
Ameliorative Effect of Annona muricata (Graviola) Extract on Hyperglycemia Induced Hepatic Damage in Type 2 Diabetic Mice



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Annona Muricata Extracts (AME)
2.2. UHPLC Analyses
2.3. Total Polyphenol Contents
2.4. Animals and Diabetes Induction
2.5. Experimental Design
2.6. Oral Glucose Tolerance Test
= {15 min × (BG0min + BG15min) × 1/2} + {15 min × (BG15min + BG30min) × 1/2} + {30 min × (BG30min + BG60min) × 1/2} + {30 min × (BG60min + BG90min) × 1/2} + {30 min × (BG90min + BG120min) × 1/2}
2.7. Plasma Hemoglobin A1c (HbA1c) Level
2.8. Plasma Insulin Level
2.9. Hepatic Damage Related Markers
2.10. Hepatic Lipid Profiles
2.11. Histological Analysis
2.12. Western Blot Assay
2.13. Statistical Analysis
3. Results
3.1. Identification of Major Natural Compounds in AME
3.2. The Total Phenolic Content
3.3. Effect of AME on Body Weight, Liver Weight and Diet Intake in Type 2 Diabetes Mice
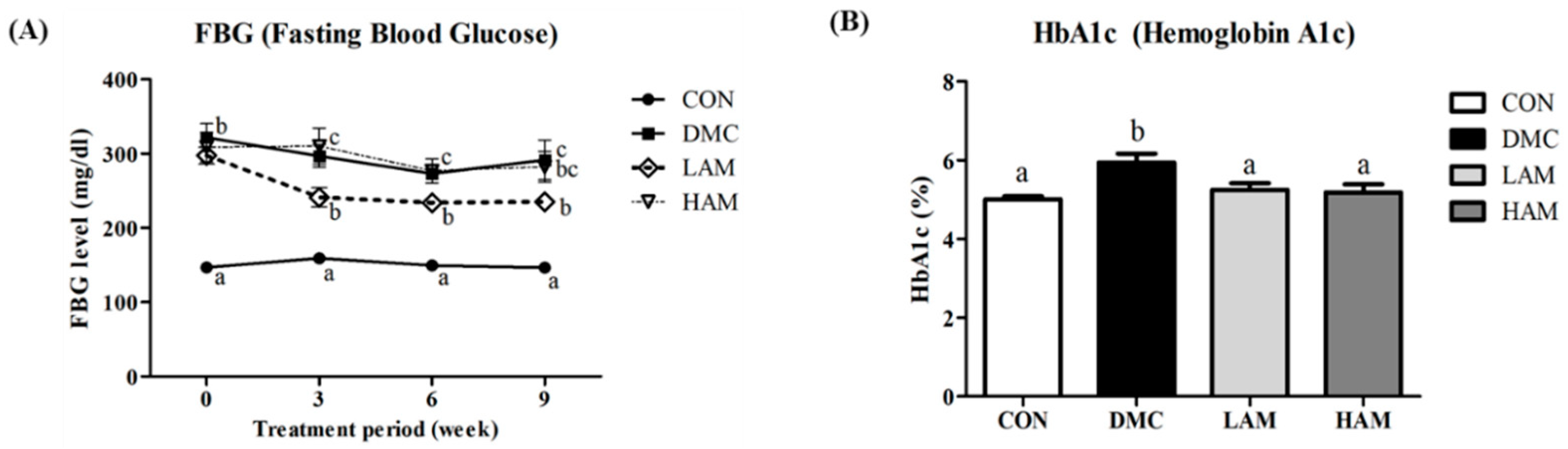
3.4. Effect of AME on Fasting Blood Glucose and Hemoglobin A1c in Type 2 Diabetes Mice
3.5. Effect of AME on Glucose Intolerance and Insulin Resistance in Type 2 Diabetes Mice
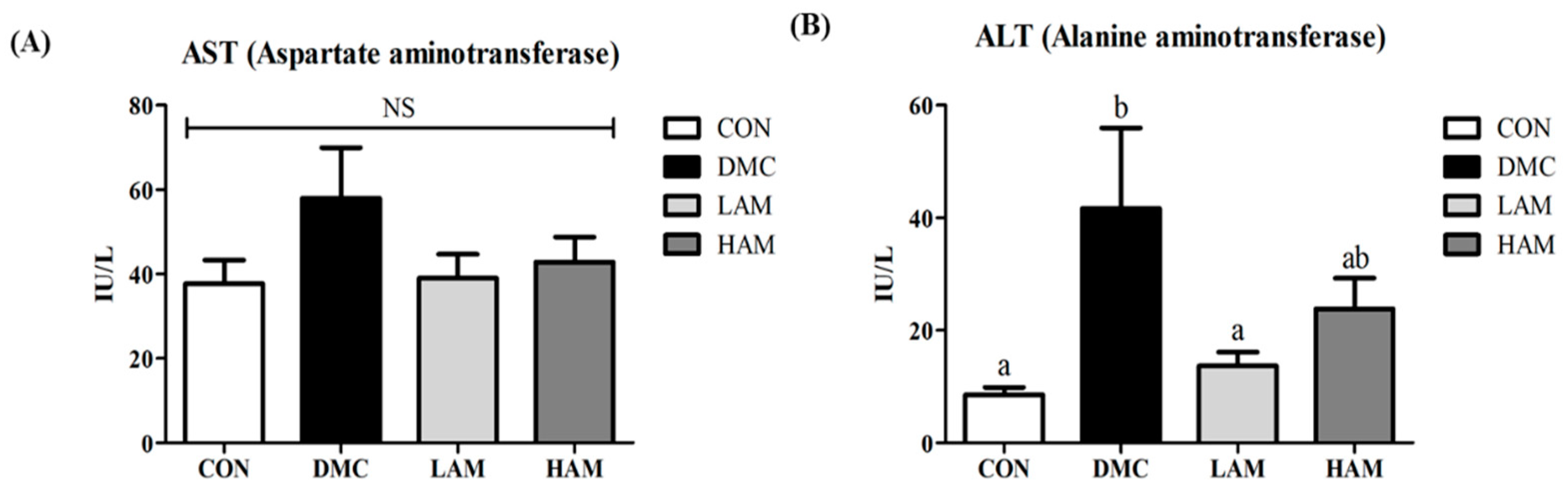
3.6. Effect of AME on Hepatic Damage in Type 2 Diabetes Mice
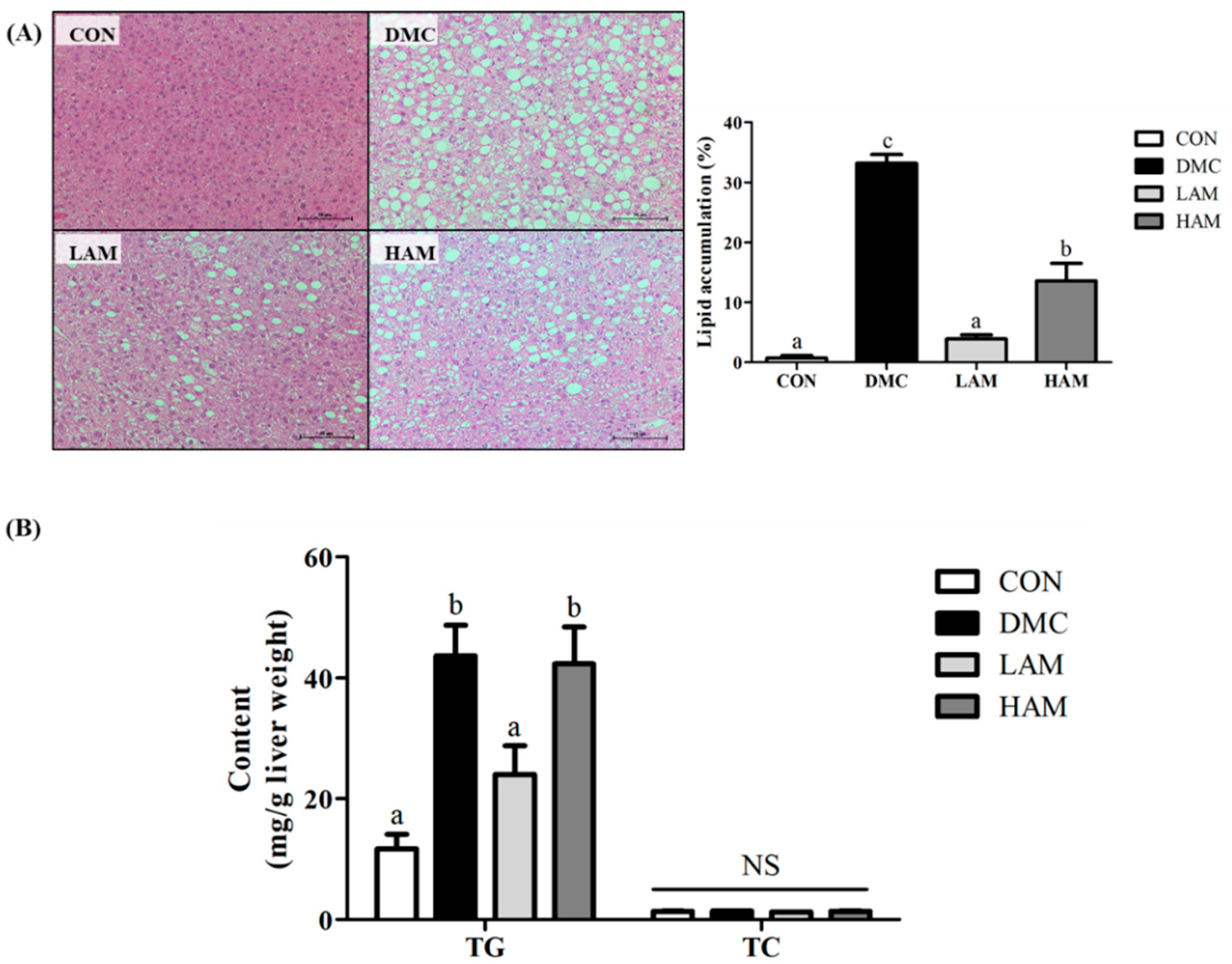
3.7. Effect of AME on Hepatic Morphology and Hepatic Triglyceride (TG) and Total Cholesterol (TC) in Type 2 Diabetes Mice
3.8. Effects of AME on Insulin Signaling in Type 2 Diabetes Mice
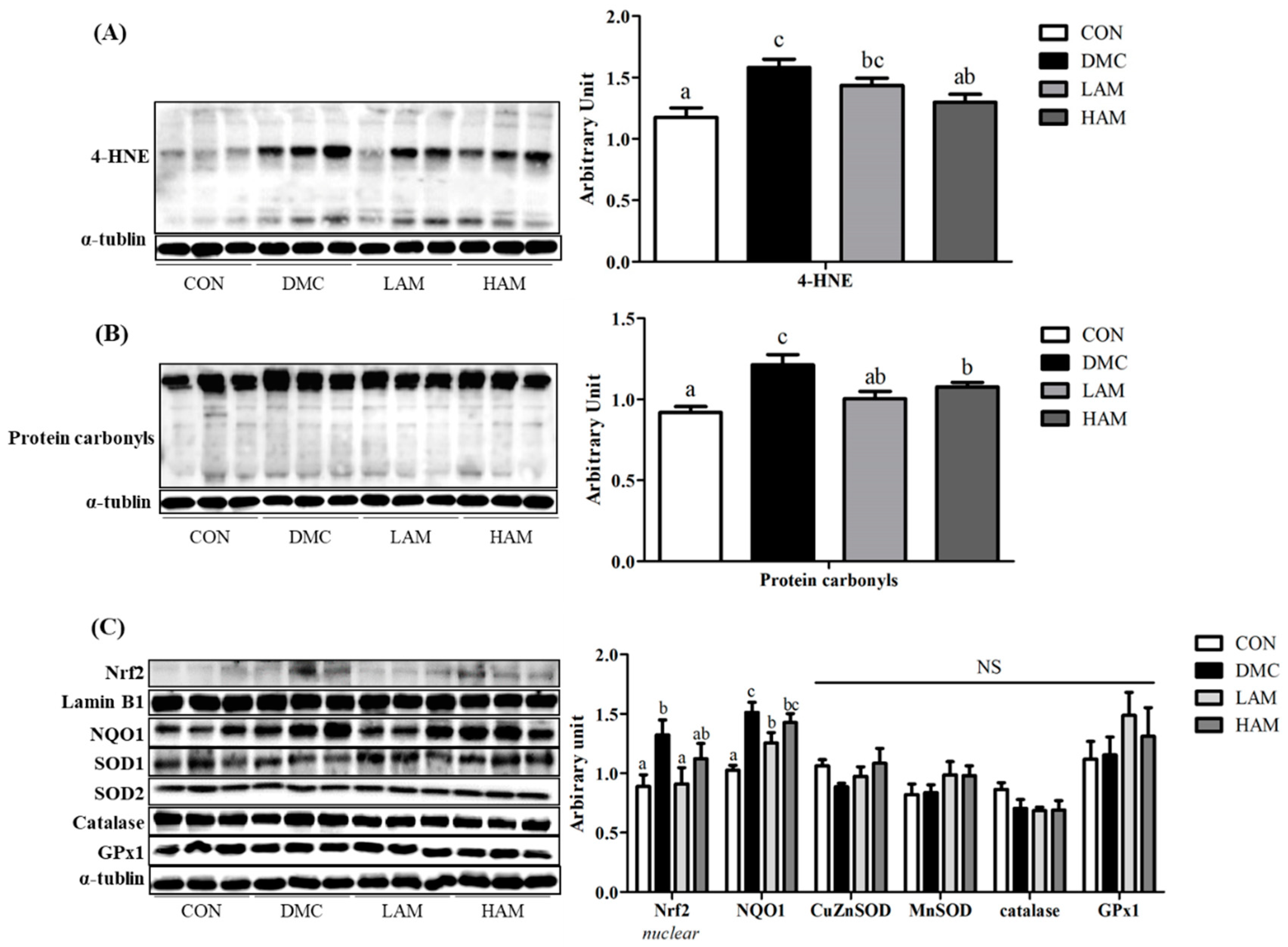
3.9. Effects of AME on Hepatic Oxidative Stress in Type 2 Diabetes Mice
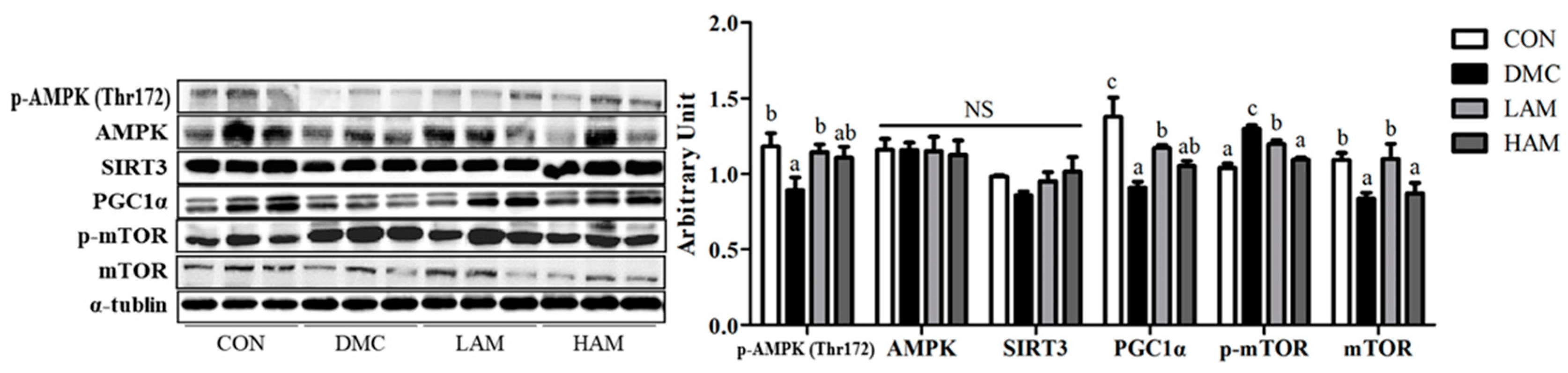
3.10. Effects of AME on Energy Metabolism in Type 2 Diabetes Mice
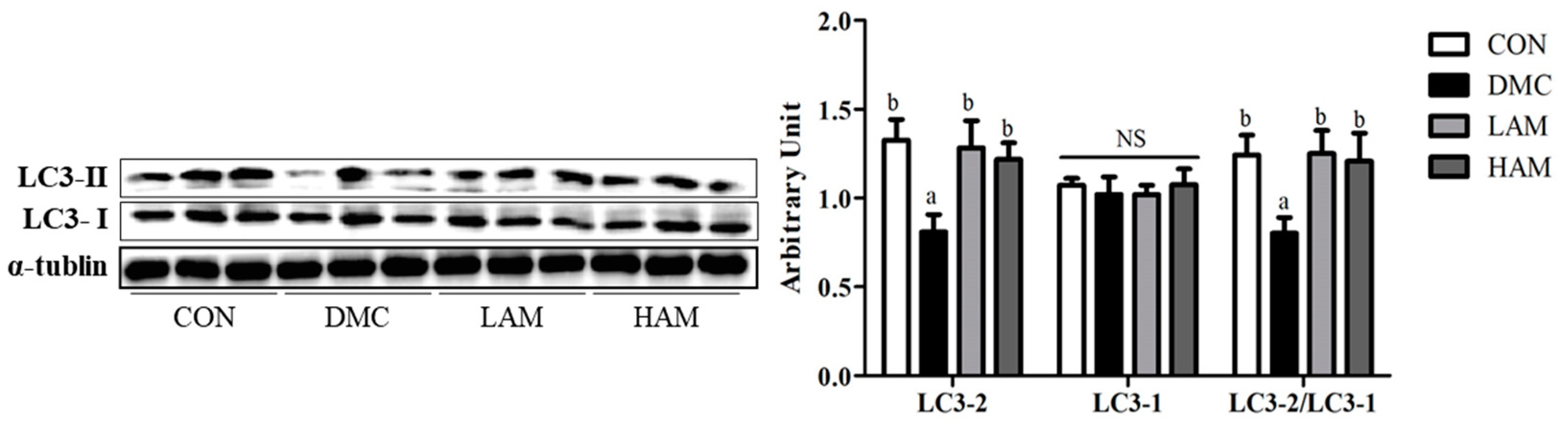
