Abstract
Kiwicha (Amaranthus caudatus) is considered one of the few multipurpose pseudocereals for its potential use not only as a source of nutrients and fiber but also for its bioactive compounds. In recent years, antioxidant peptides are commonly used as functional ingredient of food. Herein, a kiwicha protein isolate (KPI), obtained from kiwicha defatted flour (KDF), was hydrolyzed by Bioprotease LA 660, a food-grade endoprotease, under specific conditions. The resulting kiwicha protein hydrolysates (KPHs) were chemically characterized and their digestibility and antioxidant capacity were evaluated by in vitro cell-free experiments owing to their measure of capacity to sequester DPPH free radical and reducing power. KPHs showed higher digestibility and antioxidant capacity than intact proteins into KPI. Therefore, the results shown in this study indicate that KPHs could serve as an adequate source of antioxidant peptides, representing an effective alternative to the generation of functional food.
1. Introduction
According to the Food and Agriculture Organization of the United Nations (FAO) we must ensure that food is nutritive and accessible to entire population and that natural resources management preserves the functions of ecosystems to support the satisfaction of current and future human needs. Green industries, such as industrial agricultural emerging crops, offer opportunities for more long-term benefits through sustainable regenerative farming practices. Amaranth, an underutilized crop and a cheap source of proteins, minerals, vitamin A, and vitamin C, seems to be a future crop which can substantiate this demand due to its tremendous yield potential and nutritional qualities, also recently gained worldwide attention [1]. According to the World Health Organization (WHO) and FAO, dietary patterns along with lifestyle habits are important modifiable risk factors in relation to the development of diseases such as chronic noncommunicable diseases (NCDs), major contributor to the global burden of disease, and account for up to 72% of worldwide deaths [1]. Chronic low-grade inflammation, characterized by persistent elevated concentrations of circulating proinflammatory cytokines, across the life span has been associated with the development of both age and diet-related NCDs, including obesity, cardiometabolic diseases, many cancers, respiratory and auto-immune disorders, arthritis, and depression [2]. Persistent inflammation increases the production of reactive oxygen species (ROS) and activates mediators of inflammation as well as suppresses antioxidant defense mechanisms ultimately contributing to oxidative stress which promote development and progression of NCDs [3]. Dietary proteins are a source of bioactive peptides that can exert biological functions and promote health. This current trend towards the use of diet as a global strategy to reduce the incidence and severity of diseases has led to the search for new plant protein sources as an alternative therapy to classical pharmacotherapy.
Amaranths are pseudocereals which head a group of plants with great potentiality to prevent malnutrition in countries with low-income and food-deficit [4]. At the present time, Amaranth is considered one of the few versatile crops which can supply seeds in great quantities that are able to be used in different ways, as tasty leafy vegetables with high nutritional quality, as pseudocereals, and even as food and animal feed [4,5,6]. Grain amaranths are gluten-free compared to others such as wheat, rice and oat [7], and contain 30% more protein and complete set of amino acids [4]. Nowadays, the amaranth seeds are known as kiwicha [8].
The protein composition of amaranths is similar to proteins composition of legumes and crucifers, nevertheless, qualitatively and quantitatively, the protein of their seeds is higher than legumes and cereals [9,10,11,12]. Other pseudocereals such as quinoa (Chenopodium quinoa L.), considered as nonconventional sources of protein with excellent nutritional value, also showed a slightly lower protein content in comparison with amaranths grains [13]. Besides the essential role of nutrition in cancer [14], obesity, diabetes, cardiovascular [15], and even in neurodegenerative diseases, which are the leading causes of global deaths, the fact that they include good dietary sources could mitigate or prevent these disorders [1]. Thereby, in the last years, the interest of investigation in bioactive compounds from food has increased as an alternative to pharmacological treatment. The introduction of amaranth seeds into the diet has been linked not only with health promotion but also with prevention of diseases [16].
The main research activities focused on amaranths have been concentrated on their extraordinary nutritive value because of the content of proteins, good fats and bioactive compounds with antihyperlipidemic, antidiabetic [17], antihypercholesterolemic effects [18], and antioxidant [19,20] and antimicrobial activities [21]. Today, amaranth is a traditional food sold mainly as an artisanal product that has been indicated as “functional food”, due to its properties to promote health [22,23,24]. Peptides with low molecular weight (MW) extracted from food proteins can exert different biological properties such as antioxidant, anti-inflammatory, anti-hypertensive, and antibacterial activities. This has resulted in intense research into the likely applications of these kinds of peptides, as those from kiwicha, in the treatment or the prevention of different diseases [8,25].
Oxidative stress appears because of an imbalance between the amounts of produced oxidants and the antioxidant defense mechanisms. Free radicals, such as reactive nitrogen species (RNS) or ROS generated during physiological aerobic metabolism and pathological inflammatory processes, mediate oxidative stress. These ROS and RNS induce damage to DNA as base and nucleotide modifications or strand breaks and oxidation of protein amino acid residues leading to the formation of aggregates by cross-linking, loss of activity in enzymes, disturbance of metabolic pathways or even cell death. Oxidative species can also react with other molecules as low-density lipoprotein (LDL) triggering its oxidation and contributing to increased vascular permeability for example. Thus, oxidative stress contributes to many diseases such as inflammatory and cardiovascular diseases, metabolic syndrome and dementia. Cell membrane integrity can be disrupted by oxidized lipids [26].
