
Lemon Balm and Corn Silk Extracts Mitigate High-Fat Diet-Induced Obesity in Mice

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
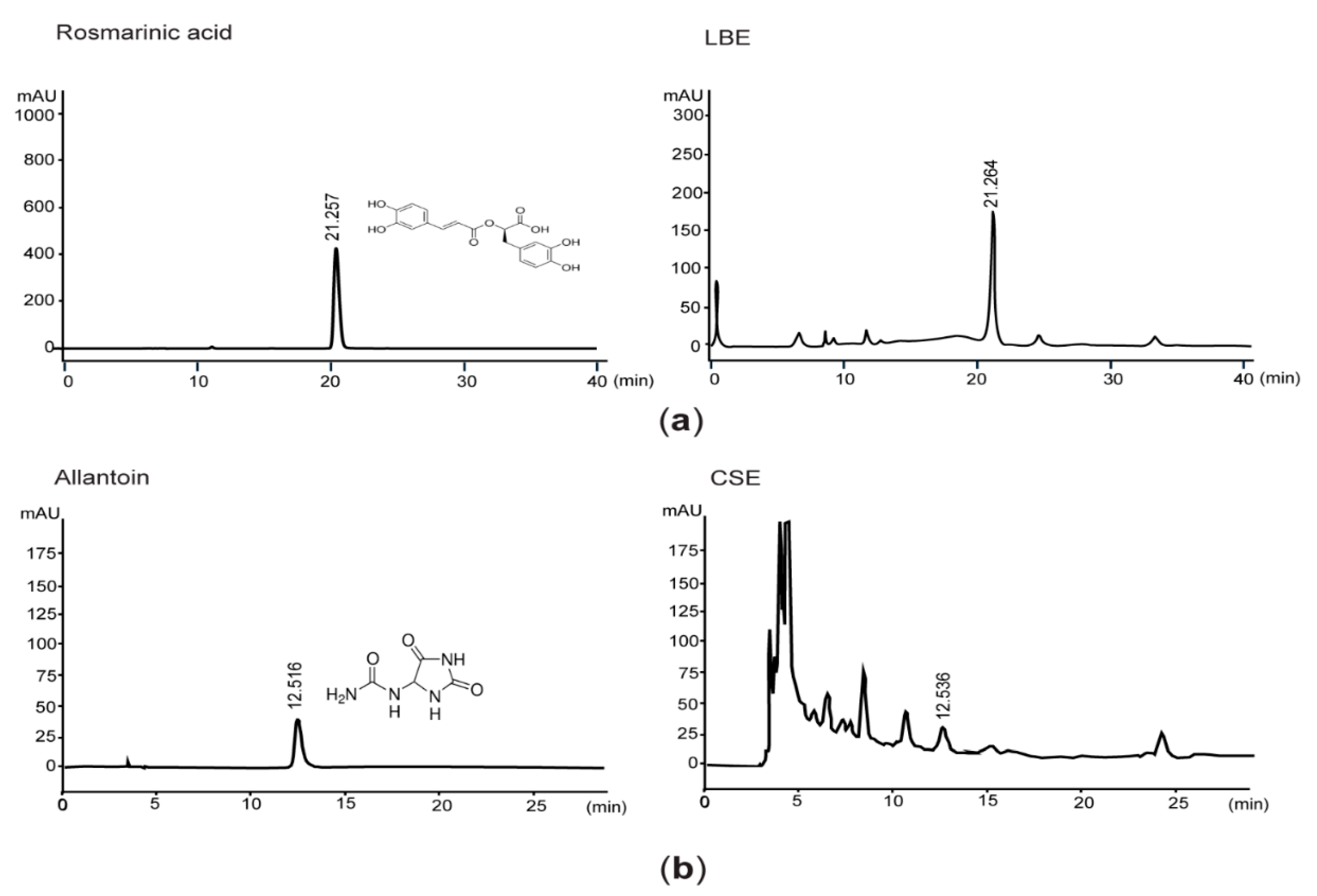
2.1. Quantification of Rosmarinic Acid and Allantoin by High-Performance Liquid Chromatography (HPLC)
2.2. Animal Husbandry and Treatment
2.3. Measurement of Body and Fat Weight
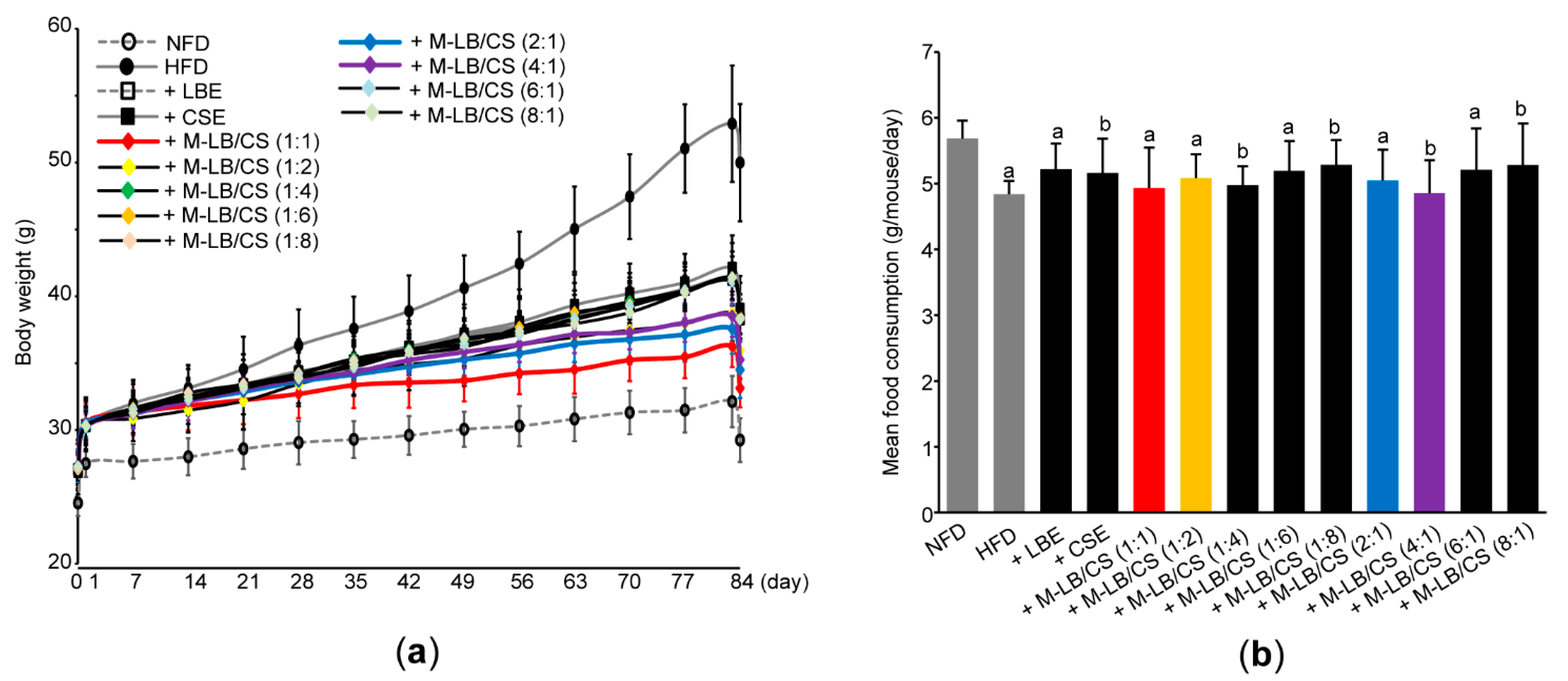