3.11. Effects of AME on Autophagy in Type 2 Diabetes Mice
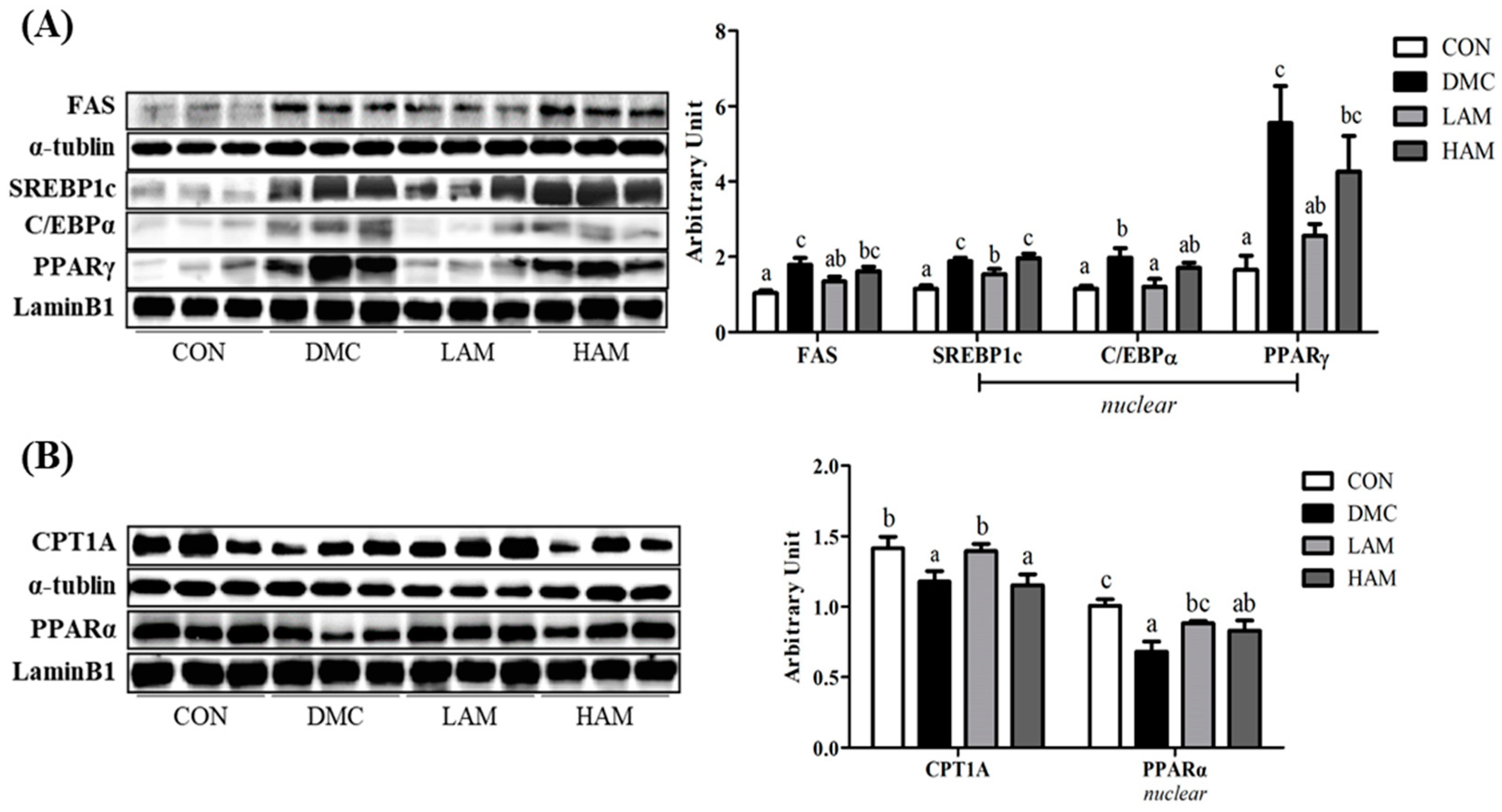
3.12. Effects of AME on Lipid Metabolism in Type 2 Diabetes Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pradeepa, R.; Mohan, V. Prevalence of type 2 diabetes and its complications in India and economic costs to the nation. Eur. J. Clin. Nutr. 2017, 71, 816–824. [Google Scholar] [CrossRef]
- Kaiser, A.B.; Zhang, N.; Der Pluijm, W. Global prevalence of type 2 diabetes over the next ten years (2018–2028). Diabetes 2018, 67, 202. [Google Scholar] [CrossRef]
- Nolan, C.J.; Damm, P.; Prentki, M. Type 2 diabetes across generations: From pathophysiology to prevention and management. Lancet 2011, 378, 169–181. [Google Scholar] [CrossRef]
- Bugianesi, E.; McCullough, A.J.; Marchesini, G. Insulin resistance: A metabolic pathway to chronic liver disease. Hepatology 2005, 42, 987–1000. [Google Scholar] [CrossRef] [PubMed]
- Guven, A.; Yavuz, O.; Cam, M. Effects of melatonin on streptozotocin-induced diabetic liver injury in rats. Acta Histochem. 2006, 108, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Palsamy, P.; Sivakumar, S.; Subramanian, S. Resveratrol attenuates hyperglycemia-mediated oxidative stress, proinflammatory cytokines and protects hepatocytes ultrastructure in streptozotocin-nicotinamide-induced experimental diabetic rats. Chem. Biol. Interact. 2010, 186, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.M.; Meki, A. Oxidative stress in streptozotocin-induced diabetic rats: Effects of garlic oil and melatonin. Comp. Biochem. Physiol.-A Mol. Integr. Physiol. 2003, 135, 539–547. [Google Scholar] [CrossRef]
