Brain Connectivity Networks and the Aesthetic Experience of Music


Abstract
1. Introduction
2. Neuroaesthetics of Music
3. Network Neuroscience and Connectomics
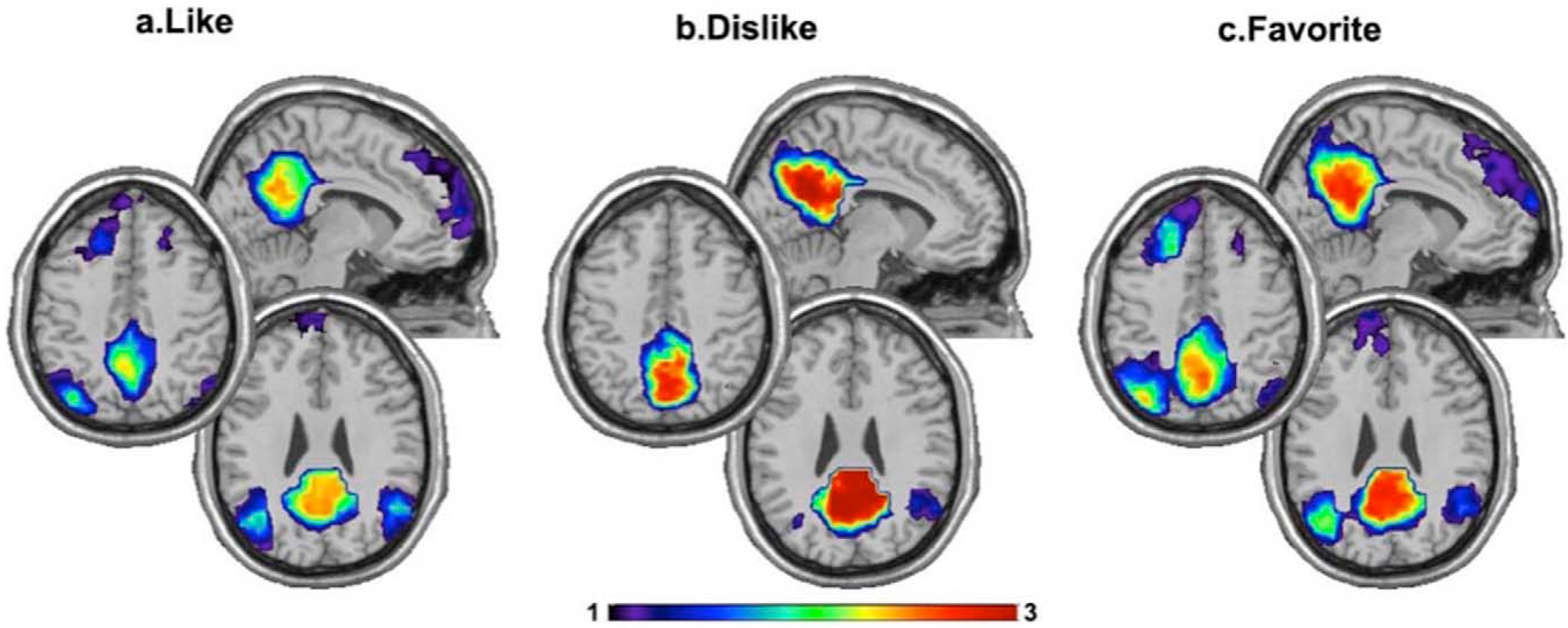
4. The Default Mode Network during Music Listening
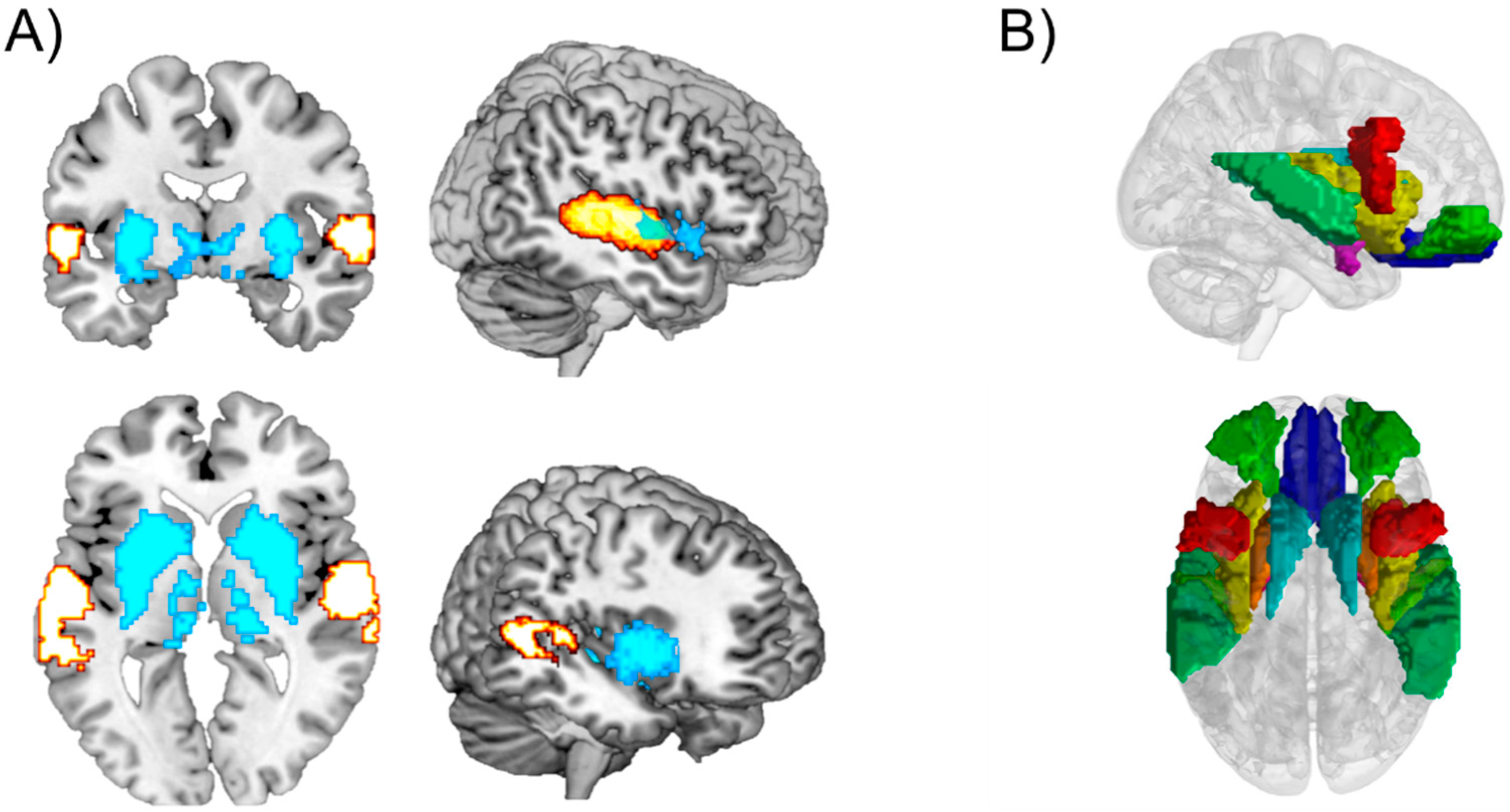
5. Brain Connections during the Experience of Musical Reward
6. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Reybrouck, M. Music cognition and real-time listening: Denotation, cue abstraction, route description and cognitive maps. Music. Sci. 2010, 2010, 187–202. [Google Scholar] [CrossRef]
- Maeder, C.; Reybrouck, M. (Eds.) Music, Analysis, Experience. New Perspectives in Musical Semiotics; Leuven University Press: Leuven, Belgium, 2015. [Google Scholar]
- Maeder, C.; Reybrouck, M. (Eds.) Sémiotique et Vécu Musical. Du sens à L’expérience, de L’expérience au sens; Leuven University Press: Leuven, Belgium, 2016. [Google Scholar]
- Laske, O. Music, Memory and Thought: Explorations in Cognitive Musicology; University Microfilms International: Ann Arbor, MI, USA, 1977. [Google Scholar]
- Honing, H. On the Growing Role of Observation, Formalization and Experimental Method in Musicology. Empir. Musicol. Rev. 2006, 1, 1–6. [Google Scholar] [CrossRef]
- Huron, D. Music and Mind: Foundations of Cognitive Musicology: Ernest Bloch Lecture; University of California: Berkeley, CA, USA, 1999. [Google Scholar]
- Lartillot, O.; Toiviainen, P. A Matlab toolbox for musical feature extraction from audio. In Proceedings of the 10th International Conference on Digital Audio Effects (DAFx-07), Bordeaux, France, 10–15 September 2007. [Google Scholar]
- Parncutt, R. Systematic Musicology and the History and Future of Western Musical Scholarship. J. Interdiscip. Music Stud. 2007, 1, 1–32. [Google Scholar]
- Pearce, M.T.; Wiggins, G.A. Expectation in melody: The influence of context and learning. Music Percept. 2006, 23, 377–405. [Google Scholar] [CrossRef]
- Purwins, H.; Herrera, P.; Grachten, M.; Hazan, A.; Marxer, R.; Serra, X. Computational Models of Music Perception and Cognitio I: The Perceptual and Cognitive Processing Chain. Phys. Life Rev. 2008, 5, 151–168. [Google Scholar] [CrossRef]
- Fazelpour, S.; Thompson, E. The Kantian brain: Brain dynamics from a neurophenomenological perspective. Curr. Opin. Neurobiol. 2015, 31, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Brattico, E. The neuroaesthetics of music: A research agenda coming of age. In The Oxford Handbook of Music and the Brain; Thaut, M., Hodges, D.A., Eds.; Oxford University Press: Oxford, UK; New York, NY, USA, 2017. [Google Scholar]
- Brattico, E.; Bogert, B.; Jacobsen, T. Toward a neural chronometry for the aesthetic experience of music. Front. Psychol. 2013, 4, 206. [Google Scholar] [CrossRef] [PubMed]
- Brattico, E.; Pearce, M. The Neuroaesthetics of Music. Psychol. Aesthet. Creat. Arts 2013, 7, 48–61. [Google Scholar] [CrossRef]
- Hodges, D.A. The neuroaesthetics of music. In The Oxford Handbook of Music Psychology; Hallam, S., Cross, I., Thaut, M., Eds.; Oxford University Press: Oxford, UK; New York, NY, USA, 2016; pp. 247–272. [Google Scholar]
