A Multidisciplinary Approach Reveals an Age-Dependent Expression of a Novel Bioactive Peptide, Already Involved in Neurodegeneration, in the Postnatal Rat Forebrain

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
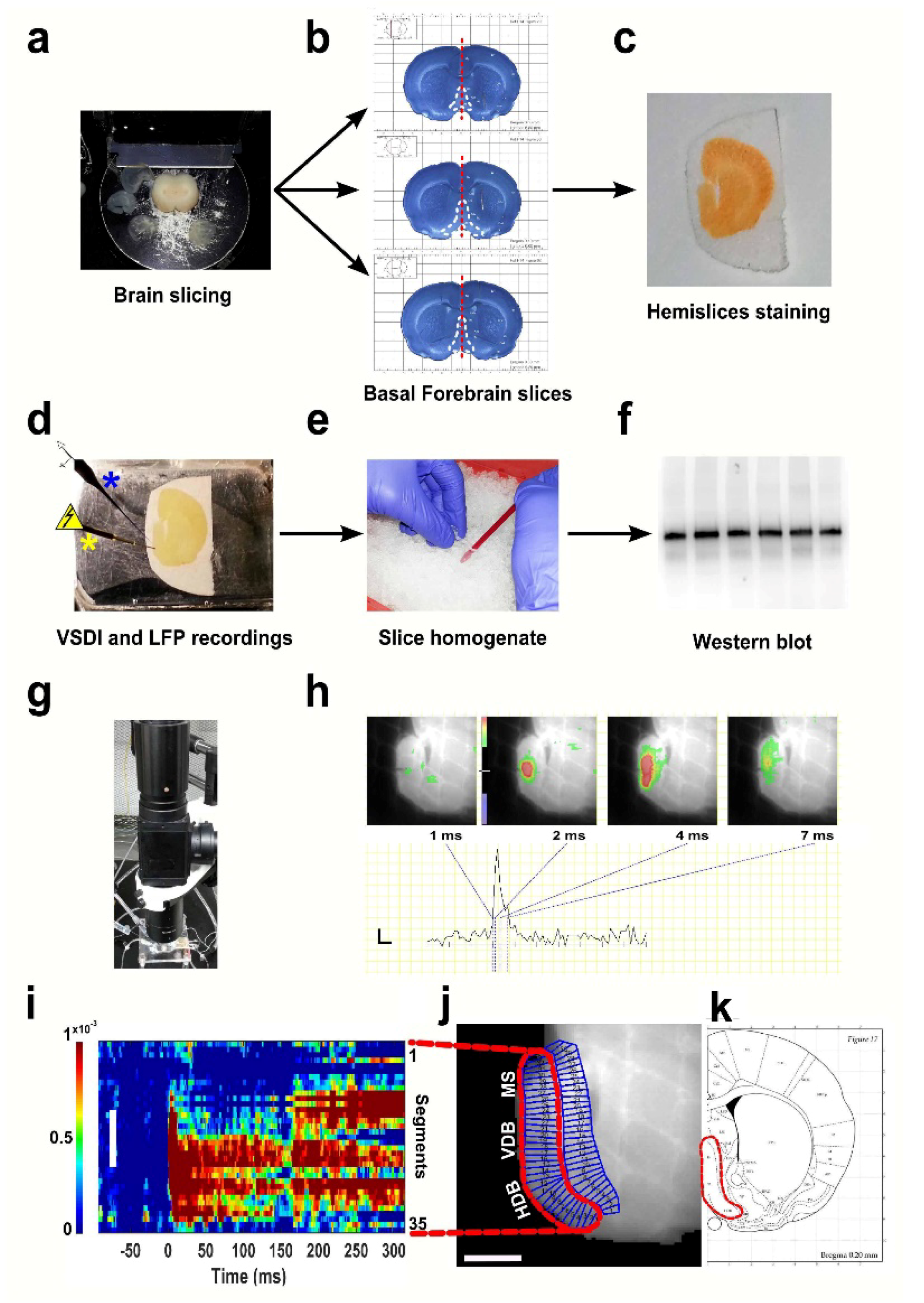
2.2. Brain Slice Preparation
2.3. Optical Recording Method
2.4. Field Recordings
2.5. Data Analysis
2.6. Tissue Homogenisation
2.7. Co-Immunoprecipitation
2.8. Protein Measurement and Western Blot Analysis
2.9. Image Processing
2.10. Statistical Analysis
3. Results
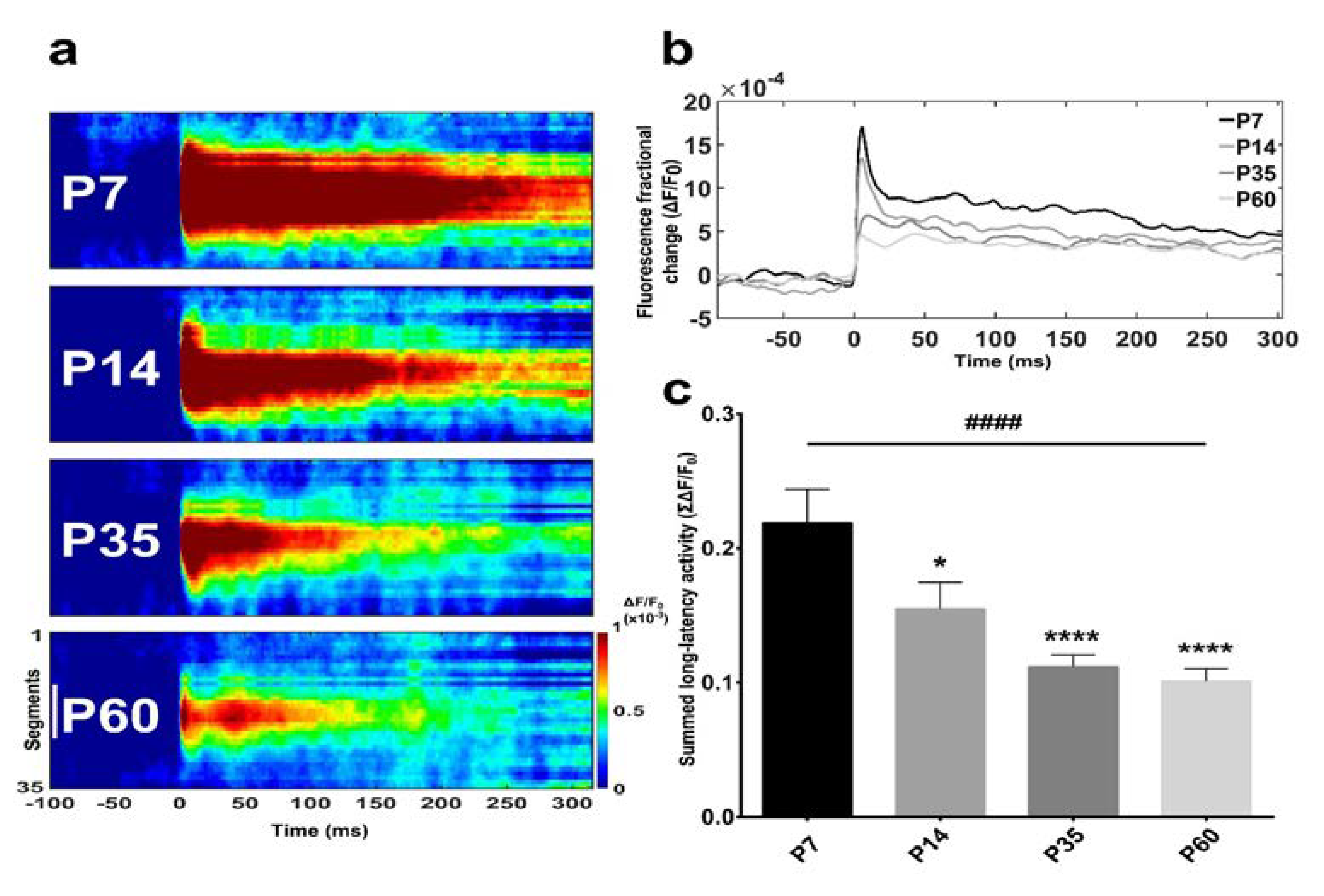
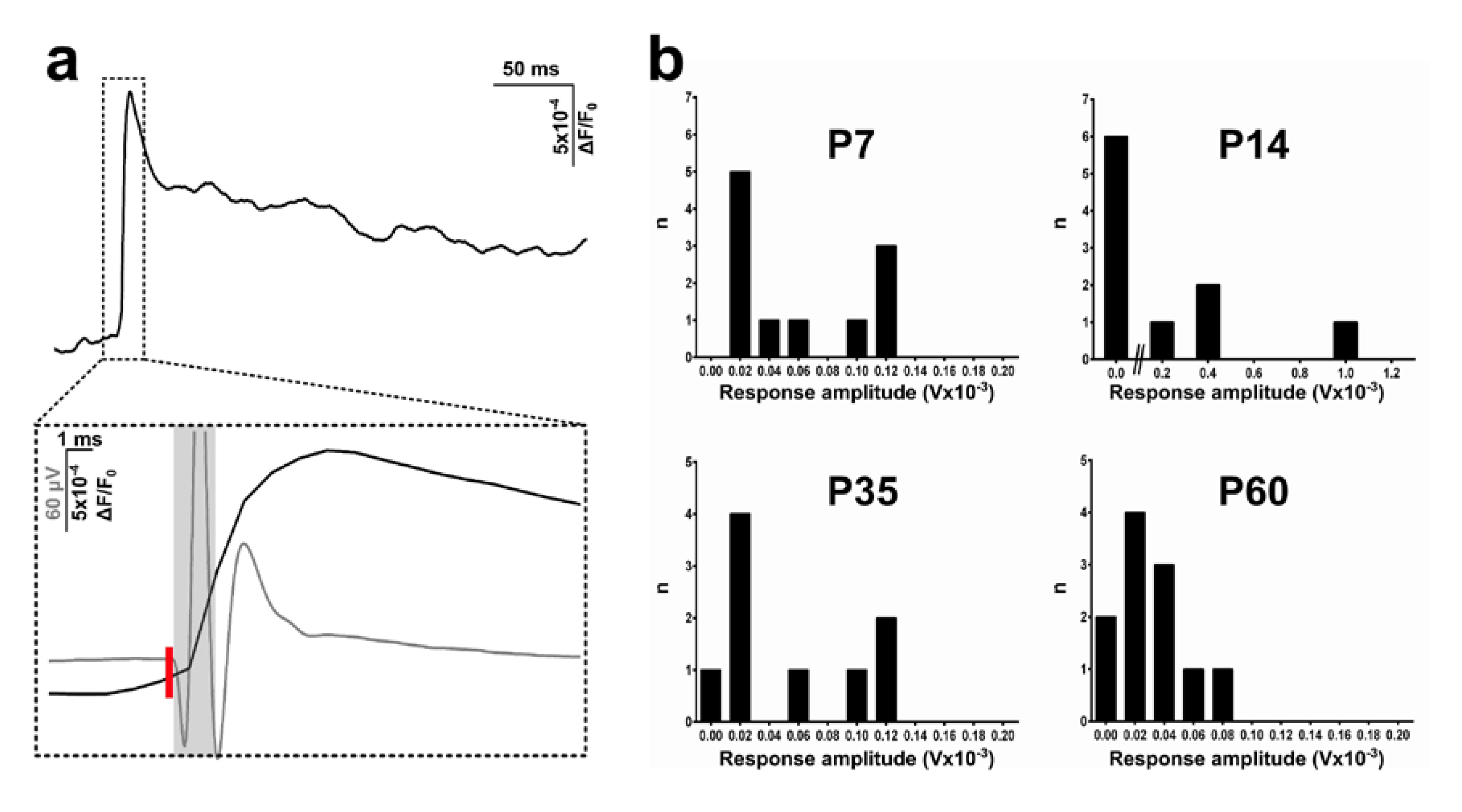
3.1. Basal Forebrain Population Activity Measured with Optical Imaging and Electrophysiology in Postnatal Development
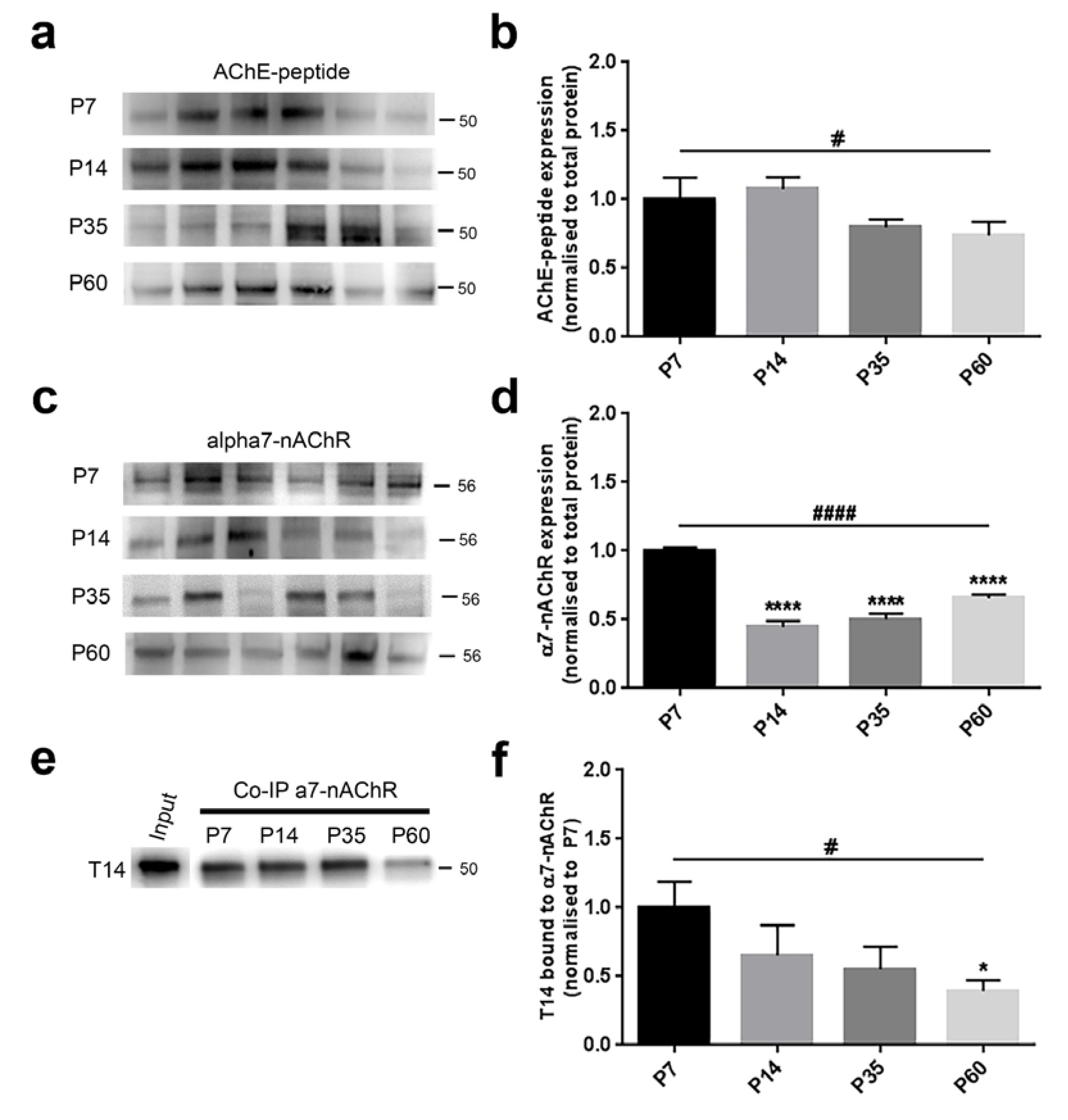
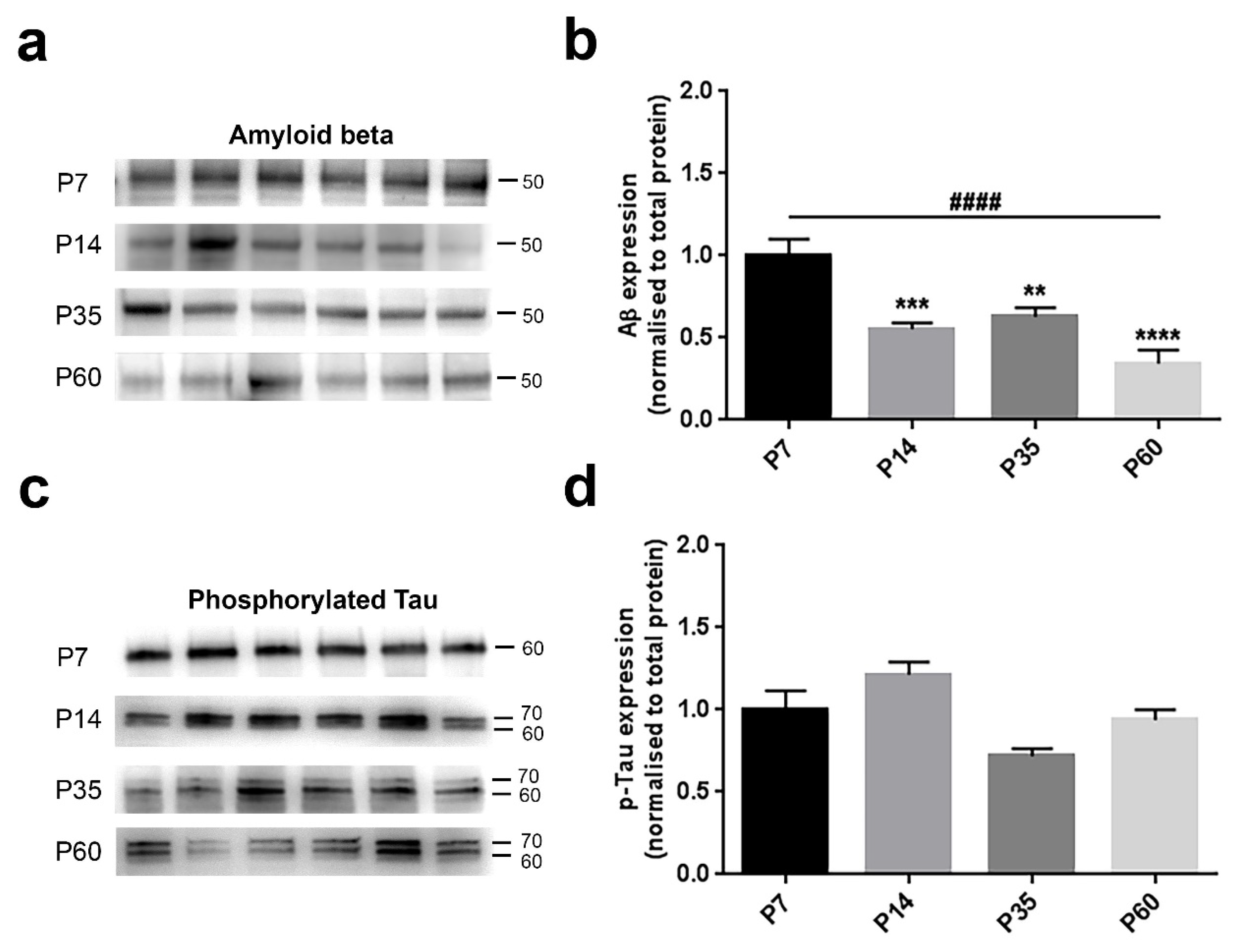
3.2. Protein Levels Change Differentially during Postnatal Development
4. Discussion
4.1. Neurodegeneration as a Recapitulation of Development: A Common T14 Signalling System
4.2. Optical Imaging of Age-Dependent Neuronal Activity
4.3. Parallel Developmental Changes in Levels of T14 and Related Key Proteins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mesulam, M.M.; Mufson, E.J.; Levey, A.I.; Wainer, B.H. Cholinergic innervation of cortex by the basal forebrain: Cytochemistry and cortical connections of the septal area, diagonal band nuclei, nucleus basalis (substantia innominata), and hypothalamus in the rhesus monkey. J. Comp. Neurol. 