Relation of Serum Plasmalogens and APOE Genotype to Cognition and Dementia in Older Persons in a Cross-Sectional Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Clinical Evaluation
2.3. Serum Biomarker Analyses
2.4. APOE Genotyping
2.5. Statistical Analyses
3. Results
3.1. Demographics
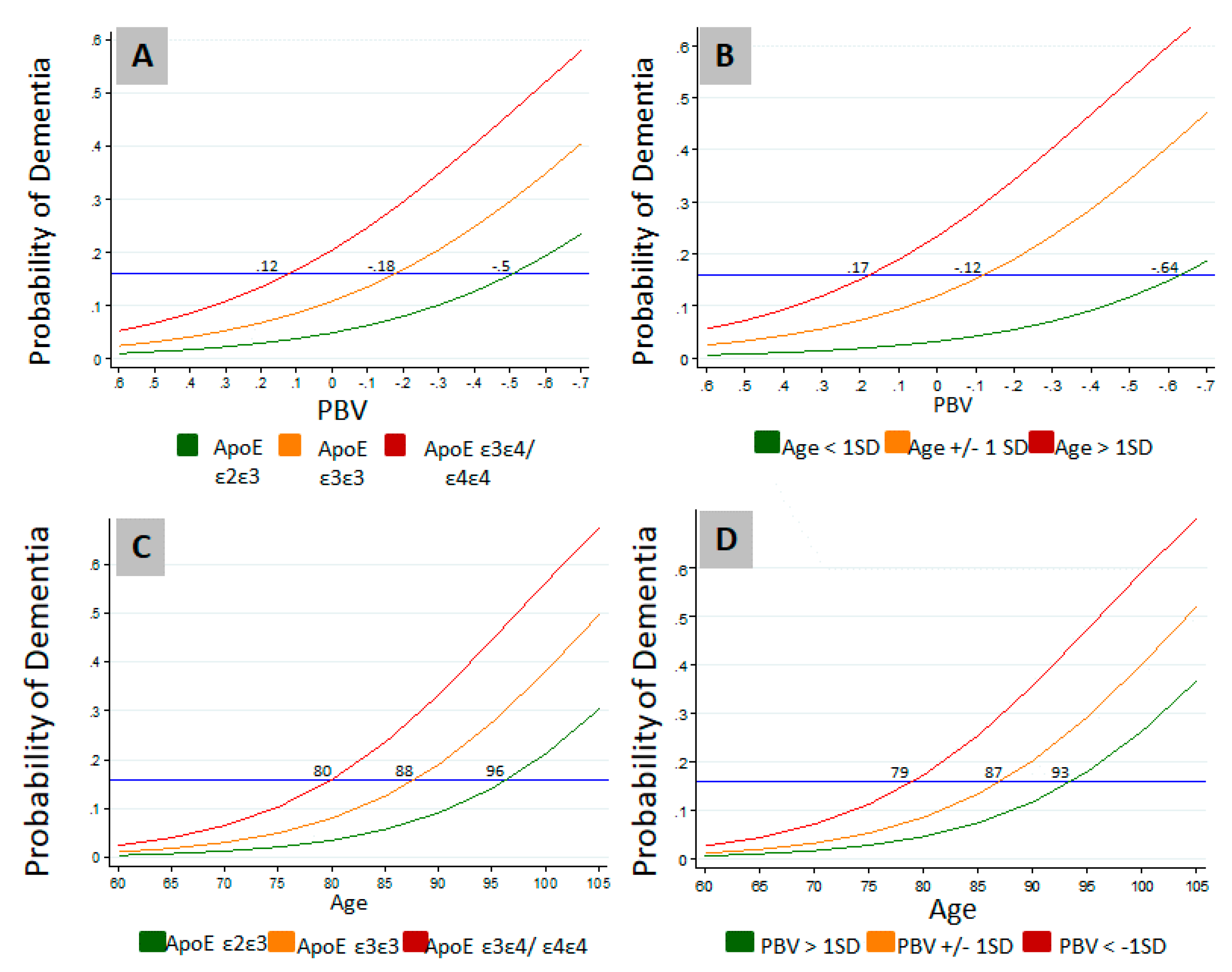
3.2. Relation of PBV and APOE to AD and Level of Cognition
3.3. Study Variable Associations on Serum Lipids
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pitas, R.E.; Boyles, J.K.; Lee, S.H.; Hui, D.; Weisgraber, K.H. Lipoproteins and their receptors in the central nervous system. Characterization of the lipoproteins in cerebrospinal fluid and identification of apolipoprotein B,E(LDL) receptors in the brain. J. Biol. Chem. 1987, 262, 14352–14360. [Google Scholar] [PubMed]
- Corder, E.H.; Saunders, A.M.; Strittmatter, W.J.; Schmechel, D.E.; Gaskell, P.C.; Small, G.W.; Roses, A.D.; Haines, J.L.; Pericak-Vance, M.A. Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer’s disease in late onset families. Science 1993, 261, 921–923. [Google Scholar] [CrossRef] [PubMed]
- Ashford, J.W. APOE genotype effects on Alzheimer’s disease onset and epidemiology. J. Mol. Neurosci. 2004, 23, 157–165. [Google Scholar] [CrossRef]
- Hauser, P.S.; Narayanaswami, V.; Ryan, R.O. Apolipoprotein E: from lipid transport to neurobiology. Prog. Lipid Res. 2011, 50, 62–74. [Google Scholar] [CrossRef]
- Kim, J.; Basak, J.M.; Holtzman, D.M. The role of apolipoprotein E in Alzheimer’s disease. Neuron 2009, 63, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Goodenowe, D.B.; Cook, L.L.; Liu, J.; Lu, Y.; Jayasinghe, D.A.; Ahiahonu, P.W.; Heath, D.; Yamazaki, Y.; Flax, J.; Krenitsky, K.F.; et al. Peripheral ethanolamine plasmalogen deficiency: a logical causative factor in Alzheimer’s disease and dementia. J. Lipid Res. 2007, 48, 2485–2498. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L.; Mankidy, R.; Ritchie, S.; Heath, D.; Wood, J.A.; Flax, J.; Goodenowe, D.B. Circulating plasmalogen levels and Alzheimer Disease Assessment Scale–Cognitive scores in Alzheimer patients. J. Psychiatry Neurosci. 2010, 35, 59–62. [Google Scholar] [CrossRef]
- Chang, C.-Y.; Ke, D.-S.; Chen, J.-Y. Essential fatty acids and human brain. Acta Neurol. Taiwanica 2009, 18, 231–241. [Google Scholar]
- Farooqui, A.A.; Farooqui, T.; Horrocks, L.A. Metabolism and Functions of Bioactive Ether Lipids in the Brain; Springer Nature: New York, NY, USA, 2008. [Google Scholar]
- Rouser, G.; Yamamoto, A. Curvilinear regression course of human brain lipid composition changes with age. Lipids 1968, 3, 284–287. [Google Scholar] [CrossRef]
