Transient Chemogenetic Inhibition of D1-MSNs in the Dorsal Striatum Enhances Methamphetamine Self-Administration


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Animals
2.2. Viral Vector Construction, Surgery and Viral Gene Transfer
2.3. Surgery for Implanting Jugular Vein Catheters
2.4. Training and Maintenance on an Extended Access Schedule (Contingent Meth)
2.5. CNO Injections
2.6. Brain Tissue Collection for Immunohistochemistry and Western Blotting
2.7. Immunohistochemistry
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
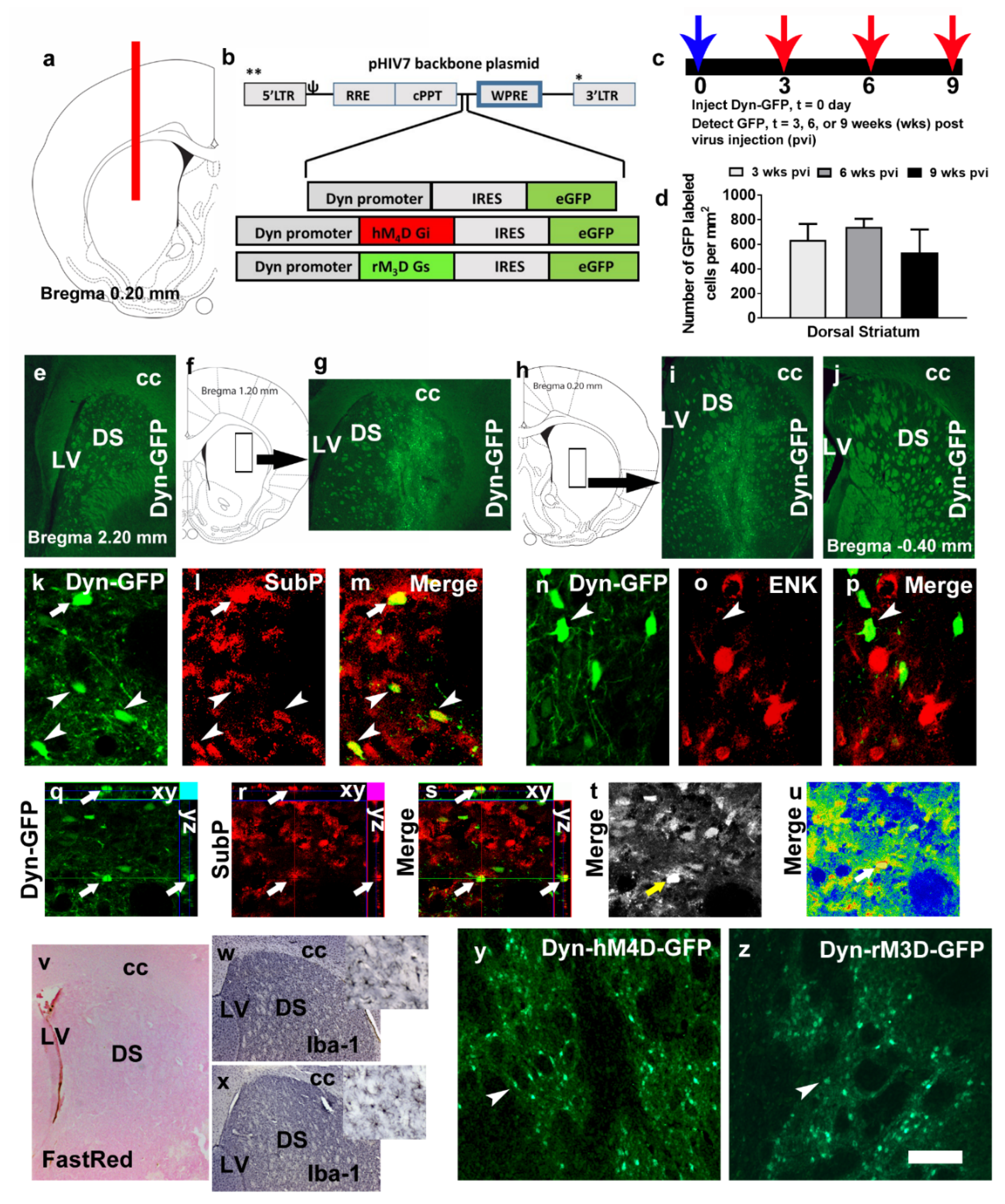
3.1. Dynorphin Promoter Driven Lentivirus Expression is Localized to D1-MSNs and not D2-MSNs
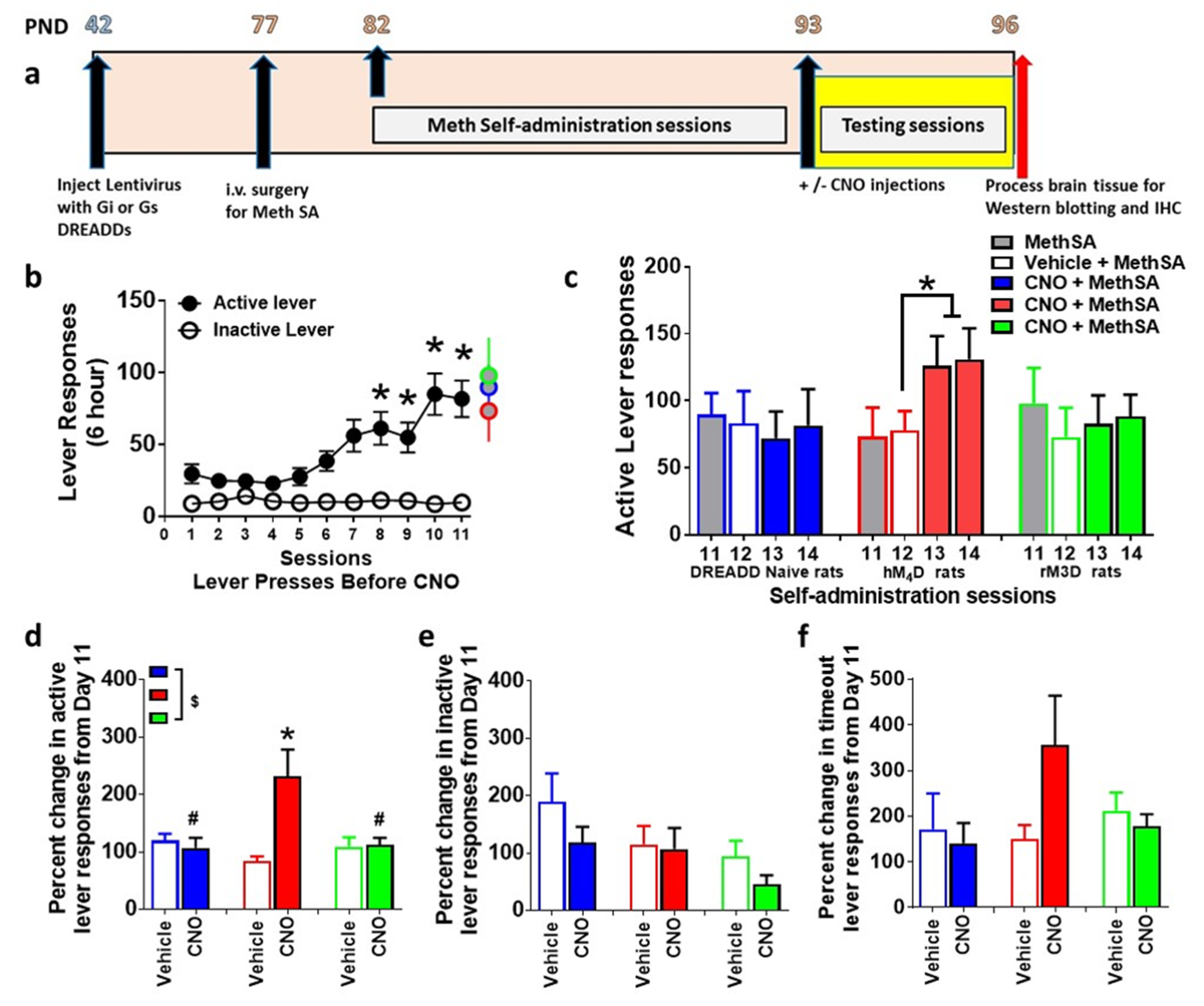
3.2. Lentivirus Expression does not Alter Extended Access Methamphetamine Self-Administration
3.3. CNO Effects Extended Access Methamphetamine Self-Administration in Animals Expressing hM4D DREADD
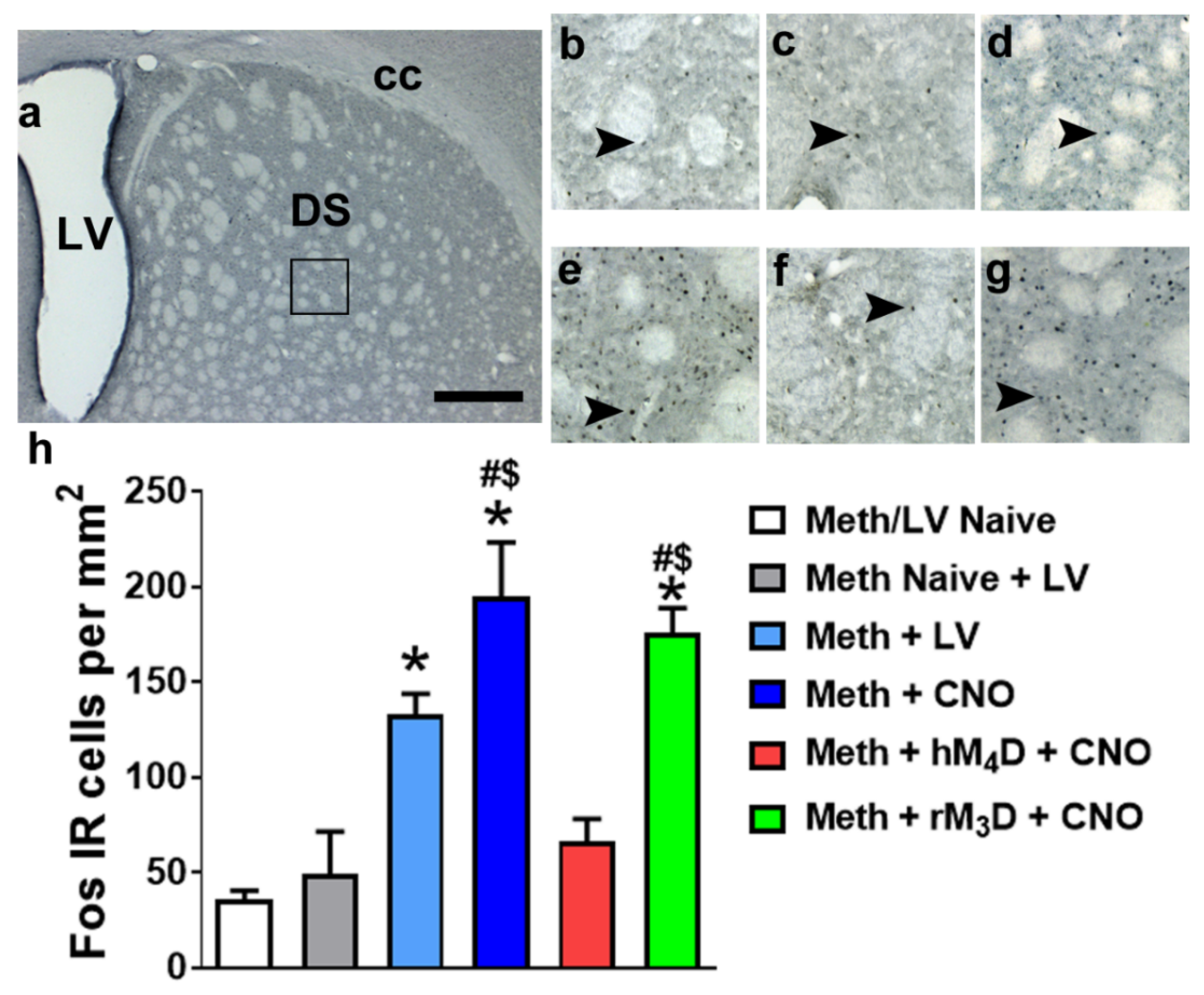
3.4. CNO Reduces Fos Expression in Animals Expressing hM4D DREADD
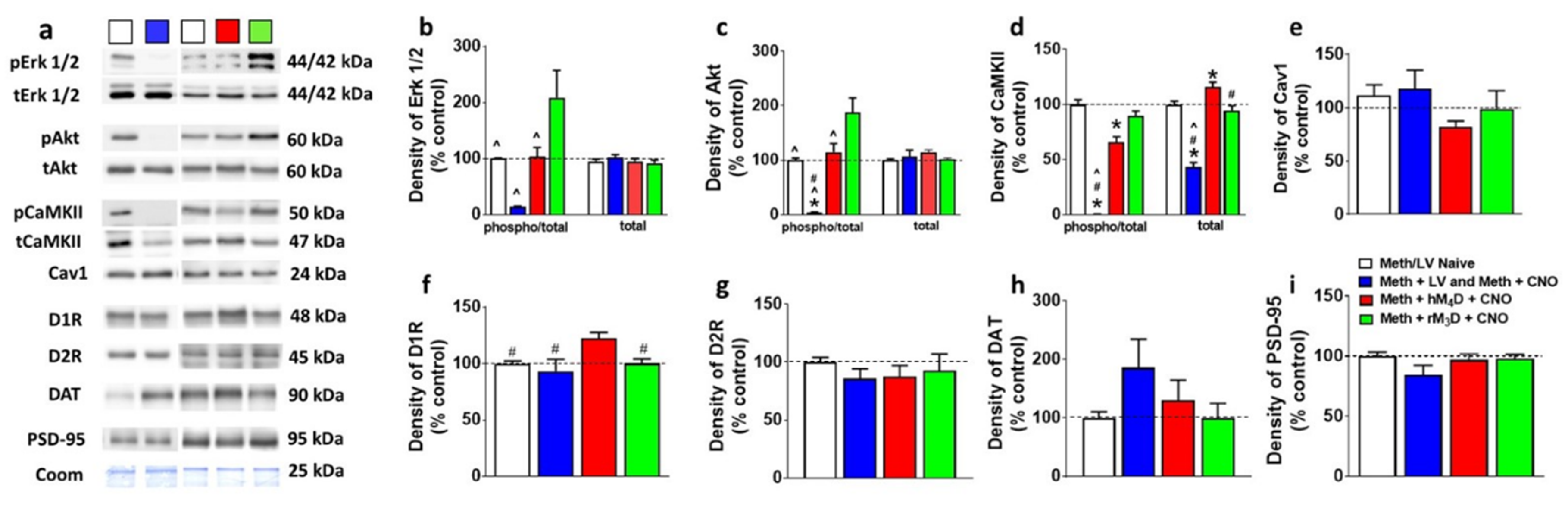
3.5. hM4D Animals Have Distinct Changes in Signaling Molecules in the Dorsal Striatum Compared with rM3D Animals
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ahmed, S.H. Transition from Moderate to Excessive Drug Intake: Change in Hedonic Set Point. Science 1998, 282, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Le Cozannet, R.; Markou, A.; Kuczenski, R. Extended-access, but not limited-access, methamphetamine self-administration induces behavioral and nucleus accumbens dopamine response changes in rats. Eur. J. Neurosci. 2013, 38, 3487–3495. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, O.; Wee, S.; Specio, S.E.; Koob, G.F.; Pulvirenti, L. Escalation of methamphetamine self-administration in rats: A dose-effect function. Psychopharmacology 2006, 186, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Balleine, B.W.; Liljeholm, M.; Ostlund, S.B. The integrative function of the basal ganglia in instrumental conditioning. Behav. Brain Res. 2009, 199, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.H.; Knowlton, B.J.; Balleine, B.W. Lesions of dorsolateral striatum preserve outcome expectancy but disrupt habit formation in instrumental learning. Eur. J. Neurosci. 2004, 19, 181–189. [Google Scholar] [CrossRef]
- Belin, D.; Everitt, B.J. Cocaine Seeking Habits Depend upon Dopamine-Dependent Serial Connectivity Linking the Ventral with the Dorsal Striatum. Neuron 2008, 57, 432–441. [Google Scholar] [CrossRef]
- Yin, H.H.; Ostlund, S.B.; Knowlton, B.J.; Balleine, B.W. The role of the dorsomedial striatum in instrumental conditioning. Eur. J. Neurosci. 2005, 22, 513–523. [Google Scholar] [CrossRef]