2.4. Measurement of Mean Daily Food Consumption
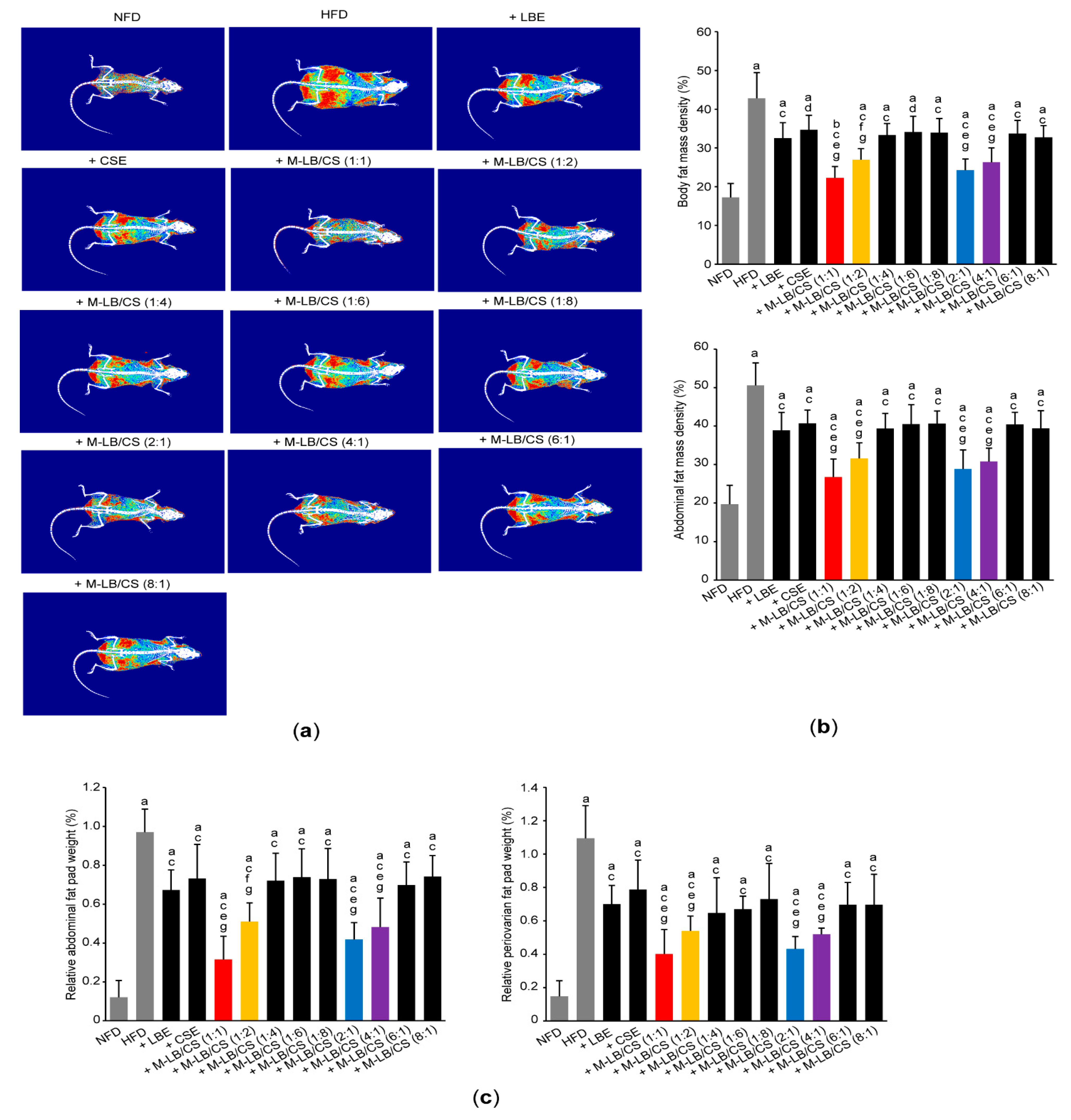
2.5. Measurement of Body and Abdominal Fat Density
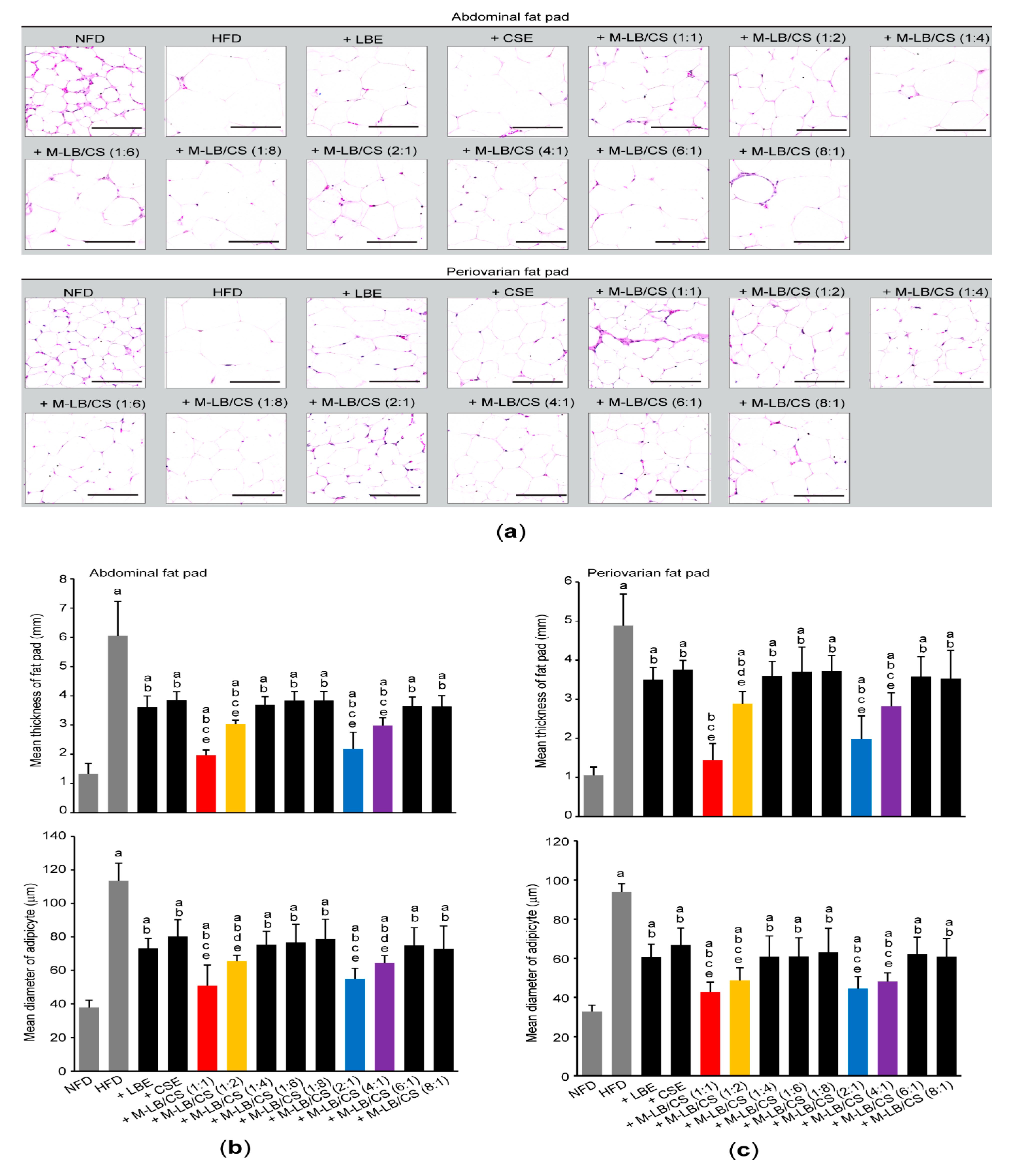
2.6. Histological Analyses
2.7. Measurement of Lipid Profiles
2.8. Quantitative Polymerase Chain Reaction (qPCR)
2.9. Measurement of Lipid Peroxidation and Antioxidant Activities