- Ginsberg, L.; Rafique, S.; Xuereb, J.H.; Rapoport, S.I.; Gershfeld, N.L. Disease and anatomic specificity of ethanolamine plasmalogen deficiency in Alzheimer’s disease brain. Brain Res. 1995, 698, 223–226. [Google Scholar] [CrossRef]
- Han, X.; Holtzman, D.M.; McKeel, D.W., Jr. Plasmalogen deficiency in early Alzheimer’s disease subjects and in animal models: molecular characterization using electrospray ionization mass spectrometry. J. Neurochem. 2001, 77, 1168–1180. [Google Scholar] [CrossRef]
- Guan, Z.; Wang, Y.; Cairns, N.J.; Lantos, P.L.; Dallner, G.; Sindelar, P.J. Decrease and Structural Modifications of Phosphatidylethanolamine Plasmalogen in the Brain with Alzheimer Disease. J. Neuropathol. Exp. Neurol. 1999, 58, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, A.A.; Rapoport, S.I.; Horrocks, L.A. “Membrane Phospholipid Alterations in Alzheimer’s Disease: Deficiency of Ethanolamine Plasmalogens. Neurochem. Res. 1997, 22, 523–527. [Google Scholar] [CrossRef]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta 2012, 1822, 1442–1452. [Google Scholar] [CrossRef]
- Bennett, D.A.; Schneider, J.A.; Aggarwal, N.T.; Arvanitakis, Z.; Shah, R.C.; Kelly, J.F.; Fox, J.H.; Cochran, E.J.; Arends, D.; Treinkman, A.D.; et al. Decision rules guiding the clinical diagnosis of Alzheimer’s disease in two community-based cohort studies compared to standard practice in a clinic-based cohort study. Neuroepidemiology 2006, 27, 169–176. [Google Scholar] [CrossRef]
- Knopman, D.S.; DeKosky, S.T.; Cummings, J.L.; Chui, H.; Corey-Bloom, J.; Relkin, N.; Small, G.W.; Miller, B.; Stevens, J.C. Practice Parameter: Diagnosis of Dementia (an Evidence-Based Review). Report of the Quality Standards Subcommittee of the American Academy of Neurology. Neurology 2001, 56, 1143–1153. [Google Scholar] [CrossRef] [PubMed]
- Poirier, J.; Miron, J.; Picard, C.; Gormley, P.; Theroux, L.; Breitner, J.; Dea, D. Apolipoprotein E and lipid homeostasis in the etiology and treatment of sporadic Alzheimer’s disease. Neurobiol. Aging 2014, 35, 3–10. [Google Scholar] [CrossRef]
- Leoni, V.; Solomon, A.; Kivipelto, M. Links between ApoE, brain cholesterol metabolism, tau and amyloid beta-peptide in patients with cognitive impairment. Biochem. Soc. Trans. 2010, 38, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Michikawa, M.; Fan, Q.W.; Isobe, I.; Yanagisawa, K. Apolipoprotein E exhibits isoform-specific promotion of lipid efflux from astrocytes and neurons in culture. J. Neurochem. 