MyomiRNAs Dysregulation in ALS Rehabilitation

Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Selection
2.2. Patient Rehabilitation Protocol
2.3. Sera Collection
2.4. RNA Extraction and MiRNA Quantification
2.5. Statistical Analysis
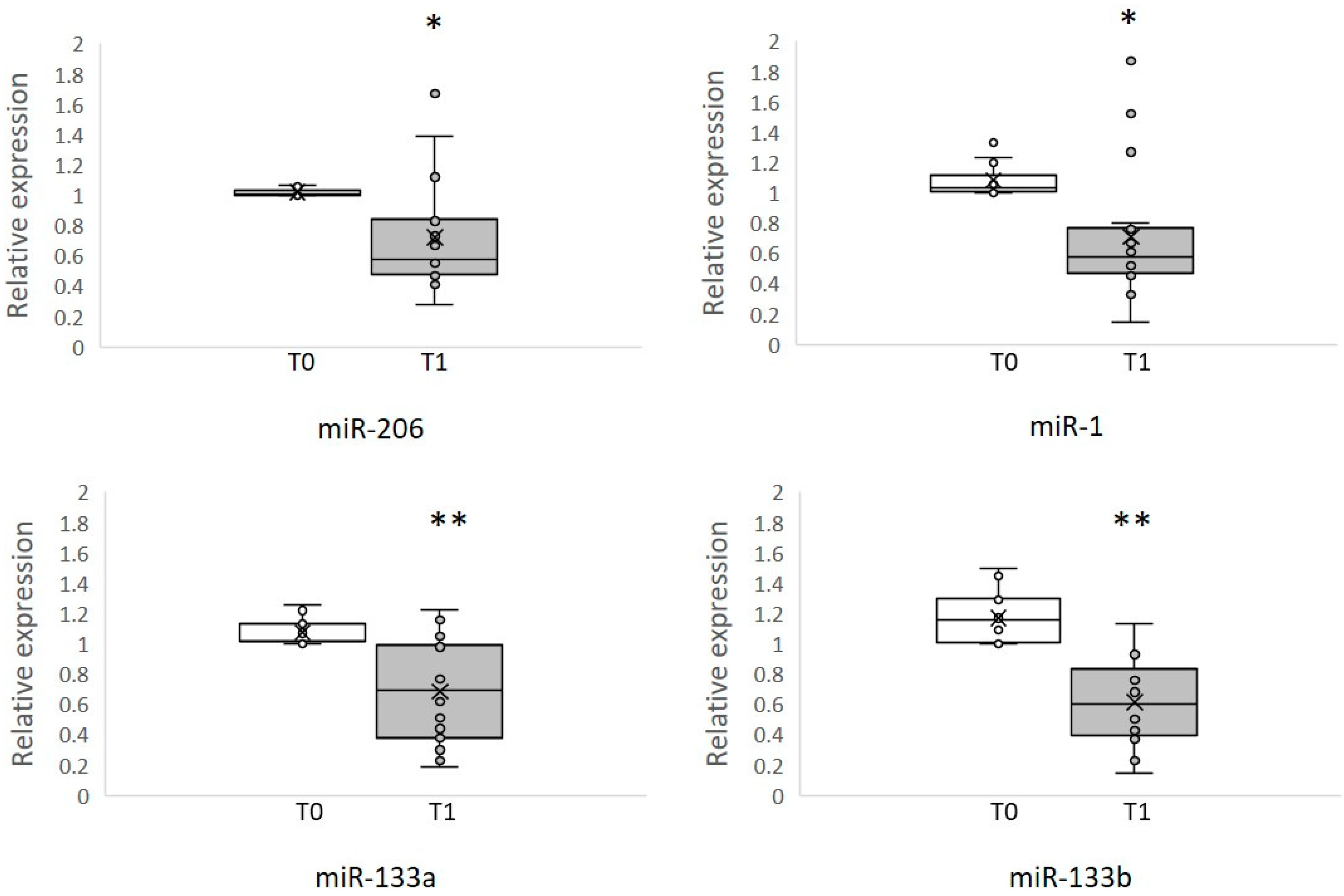
3. Results
3.1. Patient Cohort and Selection
3.2. Circulating MyomiRNAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Robberecht, W.; Philips, T. The changing scene of amyotrophic lateral sclerosis. Nat. Rev. Neurosci. 2013, 14, 248–264. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Bravver, E.K.; Langford, V.L.; Bockenek, W.L.; Lindblom, S.S. Stage of prolonged survival in ALS. Lancet Neurol. 2018, 17, 579. [Google Scholar] [CrossRef]
- Ferraiuolo, L.; Kirby, J.; Grierson, A.J.; Shaw, P.J.; Ferraiuolo, L.; Kirby, J.; Grierson, A.J.; Sendtner, M.; Shaw, P.J. Molecular pathways of motor neuron injury in amyotrophic lateral sclerosis. Nat. Rev. Neurol. 2011, 7, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.P.; Brown, R.H.; Cleveland, D.W. Decoding ALS: From genes to mechanism. Nature 2016, 539, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.H.; Al-Chalabi, A. Amyotrophic Lateral Sclerosis. N. Engl. J. Med. 2017, 377, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Recabarren-Leiva, D.; Alarcón, M. New insights into the gene expression associated to amyotrophic lateral sclerosis. Life Sci. 2018, 193, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, J.M.; Manzano, R.; Oliván, S.; Zaragoza, P.; García-Redondo, A.; Osta, R. MicroRNA-206: A potential circulating biomarker candidate for amyotrophic lateral sclerosis. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Scaramozza, A.; Marchese, V.; Papa, V.; Salaroli, R.; Sorarù, G.; Angelini, C.; Cenacchi, G. Skeletal Muscle Satellite Cells in Amyotrophic Lateral Sclerosis. Ultrastruct. Pathol. 2014, 38, 295–302. [Google Scholar] [CrossRef]
- Dadon-Nachum, M.; Melamed, E.; Offen, D. The “Dying-Back” Phenomenon of Motor Neurons in ALS. J. Mol. Neurosci. 2011, 43, 470–477. [Google Scholar] [CrossRef]
- Boyer, J.G.; Ferrier, A.; Kothary, R. More than a bystander: The contributions of intrinsic skeletal muscle defects in motor neuron diseases. Front. Physiol. 2013, 4. [Google Scholar] [CrossRef]
- Moloney, E.B.; de Winter, F.; Verhaagen, J. ALS as a distal axonopathy: Molecular mechanisms affecting neuromuscular junction stability in the presymptomatic stages of the disease. Front. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Merico, A.; Cavinato, M.; Gregorio, C.; Lacatena, A.; Gioia, E.; Piccione, F.; Angelini, C. Effects of combined endurance and resistance training in Amyotrophic Lateral Sclerosis: A pilot, randomized, controlled study. Eur. J. Transl. Myol. 2018, 28, 132–140. [Google Scholar] [CrossRef]
