
Ascorbate Peroxidase Neofunctionalization at the Origin of APX-R and APX-L: Evidence from Basal Archaeplastida

 ,
,  and
and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Protein Sequence Retrieval
2.2. Sequence Alignment and Phylogenetic Analysis
3. Results
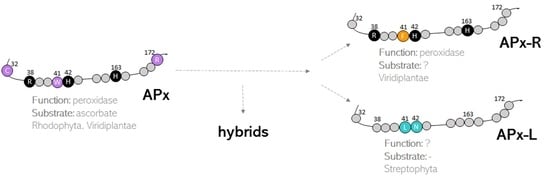
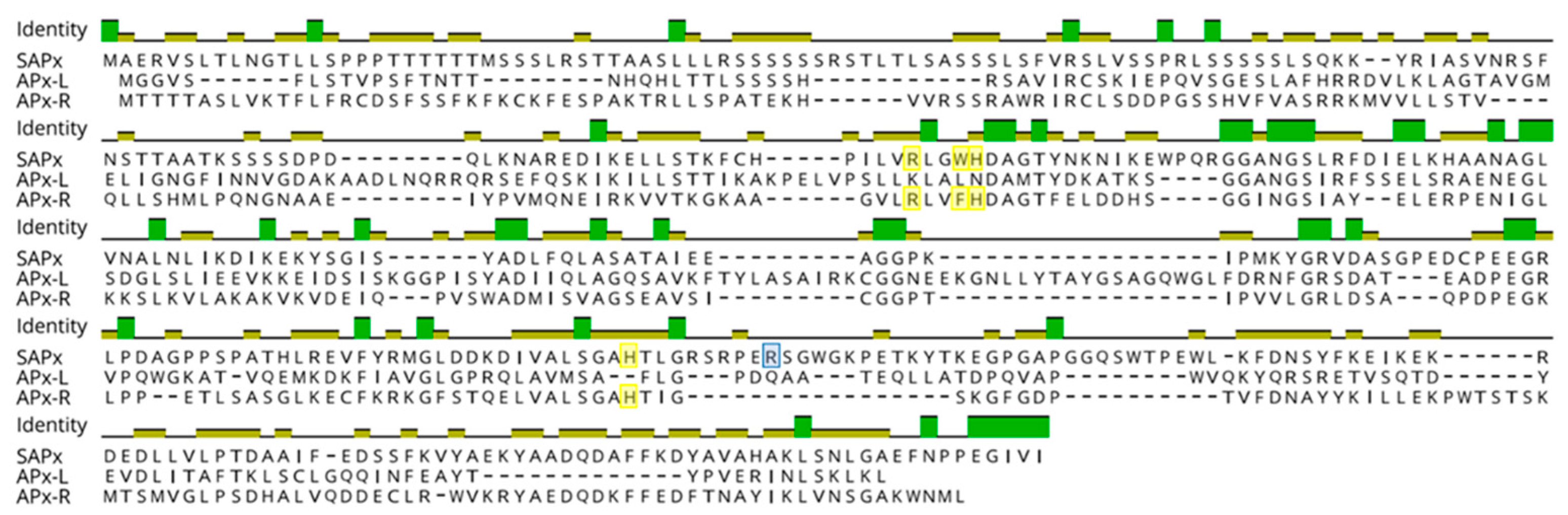
3.1. Hybrid Proteins Share Features of Distinct Families
3.2. Hybrid Proteins Are More Prevalent in Species of Green Algae
3.3. Mutations and Small Deletions Led to the Emergence of APX-R and APX-L
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Asada, K. Ascorbate Peroxidase-A Hydrogen Peroxide-Scavenging Enzyme in Plants. Physiol. Plant. 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Gest, N.; Gautier, H.; Stevens, R. Ascorbate as Seen Through Plant Evolution: The Rise of A Successful Molecule? J. Exp. Bot. 2013, 64, 33–53. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.; Hernandez, J.A.; Del Rio, L.A.; Sevilla, F. Evidence for the Presence of the Ascorbate-Glutathione Cycle in Mitochondria and Peroxisomes of Pea Leaves. Plant Physiol. 1997, 114, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, K.; Ishikawa, T.; Nakamura, Y.; Tamoi, M.; Takeda, T.; Tada, T.; Nishimura, K.; Shigeoka, S. Comparative Study on Recombinant Chloroplastic and Cytosolic Ascorbate Peroxidase Isozymes of Spinach. Arch. Bioch. Bioph. 1998, 353, 55–63. [Google Scholar] [CrossRef]
- Teixeira, F.K.; Menezes-Benavente, L.; Galvão, V.C.; Margis, R.; Margis-Pinheiro, M. Rice Ascorbate Peroxidase Gene Family Encodes Functionally Diverse Isoforms Localized in Different Subcellular Compartments. Planta 2006, 224, 300–314. [Google Scholar] [CrossRef] [PubMed]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and Function of Ascorbate Peroxidase Isoenzymes. J. Exp. Bot. 2002, 53, 1305–1319. [Google Scholar] [CrossRef] [PubMed]
- Lad, L.; Mewies, M.; Raven, E.L. Substrate Binding and Catalytic Mechanism in Ascorbate Peroxidase: Evidence for Two Ascorbate Binding Sites. Biochemistry 2002, 41, 13774–13781. [Google Scholar] [CrossRef]
- Patterson, W.R.; Poulos, T.L. Crystal Structure of Recombinant Pea Cytosolic Ascorbate Peroxidase. Biochemistry 1995, 34, 4331–4341. [Google Scholar] [CrossRef]
- Mandelman, D.; Li, H.; Poulos, T.L.; Schwarz, F.P. The Role of Quaternary Interactions on the Stability and Activity of Ascorbate Peroxidase. Protein Sci. 2009, 7, 2089–2098. [Google Scholar] [CrossRef] [PubMed]
- Raven, E.L. Understanding Functional Diversity and Substrate Specificity in Haem Peroxidases: What Can We Learn from Ascorbate Peroxidase? Nat. Prod. Rep. 2003, 20, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Sharp, K.H.; Mewies, M.; Moody, P.C.E.; Raven, E.L. Crystal Structure of the Ascorbate Peroxidase-Ascorbate Complex. Nat. Struct. Biol. 2003, 10, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, I.K.; Badyal, S.K.; Ghamsari, L.; Moody, P.C.E.; Raven, E.L. Interaction of Ascorbate Peroxidase with Substrates: A Mechanistic and Structural Analysis. Biochemistry 2006, 45, 7808–7817. [Google Scholar] [CrossRef]