- Tepper, J.M.; Abercrombie, E.D.; Bolam, J.P. Basal ganglia macrocircuits. Prog. Brain Res. 2007, 160, 3–7. [Google Scholar]
- Gerfen, C.R.; Young, W.S. Distribution of striatonigral and striatopallidal peptidergic neurons in both patch and matrix compartments: An in situ hybridization histochemistry and fluorescent retrograde tracing study. Brain Res. 1988, 460, 161–167. [Google Scholar] [CrossRef]
- Ince, E.; Ciliax, B.J.; Levey, A.I. Differential expression of D1 and D2 dopamine and m4 muscarinic acetylcholine receptor proteins in identified striatonigral neurons. Synapse 1997, 27, 357–366. [Google Scholar] [CrossRef]
- Sibley, D.R.; Monsma, F.J. Molecular biology of dopamine receptors. Trends Pharmacol. Sci. 1992, 13, 61–69. [Google Scholar] [CrossRef]
- Gerfen, C.R. The neostriatal mosaic: Multiple levels of compartmental organization. Trends Neurosci. 1992, 15, 133–139. [Google Scholar] [CrossRef]
- Deniau, J.; Mailly, P.; Maurice, N.; Charpier, S. The pars reticulata of the substantia nigra: A window to basal ganglia output. Prog. Brain Res. 2007, 160, 151–172. [Google Scholar] [PubMed]
- Graybiel, A.M. Neurotransmitters and neuromodulators in the basal ganglia. Trends Neurosci. 1990, 13, 244–254. [Google Scholar] [CrossRef]
- Féger, J.; Ohye, C. The unitary activity of the substantia nigra following stimulation of the striatum in the awake monkey. Brain Res. 1975, 89, 155–159. [Google Scholar] [CrossRef]
- Collingridge, G.L.; Davies, J. The influence of striatal stimulation and putative neurotransmitters on identified neurones in the rat substantia nigra. Brain Res. 1981, 212, 345–359. [Google Scholar] [CrossRef]
- Kravitz, A.V.; Freeze, B.S.; Parker, P.R.L.; Kay, K.; Thwin, M.T.; Deisseroth, K.; Kreitzer, A.C. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature 2010, 466, 622–626. [Google Scholar] [CrossRef]
- Beauvais, G.; Jayanthi, S.; McCoy, M.T.; Ladenheim, B.; Cadet, J.L. Differential effects of methamphetamine and SCH23390 on the expression of members of IEG families of transcription factors in the rat striatum. Brain Res. 2010, 1318, 1–10. [Google Scholar] [CrossRef]
- Brennan, K.A.; Carati, C.; Lea, R.A.; Fitzmaurice, P.S.; Schenk, S. Effect of D1-like and D2-like receptor antagonists on methamphetamine and 3,4-methylenedioxymethamphetamine self-administration in rats. Behav. Pharmacol. 2009, 20, 688–694. [Google Scholar] [CrossRef]
- Gross, N.B.; Duncker, P.C.; Marshall, J.F. Striatal dopamine D1 and D2 receptors: Widespread influences on methamphetamine-induced dopamine and serotonin neurotoxicity. Synapse 2011, 65, 1144–1155. [Google Scholar] [CrossRef]