- Bhaskaran, M.; Mohan, M. MicroRNAs: History, Biogenesis, and Their Evolving Role in Animal Development and Disease. Vet. Pathol. 2014, 51, 759–774. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef]
- Tasca, E.; Pegoraro, V.; Merico, A.; Angelini, C. Circulating microRNAs as biomarkers of muscle differentiation and atrophy in ALS. Clin. Neuropathol. 2016, 35. [Google Scholar] [CrossRef] [PubMed]
- Pegoraro, V.; Merico, A.; Angelini, C. Micro-RNAs in ALS muscle: Differences in gender, age at onset and disease duration. J. Neurol. Sci. 2017, 380, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Volonte, C.; Apolloni, S.; Parisi, C. MicroRNAs: Newcomers into the ALS picture. CNS Neurol. Disord. Drug Targets 2015, 14, 194–207. [Google Scholar] [CrossRef]
- Rajgor, D. Macro roles for microRNAs in neurodegenerative diseases. Non-Coding RNA Res. 2018, 3, 154–159. [Google Scholar] [CrossRef]
- Roser, A.E.; Caldi Gomes, L.; Schünemann, J.; Maass, F.; Lingor, P. Circulating miRNAs as Diagnostic Biomarkers for Parkinson’s Disease. Front. Neurosci. 2018, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Horak, M.; Novak, J.; Bienertova-Vasku, J. Muscle-specific microRNAs in skeletal muscle development. Dev. Biol. 2016, 410, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Alexander, M.S.; Kunkel, L.M.; Cell, S.; Children, B. Skeletal Muscle MicroRNAs: Their Diagnostic and Therapeutic Potential in Human Muscle Diseases. J. Neuromuscul. Dis. 2015, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Townley-Tilson, W.H.D.; Callis, T.E.; Wang, D. MicroRNAs 1, 133, and 206: Critical factors of skeletal and cardiac muscle development, function, and disease. Int. J. Biochem. Cell Biol. 2010, 42, 1252–1255. [Google Scholar] [CrossRef] [PubMed]
- MCCarthy, J. MicroRNA-206: The skeletal muscle-specific myomiR. Biochim. Biophys. Acta-Gene Regul. Mech. 2008, 1779, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.-Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Romero, C.; Hur, J.; Lunn, J.S.; Paez-Colasante, X.; Bender, D.E.; Yung, R.; Sakowski, S.A.; Feldman, E.L. Expression of microRNAs in human post-mortem amyotrophic lateral sclerosis spinal cords provides insight into disease mechanisms. Mol. Cell. Neurosci. 2016, 71, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.H.; Valdez, G.; Moresi, V.; Qi, X.; McAnally, J.; Elliott, J.L.; Bassel-Duby, R.; Sanes, J.R.; Olson, E.N. MicroRNA-206 Delays ALS Progression and Promotes Regeneration of Neuromuscular Synapses in Mice. Science 2009, 326, 1549–1554. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W. Circulating MicroRNA Biomarker Studies: Pitfalls and Potential Solutions. Clin. Chem. 2015, 61, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Miller, R.G.; Swash, M.; Munsat, T.L. World Federation of Neurology Research Group on Motor Neuron Diseases El Escorial revisited: Revised criteria for the diagnosis of amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Other Motor Neuron Disord. 2000, 1, 293–299. [Google Scholar] [CrossRef]
- Li, Y.; Kowdley, K.V. Method for microRNA isolation from clinical serum samples. Anal. Biochem. 2012, 431, 69–75. [Google Scholar] [CrossRef]
- Bromberg, M.B. Amyotrophic Lateral Sclerosis Treatment Update More treatments are available but with largely subclinical effects. Prat. Neurol. 2018, 66–67. [Google Scholar]
- Di Pietro, L.; Lattanzi, W.; Bernardini, C. Skeletal muscle microRNAs as key players in the pathogenesis of amyotrophic lateral sclerosis. Int. J. Mol. Sci. 2018, 19, 1534. [Google Scholar] [CrossRef] [PubMed]
- Aitkens, S.G.; McCrory, M.A.; Kilmer, D.D.; Bernauer, E.M. Moderate resistance exercise program: Its effect in slowly progressive neuromuscular disease. Arch. Phys. Med. Rehabil. 1993, 74, 711–715. [Google Scholar] [CrossRef]
- Abresch, R.T.; Carter, G.T.; Han, J.J.; McDonald, C.M. Exercise in Neuromuscular Diseases. Phys. Med. Rehabil. Clin. N. Am. 2012, 23, 653–673. [Google Scholar] [CrossRef]
- Chio, A.; Calvo, A.; Dossena, M.; Ghiglione, P.; Mutani, R.; Mora, G. ALS in Italian professional soccer players: The risk is still present and could be soccer-specific. Amyotroph. Lateral Scler. 2009, 10, 205–209. [Google Scholar] [CrossRef]
- Beghi, E.; Logroscino, G.; Chiò, A.; Hardiman, O.; Millul, A.; Mitchell, D.; Swingler, R.; Traynor, B.J. Amyotrophic lateral sclerosis, physical exercise, trauma and sports: Results of a population-based pilot case-control study. Amyotroph. Lateral Scler. 2010, 11, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Gurney, M.E.; Cutting, F.B.; Zhai, P.; Doble, A.; Taylor, C.P.; Andrus, P.K.; Hall, E.D. Benefit of vitamin E, riluzole, and gabapentin in a transgenic model of familial amyotrophic lateral sclerosis. Ann. Neurol. 1996, 39, 147–157. [Google Scholar] [CrossRef]
- Drory, V.E.; Goltsman, E.; Reznik, J.G.; Mosek, A.; Korczyn, A.D. The value of muscle exercise in patients with amyotrophic lateral sclerosis. J. Neurol. Sci. 2001, 191, 133–137. [Google Scholar] [CrossRef]