2.10. Statistical Analysis
3. Results
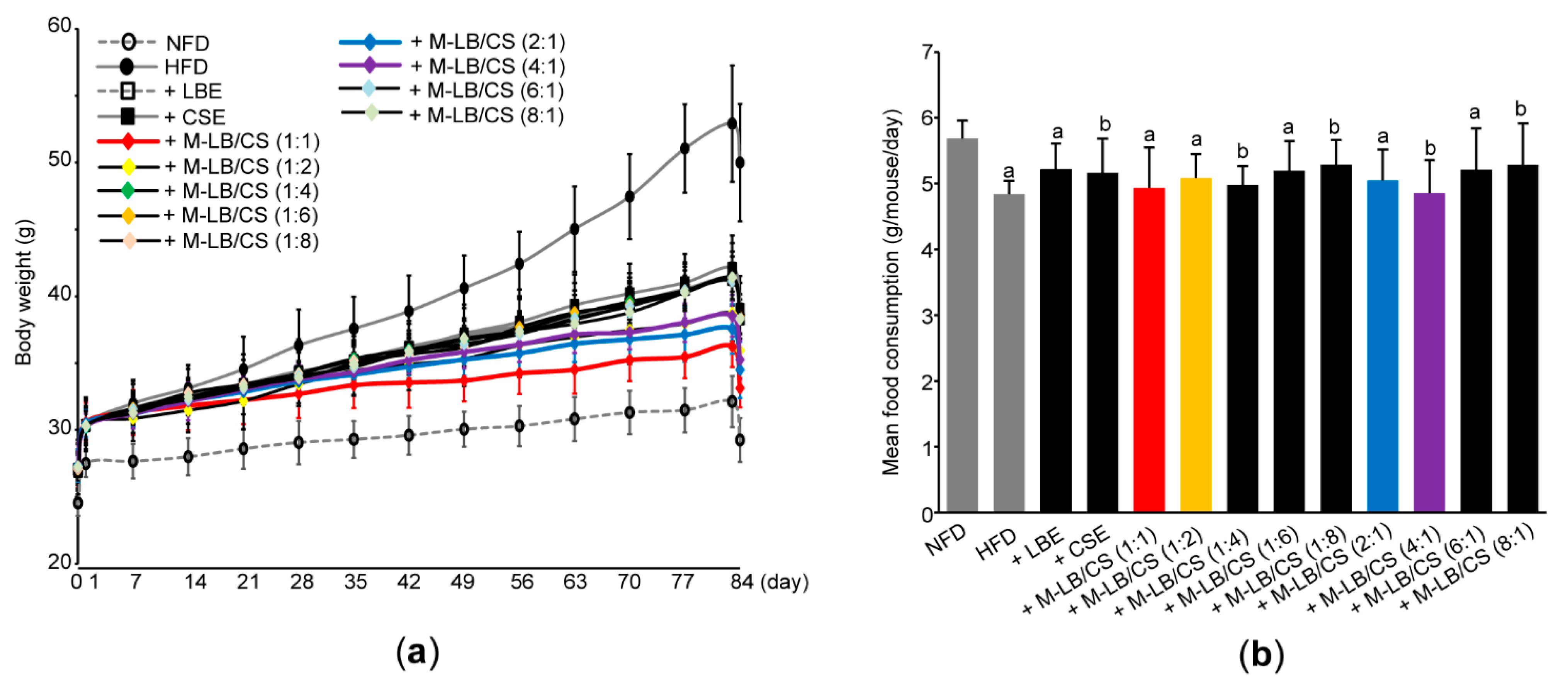
3.1. Mixture of Lemon Balm and Corn Silk Extracts Reduces Body Weight Gain in Mice Fed an HFD
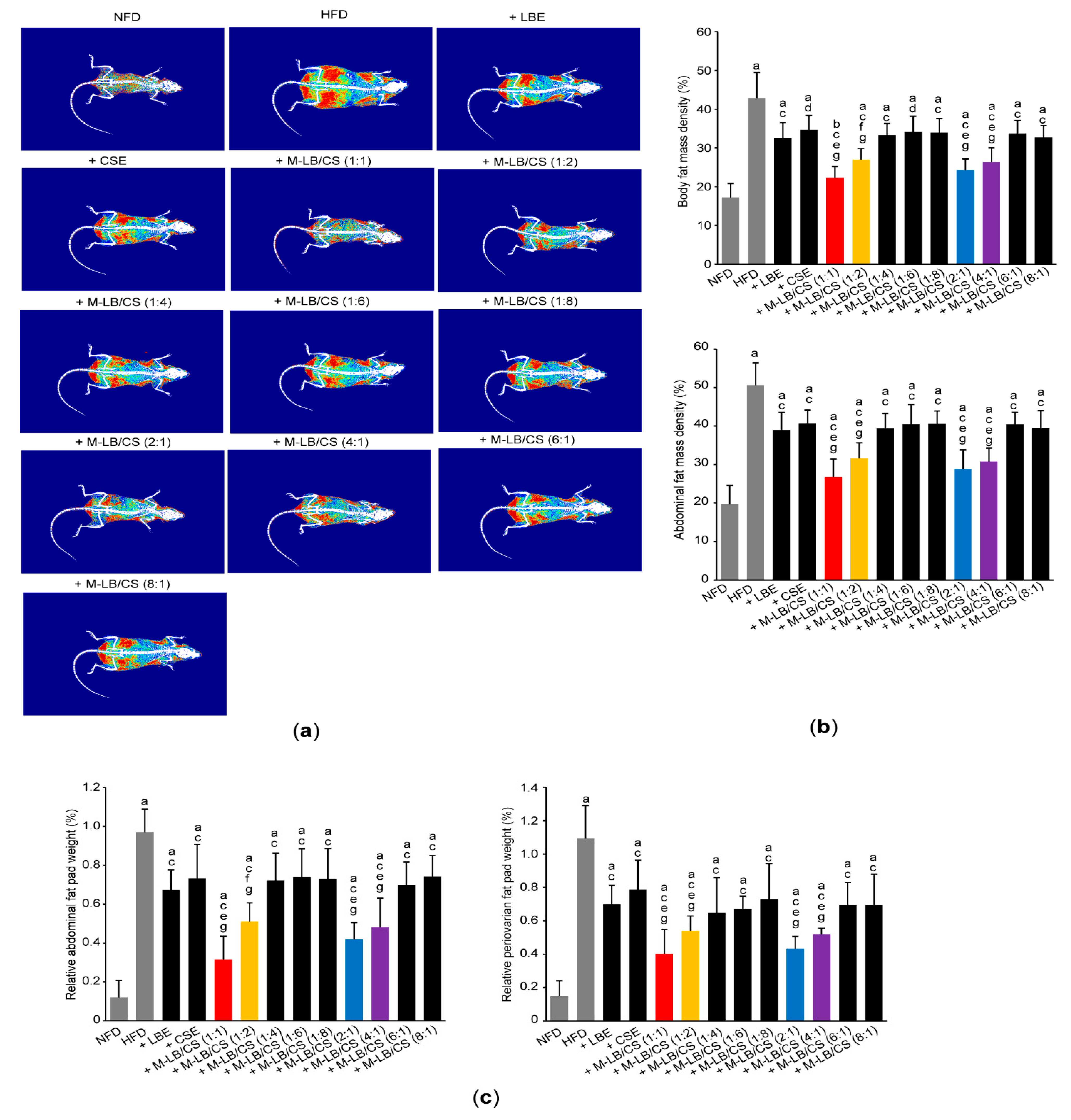
3.2. Mixture of Lemon Balm and Corn Silk Extracts Decreases Fat Deposition in Mice Fed an HFD
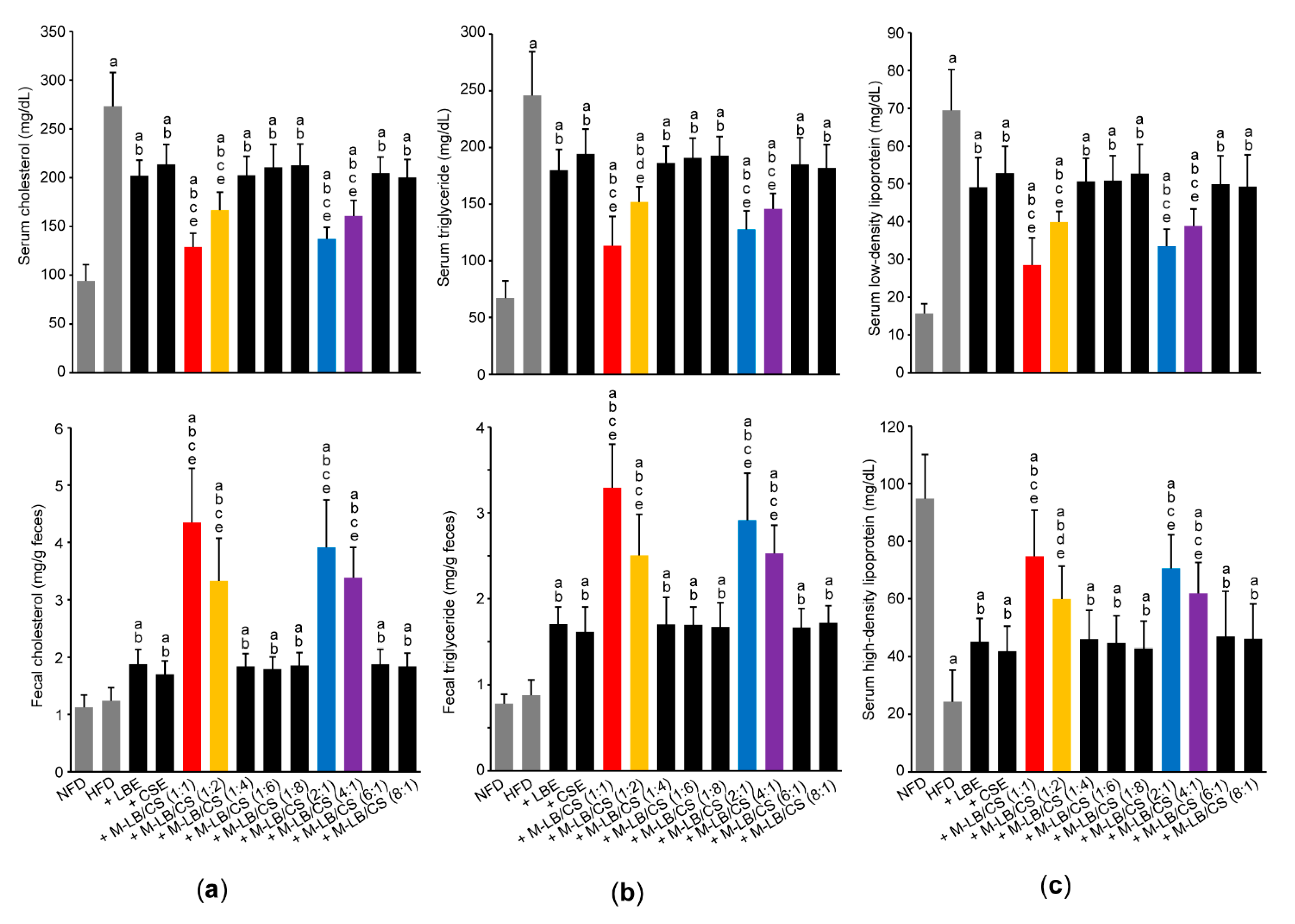
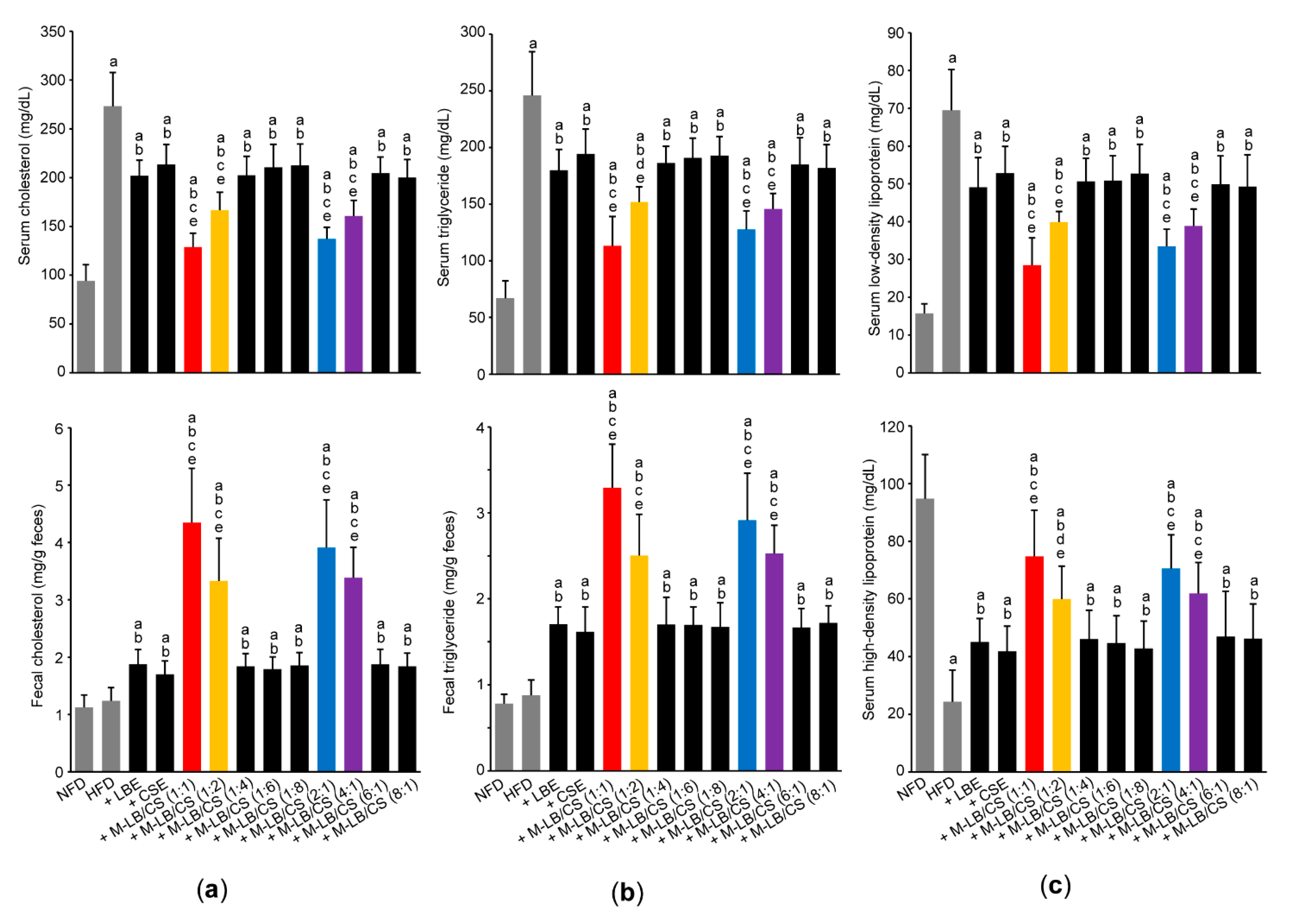
