
Maternal Supplementation with Herbal Antioxidants during Pregnancy in Swine

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Experimental Design
2.3. Evaluation of Vitamin Concentrations and Oxidant/Antioxidant Status of Sows and Piglets
2.4. Statistical Analysis
3. Results
3.1. Effects of Supplementation on Plasma Vitamin Concentrations in Sows and Piglets
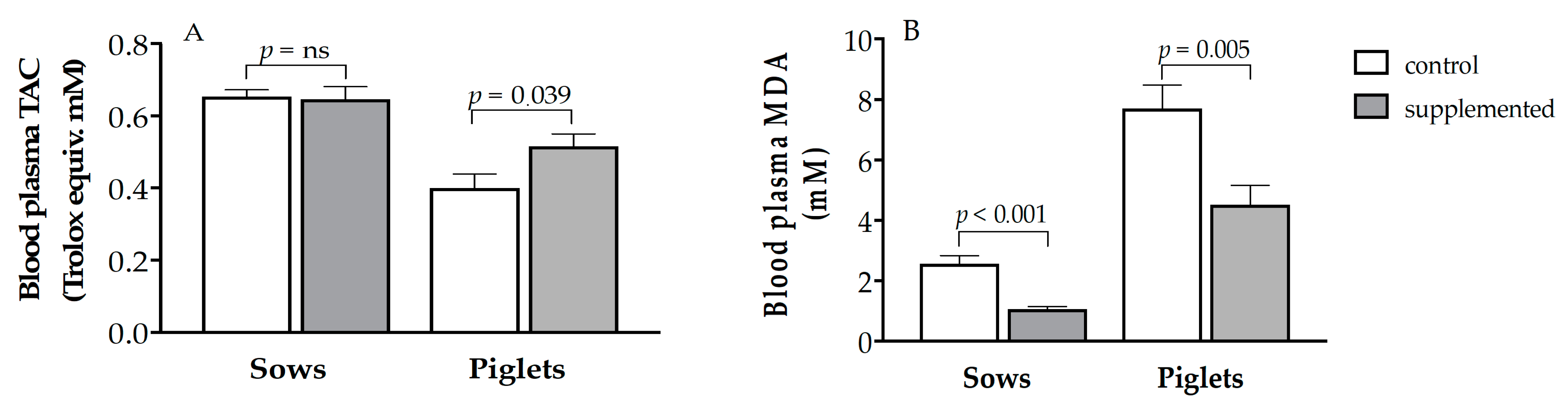
3.2. Effects of Supplementation on the Antioxidant Capacity and Oxidative Stress of Sows and Piglets
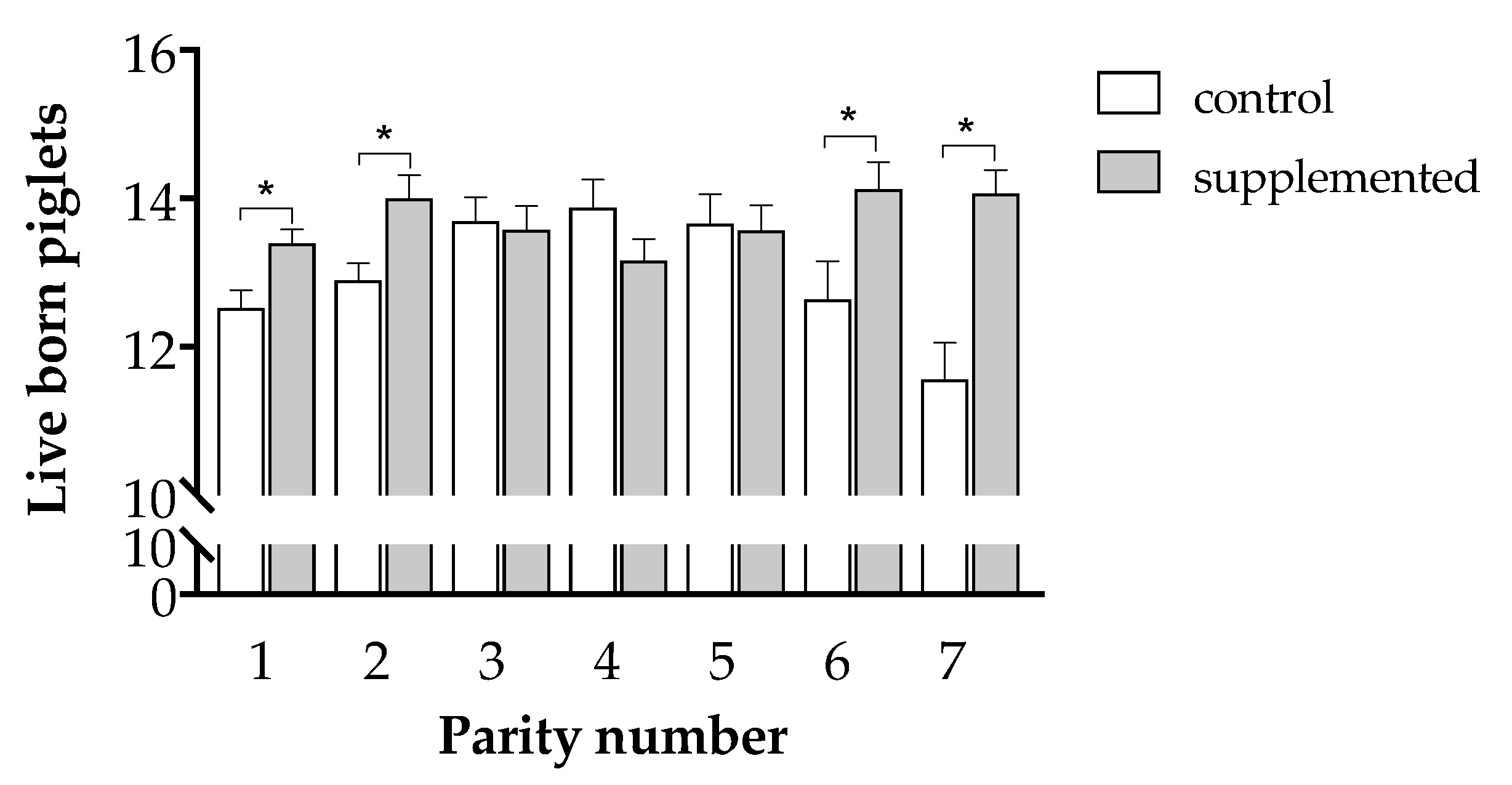
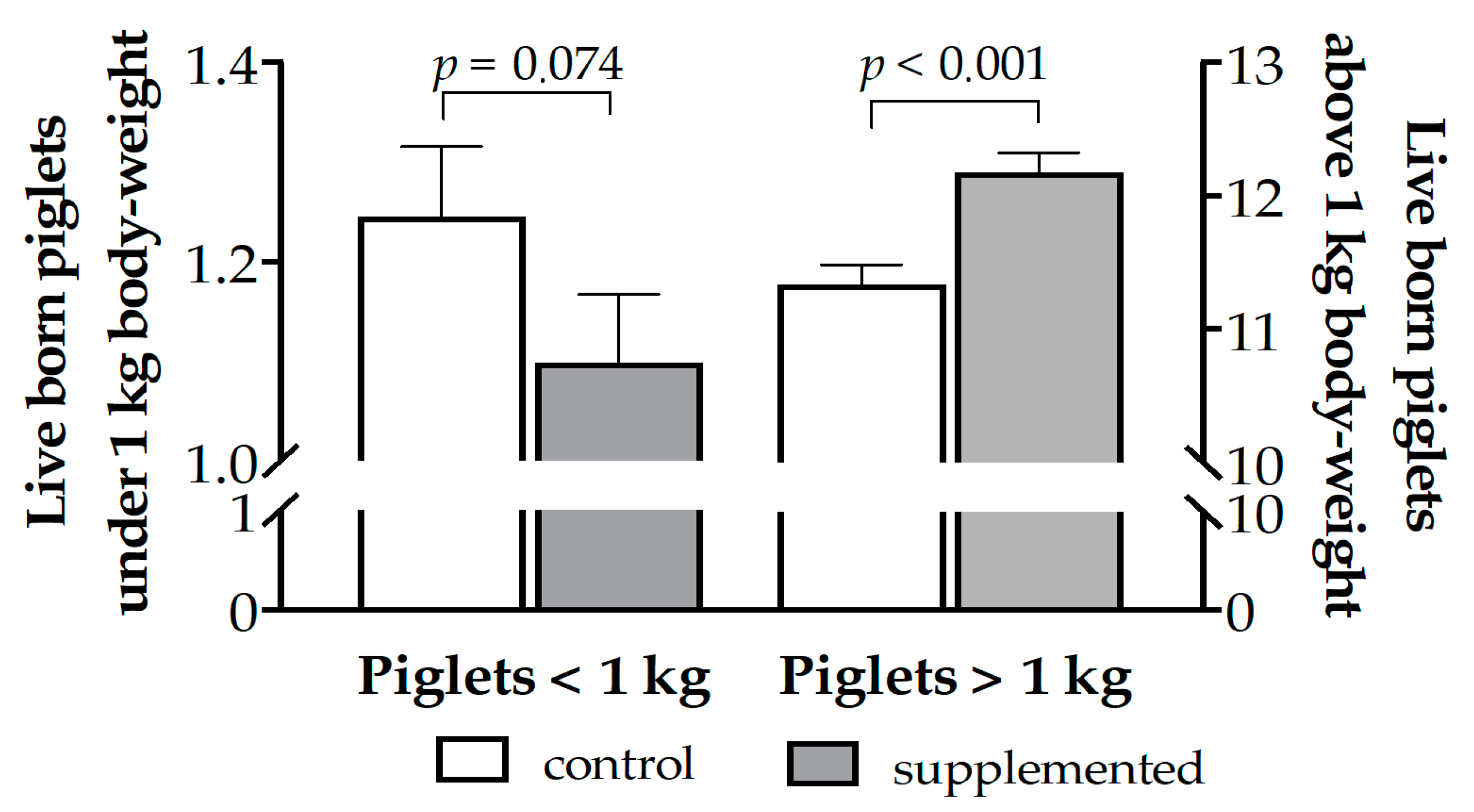
3.3. Effects of Maternal Supplementation on Litter Features at Farrowing
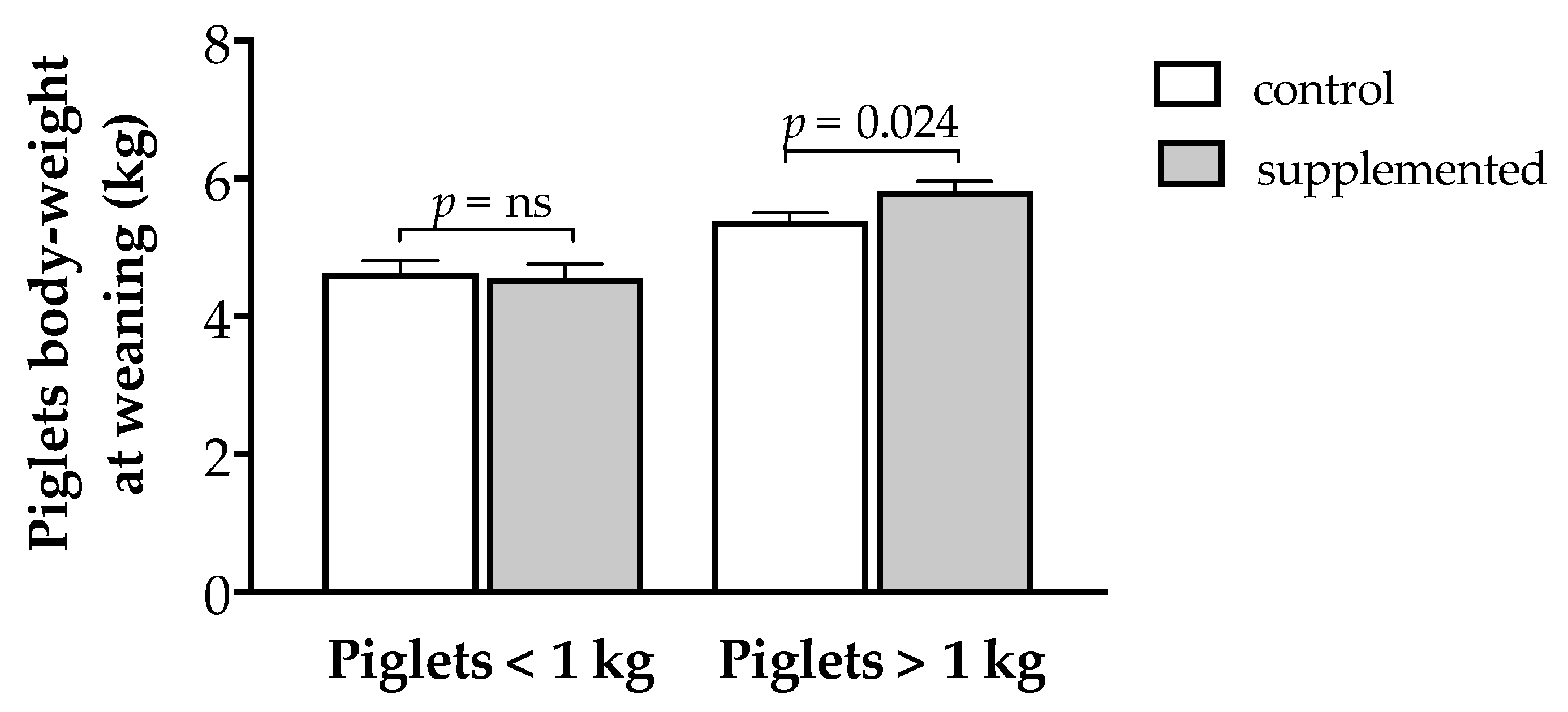
3.4. Effects of Maternal Supplementation on Postnatal Patterns of Growth and Development of Offspring
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van der Waaij, E.H.; Hazeleger, W.; Soede, N.M.; Laurenssen, B.F.A.; Kemp, B. Effect of excessive, hormonally induced intrauterine crowding in the gilt on fetal development on day 40 of pregnancy. J. Anim. Sci. 2010, 88, 2611–2619. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wu, G.; Bazer, F.; Wallace, J.; Spencer, T. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef]
- Foxcroft, G.R.; Dixon, W.T.; Novak, S.; Putman, C.T.; Town, S.C.; Vinsky, M.D.A. The biological basis for prenatal programming of postnatal performance in pigs. J. Anim. Sci. 2006, 84, E105–E112. [Google Scholar] [CrossRef]