Since studies related to antioxidant effect of amaranth-derived peptides are limited, the main objective of this study was to investigate the antioxidant properties of kiwicha protein hydrolysates (KPHs). For that, the specific goals were (i) the isolation of kiwicha protein products and their chemical characterization, (ii) the assessment of the digestibility of KPHs, and (iii) the evaluation of antioxidant effects of the KPHs.
2. Materials and Methods
2.1. Plant Material and Chemicals
Amaranthus caudatus seed flour was provided by STURLA, S.R.L. (Buenos Aires, Argentina). Bioprotease LA-660 was purchased from BIOCON (Barcelona, Spain). All chemicals compounds (reagents and solvents) were provided by Sigma Chemical (St. Louis, MO, USA), Bachem AG, and Gibco, and were of analytical grade and.
2.2. Protein Solubility Curve
Kiwicha defatted flour (KDF, 2 g) was dissolved in H2O (100 mL) and pH was adjusted to 12 with NaOH. Briefly, pH was taken successively, in 10, 8, 6, 4, and 2 until constant pH, in each point, with HCl. At each pH-point an aliquot was taken, in duplicate, which was centrifuged for 15 min at 12,000 g. The supernatants recovered were measured in nitrogen content.
2.3. Preparation of Kiwicha Protein Isolate (KPI)
The method of Lqari et al. [27] was used to obtain the KPI. Briefly, KDF was extracted with Na2SO3 (0.25% w/v) for 1 h at pH 11. Then, the extract was centrifugated at 9500 g for 15 min. On the one hand, the supernatant was recovered and on the other hand, the pellet was extracted again. The supernatants obtained in both centrifugations were adjusted to the isoelectric point (pI) of kiwicha proteins (pH 4 during 30 min) [12,28]. The precipitate was washed with distilled H2O, adjusted to pH 4 and centrifuged in order to remove other non-protein compounds and residual salts. Eventually, the proteins of this precipitate were lyophilized (stored at room temperature).
2.4. Hydrolysis of Kiwicha Protein Isolate
KPI hydrolysis was performed under continuous stirring, using a LAMBDA MINIFOR fermenter-bioreactor (Zurich, Switzerland), at controlled conditions of pH and temperature. The KPI was resuspended in distilled H2O (10% w/v) at 50 °C. Bioprotease LA-660 was added at a ratio enzyme/substrate = 0.3 AU/g protein (pH 8) for 5, 10, 15, 30, and 60 min. To ensure the complete inactivation of remaining enzyme activity each time-point sample was stopped by raising temperature at 85 °C during 20 min. KPH was constituted by the supernatant obtained after centrifugation at 9500 g during 15 min. The KPHs obtained at different times of using Bioprotease were designated as 5, 10, 15, 30, and 60, where the number indicates the hydrolysis time in minutes.
2.5. Evaluation of Hydrolysis Degree (HD)
The percentage of peptide bonds cleaved is defined as the HD. It was calculated by a method to determine α-amino groups, the 2,4,6-trinitrobenzene sulfonic acid method (TNBS) [29]. A sample that had been hydrolyzed up to 100% in HCl (6N) at 110 °C for 24 h was used as measure of the total number of free amino groups.
2.6. Compositional Analysis of Kiwicha Protein Products
The concentration of protein was determined as % nitrogen content × 6.25 by elemental microanalysis using a Leco CHNS932 analyzer (St. Joseph, MI, USA). The gravimetric method was used to determine the total dietary fiber [30]. According to the ignition method (550 °C during 36 h) ash content was determined. Measurement of total phenolic compounds and soluble sugars was undertaken using standard curves of chlorogenic acid for the first and glucose for the second [31,32].
2.7. Determination of Amino Acid Composition by Ultra-High-Performance Liquid Chromatography (HPLC)
Amino acid composition was determined according to the method of Alaiz et al. [33] with slight modifications. In this sense, samples were hydrolyzed by incubation in 6 N HCl at 110 °C for 24 h in tubes sealed under nitrogen. Amino acids were determined in the acid hydrolysate by ultra-high-performance liquid chromatography (Acquity Arc, Waters, USA), after derivatization with diethyl ethoxymethylenemalonate, using D,L-α-aminobutyric acid as internal standard, and a 3 mm × 150 mm reversed-phase column (XSelect HSS T3 XP, 2.5 μm; Waters). A binary gradient system with the solvents (A) 25 mM sodium acetate 0.02% sodium azide (pH 6.0), and (B) acetonitrile was used. Calibration curves for each amino acid were developed using a mix of amino acid standard at the same hydrolysis conditions of the samples (Merck, Madrid, Spain) and the resultant peaks were analyzed with EMPOWER software (Waters, Santa Clara, CA, USA). Besides, tryptophan content was assessed according to the Yust et al. method [34].