3.3. Mixture of Lemon Balm and Corn Silk Extracts Prevents Impaired Lipid Profiles in Mice Fed an HFD
3.4. Mixture of Lemon Balm and Corn Silk Extracts Mitigates Abnormal Changes in Specific Genes Associated with Lipid Metabolism in the Adipose Tissue
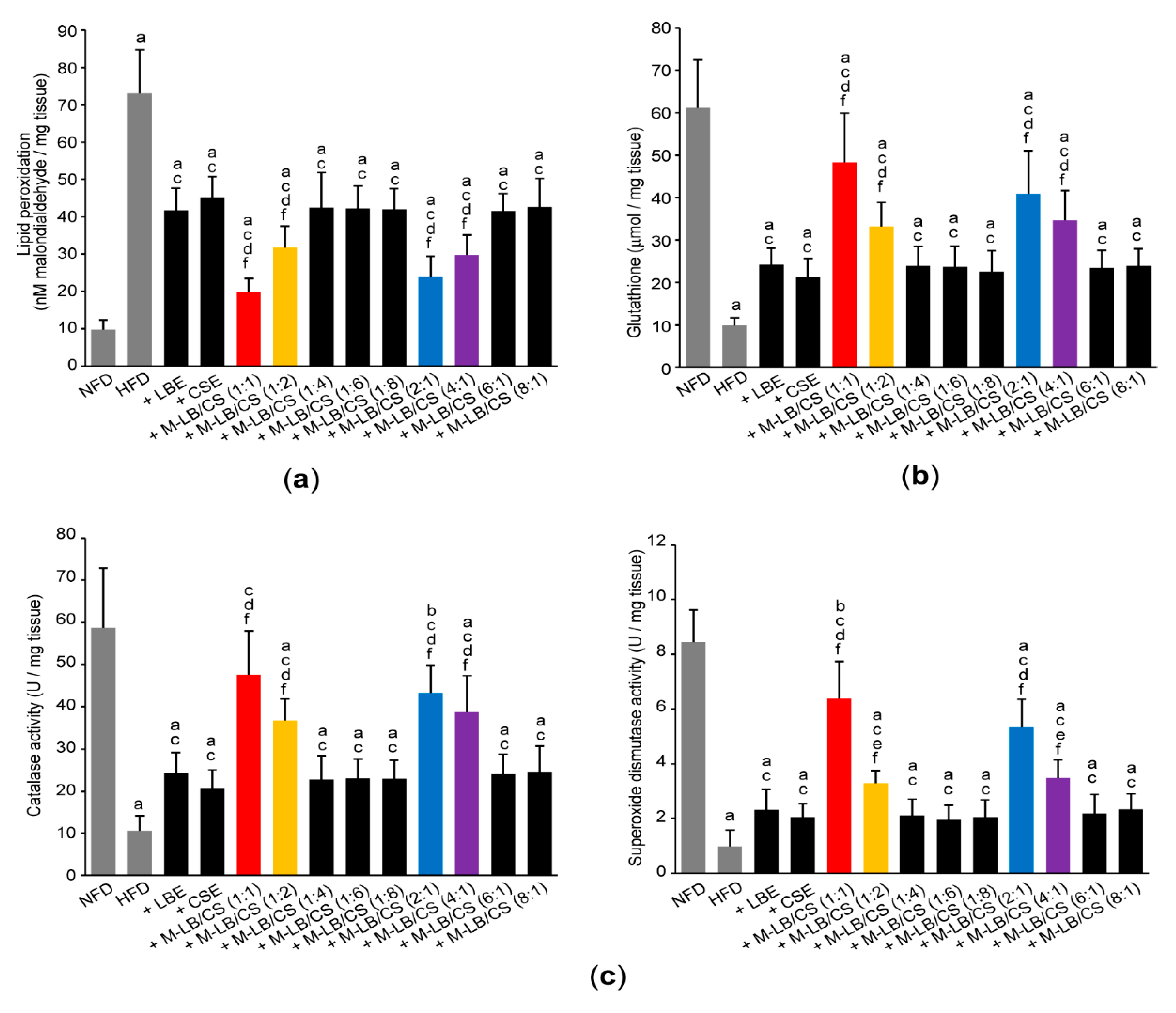
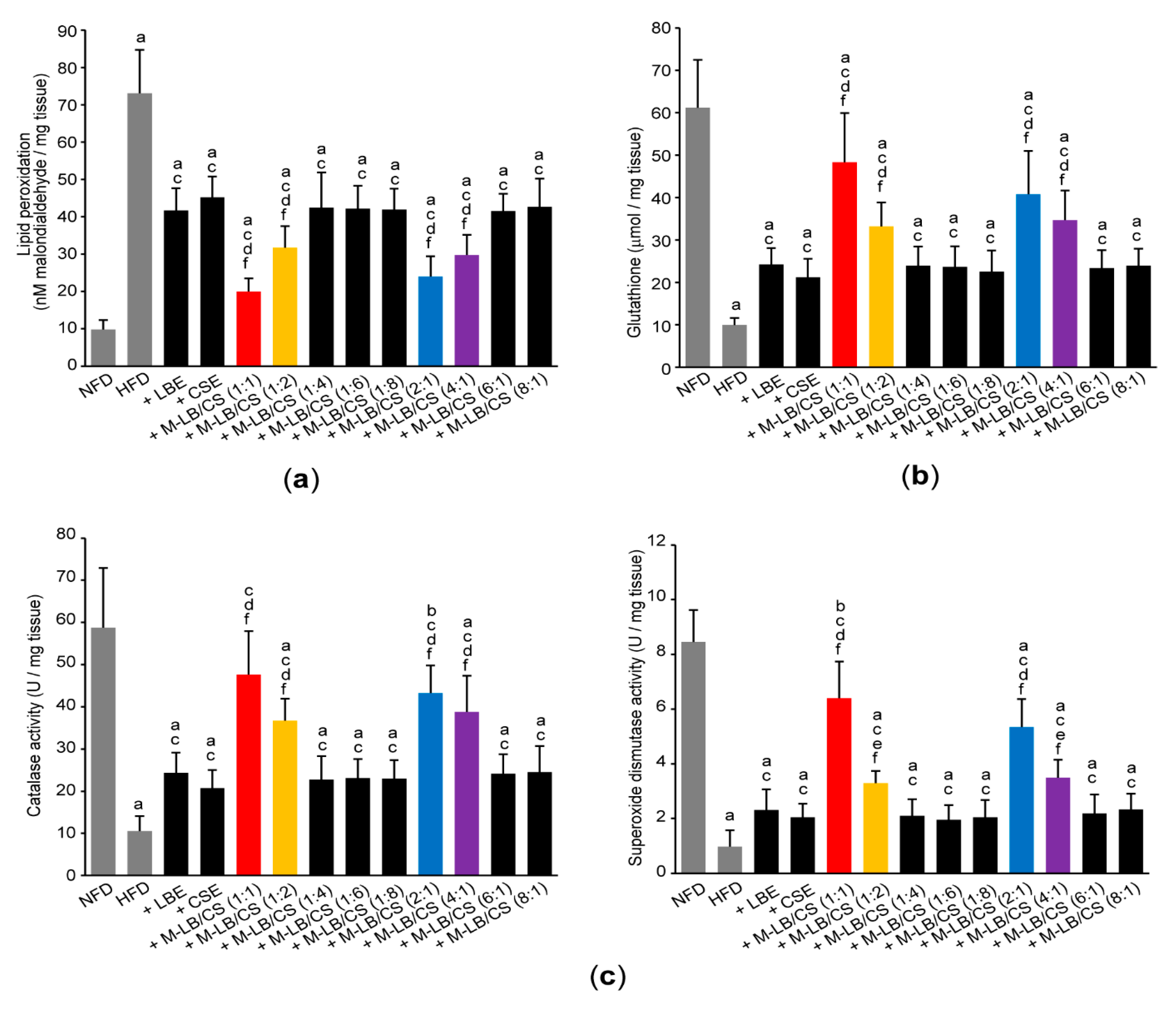
3.5. Mixture of Lemon Balm and Corn Silk Extracts Scavenges Lipid Peroxidation by Recovering Antioxidant Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Apovian, C.M. Obesity: Definition, comorbidities, causes, and burden. Am. J. Manag. Care 2016, 22, s176–s185. [Google Scholar] [PubMed]
- Obesity and Overweight. Available online: http://who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 30 October 2021).