- Abolfathi, A.A.; Mohajeri, D.; Rezaie, A.; Nazeri, M. Protective effects of green tea extract against hepatic tissue injury in streptozotocin-induced diabetic rats. Evid.-Based Complement. Altern. Med. 2012, 2012, 740671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boden, G.; Cheung, P.; Peter Stein, T.; Kresge, K.; Mozzoli, M. FFA cause hepatic insulin resistance by inhibiting insulin suppression of glycogenolysis. Am. J. Physiol.-Endocrinol. Metab. 2002, 283, E12–E19. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.D.; Shah, N.A.; Warrington, J.A. Combined analysis of oligonucleotide microarray data from transgenic and knockout mice identifies direct SREBP target genes. Proc. Natl. Acad. Sci. USA 2003, 100, 12027–12032. [Google Scholar] [CrossRef] [Green Version]
- Abu Elheiga, L.; Matzuk, M.M.; Abo Hashema, K.; Wakil, S.J. Continuous fatty acid oxidation and reduced fat storage in mice lacking acetyl-coa carboxylase 2. Science 2001, 291, 2613–2616. [Google Scholar] [CrossRef]
- Perry, R.J.; Samuel, V.T.; Petersen, K.F.; Shulman, G.I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature 2014, 510, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Lass, A.; Zimmermann, R.; Oberer, M.; Zechner, R. Lipolysis—A highly regulated multi-enzyme complex mediates the catabolism of cellular fat stores. Prog. Lipid. Res. 2011, 50, 14–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, A.S.; Coleman, R.A.; Kraemer, F.B. The role of lipid droplets in metabolic disease in rodents and humans. J. Clin. Invest. 2011, 121, 2102–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Kaushik, S.; Wang, Y. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Li, P.; Fu, S.; Calay, E.S.; Hotamisligil, G.S. Defective hepatic autophagy in obesity promotes ER stress and causes insulin resistance. Cell Metab. 