oxidative stress. Plant Cell Environ. 2008, 31, 1882–1894. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense Priming: An Adaptive Part of Induced Resistance. Ann. Rev. Plant Biol. 2017, 28, 485–512. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.J.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing Plant Defense Priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef]
- Bertini, L.; Palazzi, L.; Proietti, S.; Pollastri, S.; Arrigoni, G.; de Laureto, P.P.; Caruso, C. Proteomic Analysis of MeJa-Induced Defense Responses in Rice against Wounding. Int. J. Mol. Sci. 2019, 20, 2525. [Google Scholar] [CrossRef]
- Baccelli, I.; Benny, J.; Caruso, T.; Martinelli, F. The priming fingerprint on the plant transcriptome investigated through meta-analysis of RNA-Seq data. Eur. J. Plant Pathol. 2020, 156, 779–797. [Google Scholar] [CrossRef]
- Baccelli, I.; Mauch-Mani, B. Beta-aminobutyric acid priming of plant defense: The role of ABA and other hormones. Plant Mol. Biol. 2016, 91, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Barta, C.; Loreto, F. The relationship between the methyl-erythritol phosphate pathway leading to emission of volatile isoprenoids and abscisic acid content in leaves. Plant Physiol. 2006, 141, 1676–1683. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Frank, L.; Wenig, M.; Ghirardo, A.; van der Krol, A.; Vlot, A.C.; Schnitzler, J.P.; Rosenkranz, M. Isoprene and β-caryophyllene confer plant resistance via different plant internal signaling pathways. Plant Cell Environ. 2021. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F.; Barta, C.; Brilli, F.; Nogues, I. On the induction of volatile organic compound emissions by plants as consequence of wounding or fluctuations of light and temperature. Plant Cell Environ. 2006, 29, 1820–1828. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M.; Loreto, F. Induced plant volatiles: From genes to climate change. Trends Plant Sci. 2010, 15, 115–117. [Google Scholar] [CrossRef]
- Laothawornkitkul, J.; Taylor, J.E.; Paul, N.D.; Hewitt, C.N. Biogenic volatile organic compounds in the Earth system. New Phytol. 2009, 183, 27–51. [Google Scholar] [CrossRef]
- Brilli, F.; Barta, C.; Fortunati, A.; Lerdau, M.; Loreto, F.; Centritto, M. Response of isoprene emission and carbon metabolism to drought in white poplar (Populus alba) saplings. New Phytol. 2007, 175, 244–254. [Google Scholar] [CrossRef]
- Muller, A.; Kaling, M.; Faubert, P.; Gort, G.; Smid, H.M.; Van Loon, J.J.; Dicke, M.; Kanawati, B.; Schmitt-Kopplin, P.; Polle, A.; et al. Isoprene emission by poplar is not important for the feeding behaviour of poplar leaf beetles. BMC Plant Biol. 2015, 15, 165. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Isoprene Effect | Plant | Genes, Proteins, Metabolites Influenced by Isoprene | Condition | Reference |
|---|---|---|---|---|
| Genes | ||||
| Up-regulated | Poplar (WT and Knock-out) | Biosynthetic pathways of condensed tannins, anthocyanins and phenylpropanoids, and regulatory pathways of phenylpropanoids genes. | high light and heat stress | [50] |
| Up- and down-regulated | Arabidopsis and tobacco (WT and IspS). Arabidopsis (WT) fumigated with isoprene | Abiotic and biotic stress defence, growth regulator signaling pathways, photosynthesis, seed germination, and seedling and plant growth, JA mediated defense signaling, phenylpropanoid biosynthesis and regulation, and cytokinins (CK)-mediated wound repair genes. | non-stressful conditions | [51] |