- Milligan, B.N.; Fraser, D.; Kramer, D.L. Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights. Livest. Prod. Sci. 2002, 76, 181–191. [Google Scholar] [CrossRef]
- Edwards, S.A.; Baxter, E.M. Piglet mortality: Causes and prevention. In The Gestating and Lactating Sow; Farmer, C., Ed.; Wageningen Acad. Pub.: Wageningen, The Netherlands, 2015; Chapter 11; pp. 253–278. [Google Scholar]
- Bee, G. Effect of early gestation feeding, birth weight, and gender of progeny on muscle fiber characteristics of pigs at slaughter. J. Anim. Sci. 2004, 82, 826–836. [Google Scholar] [CrossRef]
- Gondret, F.; Lefaucheur, L.; Louveau, L.; Lebret, B.; Pichodo, X.; Le Cozler, Y. Influence of piglet birth weight on postnatal growth performance, tissue lipogenic capacity and muscle histological traits at market weight. Livest. Prod. Sci. 2005, 93, 137–146. [Google Scholar] [CrossRef]
- Gondret, F.; Lefaucheur, L.; Juin, H.; Louveau, I.; Lebret, B. Low birth weight is associated with enlarged muscle fiber area and impaired meat tenderness of the longissimus muscle in pigs. J. Anim Sci. 2006, 84, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Rehfeldt, C.; Pas, M.; Wimmers, K.; Brameld, J.M.; Nissen, P.M.; Berri, C.; Valente, L.M.P.; Power, D.M.; Picard, B.; Stickland, N.C.; et al. Advances in research on the prenatal development of skeletal muscle in animals in relation to the quality of muscle-based food. I. Regulation of myogenesis and environmental impact. Animal 2011, 5, 703–717. [Google Scholar] [CrossRef]