2010, 11, 467–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.M.; Yang, S.J.; Choi, J.H.; Park, C.S. Functional effects of a pathogenic mutation in cereblon (CRBN) on the regulation of protein synthesis via the AMPK-mTOR cascade. J. Biol. Chem. 2014, 289, 23343–23352. [Google Scholar] [CrossRef] [Green Version]
- Vucicevic, L.; Misirkic, M.; Janjetovic, K. Compound C induces protective autophagy in cancer cells through AMPK inhibition-independent blockade of Akt/mTOR pathway. Autophagy 2011, 7, 40–50. [Google Scholar] [CrossRef]
- Stewart, D.; Killeen, E.; Naquin, R.; Alam, S.; Alam, J. Degradation of transcription factor Nrf2 via the ubiquitin-proteasome pathway and stabilization by cadmium. J. Biol. Chem. 2003, 278, 2396–2402. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [Green Version]
- Kapuy, O.; Papp, D.; Vellai, T.; Bánhegyi, G.; Korcsmáros, T. Systems-level feedbacks of NRF2 controlling autophagy upon oxidative stress response. Antioxidants 2018, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florence, N.T.; Benoit, M.Z.; Jonas, K. Antidiabetic and antioxidant effects of Annona muricata (Annonaceae), aqueous extract on streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2014, 151, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Justino, A.B.; Miranda, N.C.; Franco, R.R.; Martins, M.M.; Silva, N.M.; Da Espindola, F.S. Annona muricata Linn. leaf as a source of antioxidant compounds with in vitro antidiabetic and inhibitory potential against α-amylase, α-glucosidase, lipase, non-enzymatic glycation and lipid peroxidation. Biomed. Pharmacother. 2018, 100, 83–92. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, O.V.; Vieira, G.D.V.; De Pinho, J.; De Pinho, J.d.J.R.G.; Yamamoto, C.H.; Alves, M.S. Antinociceptive and anti-inflammatory activities of the ethanol extract of Annona muricata L. leaves in animal models. Int. J. Mol. Sci. 2010, 11, 2067–2078. [Google Scholar] [CrossRef] [Green Version]