2000, 74, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Munn, N.J.; Arnio, E.; Liu, D.; Zoeller, R.A.; Liscum, L. Deficiency in ethanolamine plasmalogen leads to altered cholesterol transport. J. Lipid Res. 2003, 44, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Mankidy, R.; Ahiahonu, P.W.; Ma, H.; Jayasinghe, D.; Ritchie, S.A.; Khan, M.A.; Su-Myat, K.K.; Wood, P.L.; Goodenowe, D.B. Membrane plasmalogen composition and cellular cholesterol regulation: A structure activity study. Lipids Health Dis. 2010, 9, 9–62. [Google Scholar] [CrossRef]
- Mandel, H.; Sharf, R.; Berant, M.; Wanders, R.J.; Vreken, P.; Aviram, M. Plasmalogen phospholipids are involved in HDL-mediated cholesterol efflux: Insights from investigations with plasmalogen-deficient cells. Biochem. Biophys. Res. Comm. 1998, 250, 369–373. [Google Scholar] [CrossRef]
- Oikawa, N.; Hatsuta, H.; Murayama, S.; Suzuki, A.; Yanagisawa, K. Influence of APOE genotype and the presence of Alzheimer’s pathology on synaptic membrane lipids of human brains. J. Neurosci. Res. 2014, 92, 641–650. [Google Scholar] [CrossRef]
- Igbavboa, U.; Eckert, G.P.; Malo, T.M.; Studniski, A.E.; Johnson, L.N.; Yamamoto, N.; Kobayashi, M.; Fujita, S.C.; Appel, T.R.; Muller, W.E.; et al. Murine synaptosomal lipid raft protein and lipid composition are altered by expression of human Apoe 3 and 4 and by increasing age. J. Neurol. Sci. 2005, 229–230, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Olsen, R.H.; Merkens, L.S.; Debarber, A.; Steiner, R.D.; Sullivan, P.M.; Maeda, N.; Raber, J. Apolipoprotein E-low density lipoprotein receptor interaction affects spatial memory retention and brain ApoE levels in an isoform-dependent manner. Neurobiol. Dis. 2014, 64, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.A.; Schneider, J.A.; Arvanitakis, Z.; Wilson, R.S. Overview and findings from the religious orders study. Curr. Alzheimer. Res. 2012, 9, 628–645. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.A.; Schneider, J.A.; Buchman, A.S.; Barnes, L.L.; Boyle, P.A.; Wilson, R.S. Overview and findings from the rush Memory and Aging Project. Curr. Alzheimer. Res. 2012, 9, 646–663. [Google Scholar] [CrossRef]
- Bennett, D.A.; Schneider, J.A.; Buchman, A.S.; Mendes de Leon, C.; Bienias, J.L.; Wilson, R.S. The Rush Memory and Aging Project: Study design and baseline characteristics of the study cohort. Neuroepidemiology 2005, 25, 163–175. [Google Scholar] [CrossRef]
- Wilson, R.S.; Leurgans, S.E.; Boyle, P.A.; Bennett, D.A. Cognitive decline in prodromal Alzheimer disease and mild cognitive impairment. Arch. Neurol. 2011, 68, 351–356. [Google Scholar] [CrossRef]