2.8. Pepsin-Pancreactic Digestibility
The method established by Sindayikengera and Xia [35] was slightly modified to determine the protein digestibility in vitro. The sample (0.1 g) was added to a 50 mL tube, with 7.5 mL of HCl 0.1 M containing 1.5 mg of pepsin and then it was incubated at 37 °C for 2 h. Then the suspension was neutralized with 0.5 M NaOH, then treated with pancreatin (1.875 mg) in 7.5 mL of 0.2 M phosphate buffer (pH 8.0). The mixture was gently shaken and incubated for additional 2 h at 37 °C. After this, the sample was treated with 5 mL of 10% TCA (trichloroacetic acid) and centrifuged at 10,000 g during 20 min at room temperature. Supernatant proteins were estimated as % nitrogen content × 6.25 by elemental microanalysis in a Leco CHNS932 analyzer. Protein digestibility was calculated as percentage by the ratio of protein in supernatant to total protein in sample.
2.9. Molecular Weight (MW) Profile by Fast Protein Liquid Chromatography (FPLC)
MW profiles of protein from the kiwicha products was estimated using FPLC Akta Purifier 10 (GE Healthcare Bio-sciences AB, Switzerland). A prepacked chromatography column for high-performance size exclusion, Superose 12 10/300 GL, with a range for molecules with MWs from 1 to 300 kDa was used. Proteins with MWs known (GE Healthcare, UK) were used to calibrate the column: blue dextran (2000 kDa), aldolase (158 kDa), conalbumin (75 kDa), bovine serum albumin (67 kDa), ribonuclease A (13.7 kDa), and bacitracine (1.423 kDa). Calibration line was made using the logarithms of the MWs of these control proteins and their elution volumes. The elution was accomplished with 50 mL of sodium phosphate buffer (0.05 M), sodium chloride (0.15 M) and sodium azide (0.02% w/v) with a flow of 1 mL/min, at pH 7.5. Samples consisted of 500 μL of each one injected at a concentration of 30 mg/mL of protein. Their absorbance at 280 nm was measured to record the elution of the proteins.
2.10. Determination of Antioxidant Activity
2.10.1. DPPH Radical-Scavenging Activity
KPI and KPHs effect on scavenging of DPPH free radical was evaluated according to Wu, Chen, and Shiau [36]. In brief, 1.5 mL of each sample with 10 mg/mL of protein was added to 1.5 mL of DPPH (0.1 mmol/L in ethanol). This mix was shaken and kept at room temperature for 30 min. Then, absorbance was measured at 517 nm. Negative control was treated in the same manner using distilled water instead of sample. BHT at 0.08 mg/mL was used as positive standard. Radical DPPH has an absorption band centered at about 517 nm which disappears when an antiradical compound reduces it. Lower absorbance of the reaction mixture means higher DPPH scavenging activity. The scavenging effect was calculated as [(Acontrol − Asample)/Acontrol] × 100 (Asample and Acontrol are the absorbances respective to sample and control).
2.10.2. Reducing Power
KPI and KPHs ability to reduce iron (III) was assessed according to Oyaizu [37]. Briefly, 0.1 mL of each sample with 10 mg/mL of protein was added to 0.25 mL of sodium phosphate buffer (0.2 M) at pH 6.6 and 0.25 mL of potassiumferricyanide (0.03 M). This mixture was incubated at 50 °C during 20 min. Then 0.25 mL of trichloroacetic acid (0.6 M) was added and it was centrifuged at 1300× g for 10 min. In total, 500 µL of supernatant were mixed with 0.5 mL of distilled water and 0.1 mL of ferric chloride (3.7 mmol/L). After 10 min, the resulting solution absorbances were measured at 700 nm. Increased absorbance of the reaction mixture means increased reducing power. As positive standard BHT at 0.08 mg/mL was used.
2.11. Statistical Analysis
All values are expressed as arithmetic means ± standard deviations (SD). Graph Pad Prism Version 6.01 software (San Diego, CA, USA) was used to evaluate data. One-way analysis of variance (ANOVA) following Tukey’s multiple comparisons test as post hoc test were used to calculate the statistical significance of differences in each parameter among the groups. Less than 0.05 as p value was considered statistically significant.
3. Results and Discussion
3.1. Chemical Characterization of Kiwicha Protein Products and Protein Digestibility
Kiwicha seed flour was defatted and the resulting KDF presented a protein content of 15.88%, as shown in Table 1. The KPI was extracted from KDF in 30.87% yield by basic extraction and then by acidic precipitation. The resulting KPI showed 84.52% protein content. The chemical composition of KPHs was similar to KPI. These results are similar to those obtained in other pseudocereals of emerging crops as wheat and higher than others as quinoa [38]. The main difference between KPI and KPHs was higher ash content in the hydrolysates because of the addition of alkali to maintain the pH constant during hydrolysis.

Table 1.
Chemical composition of kiwicha protein products.