1983, 214, 170–197. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, E.C.; Ananth, M.; Talmage, D.A.; Role, L.W. Basal Forebrain Cholinergic Circuits and Signaling in Cognition and Cognitive Decline. Neuron 2016, 91, 1199–1218. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, T.W.; Nathan Spreng, R.; Weiner, M.W.; Aisen, P.; Petersen, R.; Jack, C.R.; Jagust, W.; Trojanowki, J.Q.; Toga, A.W.; Beckett, L.; et al. Basal forebrain degeneration precedes and predicts the cortical spread of Alzheimer’s pathology. Nat. Commun. 2016, 7, 13249. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Woolf, N.J.; Butcher, L.L. Postnatal development of cholinergic neurons in the rat: I. Forebrain. Brain Res. Bull. 1991, 27, 767–789. [Google Scholar] [CrossRef]
- Thal, L.J.; Gilbertson, E.; Armstrong, D.M.; Gage, F.H. Development of the basal forebrain cholinergic system: Phenotype expression prior to target innervation. Neurobiol. Aging 1992, 13, 67–72. [Google Scholar] [CrossRef]
- Silman, I.; Sussman, J.L. Acetylcholinesterase: “Classical” and “non-classical” functions and pharmacology. Curr. Opin. Pharmacol. 2005, 5, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Soreq, H.; Seidman, S. Acetylcholinesterase—New roles for an old actor. Nat. Rev. Neurosci. 