- Adeyemi, D.O.; Komolafe, O.A.; Adeole, O.S.; Martins, E.M.; Kehinde, A.T. Anti hyperglycemic activities of Annona muricata (Linn). Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 62–69. [Google Scholar]
- Adeyemi, D.; Komolafe, O.; Adewole, S.; Obuotor, E.; Adenowo, T. Effects of Annona muricata (Linn) on the morphology of pancreatic islet cells of experimentally-induced diabetic wistar rats. Internet J. Altern. Med. 2012, 5, 2209164. [Google Scholar]
- Ahalya, B.; Shankar, K.R.; Kiranmayi, G.V.N. Exploration of anti-hyperglycemic and hypolipidemic activities of ethanolic extract of Annona muricata bark in alloxan induced diabetic rats. Int. J. Pharm. Sci. Rev. Res. 2014, 25, 21–27. [Google Scholar]
- Han, B.G.; Hao, C.M.; Tchekneva, E.E.; Wang, Y.Y.; Lee, C.A.; Ebrahim, B.; Harris, R.C.; Kern, T.S.; Wasserman, D.H.; Breyer, M.D.; et al. Markers of glycemic control in the mouse: Comparisons of 6-h- and overnight-fasted blood glucoses to Hb A1c. Am. J. Physiol. 2008, 4, E981–E986. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Khan, S.; Avula, B.; Lata, H.; Yang, M.H.; Elsohly, M.A.; Khan, I.A. Assessment of total phenolic and flavonoid content, antioxidant properties, and yield of aeroponically and conventionally grown leafy vegetables and fruit crops: A comparative study. Evid. Based Complement. Alternat. Med. 2014, 253875. [Google Scholar] [CrossRef]
- Bushra, S.; Farooq, A.; Muhammad, R.A.; Shahzad, A.S.C. Antioxidant potential of extracts from different agro wastes: Stabilization of corn oil. Grasas Aceites 2008, 59, 205–217. [Google Scholar]