According to the Food and Agriculture Organization of the United Nations (FAO) to evaluate of the quality of dietary protein in human nutrition two factors must be considered: absolute and relative quantities of dietary indispensable amino acids and digestibility of protein [39]. In general, most protein from animal sources as eggs, beef, milk, casein and whey, has a higher digestibility than storage proteins of (pseudo)cereals [40]. Protein digestibility is dependent on internal and external factors of them. Some of these external factors are temperature, pH, ionic strength conditions and others as the presence of secondary molecules as antinutritional factors and emulsifiers that limit their digestibility (protease inhibitors as chymotrypsin and trypsin inhibitors), tannins and phytate. Internal factors examples are protein amino acid composition, protein folding and crosslinking. In order to increase the overall digestibility, processing of food is generally designed, affecting these external and internal factors [41]. Removal of protease inhibitors in protein isolate increases the KPI in vitro protein digestibility (77.97 ± 2.93) compared to the KDF (60.74 ± 0.04). The highest value was found for the KPH5, improving digestibility by over 20% with respect to the KDF. Then, the digestibility will decrease slightly, remaining more or less stable during the next 50 min of hydrolysis. This may be because of the generation of new peptides during hydrolysis with Bioprotease, thus, in dietary proteins some specific domains are very stable against digestion and were suggested to be able to cross the mucosal of gut barrier. It has been hypothesized that these peptides would be beneficial to human health, known as bioactive peptides [42]. Despite this, these digestibility values of KPHs are higher than those of other pseudocereals such as buckwheat and legumes such as pigeon pea [43,44].
3.2. Amino Acid Composition of Kiwicha Protein Products
The peptide activity might be influenced by amino acid profile in kiwicha products. There are nine essential amino acids, among the 20 types of standard amino acids, that adults cannot produce. The amino acid content in KDF, KPI, and KPHs is reported in Table 2, being the highest concentrations those related to glutamic acid (Glu), aspartic acid (Asp) and arginine (Arg). These profiles have similarities with the amino acid profiles reported in the literature [8]. The importance of these amino acids lies in that Asp is involved in the correct blood circulation, detoxification of toxins in liver and kidneys, and different steps of cellular metabolism and Glu is involved in the neurotransmission, energy transport, and nutrient absorption. Both of them are involved in electron donation mechanisms between ROS. Arg has been involved in nitric oxide synthesis regulation, immune regulation, and insulin, glucagon, growth hormone and prolactin release [26]. There are four essential amino acids identified as limiting amino acids: threonine and lysine in cereals, tryptophan in maize and sulfur amino acids in legumes. Legumes are frequently low in the sulfur-containing amino acids methionine and cysteine, while lysine is typically limiting in grains [45]. The analyses demonstrated that values obtained for kiwicha protein products were far higher than adults requirements established by Food and Agriculture Organization, World Health Organization and United Nations University (FAO/WHO/UNU) for indispensable dietary amino acid [39], reaching double the recommended dose for some of them. The predominant essential amino acids were aromatic amino acids (tyrosine, phenylalanine and tryptophan) and sulfur amino acids (methionine and cysteine), this being relevant since amino acids than contain sulfur represent a powerful part of antioxidant systems of cells [13]. Broadly, no significant differences were observed between protein products, indicating that the enzymatic hydrolysis process is a smooth process and does not harm the various amino acids, nor does it produce secondary metabolites.
Table 2.
Amino acid composition of kiwicha protein products and adult requirements.
3.3. Analysis of the Hydrolysis Degree and the Molecular Profile of Kiwicha Protein Products
The protein hydrolysates include free amino acids, small peptides, and large pep-tides. The extension of hydrolysis can be evaluated by the HD values and it can be used as a measure of the peptide chain length; with higher and lower values indicating shorter and longer lengths, respectively. The extensive hydrolysis of KPI was performed, as said before, using Bioprotease in the way represented in Figure 1. The rate of hydrolysis was faster during the first 5 min and then remained more stable for the following 55 min, reaching a maximum HD of 32.02% (in KPH60). Evaluation of single-stage enzymatic treatments of kiwicha protein using Bioprotease resulted in similar HD values as those obtained by other authors with Alcalase and higher than those obtained with Neutrase and Flavourzyme, all them food-grade enzymes [46].
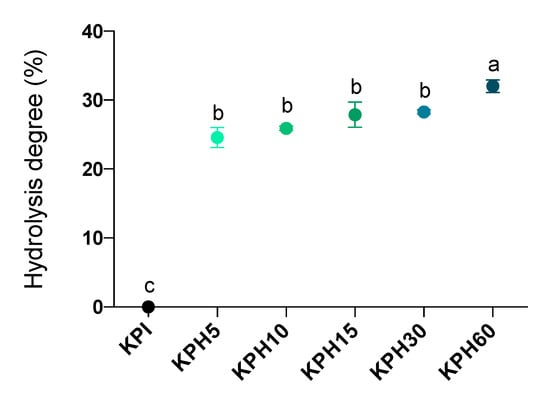
Figure 1.
Hydrolysis degree time-course of the KPI during enzymatic hydrolysis by Bioprotease at 5 (KPH5), 10 (KPH10), 15 (KPH15), 30 (KPH30) and 60 (KPH60) min. Data are expressed as the percentage of cleaved peptide bonds (left y-axis). Values are represented as means ± standard deviation of three determinations. Statistical differences are marked with different letters (p < 0.05).