- Quiniou, N.; Dagorn, J.; Gaudre, D. Variation of piglets’ birth weight and consequences on subsequent performance. Livest. Prod. Sci. 2002, 78, 63–70. [Google Scholar] [CrossRef]
- Gonzalez-Bulnes, A.; Astiz, S.; Ovilo, C.; Lopez-Bote, C.J.; Torres-Rovira, L.; Barbero, A.; MAyuso, M.; Garcia-Contreras, C.; Vazquez-Gomez, M. Developmental Origins of Health and Disease in swine: Implications for animal production and biomedical research. Theriogenology 2016, 86, 110–119. [Google Scholar] [CrossRef]
- Karowicz-Bilinska, A.; Suzin, J.; Sieroszewski, P. Evaluation of oxidative stress indices during treatment in pregnant women with intrauterine growth retardation. Med. Sci. Monit. 2002, 8, CR211–CR216. [Google Scholar]
- Sales, F.; Peralta, O.A.; Narbona, E.; McCoard, S.; De los Reyes, M.; Gonzalez-Bulnes, A.; Parraguez, V. Hypoxia and oxidative stress are associated with reduced fetal growth in twin and undernourished sheep pregnancies. Animals 2018, 8, 217. [Google Scholar] [CrossRef]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.È.; Durak, İ. Role of Oxidative Stress in Intrauterine Growth Restriction. Gynecol. Obstet. