
Potentials of Raspberry Ketone as a Natural Antioxidant


Abstract
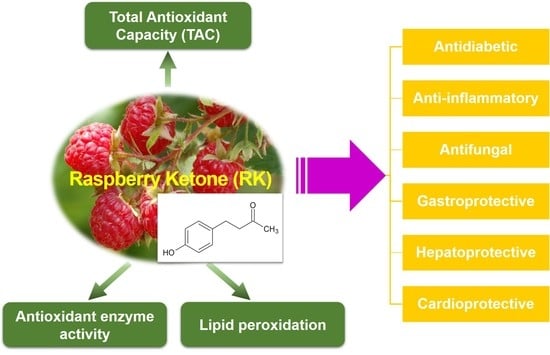
1. Introduction
2. Methodology
3. RK’s Antioxidant Activities and Their Underlying Mechanisms
3.1. Total Antioxidant Capacity (TAC)
3.2. Antioxidant Enzyme Activity
3.3. Lipid Peroxidation
4. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative Stress, Inflammation, and Cancer: How Are They Linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Persson, T.; Popescu, B.O.; Cedazo-Minguez, A. Oxidative Stress in Alzheimer’s Disease: Why Did Antioxidant Therapy Fail? Oxid. Med. Cell. Longev. 2014, 2014, 427318. [Google Scholar] [CrossRef]
- Rackova, L.; Oblozinsky, M.; Kostalova, D.; Kettmann, V.; Bezakova, L. Free Radical Scavenging Activity and Lipoxygenase Inhibition of Mahonia Aquifolium Extract and Isoquinoline Alkaloids. J. Inflamm. 2007, 4, 15. [Google Scholar] [CrossRef]
- Chanda, S.; Dave, R. In Vitro Models for Antioxidant Activity Evaluation and Some Medicinal Plants Possessing Antioxidant Properties: An Overview. Afr. J. Microbiol. Res. 2009, 3, 981–996. [Google Scholar]
- Pisoschi, A.M.; Negulescu, G.P. Methods for Total Antioxidant Activity Determination: A Review. Biochem. Anal. Biochem. 2011, 1, 106. [Google Scholar] [CrossRef]
- Rahman, K. Studies on Free Radicals, Antioxidants, and Co-Factors. Clin. Interv. Aging 2007, 2, 219–236. [Google Scholar] [PubMed]
- Kinnula, V.L.; Crapo, J.D.; Raivio, K.O. Generation and Disposal of Reactive Oxygen Metabolites in the Lung. Lab. Investig. J. Tech. Methods Pathol. 1995, 73, 3–19. [Google Scholar]
- Kinnula, V.L.; Crapo, J.D. Superoxide Dismutases in the Lung and Human Lung Diseases. Am. J. Respir. Crit. Care Med. 2003, 167, 1600–1619. [Google Scholar] [CrossRef]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide Dismutase Multigene Family: A Comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) Gene Structures, Evolution, and Expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Kinnula, V. Production and Degradation of Oxygen Metabolites During Inflammatory States in the Human Lung. Curr. Drug Target Inflamm. Allergy 2005, 4, 465–470. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Kirkman, H.N.; Rolfo, M.; Ferraris, A.M.; Gaetani, G.F. Mechanisms of Protection of Catalase by Nadph Kinetics and Stoichiometry. J. Biol. Chem. 1999, 274, 13908–13914. [Google Scholar] [CrossRef]
- Vetrano, A.M.; Heck, D.E.; Mariano, T.M.; Mishin, V.; Laskin, D.L.; Laskin, J.D. Characterization of the Oxidase Activity in Mammalian Catalase. J. Biol. Chem. 2005, 280, 35372–35381. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative Stress, Aging, and Diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Szabo, I.L.; Kenyeres, A.; Szegedi, A.; Szollosi, A.G. Heme Oxygenase and the Skin in Health and Disease. Curr. Pharm. Des. 2018, 24, 2303–2310. [Google Scholar] [CrossRef] [PubMed]
- Poljsak, B.; Šuput, D.; Milisav, I. Achieving the Balance between ROS and Antioxidants: When to Use the Synthetic Antioxidants. Oxid. Med. Cell. Longev. 2013, 2013, 956792. [Google Scholar] [CrossRef]
- Simona, C.L.; Sandra, A.V.; Mirela, D.; Andreia, T.; Gabriel Lucian, R. Biosensors Applications on Assessment of Reactive Oxygen Species and Antioxidants. J. Environ. Biosens. 2011, 9, 12–21. [Google Scholar]
- Lindsay, D.G.; Astley, S.B. European Research on the Functional Effects of Dietary Antioxidants—EUROFEDA. Mol. Aspects Med. 2002, 23, 1–38. [Google Scholar] [CrossRef]