The average molecular weight of hydrolysates is a leading factor that determines their biological properties. Reportedly, low-MW peptides, from 2 to 20 amino acids, are more biologically active when compared to their larger parent polypeptide/proteins [47]. As depicted in Figure 2, the protein molecular profile of KPHs showed that the peptide size was around 3.11–0.16 kDa (peaks d and e). This result showed that enzymatic hydrolysis using Bioprotease significantly reduced the protein MW of KPI (15.95 kDa, peak a). Nowadays, most bioactive peptides described have less than 1 kDa. The relationship between the hydrolysates MWs and their bioactivities in human health applications is essential in functional foods. It has been demonstrated that the presence of low-MW peptides enhances the absorption and the ability of amino acids to have bioactive effects [38]. The MW average of kiwicha peptides is lower than MW average of other pseudocereals’ peptides as wheat gluten protein hydrolysates, which after hydrolysis with Alcalase demonstrated to be constituted by peptides with sizes in the range of 1–300 kDa, mainly three of 30.08, 5.36, and 2.19 kDa [38]. In this sense, the smaller size of kiwicha peptides may be attributed to faster hydrolysis with Bioprotease compared to Alcalase. While Bioprotease increased HD up to 20% after 5 min, 10–15 min of hydrolysis with Alcalase are used to reach 20% HD [48].
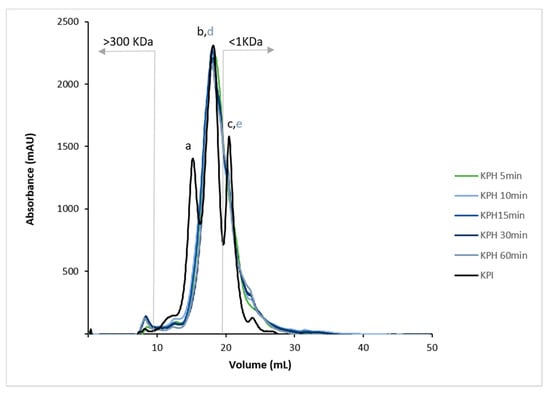
Figure 2.
Molecular weight (MW) profiles by FPLC (size-exclusion) chromatogram of KPI and KPH5, KPH10, KPH15, KPH30, and KPH60. MW ranging from 1 to 300 kDa.
3.4. Determination of the Antioxidant Activity of Kiwicha Protein Products
Antioxidant peptides are being accepted as food ingredients, as supplement in functional food and nutraceuticals. To further explore the possible antioxidant activity of kiwicha protein products, two assays were carried out: the capacity to sequester DPPH free radical (Figure 3A) and the FRAP (ferric reducing antioxidant power) (Figure 3B). After their enzymatic treatment with different proteases, the antioxidant power of kiwicha products could be enhanced due to the higher exposure of functional amino acid residues in the hydrolysate [49]. BHT was used as positive control in all of the experiments.
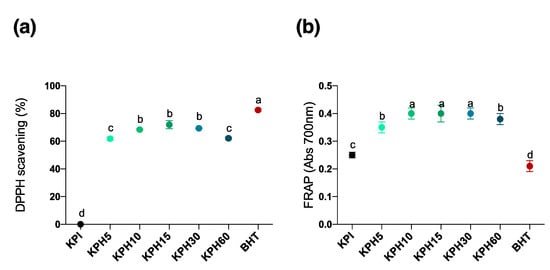
Figure 3.
(a) DPPH free radical scavenging activity and (b) FRAP (ferric reducing antioxidant power) of KPI and KPHs obtained with Bioprotease in an in vitro cell-free experiments. Values are represented as means ± standard deviation of three determinations. Statistical differences are marked with different letters (p < 0.05).
Immune response and the control of oxidative stress are two interconnected processes implicated in the maintenance of homeostasis [48]. Inflammation is an important normal immune response during lesions and infections. However, an excessive inflammation can contribute to several acute and chronic diseases characterized by uncontrolled production of proinflammatory cytokines such as tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1) and interleukin-6 (IL-6), eicosanoids derived from arachidonic acid, ROS, and adhesion molecules [50]. Hence, the fine control of the cellular pro-/anti-inflammatory microenvironment is crucial in health maintenance. In this sense, the current trend towards the use of diet as a global strategy to reduce the incidence and severity of diseases has led to the search for new plant protein sources as an alternative therapy to classical pharmacotherapy. Recent studies have shown that bioactive peptides from plant seeds such as lupin or hemp obtained by enzymatic hydrolysis can decrease oxidative stress and inflammatory response in cells of the innate immune system [51,52]. However, the targeting pathways of the immune system by these plant protein hydrolysates are unknown. Most of the food-derived peptides exhibited their anti-inflammatory activities mainly by inhibiting signaling components of either nuclear factor kappa-B (NF-κB) or mitogen-activated protein kinase (MAP)K pathway, which are the two major pathways involved in chronic inflammation following uncontrolled signal activation [53]. Inflammatory pathways impact the pathogenesis of a number of chronic diseases and involve common inflammatory mediators and regulatory pathways. Inflammatory stimuli activate intracellular signaling pathways that then activate production of inflammatory mediators. Inflammatory stimuli, including microbial products such as lipopolysaccharide (LPS), a main constituent of Gram-negative bacterial membrane, and proinflammatory cytokines such as TNF-α, mediate inflammation through interaction with the TLRs or cytokine receptor (TNFR). Receptor activation triggers important intracellular signaling pathways, including the MAPK and NF-κB, whose transcription plays important roles in inflammatory and immune response [54]. Analysis of the antioxidant activity of kiwicha protein products showed that all KPHs enhanced antioxidant activity with respect to the KPI, reaching a maximum DPPH scavenging effect of 77.91% similar to positive control (BHT), and showed much higher levels than BHT reducing power. This antioxidant bioactivity value is similar to that obtained by peptide hydrolysates derived from meat muscle as well as processed meat products like dry-cured ham [55] and better than others from plant sources such as oat, wheat or corn protein [56], which confirmed that identified KPHs may be a good source alternative to animal protein to obtain antioxidant peptides. However, further studies should be performed to elucidate the anti-inflammatory effects of kiwicha peptides.