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Narang, M.; Banerjee, B.; Basu, S. Oxidative stress in term small for gestational age neonates born to undernourished mothers: A case control study. BMC Pediatr. 2004, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Kamath, U.; Rao, G.; Kamath, S.U.; Rai, L. Maternal and fetal indicators of oxidative stress during intrauterine growth retardation (IUGR). Indian J. Clin. Biochem. 2006, 21, 111. [Google Scholar] [CrossRef]
- Parraguez, V.H.; Atlagich, M.; Araneda, O.; García, C.; Muñoz, A.; De los Reyes, M.; Urquieta, B. Effects of antioxidant vitamins on newborn and placental traits in gestations at high altitude: Comparative study in high and low altitude native sheep. Reprod. Fertil. Dev. 2011, 23, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Parraguez, V.H.; Urquieta, B.; De los Reyes, M.; González-Bulnes, A.; Astiz, S.; Muñoz, A. Steroidogenesis in sheep pregnancy with intrauterine growth retardation by high-altitude hypoxia: Effects of maternal altitudinal status and antioxidant treatment. Reprod. Fert. Dev. 2013, 25, 639–645. [Google Scholar] [CrossRef]
- Sales, F.; Peralta, O.A.; Narbona, E.; McCoard, S.; Lira, R.; De los Reyes, M.; González-Bulnes, A.; Parraguez, V.H. Maternal supplementation with antioxidant vitamins in sheep results in increased transfer to the fetus and improvement of fetal antioxidant status and development. Antioxidants 2019, 8, 59. [Google Scholar] [CrossRef]