- Bennett, D.A.; Wilson, R.S.; Schneider, J.A.; Evans, D.A.; Beckett, L.A.; Aggarwal, N.T.; Barnes, L.L.; Fox, J.H.; Bach, J. Natural history of mild cognitive impairment in older persons. Neurology 2002, 59, 198–205. [Google Scholar] [CrossRef]
- Hagve, T.A.; Christophersen, B.O. Evidence for peroxisomal retroconversion of adrenic acid (22:4(n-6)) and docosahexaenoic acids (22:6(n-3)) in isolated liver cells. Biochim. Biophys. Acta 1986, 875, 165–173. [Google Scholar] [CrossRef]
- Gronn, M.; Christensen, E.; Hagve, T.A.; Christophersen, B.O. Peroxisomal retroconversion of docosahexaenoic acid (22:6(n-3)) to eicosapentaenoic acid (20:5(n-3)) studied in isolated rat liver cells. Biochim. Biophys. Acta 1991, 1081, 85–91. [Google Scholar] [CrossRef]
- Moore, S.A.; Hurt, E.; Yoder, E.; Sprecher, H.; Spector, A.A. Docosahexaenoic acid synthesis in human skin fibroblasts involves peroxisomal retroconversion of tetracosahexaenoic acid. J. Lipid Res. 1995, 36, 2433–2443. [Google Scholar] [PubMed]
- Olazaran, J.; Gil-de-Gomez, L.; Rodriguez-Martin, A.; Valenti-Soler, M.; Frades-Payo, B.; Marin-Munoz, J.; Antunez, C.; Frank-Garcia, A.; Acedo-Jimenez, C.; Morlan-Gracia, L.; et al. A blood-based, 7-metabolite signature for the early diagnosis of Alzheimer’s disease. J. Alzheimers Dis. 2015, 45, 1157–1173. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, L.; Evans, J.E.; Emmerich, T.; Crynen, G.; Shackleton, B.; Keegan, A.P.; Luis, C.; Tai, L.; LaDu, M.J.; Mullan, M.; et al. APOE epsilon4 specific imbalance of arachidonic acid and docosahexaenoic acid in serum phospholipids identifies individuals with preclinical Mild Cognitive Impairment/Alzheimer’s Disease. Aging 2017, 9, 964–985. [Google Scholar] [CrossRef]
- Nishimukai, M.; Maeba, R.; Yamazaki, Y.; Nezu, T.; Sakurai, T.; Takahashi, Y.; Hui, S.P.; Chiba, H.; Okazaki, T.; Hara, H. Serum choline plasmalogens, particularly those with oleic acid in sn-2, are associated with proatherogenic state. J. Lipid Res. 2014, 55, 956–965. [Google Scholar] [CrossRef]
- Dorninger, F.; Brodde, A.; Braverman, N.E.; Moser, A.B.; Just, W.W.; Forss-Petter, S.; Brugger, B.; Berger, J. Homeostasis of phospholipids—The level of phosphatidylethanolamine tightly adapts to changes in ethanolamine plasmalogens. Biochim. Biophys. Acta 2015, 1851, 117–128. [Google Scholar] [CrossRef]
- Koch, M.; Jensen, M.K. HDL-cholesterol and apolipoproteins in relation to dementia. Curr. Opin. Lipidol. 2016, 27, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Anstey, K.J.; Ashby-Mitchell, K.; Peters, R. Updating the Evidence on the Association between Serum Cholesterol and Risk of Late-Life Dementia: Review and Meta-Analysis. J. Alzheimers Dis. 2017, 56, 215–228. [Google Scholar] [CrossRef]