| Up-regulated | Arabidopsis (WT) fumigated with isoprene | Chloroplast, phenylpropanoid biosynthetic and translation machinery genes, transcription factor-enriched gene network | non-stressful conditions | [52] |
| Up-regulated | Arabidopsis (WT and IspS) | RD29B gene (abscisic acid-activated signaling pathway) | exogenous ABA supplement | [53] |
| Down-regulated | Arabidopsis (WT and IspS) | COR15A gene (stromal proteins protection), and P5CS (reactive oxidative species protection) | water-stress | [53] |
| Proteins | ||||
| Higher concentrations | Poplar (WT and Knockout) | Chloroplast proteins involved in photosynthesis, light reactions, redox regulation, and oxidative stress defense and general metabolism | Control | [55] |
| Higher concentrations | Poplar (WT and Knockout) | Proteins involved in phenylpropanoid pathway and flavonoid biosynthesis | Control | [56] |
| Lower concentrations | Poplar (WT and Knockout) | Proteins involved in the biosynthesis of terpenoids, carotenoids and in the methylerythritol phosphate (MEP) and jasmonic acid pathways | Control | [56] |
| Metabolites | ||||
| Higher concentrations | Poplar (WT and Knockout) | Total phenolics and condensed tannins | high light and heat stress | [50] |
| Similar concentrations | Poplar (WT and Knockout) | Photosynthetic pigments and secondary metabolites | high light and heat stress | [50] |
| Higher concentrations | Arabidopsis and tobacco (WT and IspS) | Chlorophylls and carotenoids | non-stressful conditions | [51] |
| Higher concentrations | Poplar (WT and Knockout) | Chlorophylls, carotenes and xanthophylls | light and dark conditions | [57] |
| Higher concentrations | Poplar (WT and Knockout) | Chlorophyll content | Control | [50,58] |
| Similar concentrations | Poplar (WT and Knockout) | Carotenoids | Control | [50,58] |
| Lower concentrations | Poplar (WT and Knockout) | Zeaxanthin | Heat | [59] |
| Higher concentrations | Tobacco (WT and IspS) | Zeaxanthin, anteraxanthin and phenylpropanoids | drought conditions | [60] |
| Higher concentrations | Poplar (WT and Knockout) | Monogalactosyldiacylglycerols, digalactosyldiacylglycerols, phospholipids and unsaturated fatty acids | Control | [61] |
| Isoprene Role | Functional Expectations | Experimental Evidence |
|---|---|---|
| Antioxidant | Protection against oxidative damage by direct reaction with oxidizing molecules (ROS or NO) | PRO: Isoprene contributes to thermal and oxidative stress tolerance by quenching ROS and reactive nitrogen species [36,39]. CON: Secondary antioxidants generated by reaction with ROS (e.g., methyl vinyl ketone) being per se cytotoxic may be removed without isoprene quenching [14]. Insufficient isopreneconcentration inside membranes to carry out effective antioxidant action [17]. |
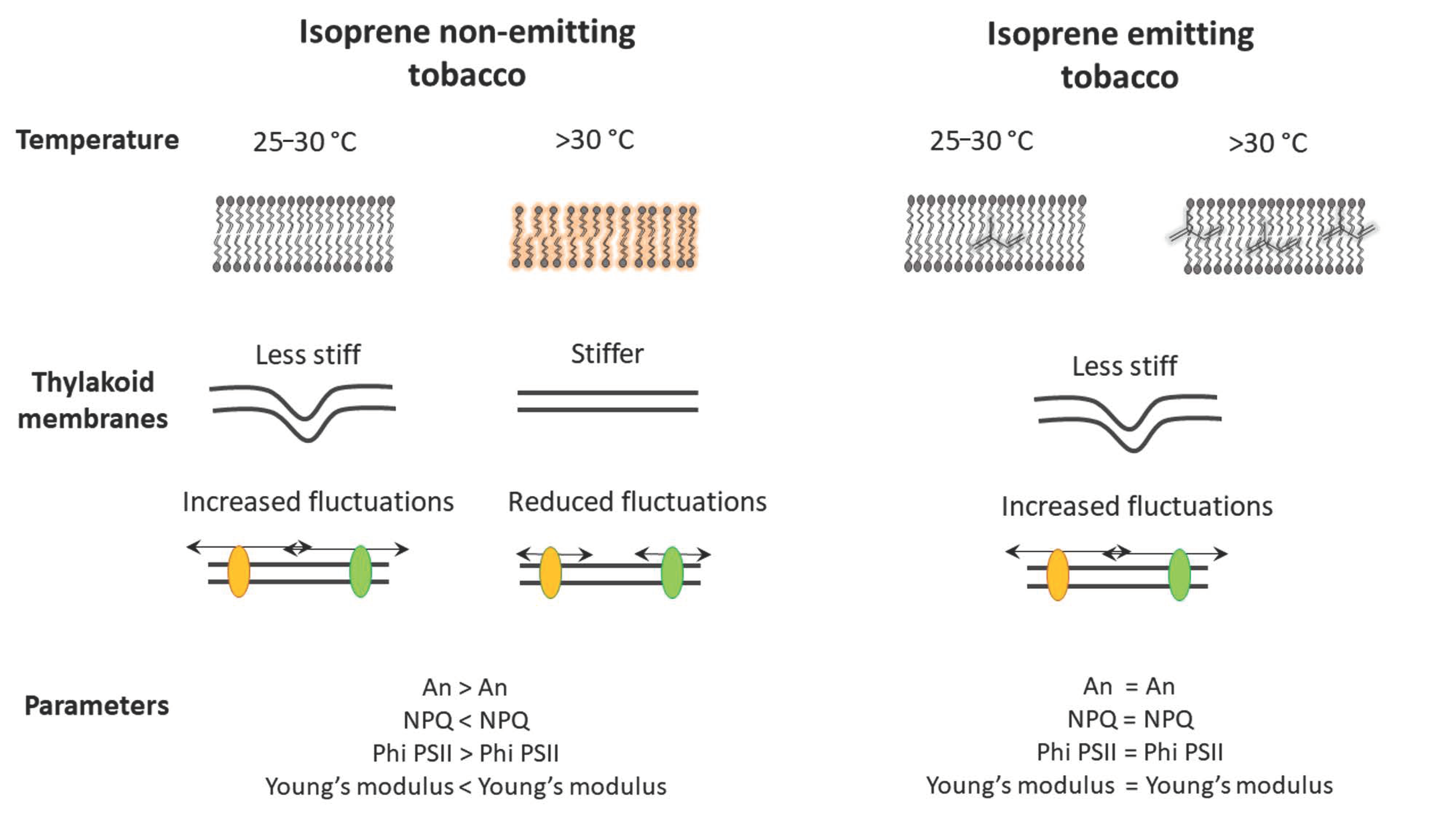
| Membrane protection | Stabilization of membrane properties when challenged by stresses by intercalating membrane lipids | PRO: Isoprene lipophylic nature favors isoprene presence in thylakoidal membranes which are kept elastic and fluid at rising temperature [29]. CON: Isoprene concentration in the membranes theoretically very low [17]. |