- Parraguez, V.H.; Sales, F.; Peralta, O.A.; Narbona, E.; Lira, R.; De los Reyes, M.; González-Bulnes, A. Supplementation of underfed twin-bearing ewes with herbal vitamins c and e: Impacts on birth weight, postnatal growth, and pre-weaning survival of the lambs. Animals 2020, 10, 652. [Google Scholar] [CrossRef]
- Szczubiał, M. Effect of supplementation with vitamins E, C and β-carotene on antioxidative/oxidative status parameters in sows during the postpartum period. Polish J. Vet. Sci. 2015, 18, 299–305. [Google Scholar] [CrossRef]
- Pinelli-Saavedra, A.; Scaife, J.R. Pre- and postnatal transfer of vitamins E and C to piglets in sows supplemented with vitamin E and vitamin C. Livest. Prod. Sci. 2005, 97, 231–240. [Google Scholar] [CrossRef]
- González-Añover, P.; González-Bulnes, A. Maternal age modulates the effects of early-pregnancy L-proline supplementation on the birth-weight of piglets. Anim. Reprod. Sci. 2017, 181, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Federation Animal Science Society. Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching, 3rd ed.; Federation Animal Science Society: Champaign, IL, USA, 2010; p. 177. [Google Scholar]
- PIC Nutrient Specifications Manual. 2016 Edition. Available online: https://www.pic.com/wp-content/uploads/sites/3/2018/10/Nutrient-Specifications-Manual_2016_English_Metric.pdf (accessed on 10 March 2021).