- Reybrouck, M.; Brattico, E. Neuroplasticity beyond Sounds: Neural Adaptations Following Long-Term Musical Aesthetic Experiences. Brain Sci. 2015, 5, 69–91. [Google Scholar] [CrossRef] [PubMed]
- Reybrouck, M. Music as Environment: An Ecological and Biosemiotic Approach. Behav. Sci. 2015, 5, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Brattico, E. From pleasure to liking and back: Bottom-up and top-down neural routes to the aesthetic enjoyment. In Art, Aesthetics, and the Brain; Huston, J., Nadal, M., Mora, F., Agnati, L., Cela Conde, C.J., Eds.; Oxford University Press: Oxford, UK, 2015; pp. 303–318. [Google Scholar]
- Brattico, E.; Bogert, B.; Alluri, V.; Tervaniemi, M.; Eerola, T.; Jacobsen, T. It’s sad but I like it: The neural dissociation between musical emotions and liking in experts and laypersons. Front. Hum. Neurosci. 2015, 9, 676. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, S.; Istok, E.; Brattico, E.; Tervaniemi, M.; Huotilainen, M. The development of aesthetic responses to music and their underlying neural and psychological mechanisms. Cortex 2011, 47, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Pearce, M.T.; Zaidel, D.W.; Vartanian, O.; Skov, M.; Leder, H.; Chatterjee, A.; Nadal, M. Neuroaesthetics: The Cognitive Neuroscience of Aesthetic Experience. Perspect. Psychol. Sci. 2016, 11, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Pelowski, M.; Markey, P.S.; Forster, M.; Gerger, G.; Leder, H. Move me, astonish me... delight my eyes and brain: The Vienna Integrated Model of top-down and bottom-up processes in Art Perception (VIMAP) and corresponding affective, evaluative, and neurophysiological correlates. Phys. Life Rev. 2017, 21, 80–125. [Google Scholar] [CrossRef] [PubMed]
- Pelowski, M.; Markey, P.S.; Lauring, J.O.; Leder, H. Visualizing the Impact of Art: An Update and Comparison of Current Psychological Models of Art Experience. Front. Hum. Neurosci. 2016, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A. Neuroaesthetics: A Coming of Age Story. J. Cogn. Neurosci. 2010, 23, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Vartanian, O. Neuroscience of aesthetics. Ann. N. Y. Acad. Sci. 2016, 1369, 172–194. [Google Scholar] [CrossRef] [PubMed]
- Leder, H. Next Steps in Neuroaesthetics: Which Processes and Processing Stages to Study? Psychol. Aesthet. Creat. Arts 2013, 7, 27–37. [Google Scholar] [CrossRef]
- Nadal, M.; Skov, M. Introduction to the Special Issue: Toward an Interdisciplinary Neuroaesthetics. Psychol. Aesthet. Creat. Arts 2013, 7, 1–12. [Google Scholar] [CrossRef]
- Zaidel, D.; Nadal, M.; Flexas, A.; Munar, E. An Evolutioary Approach to Art and Aesthetic Experience. Psychol. Aesthet. Creat. Arts 2013, 7, 100–109. [Google Scholar] [CrossRef]
- Zeki, S. Clive Bell’s “Significant Form” and the neurobiology of aesthetics. Front. Hum. Neurosci. 2013, 7, 730. [Google Scholar] [CrossRef] [PubMed]
- Alluri, V.; Toiviainen, P.; Burunat, I.; Kliuchko, M.; Vuust, P.; Brattico, E. Connectivity patterns during music listening: Evidence for action-based processing in musicians. Hum. Brain Mapp. 2017, 38, 2955–2970. [Google Scholar] [CrossRef] [PubMed]