- White, E.; Shannon, J.S.; Patterson, R.E. Relationship between Vitamin and Calcium Supplement Use and Colon Cancer. Cancer Epidemiol. Biomark. 1997, 6, 769–774. [Google Scholar]
- Descamps-Latscha, B.; Drüeke, T.; Witko-Sarsat, V. Dialysis-Induced Oxidative Stress: Biological Aspects, Clinical Consequences, and Therapy. Semin. Dial. 2001, 14, 193–199. [Google Scholar] [CrossRef]
- Sung, C.-C.; Hsu, Y.-C.; Chen, C.-C.; Lin, Y.-F.; Wu, C.-C. Oxidative Stress and Nucleic Acid Oxidation in Patients with Chronic Kidney Disease. Oxid. Med. Cell. Longev. 2013, 2013, 301982. [Google Scholar] [CrossRef]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic Acid: Chemistry, Biology and the Treatment of Cancer. Biochim. Biophys. Acta 2012, 1826, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Morris, H.; Cronin, M.T.D. Metals, Toxicity and Oxidative Stress. Curr. Med. Chem. 2005, 12, 1161–1208. [Google Scholar] [CrossRef]
- Carr, A.; Frei, B. Does Vitamin C Act as a Pro-Oxidant under Physiological Conditions? FASEB J. 1999, 13, 1007–1024. [Google Scholar] [CrossRef]
- El-Agamey, A.; Lowe, G.M.; McGarvey, D.J.; Mortensen, A.; Phillip, D.M.; Truscott, T.G.; Young, A.J. Carotenoid Radical Chemistry and Antioxidant/pro-Oxidant Properties. Arch. Biochem. Biophys. 2004, 430, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Sampson, J.; Bramley, P.M.; Holloway, D.E. Why Do We Expect Carotenoids to Be Antioxidants in Vivo? Free Radic. Res. 1997, 26, 381–398. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M. The Role of Antioxidants in Disease Prevention. Medicine 2006, 34, 533–535. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The Role of Antioxidants in the Chemistry of Oxidative Stress: A Review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Poljsak, B.; Milisav, I. The Neglected Significance of “Antioxidative Stress”. Oxid. Med. Cell. Longev. 2012, 2012, 480895. [Google Scholar] [CrossRef] [PubMed]
- Cutler, R.G.; Mattson, M.P. Measuring Oxidative Stress and Interpreting Its Clinical Relevance for Humans. In Critical Reviews of Oxidative Stress and Aging; World Scientific: Singapore, 2002; pp. 131–164. ISBN 978-981-02-4636-5. [Google Scholar]
- Cutler, R.G. Genetic Stability, Dysdifferentiation, and Longevity Determinant Genes. In Critical Reviews of Oxidative Stress and Aging; World Scientific: Singapore, 2002; pp. 1146–1235. ISBN 978-981-02-4636-5. [Google Scholar]
- Gallois, A. Quantitative evaluation of raspberry ketone using thin-layer chromatography. Sci. Aliments 1982, 2, 106. [Google Scholar]
- Ulbricht, C.; Catapang, M.; Conquer, J.; Costa, D.; Culwell, S.; D’Auria, D.; Isaac, R.; Le, C.; Marini, E.; Miller, A.; et al. Raspberry Ketone: An Evidence-Based Systematic Review by the Natural Standard Research Collaboration. Altern. Complement. Ther. 2013, 19, 98–100. [Google Scholar] [CrossRef]
- Honkanen, E.; Pyysalo, T.; Hirvi, T. The Aroma of Finnish Wild Raspberries, Rubus Idaeus, L. Z. Lebensm. Unters. Forsch. 1980, 171, 180–182. [Google Scholar] [CrossRef]
- Larsen, M.; Poll, L. Odour Thresholds of Some Important Aroma Compounds in Raspberries. Z. Lebensm. Unters. Forsch. 1990, 191, 129–131. [Google Scholar] [CrossRef]
- Larsen, M.; Poll, L.; Callesen, O.; Lewis, M. Relations between the Content of Aroma Compounds and the Sensory Evaluation of 10 Raspberry Varieties (Rubus Idaeus L.). Acta Agric. Scand. 1991, 41, 447–454. [Google Scholar] [CrossRef]
