Potential Role of the Mitochondria for the Dermatological Treatment of Papillon-Lefèvre


 ,
,  , ,
, , 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statements
2.2. Reagents
2.3. Fibroblast Cultures
2.4. Antioxidant Enzyme Activity
2.5. Measurement of CoQ10 Levels
2.6. Western Blotting
2.7. Measurement of Citrate Synthase Activity
2.8. Mitochondrial ROS Production
2.9. Oxygen Consumption Rate (OCR)
2.10. Electron Microscopy
2.11. Proliferation Rate
2.12. PCR Amplification and Sequencing
2.13. Statistical Analysis
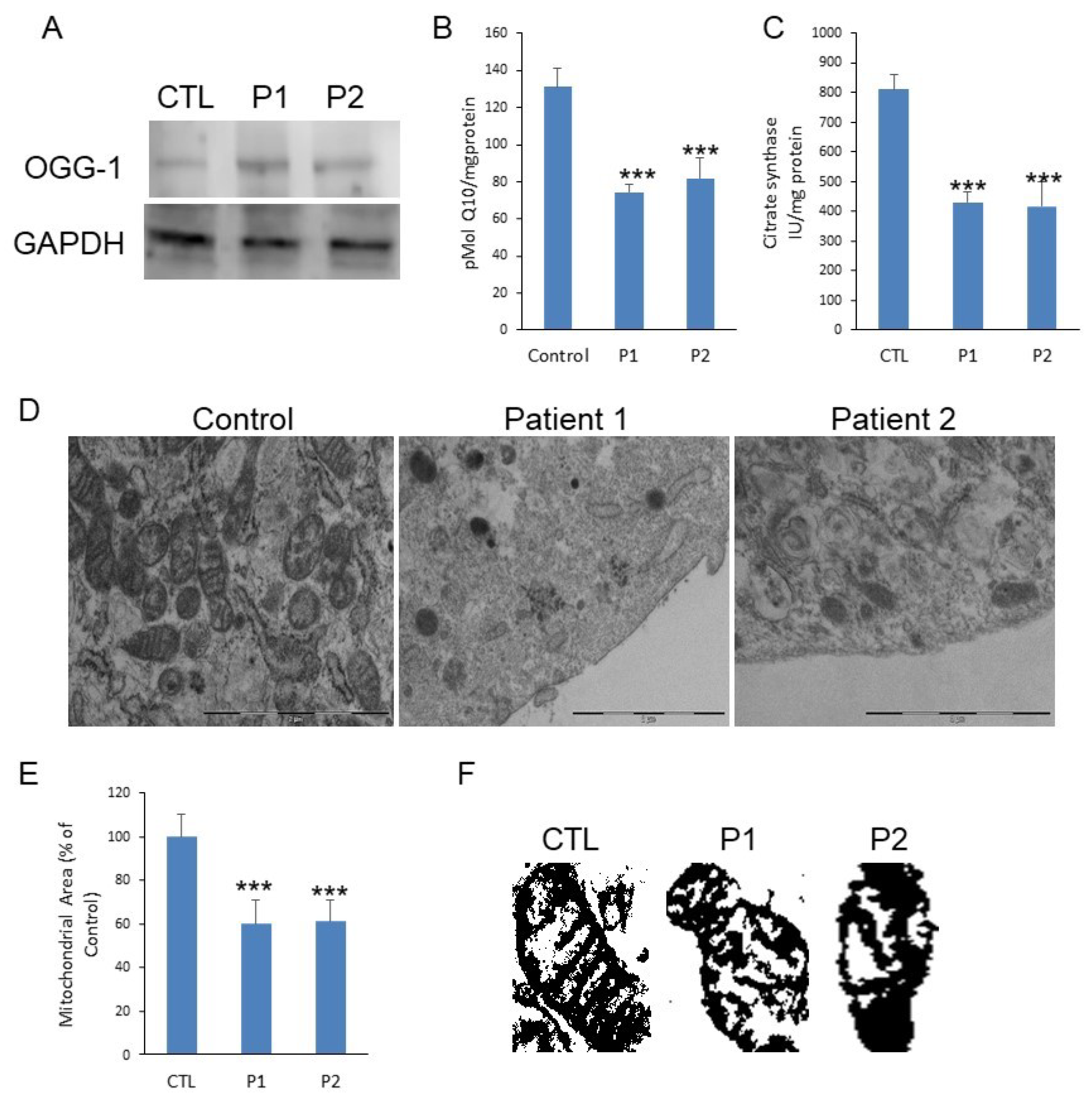
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giannetti, L.; Apponi, R.; Dello Diago, A.M.; Jafferany, M.; Goldust, M.; Sadoughifar, R. Papillon-Lefèvre syndrome: Oral aspects and treatment. Dermatol. Ther. 2020, 33, e13336. [Google Scholar] [CrossRef] [PubMed]
- Iqtadar, S.; Mumtaz, S.U.; Abaidullah, S. Papillon-Lèfevre syndrome with palmoplantar keratoderma and periodontitis, a rare cause of pyrexia of unknown origin: A case report. J. Med. Case Rep. 2015, 9, 288. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hewitt, C.; McCormick, D.; Linden, G.; Turk, D.; Stern, I.; Wallace, I.; Southern, L.; Zhang, L.; Howard, R.; Bullon, P.; et al. The role of cathepsin C in Papillon-Lefèvre syndrome, prepubertal periodontitis, and aggressive periodontitis. Hum. Mutat. 2004, 23, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.D.; Torres, V.J.; Cadwell, K. Autophagy and microbial pathogenesis. Cell Death Differ. 2020, 27, 872–886. [Google Scholar] [CrossRef] [PubMed]
- Bullón, P.; Castejón-Vega, B.; Román-Malo, L.; Jimenez-Guerrero, M.P.; Cotán, D.; Forbes-Hernandez, T.Y.; Varela-López, A.; Pérez-Pulido, A.J.; Giampieri, F.; Quiles, J.L.; et al. Autophagic dysfunction in patients with Papillon-Lefèvre syndrome is restored by recombinant cathepsin C treatment. J. Allergy Clin. Immunol. 2018, 142, 1131–1143.e7. [Google Scholar] [CrossRef] [PubMed]
- Battino, M.; Ferreiro, M.S.; Quiles, J.L.; Bompadre, S.; Leone, L.; Bullon, P. Alterations in the oxidation products, antioxidant markers, antioxidant capacity and lipid patterns in plasma of patients affected by Papillon-Lefèvre syndrome. Free Radic. Res. 2003, 37, 603–609. [Google Scholar] [CrossRef]
- Hahn, J.; Schauer, C.; Czegley, C.; Kling, L.; Petru, L.; Schmid, B.; Weidner, D.; Reinwald, C.; Biermann, M.H.C.; Blunder, S.; et al. Aggregated neutrophil extracellular traps resolve inflammation by proteolysis of cytokines and chemokines and protection from antiproteases. FASEB J. 2019, 33, 1401–1414. [Google Scholar] [CrossRef]