- Nishiguchi, M.; Kinoshita, H.; Kasuda, S.; Takahashi, M.; Yamamura, T.; Matsui, K.; Ouchi, H.; Minami, T.; Hishida, S.; Nishio, H. Effects of dopamine antagonists on methamphetamine-induced dopamine release in high and low alcohol preference rats. Toxicol. Mech. Methods 2010, 20, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; McGinty, J.F. D1 and D2 dopamine receptors differentially mediate the activation of phosphoproteins in the striatum of amphetamine-sensitized rats. Psychopharmacology 2011, 214, 653–663. [Google Scholar] [CrossRef]
- Sonsalla, P.K.; Gibb, J.W.; Hanson, G.R. Roles of D1 and D2 dopamine receptor subtypes in mediating the methamphetamine-induced changes in monoamine systems. J. Pharmacol. Exp. Ther. 1986, 238, 932–937. [Google Scholar]
- Ahmed, S.H.; Koob, G.F. Changes in response to a dopamine receptor antagonist in rats with escalating cocaine intake. Psychopharmacology 2004, 172, 450–454. [Google Scholar] [CrossRef]
- Martinez, D.; Broft, A.; Foltin, R.W.; Slifstein, M.; Hwang, D.-R.; Huang, Y.; Perez, A.; Frankel, W.G.; Cooper, T.; Kleber, H.D.; et al. Cocaine Dependence and D2 Receptor Availability in the Functional Subdivisions of the Striatum: Relationship with Cocaine-Seeking Behavior. Neuropsychopharmacology 2004, 29, 1190–1202. [Google Scholar] [CrossRef]
- Dalley, J.W.; Fryer, T.D.; Brichard, L.; Robinson, E.S.J.; Theobald, D.E.H.; Lääne, K.; Peña, Y.; Murphy, E.R.; Shah, Y.; Probst, K.; et al. Nucleus accumbens D2/3 receptors predict trait impulsivity and cocaine reinforcement. Science 2007, 315, 1267–1270. [Google Scholar] [CrossRef]
- Volkow, N.D.; Chang, L.; Wang, G.-J.; Fowler, J.S.; Ding, Y.-S.; Sedler, M.; Logan, J.; Franceschi, D.; Gatley, J.; Hitzemann, R.; et al. Low Level of Brain Dopamine D2Receptors in Methamphetamine Abusers: Association With Metabolism in the Orbitofrontal Cortex. Am. J. Psychiatry 2001, 158, 2015–2021. [Google Scholar] [CrossRef]
- Caine, S.B.; Thomsen, M.; Gabriel, K.I.; Berkowitz, J.S.; Gold, L.H.; Koob, G.F.; Tonegawa, S.; Zhang, J.; Xu, M. Lack of self-administration of cocaine in dopamine D1 receptor knock-out mice. J. Neurosci. 2007, 27, 13140–13150. [Google Scholar] [CrossRef] [PubMed]
- Lobo, M.K.; Covington, H.E., 3rd; Chaudhury, D.; Friedman, A.K.; Sun, H.; Damez-Werno, D.; Dietz, D.M.; Zaman, S.; Koo, J.W.; Kennedy, P.J.; et al. Cell type-specific loss of BDNF signaling mimics optogenetic control of cocaine reward. Science 2010, 330, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Yager, L.M.; Garcia, A.F.; Donckels, E.A.; Ferguson, S.M. Chemogenetic inhibition of direct pathway striatal neurons normalizes pathological, cue-induced reinstatement of drug-seeking in rats. Addict. Boil. 2018, 24, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, S.M.; Eskenazi, D.; Ishikawa, M.; Wanat, M.J.; Phillips, P.E.; Dong, Y.; Roth, B.L.; Neumaier, J.F. Transient neuronal inhibition reveals opposing roles of indirect and direct pathways in sensitization. Nat. Neurosci. 2011, 14, 22–24. [Google Scholar] [CrossRef] [PubMed]