- Borejsza-Wysocki, W.; Goers, S.K.; McArdle, R.N.; Hrazdina, G. (P-Hydroxyphenyl)Butan-2-One Levels in Raspberries Determined by Chromatographic and Organoleptic Methods. J. Agric. Food Chem. 1992, 40, 1176–1177. [Google Scholar] [CrossRef]
- Paterson, A.; Kassim, A.; McCallum, S.; Woodhead, M.; Smith, K.; Zait, D.; Graham, J. Environmental and Seasonal Influences on Red Raspberry Flavour Volatiles and Identification of Quantitative Trait Loci (QTL) and Candidate Genes. Theor. Appl. Genet. 2013, 126, 33–48. [Google Scholar] [CrossRef]
- Fronza, G.; Fuganti, C.; Guillou, C.; Reniero, F.; Joulain, D. Natural Abundance 2H Nuclear Magnetic Resonance Study of the Origin of Raspberry Ketone. J. Agric. Food Chem. 1998, 46, 248–254. [Google Scholar] [CrossRef]
- Fronza, G.; Fuganti, C.; Pedrocchi-Fantoni, G.; Serra, S.; Zucchi, G.; Fauhl, C.; Guillou, C.; Reniero, F. Stable Isotope Characterization of Raspberry Ketone Extracted from Taxus Baccata and Obtained by Oxidation of the Accompanying Alcohol (Betuligenol). J. Agric. Food Chem. 1999, 47, 1150–1155. [Google Scholar] [CrossRef]
- Tan, K.-H.; Nishida, R. Synomone Or Kairomone? Bulbophyllum apertum Flower Releases Raspberry Ketone To Attract Bactrocera Fruit Flies. J. Chem. Ecol. 2005, 31, 497–507. [Google Scholar] [CrossRef]
- Akiyama, M.; Murakami, K.; Ikeda, M.; Iwatsuki, K.; Wada, A.; Tokuno, K.; Onishi, M.; Iwabuchi, H. Analysis of the Headspace Volatiles of Freshly Brewed Arabica Coffee Using Solid-Phase Microextraction. J. Food Sci. 2007, 72, C388–C396. [Google Scholar] [CrossRef]
- Akiyama, M.; Murakami, K.; Hirano, Y.; Ikeda, M.; Iwatsuki, K.; Wada, A.; Tokuno, K.; Onishi, M.; Iwabuchi, H. Characterization of Headspace Aroma Compounds of Freshly Brewed Arabica Coffees and Studies on a Characteristic Aroma Compound of Ethiopian Coffee. J. Food Sci. 2008, 73, C335–C346. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.V.; Quek, S.-Y.; Stevenson, R.J.; Winz, R.A. Characterization of the Bound Volatile Extract from Baby Kiwi (Actinidia arguta). J. Agric. Food Chem. 2011, 59, 8358–8365. [Google Scholar] [CrossRef] [PubMed]
- Hugueny, P.; Dumont, B.; Ropert, F.; Belin, J.M. The Raspberry Ketone, a Biotechnological Way for Production. Bioflavor 1995, 95, 14–17. [Google Scholar]
- Morimoto, C.; Satoh, Y.; Hara, M.; Inoue, S.; Tsujita, T.; Okuda, H. Anti-Obese Action of Raspberry Ketone. Life Sci. 2005, 77, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S. Raspberry Ketone Increases Both Lipolysis and Fatty Acid Oxidation in 3T3-L1 Adipocytes. Planta Med. 2010, 76, 1654–1658. [Google Scholar] [CrossRef]
- Park, K.S. Raspberry Ketone, a Naturally Occurring Phenolic Compound, Inhibits Adipogenic and Lipogenic Gene Expression in 3T3-L1 Adipocytes. Pharm. Biol. 2015, 53, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Leu, S.-Y.; Chen, Y.-C.; Tsai, Y.-C.; Hung, Y.-W.; Hsu, C.-H.; Lee, Y.-M.; Cheng, P.-Y. Raspberry Ketone Reduced Lipid Accumulation in 3T3-L1 Cells and Ovariectomy-Induced Obesity in Wistar Rats by Regulating Autophagy Mechanisms. J. Agric. Food Chem. 2017, 65, 10907–10914. [Google Scholar] [CrossRef]
- Xiong, S.-L.; Yue, L.-M.; Lim, G.T.; Yang, J.-M.; Lee, J.; Park, Y.-D. Inhibitory Effect of Raspberry Ketone on α-Glucosidase: Docking Simulation Integrating Inhibition Kinetics. Int. J. Biol. Macromol. 2018, 113, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.B.; Jeong, H.J. Rheosmin, a Naturally Occurring Phenolic Compound Inhibits LPS-Induced INOS and COX-2 Expression in RAW264.7 Cells by Blocking NF-ΚB Activation Pathway. Food Chem. Toxicol. 2010, 48, 2148–2153. [Google Scholar] [CrossRef]