- Willems, P.H.; Rossignol, R.; Dieteren, C.E.; Murphy, M.P.; Koopman, W.J. Redox Homeostasis and Mitochondrial Dynamics. Cell Metab. 2015, 22, 207–218. [Google Scholar] [CrossRef]
- Bullon, P.; Newman, H.N.; Battino, M. Obesity, diabetes mellitus, atherosclerosis and chronic periodontitis: A shared pathology via oxidative stress and mitochondrial dysfunction? Periodontol. 2000 2014, 64, 139–153. [Google Scholar] [CrossRef]
- Feichtinger, R.G.; Sperl, W.; Bauer, J.W.; Kofler, B. Mitochondrial dysfunction: A neglected component of skin diseases. Exp. Dermatol. 2014, 23, 607–614. [Google Scholar] [CrossRef]
- Bonfield, J.K.; Rada, C.; Staden, R. Automated detection of point mutations using fluorescent sequence trace subtraction. Nucleic Acids Res. 1998, 26, 3404–3409. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Roberts, H.; White, P.; Dias, I.; McKaig, S.; Veeramachaneni, R.; Thakker, N.; Grant, M.; Chapple, I. Characterization of neutrophil function in Papillon-Lefèvre syndrome. J. Leukoc. Biol. 2016, 100, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Ledderose, C.; Seier, T.; Graf, A.F.; Brix, B.; Chong, E.; Junger, W.F. Mitochondria regulate neutrophil activation by generating ATP for autocrine purinergic signaling. J. Biol. Chem. 2014, 289, 26794–26803. [Google Scholar] [CrossRef] [PubMed]
- Haas, R.H.; Parikh, S.; Falk, M.J.; Saneto, R.P.; Wolf, N.I.; Darin, N.; Wong, L.J.; Cohen, B.H.; Naviaux, R.K. The in-depth evaluation of suspected mitochondrial disease. Mol. Genet. Metab. 2008, 94, 16–37. [Google Scholar] [CrossRef]
- Cha, M.Y.; Kim, D.K.; Mook-Jung, I. The role of mitochondrial DNA mutation on neurodegenerative diseases. Exp. Mol. Med. 2015, 47, e150. [Google Scholar] [CrossRef]
- Zhong, Z.; Liang, S.; Sanchez-Lopez, E.; He, F.; Shalapour, S.; Lin, X.J.; Wong, J.; Ding, S.; Seki, E.; Schnabl, B.; et al. New mitochondrial DNA synthesis enables NLRP3 inflammasome activation. Nature 2018, 560, 198–203. [Google Scholar] [CrossRef]
- Collins, L.V.; Hajizadeh, S.; Holme, E.; Jonsson, I.M.; Tarkowski, A. Endogenously oxidized mitochondrial DNA induces in vivo and in vitro inflammatory responses. J. Leukoc. Biol. 2004, 75, 995–1000. [Google Scholar] [CrossRef]
- Rabinowitz, J.D.; White, E. Autophagy and metabolism. Science 2010, 330, 1344–1348. [Google Scholar] [CrossRef]