2001, 2, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Arendt, T.; Brückner, M.K.; Lange, M.; Bigl, V. Changes in acetylcholinesterase and butyrylcholinesterase in Alzheimer’s disease resemble embryonic development-A study of molecular forms. Neurochem. Int. 1992, 21, 381–396. [Google Scholar] [CrossRef]
- Greenfield, S.; Vaux, D.J. Commentary Parkinson’s Disease, Alzheimer’ S Disease and Motor Neurone Disease: Identifying a Common Mechanism. Neuroscience 2002, 113, 485–492. [Google Scholar] [CrossRef]
- Greenfield, S.A.; Day, T.; Mann, E.O.; Bermudez, I. A novel peptide modulates a 7 nicotinic receptor responses: Implications for a possible trophic-toxic mechanism within the brain. J. Neurochem. 2004, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Cottingham, M.G.; Voskuil, J.L.A.; Vaux, D.J.T. The Intact Human Acetylcholinesterase C-Terminal Oligomerization Domain Is α-Helical in Situ and in Isolation, but a Shorter Fragment Forms β-Sheet-Rich Amyloid Fibrils and Protofibrillar Oligomers †. Biochemistry 2003, 42, 10863–10873. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, S. Discovering and targeting the basic mechanism of neurodegeneration: The role of peptides from the C-terminus of acetylcholinesterase: Non-hydrolytic effects of ache: The actions of peptides derived from the C-terminal and their relevance to neurodegenerat. Chem. Biol. Interact. 