- Opdyke, D.L.J. 4-(p-Hydroxyphenyl)-2-Butanone. Food Cosmet. Toxicol. 1978, 16, 781–782. [Google Scholar] [CrossRef]
- Vaughn, S.F.; Spencer, G.F.; Shasha, B.S. Volatile Compounds from Raspberry and Strawberry Fruit Inhibit Postharvest Decay Fungi. J. Food Sci. 1993, 58, 793–796. [Google Scholar] [CrossRef]
- Badr, A.M.; EL-Orabi, N.F.; Ali, R.A. The Implication of the Crosstalk of Nrf2 with NOXs, and HMGB1 in Ethanol-Induced Gastric Ulcer: Potential Protective Effect Is Afforded by Raspberry Ketone. PLoS ONE 2019, 14, e0220548. [Google Scholar] [CrossRef]
- Lin, C.-H.V.; Ding, H.-Y.; Kuo, S.-Y.; Chin, L.-W.; Wu, J.-Y.; Chang, T.-S. Evaluation of in Vitro and in Vivo Depigmenting Activity of Raspberry Ketone from Rheum Officinale. Int. J. Mol. Sci. 2011, 12, 4819–4835. [Google Scholar] [CrossRef]
- Wang, L.; Meng, X.; Zhang, F. Raspberry Ketone Protects Rats Fed High-Fat Diets Against Nonalcoholic Steatohepatitis. J. Med. Food 2012, 15, 495–503. [Google Scholar] [CrossRef]
- Fouad, D.; Badr, A.; Attia, H.A. Hepatoprotective Activity of Raspberry Ketone Is Mediated via Inhibition of the NF-ΚB/TNF-α/Caspase Axis and Mitochondrial Apoptosis in Chemically Induced Acute Liver Injury. Toxicol. Res. 2019, 8, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Akamatsu, M.; Hotta, Y.; Hosoda, A.; Tamura, H. Effect of Essential Oils, Such as Raspberry Ketone and Its Derivatives, on Antiandrogenic Activity Based on in Vitro Reporter Gene Assay. Bioorg. Med. Chem. Lett. 2010, 20, 2111–2114. [Google Scholar] [CrossRef] [PubMed]
- Khan, V.; Sharma, S.; Bhandari, U.; Sharma, N.; Rishi, V.; Haque, S.E. Suppression of Isoproterenol-Induced Cardiotoxicity in Rats by Raspberry Ketone via Activation of Peroxisome Proliferator Activated Receptor-α. Eur. J. Pharmacol. 2019, 842, 157–166. [Google Scholar] [CrossRef]
- Harada, N.; Okajima, K.; Narimatsu, N.; Kurihara, H.; Nakagata, N. Effect of Topical Application of Raspberry Ketone on Dermal Production of Insulin-like Growth Factor-I in Mice and on Hair Growth and Skin Elasticity in Humans. Growth Horm. IGF Res. 2008, 18, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.E.; Abo-ELmatty, D.M.; Mesbah, N.M.; Saleh, S.M.; Ali, A.-M.A.; Sakr, A.T. Raspberry Ketone Preserved Cholinergic Activity and Antioxidant Defense in Obesity Induced Alzheimer Disease in Rats. Biomed. Pharmacother. 2018, 107, 1166–1174. [Google Scholar] [CrossRef]
- Khan, V.; Sharma, S.; Bhandari, U.; Ali, S.M.; Haque, S.E. Raspberry Ketone Protects against Isoproterenol-Induced Myocardial Infarction in Rats. Life Sci. 2018, 194, 205–212. [Google Scholar] [CrossRef]
- Hamdy, S.M.; El-Khayat, Z.; Farrag, A.R.; Sayed, O.N.; El-Sayed, M.M.; Massoud, D. Hepatoprotective Effect of Raspberry Ketone and White Tea against Acrylamide-Induced Toxicity in Rats. Drug Chem. Toxicol. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.T.; Zaitone, S.A.; Ahmed, A.; Mehanna, E.T.; El-Sayed, N.M. Raspberry Ketones Attenuate Cyclophosphamide-Induced Pulmonary Toxicity in Mice through Inhibition of Oxidative Stress and NF-ΚB Pathway. Antioxidants 2020, 9, 1168. [Google Scholar] [CrossRef] [PubMed]
- Mehanna, E.T.; Barakat, B.M.; ElSayed, M.H.; Tawfik, M.K. An Optimized Dose of Raspberry Ketones Controls Hyperlipidemia and Insulin Resistance in Male Obese Rats: Effect on Adipose Tissue Expression of Adipocytokines and Aquaporin. Eur. J. Pharmacol. 2018, 832, 81–89. [Google Scholar] [CrossRef]