- Getz, G.S.; Reardon, C.A. Apoprotein E as a lipid transport and signaling protein in the blood, liver, and artery wall. J. Lipid Res. 2009, 50 Suppl, S156–S161. [Google Scholar] [CrossRef]
- Raber, J.; Huang, Y.; Ashford, J.W. ApoE genotype accounts for the vast majority of AD risk and AD pathology. Neurobiol. Aging 2004, 25, 641–650. [Google Scholar] [CrossRef]
- Kim, H.J.; Ye, B.S.; Yoon, C.W.; Cho, H.; Noh, Y.; Kim, G.H.; Choi, Y.S.; Kim, J.H.; Jeon, S.; Lee, J.M.; et al. Effects of APOE epsilon4 on brain amyloid, lacunar infarcts, and white matter lesions: A study among patients with subcortical vascular cognitive impairment. Neurobiol. Aging 2013, 34, 2482–2487. [Google Scholar] [CrossRef]
- Morris, J.C.; Roe, C.M.; Xiong, C.; Fagan, A.M.; Goate, A.M.; Holtzman, D.M.; Mintun, M.A. APOE predicts amyloid-beta but not tau Alzheimer pathology in cognitively normal aging. Ann. Neurol. 2010, 67, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Caselli, R.J.; Walker, D.; Sue, L.; Sabbagh, M.; Beach, T. Amyloid load in nondemented brains correlates with APOE e4. Neurosci. Lett. 2010, 473, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.A.; Wilson, R.S.; Schneider, J.A.; Evans, D.A.; Aggarwal, N.T.; Arnold, S.E.; Cochran, E.J.; Berry-Kravis, E.; Bienias, J.L. Apolipoprotein E epsilon4 allele, AD pathology, and the clinical expression of Alzheimer’s disease. Neurology 2003, 60, 246–252. [Google Scholar] [CrossRef]
- Wood, P.L.; Khan, A.M.; Mankidy, R.; Smith, T.; Goodenowe, D. Plasmalogen Deficit: A New and Testable Hypothesis for the Etiology of Alzheimer’s Disease, Alzheimer’s Disease Pathogenesis—Core Concepts, Shifting Paradigms and Therapeutic Targets, Suzanne De La Monte, IntechOpen. Available online: https://www.intechopen.com/books/alzheimer-s-disease-pathogenesis-core-concepts-shifting-paradigms-and-therapeutic-targets/plasmalogen-deficit-a-new-and-testable-hypothesis-for-the-etiology-of-alzheimer-s-disease (accessed on 22 April 2019).
- Castello, M.A.; Jeppson, J.D.; Soriano, S. Moving beyond anti-amyloid therapy for the prevention and treatment of Alzheimer’s disease. BMC Neurol. 2014, 14, 169. [Google Scholar] [CrossRef]
- Cutler, R.G.; Kelly, J.; Storie, K.; Pedersen, W.A.; Tammara, A.; Hatanpaa, K.; Troncoso, J.C.; Mattson, M.P. Involvement of oxidative stress-induced abnormalities in ceramide and cholesterol metabolism in brain aging and Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2004, 101, 2070–2075. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Tozuka, M.; Nakabayashi, T.; Sugano, M.; Hidaka, H.; Kondo, Y.; Katsuyama, T. Apolipoprotein E in cerebrospinal fluid: relation to phenotype and plasma apolipoprotein E concentrations. Clin. Chem. 1999, 45, 497–504. [Google Scholar]