- Kravitz, A.V.; Tye, L.D.; Kreitzer, A.C. Distinct roles for direct and indirect pathway striatal neurons in reinforcement. Nat. Neurosci. 2012, 15, 816–818. [Google Scholar] [CrossRef] [PubMed]
- Darvas, M.; Palmiter, R.D. Specific contributions of N-methyl-D-aspartate receptors in the dorsal striatum to cognitive flexibility. Neuroscience 2015, 284, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, S.M.; Phillips, P.E.M.; Roth, B.L.; Wess, J.; Neumaier, J.F. Direct-pathway striatal neurons regulate the retention of decision-making strategies. J. Neurosci. 2013, 33, 11668–11676. [Google Scholar] [CrossRef] [PubMed]
- Yam, P.Y.; Li, S.; Wu, J.; Hu, J.; Zaia, J.A.; Yee, J.-K. Design of HIV Vectors for Efficient Gene Delivery into Human Hematopoietic Cells. Mol. Ther. 2002, 5, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Head, B.P.; Hu, Y.; Finley, J.C.; Saldana, M.D.; Bonds, J.A.; Miyanohara, A.; Niesman, I.R.; Ali, S.S.; Murray, F.; Insel, P.A.; et al. Neuron-targeted Caveolin-1 Protein Enhances Signaling and Promotes Arborization of Primary Neurons. J. Boil. Chem. 2011, 286, 33310–33321. [Google Scholar] [CrossRef]
- Galinato, M.H.; Lockner, J.W.; Fannon-Pavlich, M.J.; Sobieraj, J.C.; Staples, M.C.; Somkuwar, S.S.; Ghofranian, A.; Chaing, S.; Navarro, A.I.; Joea, A.; et al. A synthetic small-molecule Isoxazole-9 protects against methamphetamine relapse. Mol. Psychiatry 2018, 23, 629–638. [Google Scholar] [CrossRef]
- Galinato, M.H.; Orio, L.; Mandyam, C.D. Methamphetamine differentially affects BDNF and cell death factors in anatomically defined regions of the hippocampus. Neuroscience 2015, 286, 97–108. [Google Scholar] [CrossRef]
- Sobieraj, J.C.; Kim, A.; Fannon, M.J.; Mandyam, C.D. Chronic wheel running-induced reduction of extinction and reinstatement of methamphetamine seeking in methamphetamine dependent rats is associated with reduced number of periaqueductal gray dopamine neurons. Anat. Embryol. 2014, 221, 261–276. [Google Scholar] [CrossRef]
- MacLaren, D.A.A.; Browne, R.W.; Shaw, J.K.; Radhakrishnan, S.K.; Khare, P.; España, R.A.; Clark, S.D. Clozapine N-Oxide Administration Produces Behavioral Effects in Long-Evans Rats: Implications for Designing DREADD Experiments. eNeuro 2016, 3. [Google Scholar] [CrossRef]