- Attia, R.T.; Abdel-Mottaleb, Y.; Abdallah, D.M.; El-Abhar, H.S.; El-Maraghy, N.N. Raspberry Ketone and Garcinia Cambogia Rebalanced Disrupted Insulin Resistance and Leptin Signaling in Rats Fed High Fat Fructose Diet. Biomed. Pharmacother. 2019, 110, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Oteiza, P.I.; Galleano, M. In Vitro Measurements and Interpretation of Total Antioxidant Capacity. Biochim. Biophys. Acta BBA Gen. Subj. 2014, 1840, 931–934. [Google Scholar] [CrossRef] [PubMed]
- Woodford, F.P.; Whitehead, T.P. Is Measuring Serum Antioxidant Capacity Clinically Useful? Ann. Clin. Biochem. 1998, 35, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Bartosz, G. Non-Enzymatic Antioxidant Capacity Assays: Limitations of Use in Biomedicine. Free Radic. Res. 2010, 44, 711–720. [Google Scholar] [CrossRef]
- Gutteridge, J.M.C.; Maidt, L.; Poyer, L. Superoxide Dismutase and Fenton Chemistry. Reaction of Ferric-EDTA Complex and Ferric-Bipyridyl Complex with Hydrogen Peroxide without the Apparent Formation of Iron(II). Biochem. J. 1990, 269, 169–174. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Comparison of Superoxide with Other Reducing Agents in the Biological Production of Hydroxyl Radicals. Biochem. J. 1979, 182, 625–628. [Google Scholar] [CrossRef]
- Yamazaki, I.; Piette, L.H. ESR Spin-Trapping Studies on the Reaction of Fe2+ Ions with H2O2-Reactive Species in Oxygen Toxicity in Biology. J. Biol. Chem. 1990, 265, 13589–13594. [Google Scholar] [CrossRef]
- Koracevic, D. Method for the Measurement of Antioxidant Activity in Human Fluids. J. Clin. Pathol. 2001, 54, 356–361. [Google Scholar] [CrossRef]
- Li, H.; Xie, Y.-H.; Yang, Q.; Wang, S.-W.; Zhang, B.-L.; Wang, J.-B.; Cao, W.; Bi, L.-L.; Sun, J.-Y.; Miao, S.; et al. Cardioprotective Effect of Paeonol and Danshensu Combination on Isoproterenol-Induced Myocardial Injury in Rats. PLoS ONE 2012, 7, e48872. [Google Scholar] [CrossRef]
- Hu, A.; Jiao, X.; Gao, E.; Koch, W.J.; Sharifi-Azad, S.; Grunwald, Z.; Ma, X.L.; Sun, J.-Z. Chronic β-Adrenergic Receptor Stimulation Induces Cardiac Apoptosis and Aggravates Myocardial Ischemia/Reperfusion Injury by Provoking Inducible Nitric-Oxide Synthase-Mediated Nitrative Stress. J. Pharmacol. Exp. Ther. 2006, 318, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, L.; Xu, X.; Huang, S.; Lu, B.; Ji, L.; Wang, Z. Therapeutic Detoxification of Quercetin against Carbon Tetrachloride-Induced Acute Liver Injury in Mice and Its Mechanism. J. Zhejiang Univ. Sci. B 2014, 15, 1039–1047. [Google Scholar] [CrossRef]
- Yu, H.; Deng, W.; Zhang, D.; Gao, Y.; Yang, Z.; Shi, X.; Sun, J.; Zhou, J.; Ji, H. Antioxidant Defenses of Onychostoma Macrolepis in Response to Thermal Stress: Insight from MRNA Expression and Activity of Superoxide Dismutase and Catalase. Fish Shellfish Immunol. 2017, 66, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. Inhibition of Copper-Zinc Superoxide Dismutase Activity by Selected Environmental Xenobiotics. Environ. Toxicol. Pharmacol. 2018, 58, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Fridovich, I. Biological Effects of the Superoxide Radical. Arch. Biochem. Biophys. 1986, 247, 1–11. [Google Scholar] [CrossRef]