- Mulder, M.; Ravid, R.; Swaab, D.F.; de Kloet, E.R.; Haasdijk, E.D.; Julk, J.; van der Boom, J.J.; Havekes, L.M. Reduced levels of cholesterol, phospholipids, and fatty acids in cerebrospinal fluid of Alzheimer disease patients are not related to apolipoprotein E4. Alzheimer Dis. Assoc. Disord. 1998, 12, 198–203. [Google Scholar] [CrossRef]
- Ghribi, O.; Larsen, B.; Schrag, M.; Herman, M.M. High cholesterol content in neurons increases BACE, beta-amyloid, and phosphorylated tau levels in rabbit hippocampus. Exp. Neurol. 2006, 200, 460–467. [Google Scholar] [CrossRef]
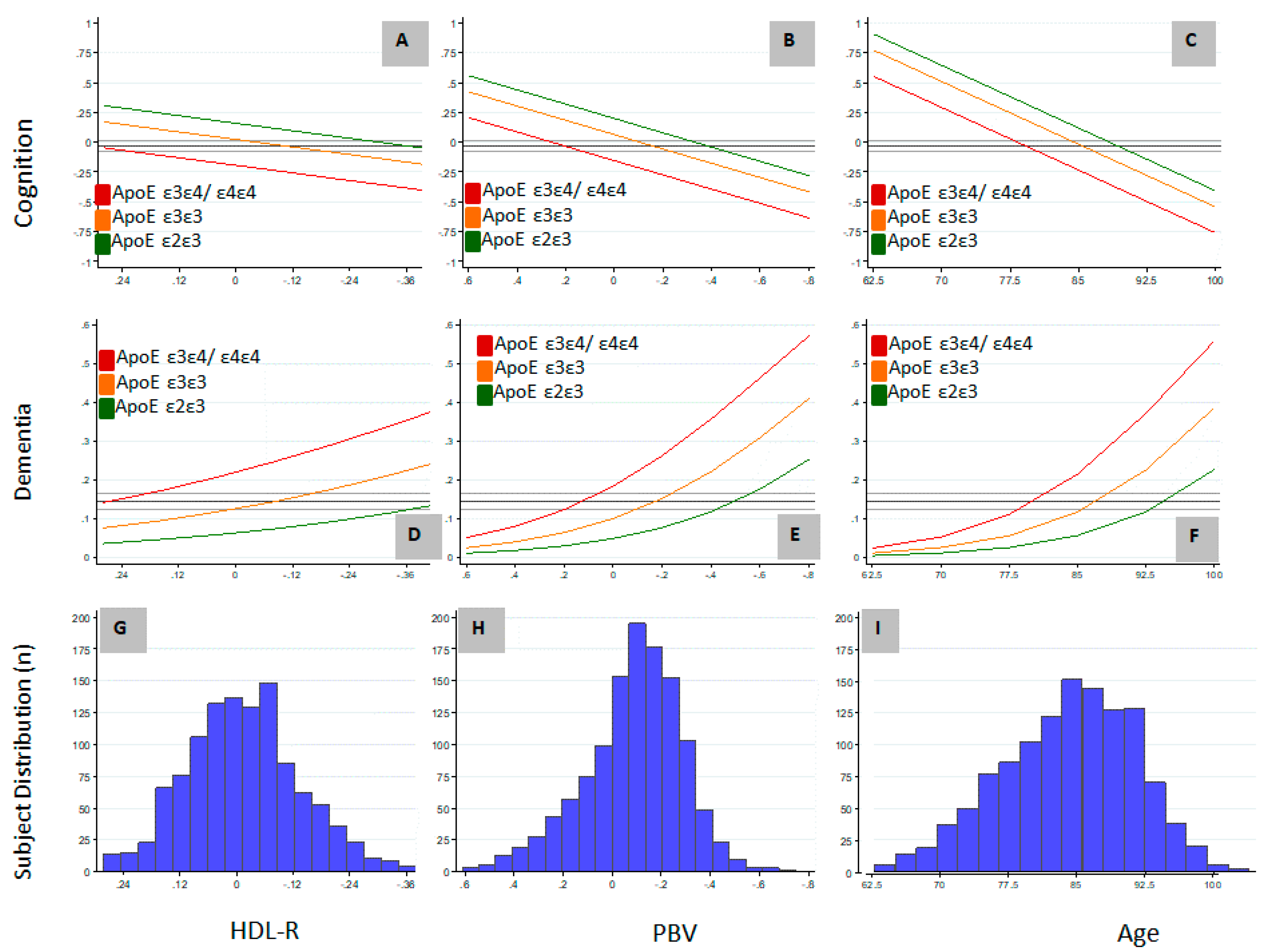

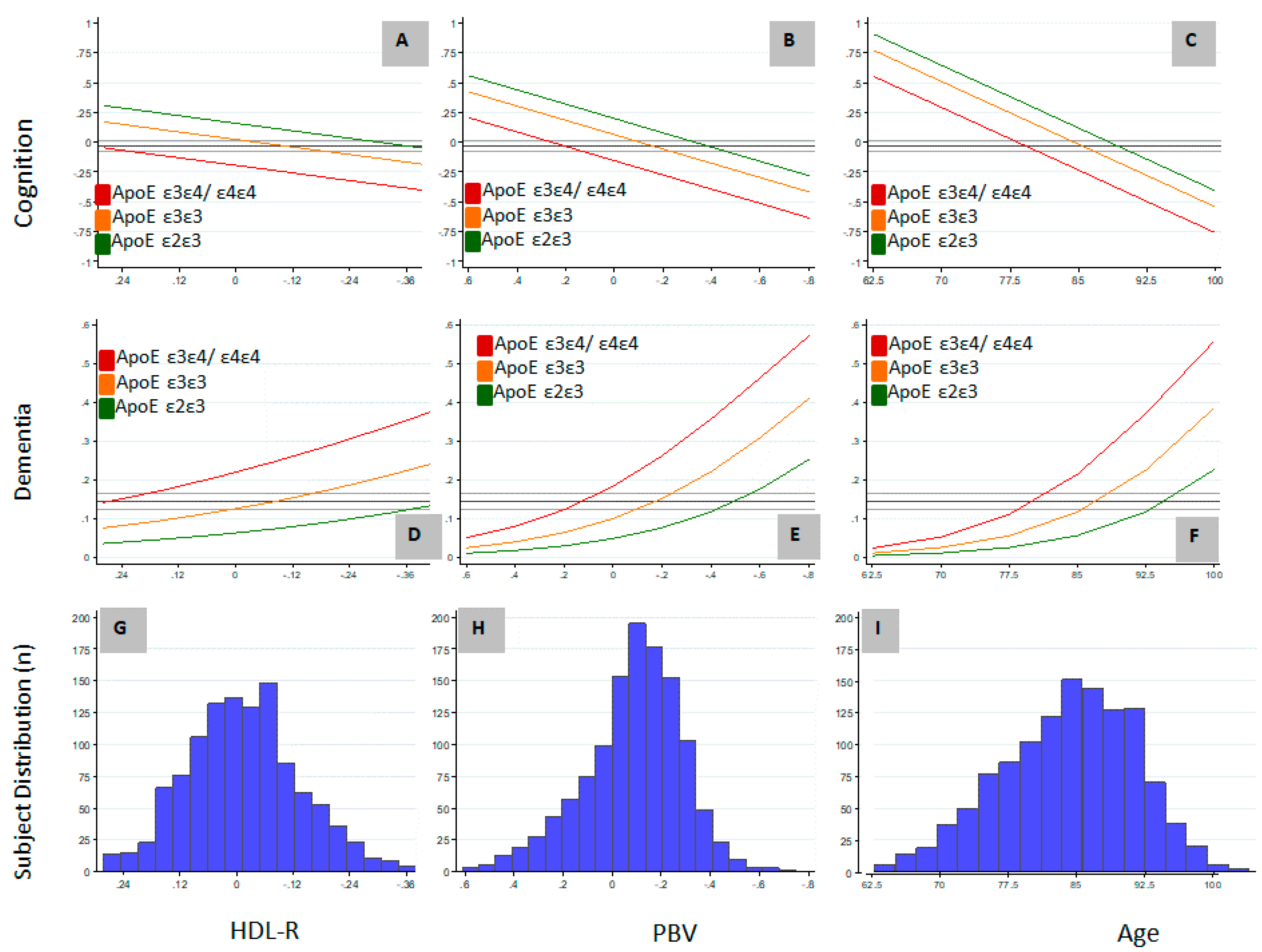{kind=link}
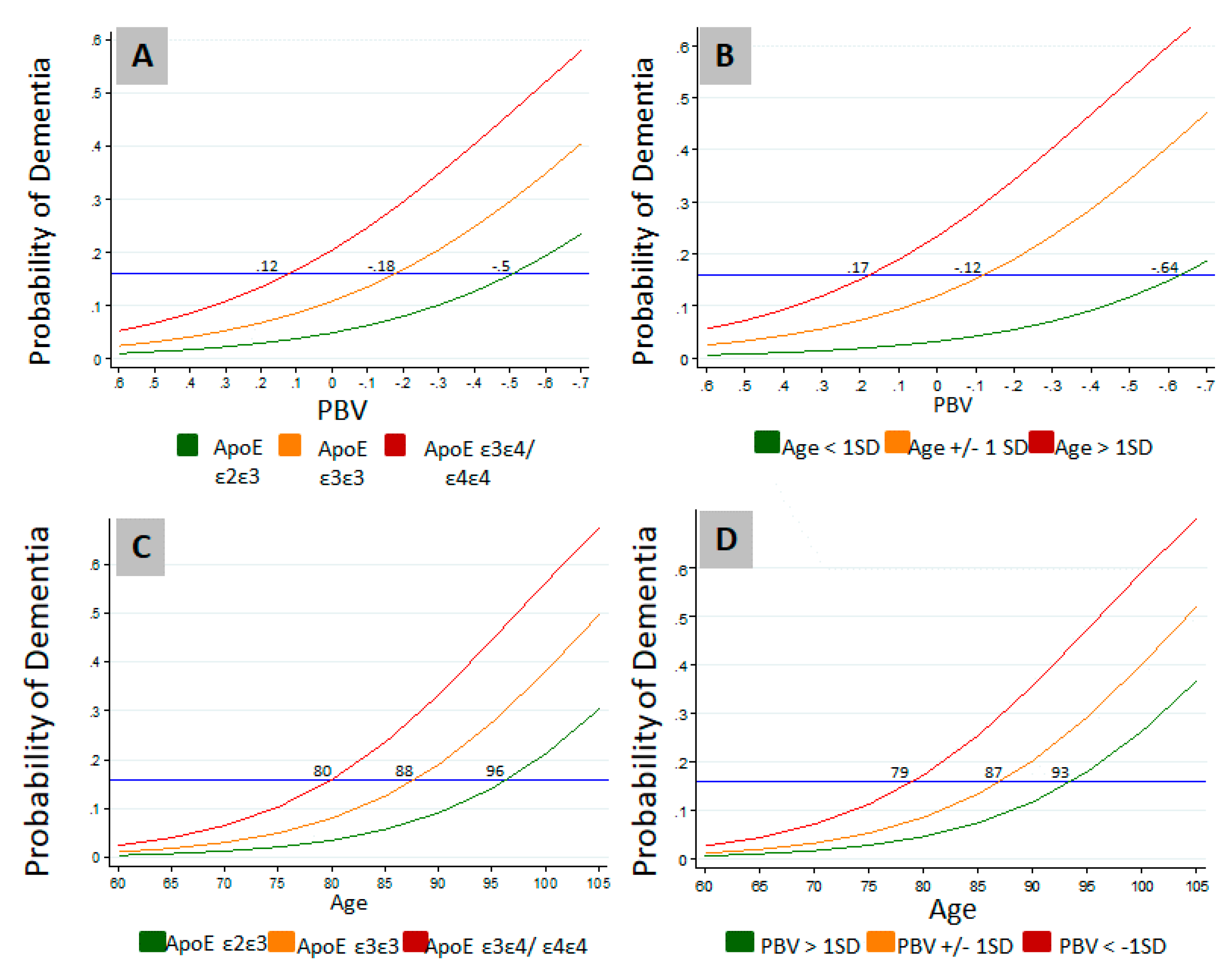{kind=link}
| Female (n = 921) | Male (n = 284) | ||||
|---|---|---|---|---|---|
| Age, Education | |||||
| Variable | Mean | SD | Mean | SD | p-Value |
| Age (range) | 83.8 (58.3–103.9) | 7.6 | 84.3 (64.2–101.0) | 7.1 | 3.8 × 10−1 |
| Education | 15.5 | 3.4 | 16.1 | 4.0 | 1.7 × 10−2 * |
| Serum Lipids | |||||
| Variable | Mean | SD | Mean | SD | p-Value |
| TG | 136.8 | 67.9 | 122.7 | 59.2 | 3.3e × 10−3 * |
| TC | 192.6 | 38.3 | 161.7 | 35.6 | 6.0 × 10−28 * |
| HDL-C | 63.9 | 18.4 | 52.5 | 15.2 | 4.9 × 10−18 * |
| LDL-C | 101.4 | 33.2 | 84.6 | 29.1 | 1.9 × 10−12 * |
| HDL-R | 0.338 | 0.092 | 0.331 | 0.088 | 3.5 × 10−1 |
| Serum Ethanolamine Glycerophospholipids | |||||
| Variable | Mean | SD | Mean | SD | p-Value |
| PE226 | 1.60 | 0.89 | 1.08 | 0.63 | 4.6 × 10−19 * |
| PL224 | 0.85 | 0.48 | 0.72 | 0.31 | 8.0 × 10−5 * |
| PL205 | 1.35 | 1.83 | 0.98 | 0.81 | 1.1 × 10−3 * |
| PL226 | 3.48 | 2.28 | 2.84 | 1.22 | 8.5 × 10−6 * |
| Plasmalogen Ratios | |||||
| Variable | Mean | SD | Mean | SD | p-Value |
| PL205/PE226 | 0.65 | 0.55 | 0.715 | 0.480 | 7.3 × 10−2 |
| PL226/PE226 | 1.81 | 0.88 | 2.18 | 1.00 | 1.6 × 10−9 * |
| PL205/PL224 | 1.94 | 2.49 | 1.67 | 1.98 | 1.0 × 10−1 |
| PL226/PL224 | 4.55 | 2.36 | 4.39 | 2.27 | 2.9 × 10−1 |
| PL205/PL226 | 0.36 | 0.23 | 0.337 | 0.186 | 1.4 × 10−1 |
| PBV | −0.09 | 0.21 | −0.075 | 0.179 | 2.9 × 10−1 |
| Clinical Diagnosis and Cognition by Key Variable Classes | |||||||
|---|---|---|---|---|---|---|---|
| Variable Class | All | No Dementia | Dementia (%) | NCI | MCI | AD | GCOG Mean (SD) |
| Female | 921 | 784 | 137(14.9) | 597 | 168 | 104 | −0.025 (0.829) |