4. Conclusions
Plant protein-based products supplemented in functional food and nutraceuticals are expected to grow considerably in the next years. Plant proteins, usually, present insufficient levels of at least one or even more essential amino acids thus limiting their biological value. However, antioxidant peptides in KPHs presented a balanced amino acid profile and a digestibility higher than intact proteins, hence, these results show that KPHs could be taken into account as an effective option within the functional food generation and would support their use in future clinical food trials.
Author Contributions
Conceptualization, S.M.-d.l.P., A.M.-L.; methodology, A.M.-L., A.V.-L.; software, S.M.-d.l.P.; validation, M.C.M.-L.; formal analysis, M.C.M.-L., S.M.-d.l.P.; investigation, M.C.M.-L., S.M.-d.l.P.; resources, A.M.-L., A.V.-L.; data curation, A.M.-L., A.V.-L.; writing—original draft preparation, M.C.M.-L.; writing—review and editing, S.M.-d.l.P.; visualization, S.M.-d.l.P.; supervision, M.C.M.-L.; project administration, M.C.M.-L.; funding acquisition, F.M., J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grant CYTED-2019/119RT0567 from the Spanish Ministry of Science, Innovation, and Universities.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rastogi, A.; Shukla, S. Amaranth: A new millennium crop of nutraceutical values. Crit. Rev. Food Sci. Nutr. 2013, 53, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.M.; Chen, L.-W.; Heude, B.; Bernard, J.Y.; Harvey, N.C.; Duijts, L.; Mensink-Bout, S.M.; Polanska, K.; Mancano, G.; Suderman, M.; et al. Dietary inflammatory index and non-communicable disease risk: A narrative review. Nutrients 2019, 11, 1873. [Google Scholar] [CrossRef]
- Jha, J.C.; Ho, F.; Dan, C.; Jandeleit-Dahm, K. A causal link between oxidative stress and inflammation in cardiovascular and renal complications of diabetes. Clin. Sci. 2018, 132, 1811–1836. [Google Scholar] [CrossRef] [PubMed]
- Das, S. Amaranths: The crop of great prospect. In Amaranthus: A Promising Crop of Future, 1st ed.; Springer: Singapore, 2016; p. 48. [Google Scholar] [CrossRef]
- Cheng, A. Shaping a sustainable food future by rediscovering long-forgotten ancient grains. Plant Sci. 2018, 269, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Coelho, L.M.; Silva, P.M.; Martins, J.T.; Pinheiro, A.C.; Vicente, A.A. Emerging opportunities in exploring the nutritional/functional value of amaranth. Food Funct. 2018, 9, 5499–5512. [Google Scholar] [CrossRef]
- Valcárcel-Yamani, B.; Lannes, S.D.S. Applications of quinoa (Chenopodium quinoa Willd.) and amaranth (Amaranthus spp.) and their influence in the nutritional value of cereal based foods. Food Public Health 2012, 2, 265–275. [Google Scholar] [CrossRef]
- Martinez-Lopez, A.; Millan-Linares, M.C.; Rodriguez-Martin, N.M.; Millan, F.; Montserrat-de la Paz, S. Nutraceutical value of kiwicha (Amaranthus caudatus L.). J. Funct. Foods 2020, 65, 103735. [Google Scholar] [CrossRef]
- Marcone, M.F. Evidence confirming the existence of a 7S globulin-like storage protein in Amaranthus hypochondriacus seed. Food Chem. 1999, 65, 533–542. [Google Scholar] [CrossRef]
- Singhal, R.S.; Kulkarni, P.R. Composition of the seeds of some Amaranthus species. J. Sci. Food Agric. 1988, 42, 325–331. [Google Scholar] [CrossRef]
- Singhal, R.S.; Kulkarni, P.R. Amaranths—An underutilized resource. Int. J. Food Sci. Technol. 1988, 23, 125–139. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Miralles, B.; Hernández-Ledesma, B. Release of multifunctional peptides from kiwicha. (Amaranthus caudatus) protein under in vitro gastrointestinal digestion. J. Sci. Food Agric. 2019, 99, 1225–1232. [Google Scholar] [CrossRef]
- Chirinos, R.; Pedrechis, R.; Velásquez-Sánchez, M.; Aquilar-Gálvez, A.; Campos, D. In vitro antioxidant and angiotensin I-converting enzyme inhibitory properties of enzymatically hydrolyzed quinoa (Chenopodium quinoa) and kiwicha (Amaranthus caudatus) proteins. Cereal. Chem. 2020, 97, 949–957. [Google Scholar] [CrossRef]
- Mayne, S.T.; Playdon, M.C.; Rock, C.L. Diet, nutrition, and cancer: Past, present and future. Nat. Rev. Clin. Oncol. 2016, 13, 504. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D. Dietary and policy priorities for cardiovascular disease, diabetes, and obesity: A comprehensive review. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef]