- Mruk, D.D.; Silvestrini, B.; Mo, M.; Cheng, C.Y. Antioxidant Superoxide Dismutase—A Review: Its Function, Regulation in the Testis, and Role in Male Fertility. Contraception 2002, 65, 305–311. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the Superoxide Anion Radical in the Autoxidation of Pyrogallol and a Convenient Assay for Superoxide Dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Claiborne, A.L. Catalase Activity. In CRC Handbook of Methods for Oxygen Radical Research; CRC Press: Boca Raton, FL, USA, 1986; pp. 283–284. [Google Scholar]
- Schrader, M.; Fahimi, H.D. Peroxisomes and Oxidative Stress. Biochim. Biophys. Acta BBA Mol. Cell Res. 2006, 1763, 1755–1766. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an Antioxidant and Disulphide Breaking Agent: The Reasons Why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Mehta, A.; Singh, S.; Ganguly, N.K. Impairment of Intestinal Mucosal Antioxidant Defense System during Salmonella Typhimurium Infection. Dig. Dis. Sci. 1998, 43, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.W.; Jang, J.Y.; Kim, Y.H.; Kim, J.-W.; Shim, J.-J. The Effects of Broccoli Sprout Extract Containing Sulforaphane on Lipid Peroxidation and Helicobacter Pylori Infection in the Gastric Mucosa. Gut Liver 2015, 9, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Deponte, M. Glutathione Catalysis and the Reaction Mechanisms of Glutathione-Dependent Enzymes. Biochim. Biophys. Acta 2013, 1830, 3217–3266. [Google Scholar] [CrossRef] [PubMed]
- Paglia, D.E.; Valentine, W.N. Studies on the Quantitative and Qualitative Characterization of Erythrocyte Glutathione Peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Yang, G.Q.; Chen, J.S.; Wen, Z.M.; Ge, K.Y.; Zhu, L.Z.; Chen, X.C.; Chen, X.S. The Role of Selenium in Keshan Disease. Adv. Nutr. Res. 1984, 6, 203–231. [Google Scholar] [CrossRef]
- Zaitone, S.A.; Barakat, B.M.; Bilasy, S.E.; Fawzy, M.S.; Abdelaziz, E.Z.; Farag, N.E. Protective Effect of Boswellic Acids versus Pioglitazone in a Rat Model of Diet-Induced Non-Alcoholic Fatty Liver Disease: Influence on Insulin Resistance and Energy Expenditure. Naunyn. Schmiedebergs Arch. Pharmacol. 2015, 388, 587–600. [Google Scholar] [CrossRef]
- Noeman, S.A.; Hamooda, H.E.; Baalash, A.A. Biochemical Study of Oxidative Stress Markers in the Liver, Kidney and Heart of High Fat Diet Induced Obesity in Rats. Diabetol. Metab. Syndr. 2011, 3, 17. [Google Scholar] [CrossRef]
- Hsiao, P.-J.; Hsieh, T.-J.; Kuo, K.-K.; Hung, W.-W.; Tsai, K.-B.; Yang, C.-H.; Yu, M.-L.; Shin, S.-J. Pioglitazone Retrieves Hepatic Antioxidant DNA Repair in a Mice Model of High Fat Diet. BMC Mol. Biol. 2008, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/Small Maf Heterodimer Mediates the Induction of Phase II Detoxifying Enzyme Genes through Antioxidant Response Elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Gómez, O.; García-Rodríguez, R.V.; Quevedo-Corona, L.; Pérez-Pastén-Borja, R.; Rivero-Ramírez, N.L.; Ríos-Castro, E.; Pérez-Gutiérrez, S.; Pérez-Ramos, J.; Chamorro-Cevallos, G.A. Amelioration of Ethanol-Induced Gastric Ulcers in Rats Pretreated with Phycobiliproteins of Arthrospira (Spirulina) Maxima. Nutrients 2018, 10, 763. [Google Scholar] [CrossRef]
- Bredsdorff, L.; Wedebye, E.B.; Nikolov, N.G.; Hallas-Møller, T.; Pilegaard, K. Raspberry ketone in food supplements—High intake, few toxicity data—A cause for safety concern? Regul. Toxicol. Pharmacol. 2015, 73, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Kshatriya, D.; Li, X.; Badrinath, A.; Szmacinski, Z.; Goedken, M.J.; Polunas, M.; Bello, N.T. Acute feeding suppression and toxicity of raspberry ketone [4-(4-hydroxyphenyl)-2-butanone] in mice. Food Chem. Toxiccol. 2020, 143, 111512. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Mechanism(s) | Dose | Study Model | Study Result(s) | Ref. | |