- Tak, Y.J.; Lee, S.Y. Long-Term Efficacy and Safety of Anti-Obesity Treatment: Where Do We Stand? Curr. Obes. Rep. 2021, 10, 14–30. [Google Scholar] [CrossRef]
- Mollica, A.; Zengin, G.; Stefanucci, A.; Ferrante, C.; Menghini, L.; Orlando, G.; Brunetti, L.; Locatelli, M.; Dimmito, M.P.; Novellino, E.; et al. Nutraceutical potential of Corylus avellana daily supplments for obesity and related dysmetabolism. J. Funct. Foods 2018, 47, 562–574. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Dinda, B.; Dinda, M.; Roy, A.; Dinda, S. Dietary plant flavonoids in prevention of obesity and diabetes. Adv. Protein Chem. Struct. Biol. 2020, 120, 159–235. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.A.; Arts, I.C.; Ambergen, T.; Brants, H.A.; Dagnelie, P.C.; Goldbohm, R.A.; van den Brandt, P.A.; Weijenberg, M.P. Netherlands Cohort Study. Higher dietary flavone, flavonol, and catechin intakes are associated with less of an increase in BMI over time in women: A longitudinal analysis from the Netherlands Cohort Study. Am. J. Clin. Nutr. 2008, 88, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, A.; Sahebkar, A.; Javadi, B. Melissa officinalis L.—A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2016, 188, 204–228. [Google Scholar] [CrossRef] [PubMed]
- Hasanudin, K.; Hashim, P.; Mustafa, S. Corn silk (Stigma maydis) in healthcare: A phytochemical and pharmacological review. Molecules 2012, 17, 9697–9715. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, Q.; Fan, S.; Yang, X.; Ming, L.; Wang, H.; Liu, J. Rapid analysis and characterization of multiple constituents of corn silk aqueous extract using ultra-high-performance liquid chromatography combined with quadrupole time-of-flight mass spectrometry. J. Sep. Sci. 2019, 42, 3054–3066. [Google Scholar] [CrossRef]
- Lee, D.; Shin, Y.; Jang, J.; Park, Y.; Ahn, J.; Jeong, S.; Shin, S.S.; Yoon, M. The herbal extract ALS-L1023 from Melissa officinalis alleviates visceral obesity and insulin resistance in obese female C57BL/6J mice. J. Ethnopharmacol. 2020, 253, 112646. [Google Scholar] [CrossRef]
- Lee, D.; Shin, Y.; Roh, J.S.; Ahn, J.; Jeoong, S.; Shin, S.S.; Yoon, M. Lemon Balm Extract ALS-L1023 Regulates Obesity and Improves Insulin Sensitivity via Activation of Hepatic PPARα in High-Fat Diet-Fed Obese C57BL/6J Mice. Int. J. Mol. Sci. 2020, 21, 4256. [Google Scholar] [CrossRef]
- Hsu, Y.A.; Kuo, Y.H.; Chen, C.S.; Chen, Y.C.; Huang, C.C.; Chang, C.Y.; Lin, C.J.; Lin, C.W.; Lin, H.J.; Liu, F.T.; et al. Galectin-12 is Involved in Corn Silk-Induced Anti-Adipogenesis and Anti-Obesity Effects. Am. J. Chin. Med. 2018, 46, 1045–1063. [Google Scholar] [CrossRef]
- Kim, J.; Lee, H.; Lim, J.; Lee, H.; Yoon, S.; Shin, S.S.; Yoon, M. The lemon balm extract ALS-L1023 inhibits obesity and nonalcoholic fatty liver disease in female ovariectomized mice. Food Chem. Toxicol. 2017, 106, 292–305. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.H.; Kim, S.R.; Kang, H.J.; Kim, M.H.; Ha, A.W.; Kim, W.K. Corn silk extract improves cholesterol metabolism in C57BL/6J mouse fed high-fat diets. Nutr. Res. Pract. 2016, 10, 501–506. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.Y.; Kim, S.L.; Kang, H.J.; Kim, M.H.; Ha, A.W.; Kim, W.K. High maysin corn silk extract reduces body weight and fat deposition in C57BL/6J mice fed high-fat diets. Nutr. Res. Pract. 2016, 10, 575–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, B.R.; Cho, I.J.; Jung, S.J.; Kim, J.K.; Park, S.M.; Lee, D.G.; Ku, S.K.; Park, K.M. Lemon balm and dandelion leaf extract synergistically alleviate ethanol-induced hepatotoxicity by enhancing antioxidant and anti-inflammatory activity. J. Food Biochem. 2020, 44, e13232. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.-R.; Cho, I.J.; Jung, S.-J.; Kim, J.K.; Lee, D.G.; Ku, S.K.; Park, K.-M. Study on the hepatoprotective effects of lemon balm and dandelion leaf extract combination in carbon tetrachloride-mediated liver injured mice. Herb. Formula Sci. 2019, 27, 199–211. [Google Scholar] [CrossRef]
- Choi, B.R.; Kim, H.J.; Lee, Y.J.; Ku, S.K. Anti-Diabetic Obesity Effects of Wasabia Japonica Matsum Leaf Extract on 45% Kcal High-Fat Diet-Fed Mice. Nutrients 2020, 12, 2837. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Lee, Y.S.; Seol, D.J.; Cho, I.J.; Ku, S.K.; Choi, J.S.; Lee, H.J. Anti-obesity and fatty liver-preventing activities of Lonicera caerulea in high-fat diet-fed mice. Int. J. Mol. Med. 2018, 42, 3047–3064. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Maksimović, Z.; Malenović, A.; Jancić, B.; Kovacević, N. Quantification of allantoin in various Zea mays L. hybrids by RP-HPLC with UV detection. Pharmazie 2004, 59, 524–527. [Google Scholar]
- Sun, C.; Mao, S.; Chen, S.; Zhang, W.; Liu, C. PPARs-Orchestrated Metabolic Homeostasis in the Adipose Tissue. Int. J. Mol. Sci. 2021, 22, 8974. [Google Scholar] [CrossRef] [PubMed]
- Payne, V.A.; Au, W.S.; Lowe, C.E.; Rahman, S.M.; Friedman, J.E.; O’Rahilly, S.; Rochford, J.J. C/EBP transcription factors regulate SREBP1c gene expression during adipogenesis. Biochem. J. 2009, 425, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bougarne, N.; Weyers, B.; Desmet, S.J.; Deckers, J.; Ray, D.W.; Staels, B.; De Bosscher, K. Molecular Actions of PPARα in Lipid Metabolism and Inflammation. Endocr. Rev. 2018, 39, 760–802. [Google Scholar] [CrossRef] [Green Version]
- Hamidi-Zad, Z.; Moslehi, A.; Rastegarpanah, M. Attenuating effects of allantoin on oxidative stress in a mouse model of nonalcoholic steatohepatitis: Role of SIRT1/Nrf2 pathway. Res. Pharm. Sci. 2021, 16, 651–659. [Google Scholar] [CrossRef]
- Xu, H.Y.; Zheng, H.C.; Zhang, H.W.; Zhang, J.Y.; Ma, C.M. Comparison of Antioxidant Constituents of Agriophyllum squarrosum Seed with Conventional Crop Seeds. J. Food Sci. 2018, 83, 1823–1831. [Google Scholar] [CrossRef]
- Adomako-Bonsu, A.G.; Chan, S.L.; Pratten, M.; Fry, J.R. Antioxidant activity of rosmarinic acid and its principal metabolites in chemical and cellular systems: Importance of physico-chemical characteristics. Toxicol. In Vitro 2017, 40, 248–255. [Google Scholar] [CrossRef]
- Anwar, S.; Shamsi, A.; Shahbaaz, M.; Queen, A.; Khan, P.; Hasan, G.M.; Islam, A.; Alajmi, M.F.; Hussain, A.; Ahmad, F.; et al. Rosmarinic Acid Exhibits Anticancer Effects via MARK4 Inhibition. Sci. Rep. 2020, 10, 10300. [Google Scholar] [CrossRef] [PubMed]