- Garza-Villarreal, E.A.; Jiang, Z.; Vuust, P.; Alcauter, S.; Vase, L.; Pasaye, E.H.; Cavazos-Rodriguez, R.; Brattico, E.; Jensen, T.S.; Barrios, F.A. Music reduces pain and increases resting state fMRI BOLD signal amplitude in the left angular gyrus in fibromyalgia patients. Front. Psychol. 2015, 6, 1051. [Google Scholar] [CrossRef] [PubMed]
- Karmonik, C.; Brandt, A.; Anderson, J.; Brooks, F.; Lytle, J.; Silverman, E. Music Listening Modulates Functional Connectivity and Information Flow in the Human Brain. Brain Connect. 2016, 6, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Skouras, S. Functional centrality of amygdala, striatum and hypothalamus in a “small-world” network underlying joy: An fMRI study with music. Hum. Brain Mapp. 2014, 35, 3485–3498. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Skouras, S.; Lohmann, G. The auditory cortex hosts network nodes influential for emotion processing: An fMRI study on music-evoked fear and joy. PLoS ONE 2018, 13, e0190057. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Abu-Jamous, B.; Brattico, E.; Nandi, A.K. Towards Tunable Consensus Clustering for Studying Functional Brain Connectivity During Affective Processing. Int. J. Neural Syst. 2017, 27, 1650042. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Brattico, E.; Abu-Jamous, B.; Pereira, C.S.; Jacobsen, T.; Nandi, A.K. Effect of Explicit Evaluation on Neural Connectivity Related to Listening to Unfamiliar Music. Front. Hum. Neurosci. 2017, 11, 611. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Molina, N.; Mas-Herrero, E.; Rodriguez-Fornells, A.; Zatorre, R.J.; Marco-Pallares, J. Neural correlates of specific musical anhedonia. Proc. Natl. Acad. Sci. USA 2016, 113, E7337–E7345. [Google Scholar] [CrossRef] [PubMed]
- Sachs, M.E.; Ellis, R.E.; Schlaug, G.; Loui, P. Brain connectivity reflects human aesthetic responses. Soc. Cogn. Affect. Neurosci. 2016, 11, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Salimpoor, V.N.; van den Bosch, I.; Kovacevic, N.; McIntosh, A.R.; Dagher, A.; Zatorre, R.J. Interactions between the nucleus accumbens and auditory cortices predict music reward value. Science 2013, 340, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Taruffi, L.; Pehrs, C.; Skouras, S.; Koelsch, S. Effects of Sad and Happy Music on Mind-Wandering and the Default Mode Network. Sci. Rep. 2017, 7, 14396. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, R.W.; Hodges, D.A.; Laurienti, P.J.; Steen, M.; Burdette, J.H. Network Science and the Effects of Music Preference on Functional Brain Connectivity: From Beethoven to Eminem. Sci. Rep. 2014, 4, 6130. [Google Scholar] [CrossRef] [PubMed]
- Zeki, S. Inner Vision: An Exploration of Art and the Brain; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Zatorre, R.; Salimpoor, V. From perception to pleasure: Music and its neural substrates. Proc. Natl. Acad. Sci. USA 2013, 18 (Suppl. S2), 10430–10437. [Google Scholar] [CrossRef] [PubMed]
- Berlyne, D.E. Aesthetics and Psychobiology; Appleton-Century-Crofts: New York, NY, USA, 1971. [Google Scholar]
- Brattico, P.; Brattico, E.; Vuust, P. Global Sensory Qualities and Aesthetic Experience in Music. Front. Neurosci. 2017, 11, 159. [Google Scholar] [CrossRef] [PubMed]