|---|---|---|---|---|---|
| TAC↑ | 25, 50, 100 and 200 mg/kg | Male Wistar rats induced hepatotoxicity by CCl4 | Dose-dependent amelioration of TAC by RK treatment, which was decreased by half in the toxic group | [57] | |
| 50, 100 and 200 mg/kg | Wistar albino rats induced cardiotoxicity with ISO | Dose-dependent upregulation of TAC (3.914 ± 0.65 to 9.509 ± 0.84 µmol/L) by RK treatment compared with that in the ISO-treated group (2.598 ± 0.77 µmol/L) | [59] | ||
| 44 mg/kg | High-fat diet-fed male Wister albino rats | Increased TAC (0.7 ± 0.05 mg/dL) by RK treatment compared with that in the obese group (0.423 ± 0.03 mg/dL) | [61] | ||
| Antioxidant Enzyme Activity | SOD↑ | 0.5%, 1% or 2% | High-fat diet-fed male female Sprague-Dawley rats | RK high-dose group normalized SOD activity, which was reduced by about 33% with a high-fat diet | [56] |
| 25, 50, 100 and 200 mg/kg | Male Wistar rats induced hepatotoxicity by CCl4 | RK at 200 mg/kg normalized the SOD activity, which was dropped by about 50% due to CCl4 | [57] | ||
| 50, 100 and 200 mg/kg | Wistar albino rats induced myocardial by ISO | Significant increases in SOD level in medium- (100 mg/kg) and high-dose (200 mg/kg) of RK groups (58.39 ± 1.28 and 59.52 ± 2.3 U/mg protein, respectively) compared with that in the ISO-treated group (28.77 ± 1.4 U/mg protein) | [62] | ||
| 6 mg/kg | Male albino rats induced toxicity by AA | RK co-treatment improved the SOD level (128.26 ± 6.34 U/mL) compared with that in AA-treated group (75.65 ± 5.87 U/mL) | [63] | ||
| 25, 50, 100 and 200 mg/kg | Adult male Swiss albino rats induced pulmonary toxicity by CP | Dose-dependent recovery of SOD level (55.32 ± 2.42, 56.28 ± 2.30, 68.36 ± 3.89, and 74.59 ± 2.15 U/mg protein, respectively) by RK pre-treatment against CP-induced toxicity (40.51 U/mg protein) | [64] | ||
| CAT↑ | 50 mg/kg | Adult male Wistar rats induced gastric lesion by EtOH | CAT was abated with EtOH treatment (3.7 ± 0.07 U/g tissue); RK treatment reversed the CAT level (6.2 ± 0.28 U/g tissue) | [54] | |
| 50, 100 and 200 mg/kg | Wistar albino rats induced myocardial by ISO | Significant increases in CAT level in medium- and high-dose of RK groups (37.44 ± 2.92 and 37.95 ± 2.35 nmol H2O2/min/mg protein, respectively) compared with that in the ISO-treated group (14.75 ± 1.98 nmol H2O2/min/mg protein) | [62] | ||
| 6 mg/kg | Adult male albino rats induced toxicity by AA | RK co-treatment improved the CAT level (4.13 ± 0.45 U/mL) compared with that in AA-treated group (2.57 ± 0.64 U/mL) | [63] | ||
| 25, 50, 100 and 200 mg/kg | Adult male Swiss albino rats induced pulmonary toxicity by CP | Dose-dependent recovery of CAT level (84.31 ± 4.75, 89.31 ± 1.70, 116.32 ± 12.25, and 128.59 ± 9.30 U/mg protein, respectively) by RK pre-treatment against CP-induced toxicity (59.86 ± 4.72 U/mg protein) | [64] | ||
| GSH-Px↑ | 50 mg/kg | Adult male Wistar rats induced gastric lesion by EtOH | Significant amelioration of GSH-Px activity (262 ± 15.7 U/g tissue) compared with that in the EtOH group (86.8 ± 5.6 U/g tissue) | [54] | |
| Lipid peroxidation | GSH↑ | 50 mg/kg | Adult male Wistar rats induced gastric lesion by EtOH | Significant decrease in GSH content in the EtOH-treated group (2.8 ± 0.18 µmol/g tissue) compared with that in the control group (7.1 ± 0.43 µmol/g tissue); RK normalized the GSH content (7.8 ± 0.21 µmol/g tissue) | [54] |