- Somkuwar, S.S.; Fannon-Pavlich, M.J.; Ghofranian, A.; Quigley, J.A.; Dutta, R.R.; Galinato, M.H.; Mandyam, C.D.; Fannon, M.J. Wheel running reduces ethanol seeking by increasing neuronal activation and reducing oligodendroglial/neuroinflammatory factors in the medial prefrontal cortex. Brain Behav. Immun. 2016, 58, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, D.; Petryszyn, S.; Sanchez, M.G.; Bories, C.; Beaulieu, J.M.; De Koninck, Y.; Parent, A.; Parent, M. Striatal Neurons Expressing D1 and D2 Receptors are Morphologically Distinct and Differently Affected by Dopamine Denervation in Mice. Sci. Rep. 2017, 7, 41432. [Google Scholar] [CrossRef] [PubMed]
- Mandyam, C.D.; Norris, R.D.; Eisch, A.J. Chronic morphine induces premature mitosis of proliferating cells in the adult mouse subgranular zone. J. Neurosci. Res. 2004, 76, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Mandyam, C.D. Methamphetamine affects cell proliferation in the medial prefrontal cortex: A new niche for toxicity. Pharmacol. Biochem. Behav. 2014, 126, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, H.; Yamada, K.; Mizuno, M.; Mizuno, T.; Nitta, A.; Noda, Y.; Nabeshima, T. Regulations of Methamphetamine Reward by Extracellular Signal-Regulated Kinase 1/2/ets-Like Gene-1 Signaling Pathway via the Activation of Dopamine Receptors. Mol. Pharmacol. 2004, 65, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Granado, N.; Moratalla, R.; Ares-Santos, S. The role of dopamine receptors in the neurotoxicity of methamphetamine. J. Intern. Med. 2013, 273, 437–453. [Google Scholar]
- Wei, Y.; Williams, J.M.; Dipace, C.; Sung, U.; Javitch, J.A.; Galli, A.; Saunders, C. Dopamine transporter activity mediates amphetamine-induced inhibition of Akt through a Ca2+/calmodulin-dependent kinase II-dependent mechanism. Mol. Pharmacol. 2007, 71, 835–842. [Google Scholar] [CrossRef]
- Ares-Santos, S.; Granado, N.; Oliva, I.; O’Shea, E.; Martín, E.D.; Colado, M.; Moratalla, R. Dopamine D1 receptor deletion strongly reduces neurotoxic effects of methamphetamine. Neurobiol. Dis. 2012, 45, 810–820. [Google Scholar] [CrossRef]
- O’Dell, S.J.; Weihmuller, F.B.; Marshall, J.F. Methamphetamine-Induced Dopamine Overflow and Injury to Striatal Dopamine Terminals: Attenuation by Dopamine D1or D2Antagonists. J. Neurochem. 1993, 60, 1792–1799. [Google Scholar] [CrossRef]
- Hiranita, T.; Yamamoto, T.; Nawata, Y. A tryptamine-derived catecholaminergic enhancer, (-)-1-(benzofuran-2-yl)-2-propylaminopentane [(-)-BPAP], attenuates reinstatement of methamphetamine-seeking behavior in rats. Neuroscience 2010, 165, 300–312. [Google Scholar] [CrossRef]
- Berke, J.D.; Hyman, S.E. Addiction, dopamine, and the molecular mechanisms of memory. Neuron 2000, 25, 515–532. [Google Scholar] [CrossRef]