- Bell, C. Art; Chatto and Windus: London, UK, 1914. [Google Scholar]
- Gebauer, L.K.; Kringelbach, M.L.; Vuust, P. Musical Pleasure Cycles: The role of anticipation and dopamine. Psychomusicology 2012, 22, 16. [Google Scholar] [CrossRef]
- Vuust, P.; Dietz, M.J.; Witek, M.; Kringelbach, M.L. Now you hear it: A predictive coding model for understanding rhythmic incongruity. Ann. N. Y. Acad. Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Brattico, E.; Vuust, P. The urge to judge: Why the judgmental attitude has anything to do with the aesthetic enjoyment of negative emotions. Behav. Brain Sci. 2017, 40, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Bundgaard, H. Feeling, meaning, and intentionality—A critique of the neuroaesthetics of beauty. Phenomenol. Cogn. Sci. 2015, 14, 781–801. [Google Scholar] [CrossRef]
- Kringelbach, M.L.; Berridge, K.C. The Affective Core of Emotion: Linking Pleasure, Subjective Well-Being, and Optimal Metastability in the Brain. Emot. Rev. 2017, 9, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Brattico, E.; Brattico, P.; Jacobsen, T. The origins of the aesthetic enjoyment of music—A review of the literature. Music. Sci. 2009, 13, 15–39. [Google Scholar] [CrossRef]
- Koelsch, S. Investigating emotion with music: An fMRI study. Hum. Brain Mapp. 2006, 27, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Reybrouck, M.; Eerola, T. Music and its inductive power: A psychobiological and evolutionary approach to musical emotions. Front. Psychol. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Redies, C. Combining universal beauty and cultural context in a unifying model of visual aesthetic experience. Front. Hum. Neurosci. 2015, 9, 218. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, D.J.; North, A.C. Experimental aesthetics and liking for music. In Handbook of Music and Emotion; Juslin, P.N., Sloboda, J.A., Eds.; Oxford University Press: Oxford, UK; New York, NY, USA, 2010; pp. 513–546. [Google Scholar]
- Vessel, E.; Starr, G.; Rubin, N. The brain on art: Intense aesthetic experience activates the default network. Front. Hum. Neurosci. 2012, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Vessel, E.A.; Starr, G.G.; Rubin, N. Art reaches within: Aesthetic experience, the self and the default network. Front. Neurosci. 2013, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O. Networks of the Brain; MIT Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Cabral, J.; Kringelbach, M.L.; Deco, G. Exploring the network dynamics underlying brain activity during rest. Prog. Neurobiol. 2014, 114, 102–131. [Google Scholar] [CrossRef] [PubMed]