| Defense priming stimulus | Improving stress resistance/tolerance by boosting stress signal perception, propagation, and/or activation of defense mechanisms. A quicker/stronger defense gene expression during and not before stress conditions is the hallmark of priming. | PRO: Isoprene-emitting transgenic lines show enhanced tolerance to dehydration and heat stress by displaying enhanced ABA-induced stomatal closure, reduced water loss, and higher ABA-induced expression of the ABA-responsive marker gene RD29B [53]. Isoprene exposure induces resistance to bacterial infection through SA-dependent defense mechanisms in Arabidopsis [74]. Isoprene fumigation induces a small reprogramming of the transcriptome (167 DEGs, fold change >2) in Arabidopsis, including up-regulation of some of the most important defensive pathway genes [52]. CON: ROS levels are reduced during thermal and oxidative stress because of direct and rapid antioxidant action of isoprene [36], without any need to invoke priming. Unclear whether isoprene-exposed plants (e.g., Arabidopsis in [74]) are more resistant to bacterial infection because of isoprene induction of SA synthesis and activation of SA-dependent defenses ahead of infection (no priming) or because isoprene also primes SA-dependent defenses for a more robust defensive behavior upon infection. |
| Stress signal | Increased production after stress and role as transmission signal of stresses to other parts of the plant and/or to neighboring plants | PRO: Isoprene emission changes rapidly in response to wounding and temperature [75]. Knocking-down (by RNAi) or inhibiting (by fosmidomycin) isoprene emission negatively impacts thermotolerance [30,33,59]. CON: Isoprene production occurs constitutively in several plants without apparent association to environmental stress [22]. Isps gene promoter activity takes place in roots and especially in developing lateral roots [65] during normal growth. Isoprene-emitting transgenic lines show positive effects on growth and development in unstressed plants [51]. |
| Hormone | Regulation, at very low concentration, of plant processes at the gene level | PRO: Isoprene acts as a negative regulator of ROS production by modulating gene expression [51,58,65,66]. Isoprene can both positively and negatively regulate plant growth and development [51,65]. CON: Isoprene is produced and emitted in very large amount by plants [22] which does not make it a hormone by definition. |
| Infochemical | Attraction of pollinators and insect predators or repulsion of phytophagous insects | PRO: Isoprene is a highly volatile molecule as are many infochemicals [76]. Tobacco plants emitting isoprene repel Manduca sexta larvae [77]. The parasitic wasp Diadegma semiclausum perceives isoprene by its chemoreceptors and is being repelled [31]. CON: Chrysomela populi (poplar leaf beetle) is unable to detect isoprene and show no preference for isoprene-emitting or non-emitting poplar plants [78,79]. Isoprene production occurs constitutively in several plants without apparent association to delivery of infochemical messages [22]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pollastri, S.; Baccelli, I.; Loreto, F. Isoprene: An Antioxidant Itself or a Molecule with Multiple Regulatory Functions in Plants? Antioxidants 2021, 10, 684. https://doi.org/10.3390/antiox10050684
Pollastri S, Baccelli I, Loreto F. Isoprene: An Antioxidant Itself or a Molecule with Multiple Regulatory Functions in Plants? Antioxidants. 2021; 10(5):684. https://doi.org/10.3390/antiox10050684
Chicago/Turabian StylePollastri, Susanna, Ivan Baccelli, and Francesco Loreto. 2021. "Isoprene: An Antioxidant Itself or a Molecule with Multiple Regulatory Functions in Plants?" Antioxidants 10, no. 5: 684. https://doi.org/10.3390/antiox10050684
APA StylePollastri, S., Baccelli, I., & Loreto, F. (2021). Isoprene: An Antioxidant Itself or a Molecule with Multiple Regulatory Functions in Plants? Antioxidants, 10(5), 684. https://doi.org/10.3390/antiox10050684