- Lee, M.Y.; Yoo, M.S.; Whang, Y.J.; Jin, Y.J.; Hong, M.H.; Pyo, Y.H. Vitamin C, total polyphenol, flavonoid contents and antioxidant capacity of several fruit peels. Korean J. Food Sci. Technol. 2012, 44, 540–544. [Google Scholar] [CrossRef] [Green Version]
- International Diabetes Foundation. International Diabetes Federation—Facts & Figures; International Diabetes Foundation: Brussels, Belgium, 2019. [Google Scholar]
- International Diabetes Federation. Global gudelines for type 2 diabetes. In Global Guideline for Type 2 Diabetes; International Diabetes Federation: Brussels, Belgium, 2012. [Google Scholar]
- Ghorbani, A. Mechanisms of antidiabetic effects of flavonoid rutin. Biomed. Pharmacother. 2017, 96, 305–312. [Google Scholar] [CrossRef]
- Alkhalidy, H.; Moore, W.; Zhang, Y.; McMillan, R.; Wang, A.; Ali, M.; Suh, K.S.; Zhen, W.; Cheng, Z.; Jia, Z.; et al. Retracted: Small molecule kaempferol promotes insulin sensitivity and preserved pancreatic β-cell mass in middle-aged obese diabetic mice. J. Diabetes Res. 2020, 2020, 1013482. [Google Scholar]
- Chen, S.; Jiang, H.; Wu, X.; Fang, J. Therapeutic effects of quercetin on inflammation, obesity, and type 2 diabetes. Mediat. Inflamm. 2016, 2016, 9340637. [Google Scholar] [CrossRef]
- Luo, C.; Yang, H.; Tang, C.; Yao, G.; Kong, L.; He, H.; Zhou, Y. Kaempferol alleviates insulin resistance via hepatic IKK/NF-κB signal in type 2 diabetic rats. Int. Immunopharmacol. 2015, 28, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Kotronen, A.; Westerbacka, J.; Bergholm, R.; Pietiläinen, K.H.; Yki-Järvinen, H. Liver fat in the metabolic syndrome. J. Clin. Endocrinol. Metab. 2007, 92, 3490–3497. [Google Scholar] [CrossRef]