- Chavez, E.R. Supplemental value of ascorbic acid during late gestation on piglet survival and early growth. Can. J. Anim. Sci. 1983, 63, 683–687. [Google Scholar] [CrossRef]
- Brown, R.G.; Harris, W.H.; Cummings, J.N. Ascorbate metabolism in swine, influence of maternal hypoxia on fetal tissue ascorbate levels. Can. J. Physiol. 1972, 50, 407–410. [Google Scholar] [CrossRef]
- Mahan, D.C.; Ching, S.; Dabrowski, K. Developmental aspects and factors influencing the synthesis and status of ascorbic acid in the pig. Annu. Rev. Nutr. 2004, 24, 79–103. [Google Scholar] [CrossRef] [PubMed]
- Ching, S.; Mahan, D.C.; Ottobre, J.S.; Dabrowski, K. Ascorbic acid synthesis in fetal and neonatal pigs and in pregnant and postpartum sows. J. Nutr. 2001, 131, 1997–2001. [Google Scholar] [CrossRef] [PubMed]
- Padayatty, S.J.; Levine, M. Vitamin C physiology: The known and the unknown and Goldilocks. Oral Dis. 2016, 22, 463–493. [Google Scholar] [CrossRef]
- Sosnowska, A.; Kawęcka, M.; Jacyno, E.; Kołodziej-Skalska, A.; Kamyczek, M.; Matysiak, B. Effect of dietary vitamins E and C supplementation on performance of sows and piglets. Acta Agric. Scand. Sect. A Anim. Sci. 2012, 61, 196–203. [Google Scholar] [CrossRef]
- Pinelli-Saavedra, A.; Calderón de la Barca, A.M.; Hernández, J.; Valenzuela, R.; Scaife, J.R. Effect of supplementing sows’ feed with a-tocopherol acetate and vitamin C on transfer of α-tocopherol to piglet tissues, colostrum, and milk: Aspects of immune status of piglets. Res. Vet. Sci. 2008, 85, 92–100. [Google Scholar] [CrossRef]
- Mavromatis, J.; Koptopoulos, G.; Kyriakis, S.C.; Papasteriadis, A.; Saoulidis, K. Effects of a-tocopherol and selenium on pregnant sows and their piglets’ immunity and performance. J. Vet. Med. A 1999, 46, 545–553. [Google Scholar] [CrossRef]
- Umesiobi, D.O. Vitamin E supplementation to sows and effects on fertility rate and subsequent body development of their weanling piglets. J. Agric. Rural Dev. Trop. Subtrop. 2009, 110, 155–168. [Google Scholar]
- Hidiroglou, M.; Farnworth, E.; Butler, G. Vitamin E and fat supplementation of sows and the effect on tissue vitamin E concentrations in their progeny. Reprod. Nutr. Dev. 1993, 33, 557–565. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hidiroglou, M.; Batra, T.R.; Farnworth, E.R.; Markham, F. Effect of vitamin E supplementation on immune status and a-tocopherol in plasma of piglets. Reprod. Nutr. Dev. 1995, 35, 443–450. [Google Scholar] [CrossRef]
- Gaykwad, C.K.; De, U.K.; Jadhavb, S.E.; Chethanc, G.E.; Akhilesha Sahood, N.R.; Mondala, D.B.; Guard, G.K.; Vermae, M.R.; Chaudhuri, P. Adding α-tocopherol-selenium and ascorbic acid to periparturient sow diets influences hemogram, lipid profile, leptin, oxidant/antioxidant imbalance, performance and neonatal piglet mortality. Res. Vet. Sci. 2019, 125, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Berchieri-Ronchi, C.B.; Kim, S.W.; Zhao, Y.; Correa, C.R.; Yeum, K.-J.; Ferreira, A.L.A. Oxidative stress status of highly prolific sows during gestation and lactation. Animal 2011, 5, 1774–1779. [Google Scholar] [CrossRef] [PubMed]