2013, 203, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Eimerl, S.; Schramm, M. The Quantity of Calcium that Appears to Induce Neuronal Death. J. Neurochem. 1994, 62, 1223–1226. [Google Scholar] [CrossRef] [PubMed]
- García-Ayllón, M.-S.; David, S.H.; Avila, J.; Sáez-Valero, J. Revisiting the role of acetylcholinesterase in Alzheimer’s disease: Cross-talk with P-tau and β-amyloid. Front. Mol. Neurosci. 2011, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ratés, S.; Morrill, P.; Tu, H.; Pottiez, G.; Badin, A.S.; Tormo-Garcia, C.; Heffner, C.; Coen, C.W.; Greenfield, S.A. (I) Pharmacological profiling of a novel modulator of the α7 nicotinic receptor: Blockade of a toxic acetylcholinesterase-derived peptide increased in Alzheimer brains. Neuropharmacology 2016, 105, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Badin, A.S.; Morrill, P.; Devonshire, I.M.; Greenfield, S.A. (II) Physiological profiling of an endogenous peptide in the basal forebrain: Age-related bioactivity and blockade with a novel modulator. Neuropharmacology 2016, 105, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Brai, E.; Stuart, S.; Badin, A.-S.; Greenfield, S.A. A novel ex vivo model to investigate the underlying mechanisms in Alzheimer’s disease. Front. Cell. Neurosci. 2017, 11, 291. [Google Scholar] [CrossRef] [PubMed]