- Nestler, E.J. Molecular basis of long-term plasticity underlying addiction. Nat. Rev. Neurosci. 2001, 2, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Krasnova, I.N.; Marchant, N.J.; Ladenheim, B.; McCoy, M.T.; Panlilio, L.V.; Bossert, J.M.; Shaham, Y.; Cadet, J.L. Incubation of Methamphetamine and Palatable Food Craving after Punishment-Induced Abstinence. Neuropsychopharmacology 2014, 39, 2008–2016. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.L.; De Santis, S.; See, R.E. Extended methamphetamine self-administration enhances reinstatement of drug seeking and impairs novel object recognition in rats. Psychopharmacology 2008, 199, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Nawaratne, V.; Leach, K.; Suratman, N.; LoIacono, R.E.; Felder, C.C.; Armbruster, B.N.; Roth, B.L.; Sexton, P.M.; Christopoulos, A. New Insights into the Function of M4 Muscarinic Acetylcholine Receptors Gained Using a Novel Allosteric Modulator and a DREADD (Designer Receptor Exclusively Activated by a Designer Drug). Mol. Pharmacol. 2008, 74, 1119–1131. [Google Scholar] [CrossRef]
- Farrell, M.S.; Pei, Y.; Wan, Y.; Yadav, P.N.; Daigle, T.L.; Urban, D.J.; Lee, H.M.; Sciaky, N.; Simmons, A.; Nonneman, R.J.; et al. A Galphas DREADD mouse for selective modulation of cAMP production in striatopallidal neurons. Neuropsychopharmacology 2013, 38, 854–862. [Google Scholar] [CrossRef]
- Armbruster, B.N.; Li, X.; Pausch, M.H.; Herlitze, S.; Roth, B.L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. USA 2007, 104, 5163–5168. [Google Scholar] [CrossRef]
- Gomez, J.L.; Bonaventura, J.; Lesniak, W.; Mathews, W.B.; Sysa-Shah, P.; Rodriguez, L.A.; Ellis, R.J.; Richie, C.T.; Harvey, B.K.; Dannals, R.F.; et al. Chemogenetics revealed: DREADD occupancy and activation via converted clozapine. Science 2017, 357, 503–507. [Google Scholar] [CrossRef]
- Mahler, S.V.; Aston-Jones, G. CNO Evil? Considerations for the Use of DREADDs in Behavioral Neuroscience. Neuropsychopharmacology 2018, 43, 934–936. [Google Scholar] [CrossRef]
- Bardo, M.T.; Valone, J.M.; Bevins, R.A. Locomotion and conditioned place preference produced by acute intravenous amphetamine: Role of dopamine receptors and individual differences in amphetamine self-administration. Psychopharmacology 1999, 143, 39–46. [Google Scholar] [CrossRef]
- Koshikawa, N.; Mori, E.; Oka, K.; Nomura, H.; Yatsushige, N.; Maruyama, Y. Effects of SCH23390 injection into the dorsal striatum and nucleus accumbens on methamphetamine-induced gnawing and hyperlocomotion in rats. J. Nihon Univ. Sch. Dent. 1989, 31, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.J.; Belin, D.; Economidou, D.; Pelloux, Y.; Dalley, J.W.; Robbins, T.W. Review. Neural mechanisms underlying the vulnerability to develop compulsive drug-seeking habits and addiction. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 3125–3135. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.J.; Robbins, T.W. From the ventral to the dorsal striatum: Devolving views of their roles in drug addiction. Neurosci. Biobehav. Rev. 2013, 37, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Tecuapetla, F.; Jin, X.; Lima, S.Q.; Costa, R.M. Complementary Contributions of Striatal Projection Pathways to Action Initiation and Execution. Cell 2016, 166, 703–715. [Google Scholar] [CrossRef]
- Isomura, Y.; Takekawa, T.; Harukuni, R.; Handa, T.; Aizawa, H.; Takada, M.; Fukai, T. Reward-Modulated Motor Information in Identified Striatum Neurons. J. Neurosci. 2013, 33, 10209–10220. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.-J.; Tomasi, D.; Baler, R.D. Unbalanced neuronal circuits in addiction. Curr. Opin. Neurobiol. 2013, 23, 639–648. [Google Scholar] [CrossRef]
- Dobbs, L.K.; Kaplan, A.R.; Bock, R.; Phamluong, K.; Shin, J.H.; Bocarsly, M.E.; Eberhart, L.; Ron, D.; Alvarez, V.A. D1 receptor hypersensitivity in mice with low striatal D2 receptors facilitates select cocaine behaviors. Neuropsychopharmacology 2019, 44, 805–816. [Google Scholar] [CrossRef]
- Shen, F.; Jin, S.; Duan, Y.; Liang, J.; Zhang, M.; Jiang, F.; Sui, N. Distinctive Changes of L-Type Calcium Channels and Dopamine Receptors in the Dorsomedial and Dorsolateral Striatum after the Expression of Habitual Cocaine-Seeking Behavior in Rats. Neuroscience 2018, 370, 139–147. [Google Scholar] [CrossRef]
- Hoffmann, H.M.; Nadal, R.; Vignes, M.; Ortiz, J. Chronic cocaine self-administration modulates ERK1/2 and CREB responses to dopamine receptor agonists in striatal slices. Addict. Biol. 2012, 17, 565–575. [Google Scholar] [CrossRef]
- Hikosaka, O.; Takikawa, Y.; Kawagoe, R. Role of the basal ganglia in the control of purposive saccadic eye movements. Physiol. Rev. 2000, 80, 953–978. [Google Scholar] [CrossRef]
- Mink, J.W. The basal ganglia: Focused selection and inhibition of competing motor programs. Prog. Neurobiol. 1996, 50, 381–425. [Google Scholar] [CrossRef]
- Bock, R.; Shin, J.H.; Kaplan, A.R.; Dobi, A.; Markey, E.; Kramer, P.F.; Gremel, C.M.; Christensen, C.H.; Adrover, M.F.; Alvarez, V.A. Strengthening the accumbal indirect pathway promotes resilience to compulsive cocaine use. Nat. Neurosci. 2013, 16, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Floresco, S.B.; West, A.R.; Ash, B.; Moore, H.; Grace, A.A. Afferent modulation of dopamine neuron firing differentially regulates tonic and phasic dopamine transmission. Nat. Neurosci. 2003, 6, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Otani, S.; Grace, A.A. The Yin and Yang of dopamine release: A new perspective. Neuropharmacology 2007, 53, 583–587. [Google Scholar] [CrossRef]