- Biswal, B.; Yetkin, F.Z.; Haughton, V.M.; Hyde, J.S. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn. Reson. Med. 1995, 34, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Fauvel, B.; Groussard, M.; Chételat, G.; Fouquet, M.; Landeau, B.; Eustache, F.; Desgranges, B.; Platel, H. Morphological brain plasticity induced by musical expertise is accompanied by modulation of functional connectivity at rest. NeuroImage 2014, 90, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.; Schaefer, R.; Bastin, M.; Roberts, N.; Overy, K. Can Musical Training Influence Brain Connectivity? Evidence from Diffusion Tensor MRI. Brain Sci. 2014, 4, 405–427. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, M.; Hulshoff Pol, H. Exploring the brain network: A review on resting-state fMRI functional connectivity. Eur. Neuropsychopharmacol. 2010, 20, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Immordino-Yang, M.H.; Christodoluou, J.; Singh, V. Rest Is Not Idleness: Implications of the Brain’s Default Mode for Human Deelopment and Education. Perspect. Psychol. Sci. 2012, 7, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.; Andrews-Hanna, J.; Schacter, D. The Brain’s Default Network. Anatomy, Function, and Relevance to Disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M. The brain’s default network. Ann. Rev. Neurosci. 2015, 8, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.; Snyder, A. A default mode of brain function: A brief history of an evolving idea. NeuroImage 2007, 37, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Kringelbach, M.; Berridge, K. Towards a functional neuroanatomy of pleasure and happiness. Trends Cogn. Sci. 2009, 13, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.; Wymbs, N.; Porter, M.; Mucha, P.; Carlson, J.; Grafton, S. Dynamic reconfiguration of human brain networks during learning. Proc. Natl. Acad. Sci. USA 2011, 3, 7641–7646. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Liem, F.; Hänggi, J.; Elmer, S.; Jäncke, L. The “Silent” Imprint of Musical Training. Hum. Brain Mapp. 2016, 37, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Cabral, J.; Vidaurre, D.; Marques, P.; Magalhães, R.; Silva Moreira, P.; Miguel Soares, J.; Deco, G.; Sousa, N.; Kringelbach, M.L. Cognitive performance in healthy older adults relates to spontaneous switching between states of functional connectivity during rest. Sci. Rep. 2017, 7, 5135. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.; Greicius, M. Clinical applications of resting state functional connectivity. Front. Syst. Neurosci. 2010, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Gusnard, D.A.; Akbudak, E.; Shulman, G.L.; Raichle, M.E. Medial prefrontal cortex and self-referential mental activity: Reation to a default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 4259–4264. [Google Scholar] [CrossRef] [PubMed]
- Saarikallio, S.; Erkkila, J. The role of music in adolescents’ mood regulation. Psychol. Music 2007, 35, 88–109. [Google Scholar] [CrossRef]
- Tanner, J.; Asbridge, M.; Wortley, S. Our favourite melodies: Musical consumption and teenage lifestyles. Br. J. Sociol. 2008, 59, 117–144. [Google Scholar] [CrossRef] [PubMed]
- Avram, M.; Gutyrchik, E.; Bao, Y.; Pöppel, E.; Reiser, M.; Blautzik, J. Neurofunctional correlates of esthetic and moral judgments. Neurosci. Lett. 2013, 534, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, N.; Peretz, I.; Johnsen, E.; Adolphs, R. Amygdala damage impairs emotion recognition from music. Neuropsychologia 2007, 45, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Zald, D. The human amygdala and the emotional evaluation of sensory stimuli. Brain Res. Rev. 2003, 41, 88–123. [Google Scholar] [CrossRef]