- Sovia, E.; Ratwita, W.; Wijayanti, D.; Novianty, D.R. Hypoglycemic and hypolipidemic effects of Annona muricata L. leaf ethanol extract. Int. J. Pharm. Pharm. Sci. 2017, 9, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Fujii, N.; Jessen, N.; Goodyear, L.J. AMP-activated protein kinase and the regulation of glucose transport. Am. J. Physiol-Endocrinol. Metab. 2006, 291, E867–E877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhao, X.; Ran, L. Resveratrol improves insulin resistance, glucose and lipid metabolism in patients with non-alcoholic fatty liver disease: A randomized controlled trial. Dig. Liver Dis. 2015, 47, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, S.; Mihaylova, M.M. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wang, B.; Wang, S.; Lu, H.; Wu, H.; Ding, M.; Ying, L.; Mao, Y.; Li, Y. Effect of quercetin on lipids metabolism through modulating the gut microbial and AMPK/PPAR signaling pathway in broilers. Front. Cell Dev. Biol. 2021, 9, 616219. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Pan, R.; Ding, L.; Zhang, F.; Hu, L.; Ding, B.; Zhu, L.; Xia, Y.; Dou, X. Rutin exhibits hepatoprotective effects in a mouse model of non-alcoholic fatty liver disease by reducing hepatic lipid levels and mitigating lipid-induced oxidative injuries. Int. Immunopharmacol. 2017, 49, 132–141. [Google Scholar] [CrossRef]
- Xiang, H.; Shao, M.; Lu, Y.; Wang, J.; Wu, T.; Ji, G. Kaempferol alleviates steatosis and inflammation during early non-alcoholic steatohepatitis associated with liver X receptor α-lysophosphatidylcholine acyltransferase 3 signaling pathway. Front. Pharmacol. 2021, 12, 690736. [Google Scholar] [CrossRef] [PubMed]