| 25, 50, 100 and 200 mg/kg | Male Wistar rats induced hepatotoxicity by CCl4 | Dose-dependent recovery in GSH levels by RK treatment, which was reduced by 40% due to CCl4 | [57] | ||
| 50, 100 and 200 mg/kg | Wistar albino rats induced myocardial by ISO | Significant elevations in GSH level in medium- and high-dose of RK groups (3.88 ± 0.08 and 4.07 ± 0.18 µmol GSH/mg protein, respectively) compared with that in the ISO-treated group (0.97 ± 0.15 µmol GSH/mg protein) | [62] | ||
| 250 and 500 mg/kg | High-fat diet-fed Male Wistar albino rats | RK treatment 2-fold increased the content of GSH, compared with that in the obese group | [65] | ||
| 55 mg/kg | High-fat diet-fed adult male Wistar rats | Normalized GSH content in the RK-treated group, compared with that in the high-fat diet group | [66] | ||
| MDA↓ | 0.5%, 1% or 2% | High-fat diet-fed male female Sprague-Dawley rats | MDA level in RK high-dose group (36.2798 ± 6.454 nmol/mg of protein) was significantly lower than that in the high-fat diet group (47.9707 ± 3.187 nmol/mg of protein) | [56] | |
| 44 mg/kg | High-fat diet-fed male Wistar albino rats | Decreased MDA level in RK-treated group (29 ± 4.2 m/dL compared with that in the obese group (48.1 ± 5.2 mg/dL) | [61] | ||
| 50, 100 and 200 mg/kg | Wistar albino rats induced myocardial by ISO | Significant reductions in MDA level in medium- and high-dose of RK groups (5.84 ± 0.33 and 5.66 ± 0.34 nmol MDA/mg protein, respectively) compared with that in the ISO-treated group (9.40 ± 0.45 nmol MDA/ mg protein) | [62] | ||
| 6 mg/kg | Male albino rats induced toxicity by AA | RK co-treatment ameliorated the MDA level (13.18 ± 2.618 nmol/mL) compared with that in AA-treated group (20.83 ± 1.81 nmol/mL) | [63] | ||
| 25, 50, 100 and 200 mg/kg | Adult male Swiss albino rats induced pulmonary toxicity by CP | Dose-dependent recovery of MDA level (537.04 ± 13.73, 456.79 ± 5.12, 424.69 ± 6.67 and 374.07 ± 5.32 U/mg protein, respectively) by RK pre-treatment against CP-induced toxicity (730.25 ± 16.07 nmol/mg protein) | [64] | ||
| 250 and 500 mg/kg | High-fat diet-fed Male Wistar albino rats | RK treatment reduced MDA by approximately 50% compared with that in the obese group | [65] | ||
| 55 mg/kg | High-fat diet-fed adult male Wistar rats | RK treatment group normalized the MDA level | [66] | ||
| Nrf-2 ↑ | 50 mg/kg | Adult male Wistar rats induced gastric lesion by EtOH | Nrf-2 expression in the RK-treated group was increased by 50% compared with that in the EtOH-treated group | [54] | |
| NOXs ↓ | 50 mg/kg | Adult male Wistar rats induced gastric lesion by EtOH | NOX-1 and NOX-4 expressions in the RK-treated group were abated by about 50% compared with that in the EtOH-treated group | [54] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, S.H.; Choi, C.-I. Potentials of Raspberry Ketone as a Natural Antioxidant. Antioxidants 2021, 10, 482. https://doi.org/10.3390/antiox10030482
Lim SH, Choi C-I. Potentials of Raspberry Ketone as a Natural Antioxidant. Antioxidants. 2021; 10(3):482. https://doi.org/10.3390/antiox10030482
Chicago/Turabian StyleLim, Sung Ho, and Chang-Ik Choi. 2021. "Potentials of Raspberry Ketone as a Natural Antioxidant" Antioxidants 10, no. 3: 482. https://doi.org/10.3390/antiox10030482
APA StyleLim, S. H., & Choi, C.-I. (2021). Potentials of Raspberry Ketone as a Natural Antioxidant. Antioxidants, 10(3), 482. https://doi.org/10.3390/antiox10030482