- Elufioye, T.O.; Habtemariam, S. Hepatoprotective effects of rosmarinic acid: Insight into its mechanisms of action. Biomed. Pharmacother. 2019, 112, 108600. [Google Scholar] [CrossRef] [PubMed]
- Guginski, G.; Luiz, A.P.; Silva, M.D.; Massaro, M.; Martins, D.F.; Chaves, J.; Mattos, R.W.; Silveira, D.; Ferreira, V.M.; Calixto, J.B.; et al. Mechanisms involved in the antinociception caused by ethanolic extract obtained from the leaves of Melissa officinalis (lemon balm) in mice. Pharmacol. Biochem. Behav. 2009, 93, 10–16. [Google Scholar] [CrossRef]
- Xavier, C.P.; Lima, C.F.; Fernandes-Ferreira, M.; Pereira-Wilson, C. Salvia fruticosa, Salvia officinalis, and rosmarinic acid induce apoptosis and inhibit proliferation of human colorectal cell lines: The role in MAPK/ERK pathway. Nutr. Cancer 2009, 61, 564–571. [Google Scholar] [CrossRef] [Green Version]
- Vasileva, L.V.; Savova, M.S.; Tews, D.; Wabitsch, M.; Georgiev, M.I. Rosmarinic acid attenuates obesity and obesity-related inflammation in human adipocytes. Food Chem. Toxicol. 2021, 149, 112002. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Tong, L.; Cheng, J.; Wang, G.; Qin, L.; Wan, Z. Rosmarinic acid suppresses adipogenesis, lipolysis in 3T3-L1 adipocytes, lipopolysaccharide-stimulated tumor necrosis factor-α secretion in macrophages, and inflammatory mediators in 3T3-L1 adipocytes. Food Nutr. Res. 2017, 61, 1330096. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.J.; Park, S.J.; Woo, H.; Lee, H.E.; Kim, H.J.; Kwon, G.; Gao, Q.; Jang, D.S.; Ryu, J.H. Effects of allantoin on cognitive function and hippocampal neurogenesis. Food Chem. Toxicol. 2014, 64, 210–216. [Google Scholar] [CrossRef]
- Chung, H.H.; Lee, K.S.; Cheng, J.T. Decrease of Obesity by Allantoin via Imidazoline I1 -Receptor Activation in High Fat Diet-Fed Mice. Evid. Based Complement. Alternat. Med. 2013, 2013, 589309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haghi, G.; Arshi, R.; Safaei, A. Improved high-performance liquid chromatography (HPLC) method for qualitative and quantitative analysis of allantoin in Zea mays. J. Agric. Food Chem. 2008, 56, 1205–1209. [Google Scholar] [CrossRef] [PubMed]
- Todd, C.D.; Tipton, P.A.; Blevins, D.G.; Piedras, P.; Pineda, M.; Polacco, J.C. Update on ureide degradation in legumes. J. Exp. Bot. 2006, 57, 5–12. [Google Scholar] [CrossRef]
- Araújo, L.U.; Grabe-Guimarães, A.; Mosqueira, V.C.; Carneiro, C.M.; Silva-Barcellos, N.M. Profile of wound healing process induced by allantoin. Acta Cir. Bras. 2010, 25, 460–461. [Google Scholar] [CrossRef]
- Eslami-Farsani, M.; Moslehi, A.; Hatami-Shahmir, A. Allantoin improves histopathological evaluations in a rat model of gastritis. Physiol. Int. 2018, 105, 325–334. [Google Scholar] [CrossRef]
- Lee, M.Y.; Lee, N.H.; Jung, D.; Lee, J.A.; Seo, C.S.; Lee, H.; Kim, J.H.; Shin, H.K. Protective effects of allantoin against ovalbumin (OVA)-induced lung inflammation in a murine model of asthma. Int. Immunopharmacol. 2010, 10, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.T.; Chiu, N.H.; Chung, H.H.; Hsu, C.T.; Lee, W.J.; Cheng, J.T. Stimulatory effect of allantoin on imidazoline I1 receptors in animal and cell line. Horm. Metab. Res. 2012, 44, 879–884. [Google Scholar] [CrossRef]
- Nyandwi, J.B.; Ko, Y.S.; Jin, H.; Yun, S.P.; Park, S.W.; Kim, H.J. Rosmarinic Acid Exhibits a Lipid-Lowering Effect by Modulating the Expression of Reverse Cholesterol Transporters and Lipid Metabolism in High-Fat Diet-Fed Mice. Biomolecules 2021, 11, 1470. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Sun, H.; Peng, J.; Gao, C.; Bao, L.; Ji, R.; Zhang, C.; Zhu, W.; Jin, Y. Rosmarinic acid exerts an antagonistic effect on nonalcoholic fatty liver disease by regulating the YAP1/TAZ-PPARγ/PGC-1α signaling pathway. Phytother. Res. 2021, 35, 1010–1022. [Google Scholar] [CrossRef] [PubMed]
- Seyedan, A.; Alshawsh, M.A.; Alshagga, M.A.; Mohamed, Z. Antiobesity and Lipid Lowering Effects of Orthosiphon stamineus in High-Fat Diet-Induced Obese Mice. Planta Med. 2017, 83, 684–692. [Google Scholar] [CrossRef]
- Sun, A.; Hu, X.; Chen, H.; Ma, Y.; Yan, X.; Peng, D.; Ping, J.; Yan, Y. Ursolic acid induces white adipose tissue beiging in high-fat-diet obese male mice. Food Funct. 2021, 12, 6490–6501. [Google Scholar] [CrossRef] [PubMed]
- Sousa, G.M.; Cazarin, C.B.B.; Maróstica Junior, M.R.; Lamas, C.A.; Quitete, V.H.A.C.; Pastore, G.M.; Bicas, J.L. The effect of α-terpineol enantiomers on biomarkers of rats fed a high-fat diet. Heliyon 2020, 6, e03752. [Google Scholar] [CrossRef]
- Kim, H.M.; Kim, Y.; Lee, E.S.; Huh, J.H.; Chung, C.H. Caffeic acid ameliorates hepatic steatosis and reduces ER stress in high fat diet-induced obese mice by regulating autophagy. Nutrition 2018, 55–56, 63–70. [Google Scholar] [CrossRef]
- Lee, C.W.; Seo, J.Y.; Kim, S.L.; Lee, J.; Choi, J.W.; Park, Y.I. Corn silk maysin ameliorates obesity in vitro and in vivo via suppression of lipogenesis, differentiation, and function of adipocytes. Biomed. Pharmacother. 2017, 93, 267–275. [Google Scholar] [CrossRef]
- Kwon, E.Y.; Jung, U.J.; Park, T.; Yun, J.W.; Choi, M.S. Luteolin attenuates hepatic steatosis and insulin resistance through the interplay between the liver and adipose tissue in mice with diet-induced obesity. Diabetes 2015, 64, 1658–1669. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Ma, Y.; Liu, D. Rutin suppresses palmitic acids-triggered inflammation in macrophages and blocks high fat diet-induced obesity and fatty liver in mice. Pharm. Res. 2013, 30, 2940–2950. [Google Scholar] [CrossRef] [Green Version]
- de Melo, C.L.; Queiroz, M.G.; Fonseca, S.G.; Bizerra, A.M.; Lemos, T.L.; Melo, T.S.; Santos, F.A.; Rao, V.S. Oleanolic acid, a natural triterpenoid improves blood glucose tolerance in normal mice and ameliorates visceral obesity in mice fed a high-fat diet. Chem. Biol. Interact. 2010, 185, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Garawi, F.; Devries, K.; Thorogood, N.; Uauy, R. Global differences between women and men in the prevalence of obesity: Is there an association with gender inequality? Eur. J. Clin. Nutr. 2014, 68, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.M.; Yi, S.J.; Cho, I.J.; Ku, S.K. Red-koji fermented red ginseng ameliorates high fat diet-induced metabolic disorders in mice. Nutrients 2013, 5, 4316–4332. [Google Scholar] [CrossRef] [Green Version]
- Hua, H.; Zhang, Y.; Zhao, F.; Chen, K.; Wu, T.; Liu, Q.; Huang, S.; Zhang, A.; Jia, Z. Celastrol inhibits intestinal lipid absorption by reprofiling the gut microbiota to attenuate high-fat diet-induced obesity. iScience 2021, 24, 102077. [Google Scholar] [CrossRef]