- Waterman, A.S. Two conceptions of happiness: Contrasts of personal expressiveness (eudaimonia) and hedonic enjoyment. J. Personal. Soc. Psychol. 1993, 64, 678–691. [Google Scholar] [CrossRef]
- Berridge, K.C.; Robinson, T.E. Parsing reward. Trends Neurosci. 2003, 26, 507–513. [Google Scholar] [CrossRef]
- Eklund, A.; Nichols, T.E.; Knutsson, H. Cluster failure: Why fMRI inferences for spatial extent have inflated false-positive rates. Proc. Natl. Acad. Sci. USA 2016, 113, 7900–7905. [Google Scholar] [CrossRef] [PubMed]
- Abu-Jamous, B.; Fa, R.; Roberts, D.; Nandi, K. Paradigm of tunable clustering using binarization of consensus partition matrices (Bi-CoPaM) for gene discovery. PLoS ONE 2013, 8, e56432. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, M.P.; Mandl, R.C.; Hulshoff Pol, H.E. Normalized cut group clustering of resting-state fMRI data. PLoS ONE 2008, 3, e2001. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Suzukamo, Y.; Sato, M.; Izumi, S. Effects of music therapy on behavioral and psychological symptoms of dementia: A systematic review and meta-analysis. Aging Res. Rev. 2013, 12, 628–641. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Yang, M.; Duan, M.; Chen, X.; Lai, Y.; Xia, Y.; Shao, J.; Biswal, B.B.; Luo, C.; Yao, D. Music Intervention Leads to Increased Insular Connectivity and Improved Clinical Symptoms in Schizophrenia. Front. Neurosci. 2017, 11, 744. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Study | Method | Participants | Major Findings |
|---|---|---|---|
| Alluri et al., 2017 [30] | Task-free music listening; fMRI; whole-brain graph-theory analyses | Musicians (n = 18) and non-musicians (n = 18) | Musicians primary hubs: cerebral and cerebellar sensorimotor regions, non-musicians: DMN-related regions. |
| Garza-Villareal et al., 2015 [31] | Resting state fMRI; BOLD signal | Non-musicians fibromyalgia patients (n = 22) | Increased amplitude of BOLD signal after listening; higher connectivity with right dorsolateral prefrontal cortex and left caudate; decreased connectivity with right anterior cingulate cortex, right supplementary motor area, precuneus and right precentral gyrus. |
| Karmonik et al., 2016 [32] | Task-free music listening; fMRI; graph network analysis | Non-musicians (n = 12) | Variation in functional connectivity after listening; most intense connections between brain areas were found for processing self-selected emotional music or culturally unfamiliar music. |
| Koelsch & Skouras, 2014 [33] | Mixed-block design; fMRI; eigenvector centrality mapping; functional connectivity analysis | Non-musicians, n = 20 | Superficial amygdala, laterobasal amygdala, striatum, and hypothalamus function as computational hubs during joy evoked by music. |
| Koelsch, Skouras & Lohmann, 2018 [34] | Mixed-block design; fMRI | Non-musicians, n = 24 | Anterior and posterior regions of auditory association cortex show functional connectivity with limbic/paralimbic, somatosensory, visual, motor-related, and attentional structures; primary auditory fields show strong functional connectivity with intra-auditory regions. |