- Kovacs, F.A.; Sarath, G.; Woodworth, K.; Twigg, P.; Tobias, C.M. Abolishing Activity Against Ascorbate in A Cytosolic Ascorbate Peroxidase from Switchgrass. Phytochemistry 2013, 94, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Çelik, A.; Cullis, P.M.; Sutcliffe, M.J.; Sangar, R.; Raven, E.L. Engineering the Active Site of Ascorbate Peroxidase. Eur. J. Biochem. 2001, 268, 78–85. [Google Scholar] [CrossRef]
- Battistuzzi, G.; Bellei, M.; Bortolotti, C.A.; Sola, M. Redox Properties of Heme Peroxidases. Arch. Biochem. Biophys. 2010, 500, 21–36. [Google Scholar] [CrossRef]
- Pipirou, Z.; Bottrill, A.R.; Metcalfe, C.M.; Mistry, S.C.; Badyal, S.K.; Rawlings, B.J.; Raven, E.L. Autocatalytic Formation of A Covalent Link between Tryptophan 41 and the Heme in Ascorbate Peroxidase. Biochemistry 2007, 46, 2174–2180. [Google Scholar] [CrossRef]
- Banci, L. Structural Properties of Peroxidases. J. Biotechnol. 1997, 53, 253–263. [Google Scholar] [CrossRef]
- Chary, K.V.R.; Srivastava, A.K. Encyclopedia of Biophysics; Springer: Berlin, Germany, 2013; Volume 50, ISBN 978-3-642-16711-9. [Google Scholar]
- Lad, L.; Mewies, M.; Basran, J.; Scrutton, N.S.; Raven, E.L. Role of Histidine 42 in Ascorbate Peroxidase: Kinetic Analysis of the H42A and H42E Variants. Eur. J. Biochem. 2002, 269, 3182–3192. [Google Scholar] [CrossRef]
- Meharenna, Y.T.; Oertel, P.; Bhaskar, B.; Poulos, T.L. Engineering Ascorbate Peroxidase Activity into Cytochrome c Peroxidase. Biochemistry 2008, 47, 10324–10332. [Google Scholar] [CrossRef]
- Welinder, K.G. Superfamily of Plant, Fungal and Bacterial Peroxidases. Curr. Opin. Struct. Biol. 1992, 2, 388–393. [Google Scholar] [CrossRef]
- Zámocký, M.; Hofbauer, S.; Schaffner, I.; Gasselhuber, B.; Nicolussi, A.; Soudi, M.; Pirker, K.F.; Furtmüller, P.G.; Obinger, C. Independent Evolution of Four Heme Peroxidase Superfamilies. Arch. Biochem. Biophys. 2015, 574, 108–119. [Google Scholar] [CrossRef]
- Wada, N.; Kinoshita, S.; Matsuo, M.; Amako, K.; Miyake, C.; Asada, K. Purification and Molecular Properties of Ascorbate Peroxidase from Bovine Eye. Biochem. Biophys. Res. Commun. 1998, 242, 256–261. [Google Scholar] [CrossRef]
- Adak, S.; Datta, A.K. Leishmania Major Encodes An Unusual Peroxidase that is A Close Homologue of Plant Ascorbate Peroxidase: A Novel Role of the Transmembrane Domain. Biochem. J. 2005, 390, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Passardi, F.; Bakalovic, N.; Teixeira, F.K.; Margis-Pinheiro, M.; Penel, C.; Dunand, C. Prokaryotic Origins of the Non-Animal Peroxidase Superfamily and Organelle-Mediated Transmission to Eukaryotes. Genomics 2007, 89, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Zámocký, M.; Furtmüller, P.G.; Obinger, C. Evolution of Structure and Function of Class I Peroxidases. Arch. Biochem. Biophys. 2010, 500, 45–57. [Google Scholar] [CrossRef]
- Lazzarotto, F.; Turchetto-Zolet, A.C.; Margis-Pinheiro, M. Revisiting the Non-Animal Peroxidase Superfamily. Trends. Plant. Sci. 2015, 20, 807–813. [Google Scholar] [CrossRef]
- Lazzarotto, F.; Teixeira, F.K.; Rosa, S.B.; Dunand, C.; Fernandes, C.L.; de Vasconcelos Fontenele, A.; Silveira, J.A.G.; Verli, H.; Margis, R.; Margis-Pinheiro, M. Ascorbate Peroxidase-Related (APX-R) is A New Heme-Containing Protein Functionally Associated with Ascorbate Peroxidase but Evolutionarily Divergent. New Phytol. 2011, 191, 234–250. [Google Scholar] [CrossRef]