- Xu, P.; Dai, S.; Wang, J.; Zhang, J.; Liu, J.; Wang, F.; Zhai, Y. Preventive obesity agent montmorillonite adsorbs dietary lipids and enhances lipid excretion from the digestive tract. Sci. Rep. 2016, 6, 19659. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, S.; Taniguchi, Y.; Saka, A.; Yoshida, A.; Yajima, H. Prevention of diet-induced obesity by dietary black tea polyphenols extract in vitro and in vivo. Nutrition 2011, 27, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.F.; Kusumoto, A.; Abe, K.; Hosoda, K.; Kiso, Y.; Wang, M.F.; Yamamoto, S. Polyphenol-enriched oolong tea increases fecal lipid excretion. Eur. J. Clin. Nutr. 2006, 60, 1330–1336. [Google Scholar] [CrossRef]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [Green Version]
- Weidner, C.; Wowro, S.J.; Freiwald, A.; Kodelja, V.; Abdel-Aziz, H.; Kelber, O.; Sauer, S. Lemon balm extract causes potent antihyperglycemic and antihyperlipidemic effects in insulin-resistant obese mice. Mol. Nutr. Food Res. 2014, 58, 903–907. [Google Scholar] [CrossRef] [PubMed]
- Koska, J.; Stefan, N.; Permana, P.A.; Weyer, C.; Sonoda, M.; Bogardus, C.; Smith, S.R.; Joanisse, D.R.; Funahashi, T.; Krakoff, J.; et al. Increased fat accumulation in liver may link insulin resistance with subcutaneous abdominal adipocyte enlargement, visceral adiposity, and hypoadiponectinemia in obese individuals. Am. J. Clin. Nutr. 2008, 87, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, J.H.; Rutkowski, J.M.; Scherer, P.E. Adiponectin, Leptin, and Fatty Acids in the Maintenance of Metabolic Homeostasis through Adipose Tissue Crosstalk. Cell Metab. 2016, 23, 770–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campfield, L.A.; Smith, F.J.; Guisez, Y.; Devos, R.; Burn, P. Recombinant mouse OB protein: Evidence for a peripheral signal linking adiposity and central neural networks. Science 1995, 269, 546–549. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Backward Primer | RefSeq Accession No. | Amplicon Size (bp) |
|---|---|---|---|---|
| Adiponectin | 5′-CCCAAGGGAACTTGTGCAGGTTGGATG-3′ | 5′-GTTGGTATCATGGTAGAGAAGAAAGCC-3′ | NM_009605.5 | 639 |
| C/EBPα | 5′-TGGACAAGAACAGCAACGAGTAC-3′ | 5′-CGGTCATTGTCACTGGTCAACT-3′ | NM_001287514.1 | 136 |
| C/EBPβ | 5′-AAGCTGAGCGACGAGTACAAGA-3′ | 5′-GTCAGCTCCAGCACCTTGTG-3′ | NM_001287738.1 | 116 |
| GAPDH | 5′-CATCTTCCAGGAGCGAGACC-3′ | 5′-TCCACCACCCTGTTGCTGTA-3′ | NM_001289726.1 | 753 |
| Leptin | 5′-CCAAAACCCTCATCAAGACC-3′ | 5′-GTCCAACTGTTGAAGAATGTCCC-3′ | NM_008493.3 | 390 |
| PPARα | 5′-ATGCCAGTACTGCCGTTTTC-3′ | 5′-GGCCTTGACCTTGTTCATGT-3′ | NM_011144.6 | 220 |
| PPARγ | 5′-AGTGGAGACCGCCCAGG-3′ | 5′-GCAGCAGGTTGTCTTGGATGT-3′ | NM_00127330.2 | 64 |
| SREBP-1c | 5′-AGCCTGGCCATCTGTGAGAA-3′ | 5′-CAGACTGGTACGGGCCACAA-3′ | NM_011480.4 | 132 |
| UCP2 | 5′-CCGCATTGGCCTCTACGACTCT-3′ | 5′-CCCCGAAGGCAGAAGTGAAGTG-3′ | NM_011671.5 | 386 |
| Experimental Group | Relative mRNA Level (Folds) | |||||||
|---|---|---|---|---|---|---|---|---|
| C/BEPα | C/BEPβ | PPARγ | SREBP-1c | PPARα | UCP2 | Leptin | Adiponectin | |
| NFD | 1.00 ± 0.06 | 1.01 ± 0.06 | 1.00 ± 0.07 | 1.01 ± 0.06 | 1.00 ± 0.04 | 1.02 ± 0.08 | 1.03 ± 0.08 | 0.99 ± 0.16 |
| HFD | 2.05 ± 0.34 a | 3.74 ± 0.86 a | 7.17 ± 1.01 a | 2.46 ± 0.54 a | 0.22 ± 0.05 a | 0.20 ± 0.06 a | 6.94 ± 0.99 a | 0.14 ± 0.07 a |
| HFD + LBE | 1.53 ± 0.07 a,b | 2.48 ± 0.41 a,b | 4.79 ± 0.71 a,b | 1.75 ± 0.15 a,b | 0.36 ± 0.08 a,b | 0.36 ± 0.04 a,b | 4.32 ± 0.55 a,b | 0.30 ± 0.06 a,b |
| HFD + CSE | 1.63 ± 0.17 a,b | 2.76 ± 0.31 a,b | 5.02 ± 0.89 a,b | 1.87 ± 0.11 a,b | 0.31 ± 0.05 a,b | 0.30 ± 0.05 a,c | 4.71 ± 0.63 a,b | 0.26 ± 0.08 a,c |
| HFD + M-LB/CS (1:1) | 1.17 ± 0.11 a,b,d,f | 1.34 ± 0.29 a,b,d,f | 2.17 ± 0.49 a,b,d,f | 1.26 ± 0.17 a,b,d,f | 0.69 ± 0.14 a,b,d,f | 0.69 ± 0.10 a,b,d,f | 1.51 ± 0.37 a,b,d,f | 0.58 ± 0.15 a,b,d,f |
| HFD + M-LB/CS (1:2) | 1.39 ± 0.08 a,b,d,g | 1.87 ± 0.37 a,b,e,f | 3.89 ± 0.84 a,b,e,g | 1.56 ± 0.08 a,b,d,f | 0.53 ± 0.11 a,b,d,f | 0.51 ± 0.11 a,b,d,f | 3.03 ± 0.30 a,b,d,f | 0.39 ± 0.07 a,b,e,f |
| HFD + M-LB/CS (1:4) | 1.52 ± 0.16 a,b | 2.64 ± 0.39 a,b | 4.86 ± 0.83 a,b | 1.76 ± 0.22 a,b | 0.34 ± 0.08 a,b | 0.35 ± 0.08 a,b | 4.24 ± 0.68 a,b | 0.30 ± 0.07 a,b |
| HFD + M-LB/CS (1:6) | 1.54 ± 0.12 a,b | 2.62 ± 0.37 a,b | 4.92 ± 0.93 a,b | 1.77 ± 0.24 a,b | 0.34 ± 0.08 a,b | 0.33 ± 0.10 a,b | 4.30 ± 0.78 a,b | 0.30 ± 0.09 a,b |
| HFD + M-LB/CS (1:8) | 1.55 ± 0.16 a,b | 2.74 ± 0.34 a,b | 4.98 ± 1.22 a,b | 1.86 ± 0.18 a,b | 0.32 ± 0.06 a,b | 0.34 ± 0.11 a,b | 4.30 ± 0.90 a,b | 0.30 ± 0.09 a,b |
| HFD + M-LB/CS (2:1) | 1.25 ± 0.09 a,b,d,f | 1.52 ± 0.31 a,b,d,f | 2.55 ± 0.59 a,b,d,f | 1.36 ± 0.11 a,b,d,f | 0.63 ± 0.12 a,b,d,f | 0.61 ± 0.11 a,b,d,f | 2.05 ± 0.59 a,b,d,f | 0.51 ± 0.10 a,b,d,f |
| HFD + M-LB/CS (4:1) | 1.34 ± 0.12 a,b,d,f | 1.84 ± 0.26 a,b,d,f | 3.85 ± 0.53 a,b,d,f | 1.51 ± 0.16 a,b,e,f | 0.55 ± 0.09 a,b,d,f | 0.53 ± 0.09 a,b,d,f | 2.81 ± 0.42 a,b,d,f | 0.41 ± 0.08 a,b,e,f |
| HFD + M-LB/CS (6:1) | 1.53 ± 0.20 a,b | 2.50 ± 0.48 a,b | 4.77 ± 1.02 a,b | 1.79 ± 0.12 a,b | 0.34 ± 0.06 a,b | 0.35 ± 0.10 a,b | 4.22 ± 0.76 a,b | 0.32 ± 0.04 a,b |
| HFD + M-LB/CS (8:1) | 1.54 ± 0.18 a,b | 2.45 ± 0.43 a,b | 4.80 ± 0.27 a,b | 1.76 ± 0.19 a,b | 0.36 ± 0.09 a,b | 0.37 ± 0.11 a,b | 4.32 ± 0.79 a,b | 0.32 ± 0.11 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, I.-J.; Kim, S.-E.; Choi, B.-R.; Park, H.-R.; Park, J.-E.; Hong, S.-H.; Kwon, Y.-S.; Oh, W.-S.; Ku, S.-K. Lemon Balm and Corn Silk Extracts Mitigate High-Fat Diet-Induced Obesity in Mice. Antioxidants 2021, 10, 2015. https://doi.org/10.3390/antiox10122015
Cho I-J, Kim S-E, Choi B-R, Park H-R, Park J-E, Hong S-H, Kwon Y-S, Oh W-S, Ku S-K. Lemon Balm and Corn Silk Extracts Mitigate High-Fat Diet-Induced Obesity in Mice. Antioxidants. 2021; 10(12):2015. https://doi.org/10.3390/antiox10122015
Chicago/Turabian StyleCho, Il-Je, Sung-Eon Kim, Beom-Rak Choi, Hye-Rim Park, Jeong-Eun Park, Seong-Hwa Hong, Young-Sam Kwon, Won-Seok Oh, and Sae-Kwang Ku. 2021. "Lemon Balm and Corn Silk Extracts Mitigate High-Fat Diet-Induced Obesity in Mice" Antioxidants 10, no. 12: 2015. https://doi.org/10.3390/antiox10122015
APA StyleCho, I.-J., Kim, S.-E., Choi, B.-R., Park, H.-R., Park, J.-E., Hong, S.-H., Kwon, Y.-S., Oh, W.-S., & Ku, S.-K. (2021). Lemon Balm and Corn Silk Extracts Mitigate High-Fat Diet-Induced Obesity in Mice. Antioxidants, 10(12), 2015. https://doi.org/10.3390/antiox10122015