- Demchuk, M.; Buchko, O.; Lis, M.W.; Niedziółka, J.W. Performance of the antioxidant protection in blood of highly prolific sows before and after farrowing. Performance of the antioxidant protection in blood of highly prolific sows before and after farrowing. Large Anim. Rev. 2014, 20, 135–139. [Google Scholar]
- Yin, J.; Ren, W.; Liu, G.; Duan, J.; Yang, G.; Wu, L.; Li, T.; Yin, Y. Birth oxidative stress and the development of an antioxidant system in newborn piglets. Free Radic. Res. 2013, 47, 1027–1035. [Google Scholar] [CrossRef]
- Zhao, Y.; Kim, S.W. Oxidative stress status and reproductive performance of sows during gestation and lactation under different thermal environments. Asian Australas. J. Anim. Sci. 2020, 33, 722–731. [Google Scholar] [CrossRef]
- Pasqualotto, E.B.; Agarwal, A.; Sharma, R.K.; Izzo, V.M.; Pinotti, J.A.; Joshi, N.J.; Rose, B.I. Effect of oxidative stress in follicular fluid on the outcome of assisted reproductive procedures. Fertil. Steril. 2004, 81, 973–976. [Google Scholar] [CrossRef]
- Tamura, H.; Takasaki, A.; Miwa, I.; Taniguchi, K.; Maekawa, R.; Asada, H.; Taketani, T.; Matsuoka, A.; Yamagata, Y.; Shimamura, K.; et al. Oxidative stress impairs oocyte quality and melatonin protects oocytes from free radical damage and improves fertilization rate. J. Pineal Res. 2008, 44, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Soedea, N.M.; Langendijkb, P.; Kemp, B. Reproductive cycles in pigs. Anim. Reprod. Sci. 2011, 124, 251–258. [Google Scholar] [CrossRef]
- Bazer, F.W.; Thatcher, W.W.; Martinat-Botte, F.; Terqui, M. Conceptus development in large white and prolific Chinese Meishan pigs. J. Reprod. Fertil. 1988, 84, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Youngs, C.R.; Ford, S.P.; McGinnis, L.K.; Anderson, L.H. Investigations into the control of litter size in swine: I. Comparative studies on in vitro development of Meishan and Yorkshire preimplantation embryos. J. Anim. Sci. 1993, 71, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.E.; Biensen, N.J.; Ford, S.P. Novel insight into the control of litter size in pigs, using placental efficiency as a selection tool. J. Anim. Sci. 1999, 77, 1654–1658. [Google Scholar] [CrossRef]
- Webel, S.K.; Dziuk, P.J. Effect of stage of gestation and uterine space on prenatal survival in the pig. J. Anim. Sci. 1974, 38, 960–963. [Google Scholar] [CrossRef] [PubMed]
- Young, M.G.; Tokach, M.D.; Aherne, F.X.; Main, R.G.; Dritz, S.S.; Goodband, R.D.; Nelssen, J.L. Effect of sow parity and weight at service on target maternal weight and energy for gain in gestation. J. Anim. Sci. 2005, 83, 255–261. [Google Scholar] [CrossRef]
- Kim, S.W.; Weaver, A.C.; Shen, Y.B.; Zhao, Y. Improving efficiency of sow productivity: Nutrition and health. J. Anim. Sci. Biotechnol. 2013, 4, 26. [Google Scholar] [CrossRef]
- Hoving, L.L.; Soede, N.M.; van der Peet-Schwering, C.M.C.; Graat, E.A.M.; Feitsma, H.; Kemp, B. An increased feed intake during early pregnancy improves sow body weight recovery and increases litter size in young sows. J. Anim. Sci. 2011, 89, 3542–3550. [Google Scholar] [CrossRef]
- Noblet, J.; Etienne, M. Metabolic utilization of energy and maintenance requirements in pregnant sows. Livest. Prod. Sci. 1987, 16, 243–257. [Google Scholar] [CrossRef]