- Bello-Haas, V.D.; Florence, J.M.; Kloos, A.D.; Scheirbecker, J.; Lopate, G.; Hayes, S.M.; Pioro, E.P.; Mitsumoto, H. A randomized controlled trial of resistance exercise in individuals with ALS. Neurology 2007, 68, 2003–2007. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.C.; Alves, M.; Nogueira, A.; Evangelista, T.; Carvalho, J.; Coelho, A.; de Carvalho, M.; Sales-Luís, M.L. Can amyotrophic lateral sclerosis patients with respiratory insufficiency exercise? J. Neurol. Sci. 1999, 169, 69–75. [Google Scholar] [CrossRef]
- Clawson, L.L.; Cudkowicz, M.; Krivickas, L.; Brooks, B.R.; Sanjak, M.; Allred, P.; Atassi, N.; Swartz, A.; Steinhorn, G.; Uchil, A.; et al. Neals consortium A randomized controlled trial of resistance and endurance exercise in amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Front. Degener. 2018, 19, 250–258. [Google Scholar] [CrossRef]
- Lunetta, C.; Lizio, A.; Sansone, V.A.; Cellotto, N.M.; Maestri, E.; Bettinelli, M.; Gatti, V.; Melazzini, M.G.; Meola, G.; Corbo, M. Strictly monitored exercise programs reduce motor deterioration in ALS: Preliminary results of a randomized controlled trial. J. Neurol. 2016, 263, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.R.; Kent-Braun, J.A.; Majumdar, S.; Huang, Y.; Mynhier, M.; Weiner, M.W.; Miller, R.G. Physiology of fatigue in amyotrophic lateral sclerosis. Neurology 1995, 45, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H. MicroRNA in myogenesis and muscle atrophy. Curr. Opin. Clin Nutr Metab. Care 2013, 16, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Wang, Y.; Li, Y.; Cui, L.; Zhao, Y.; Zhao, B.; Li, K. MiR-206, a key modulator of skeletal muscle development and disease. Int. J. Biol. Sci. 2015, 11, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Valsecchi, V.; Boido, M.; De Amicis, E.; Piras, A.; Vercelli, A. Expression of Muscle-Specific MiRNA 206 in the Progression of Disease in a Murine SMA Model. PLoS ONE 2015, 10, e0128560. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.; Jørgensen, L.H.; Bech, R.D.; Frandsen, U.; Schrøder, H.D. Skeletal Muscle Remodelling as a Function of Disease Progression in Amyotrophic Lateral Sclerosis. BioMed Res. Int. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Chen, J. MicroRNAs in skeletal myogenesis. Cell Cycle 2011, 10, 441–448. [Google Scholar] [CrossRef]