- Xi, Z.-X.; Kleitz, H.K.; Deng, X.; Ladenheim, B.; Peng, X.-Q.; Li, X.; Gardner, E.L.; Stein, E.A.; Cadet, J.L. A single high dose of methamphetamine increases cocaine self-administration by depletion of striatal dopamine in rats. Neuroscience 2009, 161, 392–402. [Google Scholar] [CrossRef]
- Stefanski, R.; Ladenheim, B.; Lee, S.H.; Cadet, J.L.; Goldberg, S.R. Neuroadaptations in the dopaminergic system after active self-administration but not after passive administration of methamphetamine. Eur. J. Pharmacol. 1999, 371, 123–135. [Google Scholar] [CrossRef]
- Steinkellner, T.; Mus, L.; Eisenrauch, B.; Constantinescu, A.; Leo, D.; Konrad, L.; Rickhag, M.; Sørensen, G.; Efimova, E.V.; Kong, E.; et al. In vivo amphetamine action is contingent on alphaCaMKII. Neuropsychopharmacology 2014, 39, 2681–2693. [Google Scholar] [CrossRef]
- Steinkellner, T.; Yang, J.W.; Montgomery, T.R.; Chen, W.Q.; Winkler, M.T.; Sucic, S.; Lubec, G.; Freissmuth, M.; Elgersma, Y.; Sitte, H.H.; et al. Ca(2+)/calmodulin-dependent protein kinase IIalpha (alphaCaMKII) controls the activity of the dopamine transporter: Implications for Angelman syndrome. J. Biol. Chem. 2012, 287, 29627–29635. [Google Scholar] [CrossRef]
- Fog, J.U.; Khoshbouei, H.; Holy, M.; Owens, W.A.; Vaegter, C.B.; Sen, N.; Nikandrova, Y.; Bowton, E.; McMahon, D.G.; Colbran, R.J.; et al. Calmodulin Kinase II Interacts with the Dopamine Transporter C Terminus to Regulate Amphetamine-Induced Reverse Transport. Neuron 2006, 51, 417–429. [Google Scholar] [CrossRef]
- Giros, B.; Caron, M.G. Molecular characterization of the dopamine transporter. Trends Pharmacol. Sci. 1993, 14, 43–49. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliver, R.J.; Purohit, D.C.; Kharidia, K.M.; Mandyam, C.D. Transient Chemogenetic Inhibition of D1-MSNs in the Dorsal Striatum Enhances Methamphetamine Self-Administration. Brain Sci. 2019, 9, 330. https://doi.org/10.3390/brainsci9110330
Oliver RJ, Purohit DC, Kharidia KM, Mandyam CD. Transient Chemogenetic Inhibition of D1-MSNs in the Dorsal Striatum Enhances Methamphetamine Self-Administration. Brain Sciences. 2019; 9(11):330. https://doi.org/10.3390/brainsci9110330
Chicago/Turabian StyleOliver, Robert J., Dvijen C. Purohit, Khush M. Kharidia, and Chitra D. Mandyam. 2019. "Transient Chemogenetic Inhibition of D1-MSNs in the Dorsal Striatum Enhances Methamphetamine Self-Administration" Brain Sciences 9, no. 11: 330. https://doi.org/10.3390/brainsci9110330
APA StyleOliver, R. J., Purohit, D. C., Kharidia, K. M., & Mandyam, C. D. (2019). Transient Chemogenetic Inhibition of D1-MSNs in the Dorsal Striatum Enhances Methamphetamine Self-Administration. Brain Sciences, 9(11), 330. https://doi.org/10.3390/brainsci9110330