- Noblet, J.; Dourmad, J.Y.; Etienne, M. Energy utilization in pregnant and lactating sows: Modeling of energy requirements. J. Anim. Sci. 1990, 68, 562–572. [Google Scholar] [CrossRef]
- Morrow, W.E.M.; Leman, A.D.; Williamson, N.B.; Morrison, R.B.; Robinson, R.A. An epidemiological investigation of reduced second-litter size in swine. Prev. Vet. Med. 1992, 12, 15–26. [Google Scholar] [CrossRef]
- Hoving, L.L.; Soede, N.M.E.; Graat, A.M.; Feitsma, H.; Kemp, B. Effect of live weight development and reproduction in first parity on reproductive performance of second-parity sows. Anim. Reprod. Sci. 2010, 122, 82–89. [Google Scholar] [CrossRef] [PubMed]
- de Jong, E.; Appeltant, R.; Cools, A.; Beek, J.; Boyen, F.; Chiers, K.; Maes, D. Slaughterhouse examination of culled sows in commercial pig herds. Livest. Sci. 2014, 167, 362–369. [Google Scholar] [CrossRef]
- Roy, C.; Lavoie, M.; Richard, G.; Archambault, A.; Lapointe, J. Evidence that oxidative stress is higher in replacement gilts than in multiparous sows. J. Anim. Physiol. Anim. Nutr. 2016, 100, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Flowers, W.L.; Saraiva, A.; Yeum, K.-J.; Kim, S.W. Effect of social ranks and gestation housing systems on oxidative stress status, reproductive performance, and immune status of sows. J. Anim. Sci. 2013, 91, 5848–5858. [Google Scholar] [CrossRef] [PubMed]
- Smit, M.; Spencer, J.; Almeida, F.; Patterson, J.; Chiarini, H.; Dyck, M.; Foxcroft, G. Consequences of a low litter birth weight phenotype for postnatal lean growth performance and neonatal testicular morphology in the pig. Animal 2013, 7, 1681–1689. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of Parity | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Control | 148 | 111 | 75 | 50 | 54 | 47 | 38 |
| Treated | 136 | 86 | 73 | 70 | 51 | 47 | 41 |
| Total | 284 | 197 | 148 | 120 | 105 | 94 | 79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parraguez, V.H.; Sales, F.; Peralta, O.A.; De los Reyes, M.; Campos, A.; González, J.; Peralta, W.; Cabezón, C.; González-Bulnes, A. Maternal Supplementation with Herbal Antioxidants during Pregnancy in Swine. Antioxidants 2021, 10, 658. https://doi.org/10.3390/antiox10050658
Parraguez VH, Sales F, Peralta OA, De los Reyes M, Campos A, González J, Peralta W, Cabezón C, González-Bulnes A. Maternal Supplementation with Herbal Antioxidants during Pregnancy in Swine. Antioxidants. 2021; 10(5):658. https://doi.org/10.3390/antiox10050658
Chicago/Turabian StyleParraguez, Víctor H., Francisco Sales, Oscar A. Peralta, Mónica De los Reyes, Alfonso Campos, Javier González, Wolfgang Peralta, Camila Cabezón, and Antonio González-Bulnes. 2021. "Maternal Supplementation with Herbal Antioxidants during Pregnancy in Swine" Antioxidants 10, no. 5: 658. https://doi.org/10.3390/antiox10050658
APA StyleParraguez, V. H., Sales, F., Peralta, O. A., De los Reyes, M., Campos, A., González, J., Peralta, W., Cabezón, C., & González-Bulnes, A. (2021). Maternal Supplementation with Herbal Antioxidants during Pregnancy in Swine. Antioxidants, 10(5), 658. https://doi.org/10.3390/antiox10050658