- Codogno, P.; Meijer, A.J. Autophagy and signaling: Their role in cell survival and cell death. Cell Death Differ. 2005, 12, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, L.; Manti, S.; D’Angelo, G. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2015, 16, 378–400. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CON | DMC | LAM | HAM | |
|---|---|---|---|---|
| Body weight (g) | ||||
| before treatment | 26.91 ± 0.61 a | 33.77 ± 1.56 b | 33.46 ± 0.92 b | 34.07 ± 1.30 b |
| after treatment | 30.71 ± 0.79 a | 38.67 ± 2.02 b | 38.49 ± 1.29 b | 40.23 ± 1.26 b |
| change | 3.80 ± 0.33 a | 4.90 ± 0.65 ab | 5.02 ± 0.76 ab | 6.16 ± 0.59 b |
| Liver weight (% BW) | 3.55 ± 0.08 a | 4.15 ± 0.21 ab | 3.78 ± 0.26 a | 4.63 ± 0.41 b |
| Food intake (g/day) | 2.72 ± 0.09 | 2.53 ± 0.11 | 2.68 ± 0.09 | 2.66 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, Y.; Lee, H.; Son, S.-Y.; Lee, C.-H.; Kim, S.-Y.; Lim, Y. Ameliorative Effect of Annona muricata (Graviola) Extract on Hyperglycemia Induced Hepatic Damage in Type 2 Diabetic Mice. Antioxidants 2021, 10, 1546. https://doi.org/10.3390/antiox10101546
Son Y, Lee H, Son S-Y, Lee C-H, Kim S-Y, Lim Y. Ameliorative Effect of Annona muricata (Graviola) Extract on Hyperglycemia Induced Hepatic Damage in Type 2 Diabetic Mice. Antioxidants. 2021; 10(10):1546. https://doi.org/10.3390/antiox10101546
Chicago/Turabian StyleSon, Yiseul, Heaji Lee, Su-Young Son, Choong-Hwan Lee, Sun-Yeou Kim, and Yunsook Lim. 2021. "Ameliorative Effect of Annona muricata (Graviola) Extract on Hyperglycemia Induced Hepatic Damage in Type 2 Diabetic Mice" Antioxidants 10, no. 10: 1546. https://doi.org/10.3390/antiox10101546
APA StyleSon, Y., Lee, H., Son, S.-Y., Lee, C.-H., Kim, S.-Y., & Lim, Y. (2021). Ameliorative Effect of Annona muricata (Graviola) Extract on Hyperglycemia Induced Hepatic Damage in Type 2 Diabetic Mice. Antioxidants, 10(10), 1546. https://doi.org/10.3390/antiox10101546