- Chen, C.; Letnik, I.; Hacham, Y.; Dobrev, P.; Ben-Daniel, B.-H.; Vanková, R.; Amir, R.; Miller, G. ASCORBATE PEROXIDASE6 Protects Arabidopsis Desiccating and Germinating Seeds from Stress and Mediates Cross Talk between Reactive Oxygen Species, Abscisic Acid, and Auxin. Plant Physiol. 2014, 166, 370–383. [Google Scholar] [CrossRef]
- Lazzarotto, F.; Wahni, K.; Piovesana, M.; Maraschin, F.; Messens, J.; Margis-Pinheiro, M. Arabidopsis APX-R Is A Plastidial Ascorbate-Independent Peroxidase Regulated by Photomorphogenesis. Antioxidants 2021, 10, 65. [Google Scholar] [CrossRef]
- Lundberg, E.; Storm, P.; Schröder, W.P.; Funk, C. Crystal Structure of the TL29 Protein from Arabidopsis Thaliana: An APX Homolog without Peroxidase Activity. J. Struct. Biol. 2011, 176, 24–31. [Google Scholar] [CrossRef]
- Granlund, I.; Storm, P.; Funk, C.; Schröder, W.P.; García-Cerdán, J.G.; Schubert, M.; Granlund, I. The TL29 Protein is Lumen Located, Associated with PSII and Not an Ascorbate Peroxidase. Plant Cell Physiol. 2009, 50, 1898–1910. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hecker, A.G.; Hauser, B.A. The APX4 Locus Regulates Seed Vigor and Seedling Growth in Arabidopsis Thaliana. Planta 2014, 239, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Savelli, B.; Li, Q.; Webber, M.; Jemmat, A.M.; Robitaille, A.; Zámocký, M.; Mathé, C.; Dunand, C. RedoxiBase: A Database for ROS Homeostasis Regulated Proteins. Redox Biol. 2019, 26, 101247. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Drive, R.M.; Valley, M. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, T.; Wu, C.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, 1–6. [Google Scholar]
- Abascal, F.; Zardoya, R.; Posada, D. ProtTest: Selection of Best-Fit Models of Protein Evolution. Bioinformatics 2005, 21, 2104–2105. [Google Scholar] [CrossRef]
- Gilks, W.R. Markov Chain Monte Carlo. Encycl. Biostat. 2005, 4, 1–7. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; von Heijne, G. ChloroP, A Neural Network-Based Method for Predicting Chloroplast Transit Peptides and Their Cleavage Sites. Protein Sci. 1999, 8, 978–984. [Google Scholar] [CrossRef]
- Sperschneider, J.; Catanzariti, A.M.; DeBoer, K.; Petre, B.; Gardiner, D.M.; Singh, K.B.; Dodds, P.N.; Taylor, J.M. LOCALIZER: Subcellular Localization Prediction of Both Plant and Effector Proteins in the Plant Cell. Sci. Rep. 2017, 7, 1–14. [Google Scholar]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein Localization Predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Ishikawa, T.; Yoshimura, K.; Tamoi, M.; Takeda, T.; Shigeoka, S. Alternative mRNA Splicing of 3′-Terminal Exons Generates Ascorbate Peroxidase Isoenzymes in Spinach (Spinacia Oleracea) Chloroplasts. Biochem. J. 1997, 328, 795–800. [Google Scholar] [CrossRef]