- Montoya-Rodríguez, A.; Gómez-Favela, M.A.; Reyes-Moreno, C.; Milán-Carrillo, J.; González de Mejía, E. Identification of bioactive peptide sequences from amaranth (Amaranthus hypochondriacus) seed proteins and their potential role in the prevention of chronic diseases. Compr. Rev. Food Sci. Food Saf. 2015, 14, 139–158. [Google Scholar] [CrossRef]
- Sangameswaran, B.; Jayakar, B. Anti-diabetic, anti-hyperlipidemic and spermatogenic effects of Amaranthus spinosus Linn. on streptozotocin-induced diabetic rats. J. Nat. Med. 2008, 62, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Girija, K.; Lakshman, K.; Udaya, C.; Sachi, G.S.; Divya, T. Anti–diabetic and anti–cholesterolemic activity of methanol extracts of three species of Amaranthus. Asian Pac. J. Trop. Biomed. 2011, 1, 133–138. [Google Scholar] [CrossRef]
- Alvarez-Jubete, L.; Wijngaard, H.; Arendt, E.K.; Gallagher, E. Polyphenol composition and in vitro antioxidant activity of amaranth, quinoa buckwheat and wheat as affected by sprouting and baking. Food Chem. 2010, 119, 770–778. [Google Scholar] [CrossRef]
- Tironi, V.A.; Añón, M.C. Amaranth proteins as a source of antioxidant peptides: Effect of proteolysis. Food Res. Int. 2010, 43, 315–322. [Google Scholar] [CrossRef]
- Maiyo, Z.C.; Ngure, R.M.; Matasyoh, J.C.; Chepkorir, R. Phytochemical constituents and antimicrobial activity of leaf extracts of three Amaranthus plant species. Afr. J. Biotechnol. 2010, 9, 3178–3182. [Google Scholar] [CrossRef]
- Loaiza, M.A.P.P.; López-Malo, A.; Jiménez-Munguía, M.T. Nutraceutical Properties of Amaranth and Chia Seeds. In Functional Properties of Traditional Foods, 1st ed.; Springer: Boston, MA, USA, 2016; pp. 189–198. [Google Scholar] [CrossRef]
- Rojas-Rivas, E.; Espinoza-Ortega, A.; Thomé-Ortiz, H.; Moctezuma-Pérez, S. Consumers’ perception of amaranth in Mexico. A traditional food with characteristics of functional foods. Br. Food J. 2019, 121, 1190–1202. [Google Scholar] [CrossRef]
- Rojas-Rivas, E.; Espinoza-Ortega, A.; Thomé-Ortíz, H.; Moctezuma-Pérez, S.; Cuffia, F. Understanding consumers’ perception and consumption motives towards amaranth in Mexico using the Pierre Bourdieu’s theoretical concept of Habitus. Appetite 2019, 139, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.; Fogacci, F.; Colletti, A. Potential role of bioactive peptides in prevention and treatment of chronic diseases: A narrative review. Br. J. Pharmacol. 2017, 174, 1378–1394. [Google Scholar] [CrossRef] [PubMed]
- Esfandi, R.; Walters, M.E.; Tsopmo, A. Antioxidant properties and potential mechanisms of hydrolyzed proteins and peptides from cereals. Heliyon 2019, 5, e01538. [Google Scholar] [CrossRef] [PubMed]
- Lqari, H.; Vioque, J.; Pedroche, J.; Millán, F. Lupinus angustifolius protein isolates: Chemical composition, functional properties and protein characterization. Food Chem. 2002, 76, 349–356. [Google Scholar] [CrossRef]
- Tapan, K.M.; Abdullatif, K.; Abeer, H.; Elsayed, F.A.A.; Ahmed, A.-H. The molecular mass and isoelectric point of plant proteomes. BMC Genom. 2019, 20, 631. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Prosky, L.; Asp, N.G.; Schweizer, T.F.; Devries, J.W.; Furda, I. Determination of insoluble, soluble, and total dietary fiber in foods and food products: Interlaboratory study. J. Assoc. Off. Anal. Chem. 1988, 71, 1017–1023. [Google Scholar] [CrossRef]
- Moores, R.G.; McDermott, D.L.; Wood, T.R. Determination of chlorogenic acid in coffee. Anal. Chem. 1948, 20, 620–624. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Alaiz, M.; Navarro, J.L.; Girón, J.; Vioque, E. Amino acid analysis by high-performance liquid chromatography after derivatization with diethyl ethoxymethylenemalonate. J. Chromatogr. A 1992, 591, 181–186. [Google Scholar] [CrossRef]
- Yust, M.M.; Pedroche, J.; Girón-Calle, J.; Vioque, J.; Millán, F.; Alaiz, M. Determination of tryptophan by high-performance liquid chromatography of alkaline hydrolysates with spectrophotometric detection. Food Chem. 2004, 85, 317–320. [Google Scholar] [CrossRef]