- Grinvald, A.; Hildesheim, R. VSDI: A new era in functional imaging of cortical dynamics. Nat. Rev. Neurosci. 2004, 5, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Badin, A.-S.P.; Fermani, F.; Greenfield, S.A. The Features and Functions of Neuronal Assemblies: Possible Dependency on Mechanisms beyond Synaptic Transmission. Front. Neural Circuits 2017, 10, 114. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, S.A.; Badin, A.-S.; Ferrati, G.; Devonshire, I.M. Optical imaging of the rat brain suggests a previously missing link between top-down and bottom-up nervous system function. Neurophotonics 2017, 4, 31213. [Google Scholar] [CrossRef] [PubMed]
- Molas, S.; Dierssen, M. The role of nicotinic receptors in shaping and functioning of the glutamatergic system: A window into cognitive pathology. Neurosci. Biobehav. Rev. 2014, 2, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, E.; Small, D.H. Insights into the physiological function of the β-amyloid precursor protein: Beyond Alzheimer’s disease. J. Neurochem. 2014, 129, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Hiltunen, M.; Van Groen, T.; Jolkkonen, J. Functional roles of amyloid-β protein precursor and amyloid-β peptides: Evidence from experimental studies. J. Alzheimer’s Dis. 2009, 18, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Bramblett, G.T.; Goedert, M.; Jakes, R.; Merrick, S.E.; Trojanowski, J.Q.; Lee, V.M.Y. Abnormal tau phosphorylation at Ser396 in alzheimer’s disease recapitulates development and contributes to reduced microtubule binding. Neuron 1993, 10, 1089–1099. [Google Scholar] [CrossRef]
- Brai, E.; Simon, F.; Cogoni, A.; Greenfield, S.A. Modulatory effects of a novel cyclized peptide in reducing the expression of markers linked to Alzheimer’s disease. Front. Neurosci. 2018, 12, 362. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Brai, E.; Cogoni, A.; Greenfield, S.A. An Alternative Approach to Study Primary Events in Neurodegeneration Using Ex Vivo Rat Brain Slices Video Link. J. Vis. Exp. 2018, 134. [Google Scholar] [CrossRef]
- Tominaga, T.; Tominaga, Y.; Yamada, H.; Matsumoto, G.; Ichikawa, M. Quantification of optical signals with electrophysiological signals in neural activities of Di-4-ANEPPS stained rat hippocampal slices. J. Neurosci. Methods 2000, 102, 11–23. [Google Scholar] [CrossRef]
- Grandy, T.H.; Greenfield, S.A.; Devonshire, I.M. An evaluation of in vivo voltage-sensitive dyes: Pharmacological side effects and signal-to-noise ratios after effective removal of brain-pulsation artifacts. J. Neurophysiol. 2012, 108, 2931–2945. [Google Scholar] [CrossRef] [PubMed]