- Poulos, T.L. Thirty Years of Heme Peroxidase Structural Biology. Arch Biochem. Biophys. 2011, 500, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.K.; Menezes-Benavente, L.; Margis, R.; Margis-Pinheiro, M. Analysis of the Molecular Evolutionary History of the Ascorbate Peroxidase Gene Family: Inferences from the Rice Genome. J. Mol. Evol. 2004, 59, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Begara-Morales, J.C.; Sánchez-Calvo, B.; Chaki, M.; Valderrama, R.; Mata-Pérez, C.; López-Jaramillo, J.; Padilla, M.N.; Carreras, A.; Corpas, F.J.; Barroso, J.B. Dual Regulation of Cytosolic Ascorbate Peroxidase (APX) by Tyrosine Nitration and S-Nitrosylation. J. Exp. Bot. 2014, 65, 527–538. [Google Scholar] [CrossRef]
- Turner, D.D.; Lad, L.; Kwon, H.; Basran, J.; Carr, K.H.; Moody, P.C.E.; Raven, E.L. The Role of Ala134 in Controlling Substrate Binding and Reactivity in Ascorbate Peroxidase. J. Inorg. Biochem. 2018, 180, 230–234. [Google Scholar] [CrossRef]
- Barros, J.; Escamilla-Trevino, L.; Song, L.; Rao, X.; Serrani-Yarce, J.C.; Palacios, M.D.; Engle, N.; Choudhury, F.K.; Tschaplinski, T.J.; Venables, B.J.; et al. 4-Coumarate 3-Hydroxylase in the Lignin Biosynthesis Pathway is A Cytosolic Ascorbate Peroxidase. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mcwhite, C.D.; Papoulas, O.; Drew, K.; Chen, Z.J.; Ronald, P.C.; Marcotte Correspondence, E.M. A Pan-Plant Protein Complex Map Reveals Deep Conservation and Novel Assemblies. Cell 2020, 181, 460–474. [Google Scholar] [CrossRef]
- Jasion, V.S.; Polanco, J.A.; Meharenna, Y.T.; Li, H.; Poulos, T.L. Crystal Structure of Leishmania Major Peroxidase and Characterization of the Compound I Tryptophan Radical. J. Biol. Chem. 2011, 286, 24608–24615. [Google Scholar] [CrossRef]
- Zámocký, M.; Kamlárová, A.; Maresch, D.; Chovanová, K.; Harichová, J.; Furtmüller, P.G. Hybrid Heme Peroxidases from Rice Blast Fungus Magnaporthe Oryzae Involved in Defence Against Oxidative Stress. Antioxidants 2020, 9, 655. [Google Scholar] [CrossRef]
- Chen, C.; Galon, Y.; Ishka, M.R.; Malihi, S.; Shimanovsky, V.; Twito, S.; Rath, A.; Vatamaniuk, O.K.; Miller, G. ASCORBATE PEROXIDASE6 Delays the Onset of Age-Dependent Leaf Senescence. Plant Physiol. 2021, 185, 441–456. [Google Scholar] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazzarotto, F.; Menguer, P.K.; Del-Bem, L.-E.; Zámocký, M.; Margis-Pinheiro, M. Ascorbate Peroxidase Neofunctionalization at the Origin of APX-R and APX-L: Evidence from Basal Archaeplastida. Antioxidants 2021, 10, 597. https://doi.org/10.3390/antiox10040597
Lazzarotto F, Menguer PK, Del-Bem L-E, Zámocký M, Margis-Pinheiro M. Ascorbate Peroxidase Neofunctionalization at the Origin of APX-R and APX-L: Evidence from Basal Archaeplastida. Antioxidants. 2021; 10(4):597. https://doi.org/10.3390/antiox10040597
Chicago/Turabian StyleLazzarotto, Fernanda, Paloma Koprovski Menguer, Luiz-Eduardo Del-Bem, Marcel Zámocký, and Márcia Margis-Pinheiro. 2021. "Ascorbate Peroxidase Neofunctionalization at the Origin of APX-R and APX-L: Evidence from Basal Archaeplastida" Antioxidants 10, no. 4: 597. https://doi.org/10.3390/antiox10040597
APA StyleLazzarotto, F., Menguer, P. K., Del-Bem, L.-E., Zámocký, M., & Margis-Pinheiro, M. (2021). Ascorbate Peroxidase Neofunctionalization at the Origin of APX-R and APX-L: Evidence from Basal Archaeplastida. Antioxidants, 10(4), 597. https://doi.org/10.3390/antiox10040597