| Liu et al., 2017a [35] | Mixed-block design; fMRI; consensus clustering with Bi-CoPam algorithm | Musicians (n = 13) and non-musicians (n = 16) | Brain structures related to visual, reward, and auditory processing show robust spatial patterns of coherent neuroactivity during affective processing. |
| Liu et al., 2017b [36] | Mixed-block design; fMRI; consensus clustering with Bi-CoPam algorithm | Non-musicians (n = 25) | Impact of explicit evaluative judgment on neural auditory-limbic connectivity during affective processing of music. |
| Martinez-Molina et al., 2016 [37] | Barcelona Musical Reward Questionnaire (BMRQ); skin conductance response (SCR); fMRI; psychophysiological interaction (PPI) | Non-musicians (n = 45) | Music anhedonic participants show selective reduction of activity for music in nucleus accumbens and decreased functional connectivity between right auditory cortex and ventral striatum. |
| Sachs et al., 2016 [38] | Survey data; behavioral, psychophysiological Measures; diffusion tensor imaging (DTI) | Experiencing chills to music (n = 10), not experiencing chills to music (n = 10) | White matter connectivity between sensory processing areas in superior temporal gyrus and emotional and social processing areas in insula and medial prefrontal cortex explains individual differences in reward sensitivity to music. |
| Salimpoor et al., 2013 [39] | Event-related design with decision-making after listening to music excerpts; fMRI; partial least-squares analysis | Not specified (n = 19) | During listening to purchased (vs. non-purchased) music clips, the nucleus accumbens increased its connectivity with superior temporal gyrus, orbitofrontal cortex, amygdala, ventromedial prefrontal cortex, anterior cingulate, and inferior frontal gyrus. |
| Taruffi et al., 2017 [40] | Mixed-block design; fMRI; ECM analysis | Not specified. Three experiments: n = 224; n = 140, n = 24 | Sad music, compared with happy music, is associated with stronger mind-wandering and greater centrality of the nodes of the Default Mode Network. |
| Wilkins et al., 2014 [41] | Free listening; fMRI | Not specified (n = 21) | Circuit important for internally-focused thoughts, known as the default mode network is most connected when listening to preferred music. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reybrouck, M.; Vuust, P.; Brattico, E. Brain Connectivity Networks and the Aesthetic Experience of Music. Brain Sci. 2018, 8, 107. https://doi.org/10.3390/brainsci8060107
Reybrouck M, Vuust P, Brattico E. Brain Connectivity Networks and the Aesthetic Experience of Music. Brain Sciences. 2018; 8(6):107. https://doi.org/10.3390/brainsci8060107
Chicago/Turabian StyleReybrouck, Mark, Peter Vuust, and Elvira Brattico. 2018. "Brain Connectivity Networks and the Aesthetic Experience of Music" Brain Sciences 8, no. 6: 107. https://doi.org/10.3390/brainsci8060107
APA StyleReybrouck, M., Vuust, P., & Brattico, E. (2018). Brain Connectivity Networks and the Aesthetic Experience of Music. Brain Sciences, 8(6), 107. https://doi.org/10.3390/brainsci8060107