| Male | 284 | 246 | 38 (13.4) | 184 | 68 | 26 | −0.051 (0.756) |
| PBV > 1 SD | 183 | 172 | 11 (6.0) | 143 | 26 | 9 | 0.238 (0.647) |
| PBV ± 1 SD | 851 | 740 | 111 (13.0) | 556 | 177 | 80 | −0.243 (0.782) |
| PBV < 1 SD | 171 | 118 | 53 (31.0) | 82 | 33 | 41 | −0.374 (0.996) |
| ApoE e2e3 | 149 | 139 | 10 (6.7) | 116 | 24 | 3 | 0.178 (0.589) |
| + PBV > 1 SD | 25 | 25 | 0 (0) | 23 | 2 | 0 | 0.370 (0.442) |
| + PBV ± 1 SD | 106 | 100 | 6 (5.7) | 82 | 18 | 2 | 0.184 (0.612) |
| + PBV < 1 SD | 18 | 14 | 4 (22.2) | 11 | 4 | 1 | −0.139 (0.528) |
| ApoE e3e3 | 745 | 641 | 104 (14.0) | 480 | 152 | 75 | −0.028 (0.817) |
| + PBV > 1 SD | 122 | 115 | 7 (5.7) | 96 | 17 | 5 | 0.262 (0.606) |
| + PBV ± 1 SD | 511 | 448 | 63 (12.3) | 330 | 113 | 42 | −0.019 (0.771) |
| + PBV < 1 SD | 112 | 78 | 34 (30.4) | 54 | 22 | 28 | −0.406 (1.063) |
| ApoE e3e4/e4e4 | 269 | 210 | 59 (21.9) | 151 | 55 | 51 | −0.181 (0.900) |
| + PBV > 1 SD | 28 | 24 | 4 (14.3) | 18 | 6 | 4 | −0.010 (0.917) |
| + PBV ± 1 SD | 206 | 164 | 42 (20.4) | 119 | 43 | 36 | −0.173 (0.881) |
| + PBV < 1 SD | 35 | 22 | 13 (37.1) | 14 | 6 | 11 | −0.390 (1.007) |
| Dementia | NCI-MCI-AD | Global Cognition | Episodic Memory | Visuospatial Ability | Perceptual Speed | Semantic Memory | Working Memory | |
|---|---|---|---|---|---|---|---|---|
| (n = 1085) | (n = 1030) | (n = 1049) | (n = 1026) | (n = 992) | (n = 1005) | (n = 1022) | (n = 1049) | |
| Variable | OR 3 (p) | OR 2 (p) | Coef. 1 (p) | Coef. 1 (p) | Coef. 1 (p) | Coef. 1 (p) | Coef. 1 (p) | Coef. 1 (p) |
| PBV | 0.607 (3.3 × 10−6) * | 0.699 (2.5 × 10−6) * | 0.118 (1.7 × 10−7) * | 0.134 (5.3 × 10−6) * | 0.051 (4.4 × 10−2) * | 0.111 (5.2 × 10−5) * | 0.107 (3.1 × 10−5) * | 0.079 (1.6 × 10−3) * |
| HDL-R4 | 0.773 (5.9 × 10−3) * | 0.904 (1.6 × 10−1) | 0.062 (5.6 × 10−3) * | 0.078 (8.5 × 10−3) * | 0.018 (4.8 × 10−1) | 0.019 (4.8 × 10−1) | 0.042 (1.0 × 10−1) | 0.060 (1.6 × 10−2) * |
| APOE ε3ε3 | Reference | Reference | Reference | Reference | Reference | Reference | Reference | Reference |
| APOE ε2ε3 | 0.442 (3.0 × 10−2) * | 0.462 (1.8 × 10−3) * | 0.136 (4.2 × 10−2) * | 0.230 (8.1 × 10−3) * | 0.062 (4.1 × 10−1) | −0.033 (5.2 × 10−5) * | 0.112 (1.4 × 10−1) | 0.048 (5.2 × 10−1) |
| APOE ε3ε4, ε4ε4 | 2.125 (2.2 × 10−4) * | 1.996 (1.9 × 10−5) * | −0.217 (6.2 × 10−5) * | −0.319 (7.6 × 10−6) * | −0.076 (2.2 × 10−1) | −0.276 (3.1 × 10−5) * | −0.190 (2.0 × 10−3) * | −0.105 (7.9 × 10−2) |
| Age * | 2.271 (1.4 × 10−13) * | 2.056 (2.0 × 10−19) * | −0.262 (4.1 × 10−3) * | −0.290 (2.6 × 10−22) * | −0.137 (8.6 × 10−8) * | −0.352 (1.6 × 10−35) * | −0.191 (1.0 × 10−13) * | −0.177 (1.9 × 10−12) * |