- Cornelius, N.; Byron, C.; Hargreaves, I.; Guerra, P.F.; Furdek, A.K.; Land, J.; Radford, W.W.; Frerman, F.; Corydon, T.J.; Gregersen, N.; et al. Secondary coenzyme Q10 deficiency and oxidative stress in cultured fibroblasts from patients with riboflavin responsive multiple Acyl-CoA dehydrogenation deficiency. Hum. Mol. Genet. 2013, 22, 3819–3827. [Google Scholar] [CrossRef]
- Bräsen, J.H.; Koenig, K.; Bach, H.; Kontush, A.; Heinle, H.; Witting, P.K.; Ylä-Herttuala, S.; Stocker, R.; Beisiegel, U. Comparison of the effects of alpha-tocopherol, ubiquinone-10 and probucol at therapeutic doses on atherosclerosis in WHHL rabbits. Atherosclerosis 2002, 163, 249–259. [Google Scholar] [CrossRef]
- Hernández-Camacho, J.D.; Bernier, M.; López-Lluch, G.; Navas, P. Coenzyme Q10 Supplementation in Aging and Disease. Front. Physiol. 2018, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Casado, M.E.; Quiles, J.L.; Barriocanal-Casado, E.; González-García, P.; Battino, M.; López, L.C.; Varela-López, A. The Paradox of Coenzyme Q10 in Aging. Nutrients 2019, 11, 2221. [Google Scholar] [CrossRef] [PubMed]
- Kharaeva, Z.; Gostova, E.; De Luca, C.; Raskovic, D.; Korkina, L. Clinical and biochemical effects of coenzyme Q(10), vitamin E, and selenium supplementation to psoriasis patients. Nutrition 2009, 25, 295–302. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castejón-Vega, B.; Battino, M.; Quiles, J.L.; Bullon, B.; Cordero, M.D.; Bullón, P. Potential Role of the Mitochondria for the Dermatological Treatment of Papillon-Lefèvre. Antioxidants 2021, 10, 95. https://doi.org/10.3390/antiox10010095
Castejón-Vega B, Battino M, Quiles JL, Bullon B, Cordero MD, Bullón P. Potential Role of the Mitochondria for the Dermatological Treatment of Papillon-Lefèvre. Antioxidants. 2021; 10(1):95. https://doi.org/10.3390/antiox10010095
Chicago/Turabian StyleCastejón-Vega, Beatriz, Maurizio Battino, José L. Quiles, Beatriz Bullon, Mario D. Cordero, and Pedro Bullón. 2021. "Potential Role of the Mitochondria for the Dermatological Treatment of Papillon-Lefèvre" Antioxidants 10, no. 1: 95. https://doi.org/10.3390/antiox10010095
APA StyleCastejón-Vega, B., Battino, M., Quiles, J. L., Bullon, B., Cordero, M. D., & Bullón, P. (2021). Potential Role of the Mitochondria for the Dermatological Treatment of Papillon-Lefèvre. Antioxidants, 10(1), 95. https://doi.org/10.3390/antiox10010095