- Nielsen, S.; Scheele, C.; Yfanti, C.; Thorbjörnåkerström, T.T.; Nielsen, A.R.; Pedersen, B.K.; Laye, M.; Nielsen, S. Muscle specific microRNAs are regulated by endurance exercise in human skeletal muscle. J. Physiol. 2010, 588, 4029–4037. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.P.; Lamon, S.; Boon, H.; Wada, S.; Güller, I.; Brown, E.L.; Chibalin, A.V.; Zierath, J.R.; Snow, R.J.; Stepto, N.; et al. Regulation of miRNAs in human skeletal muscle following acute endurance exercise and short-term endurance training. Authors J. Physiol. C 2013, 591, 4637–4653. [Google Scholar] [CrossRef]


{kind=link}
| ALS Features | Mean ± SD |
|---|---|
| Age (years) | 61.1 ± 12.8 |
| Sex M/F | 11/7 |
| Disease duration (years) | 4.3 ± 3 |
| ALSFRS-R | 34.6 ± 4.9 |
| Patient | Age | Onset | Sex | ALSFRS-R T0 | ALSFRS-R T1 | FSS T0 | FSS T1 | FIM T0 | FIM T1 | Barthel T0 | Barthel T1 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pt 01 | 74 | N.A. | M | 29 | 29 | 5.40 | 3.90 | 78 | 80 | 45 | 60 |
| Pt 02 | 51 | 39 | M | 40 | 41 | 5.50 | 3.90 | 77 | 82 | 50 | 55 |
| Pt 03 | 57 | 53 | F | 38 | 40 | 5.60 | 4.20 | 86 | 90 | 55 | 70 |
| Pt 04 | 79 | 72 | M | 29 | 29 | 5.30 | 5.20 | 85 | 87 | 30 | 40 |
| Pt 05 | 50 | 45 | M | 40 | 41 | 5.50 | 3.90 | 77 | 82 | 50 | 55 |
| Pt 06 | 61 | 58 | M | 31 | 31 | 5.50 | 5.50 | 37 | 36 | N.A. | N.A. |
| Pt 07 | 45 | 40 | F | 28 | 28 | 5.50 | 5.50 | 88 | 91 | 70 | 80 |
| Pt 08 | 33 | 30 | F | 32 | 32 | 5.20 | 4.80 | 65 | 69 | 35 | 40 |
| Pt 09 | 76 | 70 | F | 34 | 34 | 5.50 | 4.90 | N.A. | 65 | 20 | 35 |
| Pt 10 | 44 | 43 | M | 32 | 32 | 5.40 | 5.00 | 83 | 87 | 60 | 65 |
| Pt 11 | 71 | 69 | M | 36 | 36 | 5.50 | 5.10 | N.A. | 69 | 20 | 35 |
| Pt 12 | 71 | 57 | M | 32 | 33 | 5.40 | 3.90 | 78 | 84 | 45 | 55 |
| Pt 13 | 61 | 57 | F | 33 | 34 | 5.10 | 4.00 | 60 | 70 | 35 | 45 |
| Pt 14 * | 71 | 66 | M | 28 | 28 | 5.60 | 4.40 | 65 | N.A. | 40 | 45 |
| Pt 15 | 65 | 63 | F | 39 | 40 | 4.50 | 3.20 | 83 | 85 | 50 | 55 |
| Pt 16 | 65 | 65 | F | 42 | 43 | 5.40 | 4.20 | 73 | 79 | 45 | 50 |
| Pt 17 | 72 | 62 | M | 36 | 36 | 5.60 | 4.00 | 81 | 89 | 60 | 90 |
| Pt 18 | 54 | 51 | M | 43 | 44 | 4.80 | 3.20 | 80 | 88 | 60 | 70 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pegoraro, V.; Merico, A.; Angelini, C. MyomiRNAs Dysregulation in ALS Rehabilitation. Brain Sci. 2019, 9, 8. https://doi.org/10.3390/brainsci9010008
Pegoraro V, Merico A, Angelini C. MyomiRNAs Dysregulation in ALS Rehabilitation. Brain Sciences. 2019; 9(1):8. https://doi.org/10.3390/brainsci9010008
Chicago/Turabian StylePegoraro, Valentina, Antonio Merico, and Corrado Angelini. 2019. "MyomiRNAs Dysregulation in ALS Rehabilitation" Brain Sciences 9, no. 1: 8. https://doi.org/10.3390/brainsci9010008
APA StylePegoraro, V., Merico, A., & Angelini, C. (2019). MyomiRNAs Dysregulation in ALS Rehabilitation. Brain Sciences, 9(1), 8. https://doi.org/10.3390/brainsci9010008