| Education * | 0.957 (6.5 × 10−1) | 0.917 (2.3 × 10−1) | 0.183 (4.8 × 10−16) * | 0.165 (1.9 × 10−8) * | 0.255 (1.0 × 10−22) * | 0.204 (1.2 × 10−13) * | 0.229 (6.7 × 10−19) * | 0.174 (3.2 × 10−12) * |
| Female | Reference | Reference | Reference | Reference | Reference | Reference | Reference | Reference |
| Male | 1.035 (8.8 × 10−1) | 1.104 (5.5 × 10−1) | −0.048 (3.7 × 10−1) | −0.159 (2.3 × 10−2) * | 0.205 (6.9 × 10−4) * | −0.156 (1.6 × 10−2) * | −0.013 (8.2 × 10−1) | 0.066 (2.7 × 10−1) |
| Total Cholesterol (n = 1085) | LDL-Cholesterol (n = 1078) | HDL-Cholesterol (n = 909) | HDL-R (n = 909) | Trigylcerides (n = 909) | |
|---|---|---|---|---|---|
| Variable | Coef. 1 (p) | Coef. 1 (p) | Coef. 1 (p) | Coef. 1 (p) | Coef. 1 (p) |
| PBV | 0.009 (5.7 × 10−4) * | 0.014 (2.6 × 10−3) * | 0.029 (3.8 × 10−15) * | 0.020 (5.0 × 10−8) * | −0.043 (3.8 × 10−14) * |
| APOE ε3ε3 | Reference | Reference | Reference | Reference | Reference |
| APOE ε2ε3 | −0.009 (2.8 × 10−1) | −0.036 (7.6 × 10−3) * | 0.018 (9.7 × 10−2) | 0.027 (1.3 × 10−2) * | 0.014 (4.2 × 10−1) |
| APOE ε3ε4, ε4ε4 | 0.021 (9.7 × 10−4) * | 0.042 (1.2 × 10−4) * | −0.002 (8.1 × 10−1) | −0.023 (6.5 × 10−3) * | 0.002 (8.8 × 10−1) |
| Age | −0.009 (1.1 × 10−3) * | −0.014 (2.0 × 10−3) * | −0.002 (5.6 × 10−1) | 0.007 (6.1 × 10−2) | −0.006 (2.6 × 10−1) |
| Education | −0.0004 (8.8 × 10−1) | 0.0001 (9.9 × 10−1) | 0.003 (3.8 × 10−1) | 0.004 (3.2 × 10−1) | −0.008 (1.4 × 10−1) |
| Female | Reference | Reference | Reference | Reference | Reference |
| Male | −0.001 (8.5 × 10−1) | −0.001 (9.4 × 10−1) | −0.002 (8.1 × 10−1) | −0.001 (8.7 × 10−1) | 0.007 (6.3 × 10−1) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goodenowe, D.B.; Senanayake, V. Relation of Serum Plasmalogens and APOE Genotype to Cognition and Dementia in Older Persons in a Cross-Sectional Study. Brain Sci. 2019, 9, 92. https://doi.org/10.3390/brainsci9040092
Goodenowe DB, Senanayake V. Relation of Serum Plasmalogens and APOE Genotype to Cognition and Dementia in Older Persons in a Cross-Sectional Study. Brain Sciences. 2019; 9(4):92. https://doi.org/10.3390/brainsci9040092
Chicago/Turabian StyleGoodenowe, Dayan B., and Vijitha Senanayake. 2019. "Relation of Serum Plasmalogens and APOE Genotype to Cognition and Dementia in Older Persons in a Cross-Sectional Study" Brain Sciences 9, no. 4: 92. https://doi.org/10.3390/brainsci9040092
APA StyleGoodenowe, D. B., & Senanayake, V. (2019). Relation of Serum Plasmalogens and APOE Genotype to Cognition and Dementia in Older Persons in a Cross-Sectional Study. Brain Sciences, 9(4), 92. https://doi.org/10.3390/brainsci9040092