- Sindayikengera, S.; Xia, W.-S. Nutritional evaluation of caseins and whey proteins and their hydrolysates from Protamex. Zhejiang Univ. Sci. B 2006, 7, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Chen, H.M.; Shiau, C.Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reactions: Antioxidative activities of product of browning reaction prepared from glucosamine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Rodriguez-Martin, N.M.; Villanueva, A.; Pedroche, J.; Cruz-Chamorro, I.; Millan, F.; Millan-Linares, M. Evaluation of anti-inflammatory and atheroprotective properties of wheat gluten protein hydrolysates in primary human monocytes. Foods 2020, 9, 854. [Google Scholar] [CrossRef]
- FAO FINNUT. Dietary Protein Quality Evaluation in Human Nutrition; FAO of the UN: Auckland, New Zealand, 2017. [Google Scholar]
- Hertzler, S.R.; Lieblen-Boff, C.; Weiler, M.; Allgeier, C. Plant proteins: Assessing their nutritional quality and effects on health and physical function. Nutrients 2020, 12, 3704. [Google Scholar] [CrossRef]
- Joye, I. Protein digestibility of cereal products. Foods 2019, 8, 199. [Google Scholar] [CrossRef]
- Shumoy, H.; Pattyn, S.; Raes, K. Tef protein: Solubility characterization, in vitro digestibility and its suitability as a gluten free ingredient. LWT Food Sci. Technol. 2018, 89, 697–703. [Google Scholar] [CrossRef]
- Tapal, A.; Vegarud, G.E.; Sreedharab, A.; Kaul-Tico, P. Nutraceutical protein isolate from pigeon pea (Cajanus cajan) milling waste by-product: Functional aspects and digestibility. Food Funct. 2019, 10, 2710. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Principles of Biochemistry, 7th ed.; Lehninger, W.H., Ed.; Freeman & Company: New York, NY, USA, 2017. [Google Scholar]
- Colovic, M.B.; Vesna, M.V.; Dragan, M.D.; Krstict, D.Z. Sulfur-containing amino acids: Protective role against free radicals and heavy metals. Curr. Med. Chem. 2018, 25, 324–335. [Google Scholar] [CrossRef]
- Hong, G.P.; Min, S.G.; Jo, Y.J. Anti-Oxidative and anti-aging activities of porcine by-product collagen hydrolysates produced by commercial proteases: Effect of hydrolysis and ultrafiltration. Molecules 2019, 24, 1104. [Google Scholar] [CrossRef]
- Cruz-Chamorro, I.; Álvarez-Sánchez, N.; Santos-Sánchez, G.; Pedroche, J.; Fernández-Pachón, M.S.; Millán, F.; Carrillo-Vico, A. Immunomodulatory and antioxidant properties of wheat gluten protein hydrolysates in human peripheral blood mononuclear cells. Nutrients 2020, 12, 1673. [Google Scholar] [CrossRef]
- Hou, C.; Wu, L.; Wang, Z.; Saguer, E.; Zhang, D. Purification and Identification of Antioxidant Alcalase-Derived Peptides from Sheep Plasma Proteins. Antioxidants 2019, 8, 592. [Google Scholar] [CrossRef] [PubMed]
- Millán-Linares, M.C.; Bermúdez, B.; Yust, M.M.; Millán, F.; Pedroche, J. Anti-inflammatory activity of lupine (Lupinus angustifolius L.) protein hydrolysates in THP-1- derived macrophages. J. Funct. Foods 2014, 8, 224–233. [Google Scholar] [CrossRef]
- Millán-Linares, M.C.; Millán, F.; Pedroche, J.; Yust, M.M. GPETAFLR: A new anti-inflammatory peptide from Lupinus angustifolius L. protein hydrolysate. J. Funct. Foods 2015, 18, 358–367. [Google Scholar] [CrossRef]
- Rodriguez-Martin, N.M.; Toscano, R.; Villanueva, A.; Pedroche, J.; Millan, F.; Montserrat-de la Paz, S.; Millan-Linares, M.C. Neuroprotective protein hydrolysates from hemp (Cannabis sativa L.) seeds. Food Funct. 2019, 10, 6732–6739. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martin, N.M.; Montserrat-de la Paz, S.; Toscano, R.; Grao-Cruces, E.; Villanueva, A.; Pedroche, J.; Millan, F.; Millan-Linares, M.C. Hemp (Cannabis sativa L.) protein hydrolysates promote anti-inflammatory response in primary human monocytes. Biomolecules 2020, 10, 803. [Google Scholar] [CrossRef] [PubMed]
- Snigdha, G.; Kaustav, M. Structural-features of food-derived bioactive peptides with anti-inflammatory activity: A brief review. J. Nutr. Biochem. 2018, 2019, e12531. [Google Scholar] [CrossRef]
- Linlin, C.; Huidan, D.; Hengmin, C.; Jing, F.; Zhicai, Z.; Junliang, D.; Yinglun, L.; Xun, W.; Ling, Z. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Lujuan, X.; Qingquan, F.; Guanghong, Z.; Wangang, Z. A review of antioxidant peptides derived from meat muscle and by-products. Antioxidants 2016, 5, 32. [Google Scholar] [CrossRef]
- Hambraeus, L. Protein and Amino Acids in Human Nutrition. In Reference Module in Biomedical Sciences. Amaranthus: A Promising Crop of Future, 1st ed.; Elsevier: Amsterdam, The Netherland, 2014; p. 48. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).