- Khazipov, R.; Zaynutdinova, D.; Ogievetsky, E.; Valeeva, G.; Mitrukhina, O.; Manent, J.-B.; Represa, A. Atlas of the Postnatal Rat Brain in Stereotaxic Coordinates. Front. Neuroanat. 2015, 9, 161. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, E.B.; Johnson, B.N.; McCoy, A.J.; Trippa, L.; Cohen, A.S.; Marsh, E.D. A toolbox for spatiotemporal analysis of voltage-sensitive dye imaging data in brain slices. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Aldridge, G.M.; Podrebarac, D.M.; Greenough, W.T.; Weiler, I.J. The use of total protein stains as loading controls: An alternative to high-abundance single protein controls in semi-quantitative immunoblotting. J. Neurosci. Methods 2008, 172, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.A.; An, J.; Peller, D.; Bowser, R. Total protein is an effective loading control for cerebrospinal fluid western blots. J. Neurosci. Methods 2015, 251, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Guo, J.; Xu, H.-B.; Huang, R.; Shao, W.; Yang, L.; Wang, M.; Chen, J.; Xie, P. Direct Blue 71 staining as a destaining-free alternative loading control method for Western blotting. Electrophoresis 2013, 34, 2234–2239. [Google Scholar] [CrossRef] [PubMed]
- Using ImageJ to Quantify Protein Bands on a PAGE Gel. Available online: https://www.youtube.com/watch?v=JlR5v-DsTds (accessed on 27 Marth 2014).
- García-Ayllón, M.S.; Riba-Llena, I.; Serra-Basante, C.; Alom, J.; Boopathy, R.; Sáez-Valero, J. Altered levels of acetylcholinesterase in Alzheimer plasma. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Egorov, A.V.; Draguhn, A. Development of coherent neuronal activity patterns in mammalian cortical networks: Common principles and local hetereogeneity. Mech. Dev. 2013, 130, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Palmer, C.R.; Seidemann, E. The relationship between voltage-sensitive dye imaging signals and spiking activity of neural populations in primate V1. J. Neurophysiol. 2012, 107, 3281–3295. [Google Scholar] [CrossRef] [PubMed]
- Chemla, S.; Chavane, F. Voltage-sensitive dye imaging: Technique review and models. J. Physiol. Paris 2010, 104, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.P.; Dengler, C.G.; Frausto, S.F.; Putt, M.E.; Yue, C.; Takano, H.; Coulter, D. A Protracted postnatal development of sparse, specific dentate granule cell activation in the mouse hippocampus. J. Neurosci. 2013, 33, 2947–2960. [Google Scholar] [CrossRef] [PubMed]
- Cottingham, M.G.; Hollinshead, M.S.; Vaux, D.J.T. Amyloid fibril formation by a synthetic peptide from a region of human acetylcholinesterase that is homologous to the Alzheimer’s amyloid-β peptide. Biochemistry 2002, 41, 13539–13547. [Google Scholar] [CrossRef] [PubMed]
- Tribollet, E.; Bertrand, D.; Marguerat, A.; Raggenbass, M. Comparative distribution of nicotinic receptor subtypes during development, adulthood and aging: An autoradiographic study in the rat brain. Neuroscience 2004, 124, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Broide, R.S.; Leslie, F.M. The alpha7 nicotinic acetylcholine receptor in neuronal plasticity. Mol. Neurobiol. 1999, 20, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Aguayo, M.C.; Silva-Lucero, M.C.; García, U. Physiological Role of Amyloid Beta in Neural Cells: The Cellular Trophic Activity. Neurochemistry 2014, 1–26. [Google Scholar] [CrossRef]
- Arendt, T.; Stieler, J.; Ueberham, U. Is sporadic Alzheimer′s disease a developmental disorder? J. Neurochem. 2017, 1–13. [Google Scholar] [CrossRef]
- Sadot, E.; Gurwitz, D.; Barg, J.; Behar, L.; Ginzburg, I.; Fisher, A. Activation of m1 muscarinic acetylcholine receptor regulates tau phosphorylation in transfected PC12 cells. J. Neurochem. 1996, 66, 877–880. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Crowther, R.A.; Garner, C.C. Molecular characterization of microtubule-associated proteins tau and map2. Trends Neurosci. 1991, 14, 193–199. [Google Scholar] [CrossRef]
- Georgieff, I.S.; Liem, R.K.; Mellado, W.; Nunez, J.; Shelanski, M.L. High molecular weight tau: Preferential localization in the peripheral nervous system. J. Cell Sci. 1991, 100, 55–60. [Google Scholar] [PubMed]
- Dickinson, J.A.; Hanrott, K.E.; Mok, M.H.S.; Kew, J.N.C.; Wonnacott, S. Differential coupling of alpha7 and non-alpha7 nicotinic acetylcholine receptors to calcium-induced calcium release and voltage-operated calcium channels in PC12 cells. J. Neurochem. 2007, 100, 1089–1096. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrati, G.; Brai, E.; Stuart, S.; Marino, C.; Greenfield, S.A. A Multidisciplinary Approach Reveals an Age-Dependent Expression of a Novel Bioactive Peptide, Already Involved in Neurodegeneration, in the Postnatal Rat Forebrain. Brain Sci. 2018, 8, 132. https://doi.org/10.3390/brainsci8070132
Ferrati G, Brai E, Stuart S, Marino C, Greenfield SA. A Multidisciplinary Approach Reveals an Age-Dependent Expression of a Novel Bioactive Peptide, Already Involved in Neurodegeneration, in the Postnatal Rat Forebrain. Brain Sciences. 2018; 8(7):132. https://doi.org/10.3390/brainsci8070132
Chicago/Turabian StyleFerrati, Giovanni, Emanuele Brai, Skye Stuart, Celia Marino, and Susan A. Greenfield. 2018. "A Multidisciplinary Approach Reveals an Age-Dependent Expression of a Novel Bioactive Peptide, Already Involved in Neurodegeneration, in the Postnatal Rat Forebrain" Brain Sciences 8, no. 7: 132. https://doi.org/10.3390/brainsci8070132
APA StyleFerrati, G., Brai, E., Stuart, S., Marino, C., & Greenfield, S. A. (2018). A Multidisciplinary Approach Reveals an Age-Dependent Expression of a Novel Bioactive Peptide, Already Involved in Neurodegeneration, in the Postnatal Rat Forebrain. Brain Sciences, 8(7), 132. https://doi.org/10.3390/brainsci8070132