
The Interplay of Hippocampus and Ventromedial Prefrontal Cortex in Memory-Based Decision Making


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
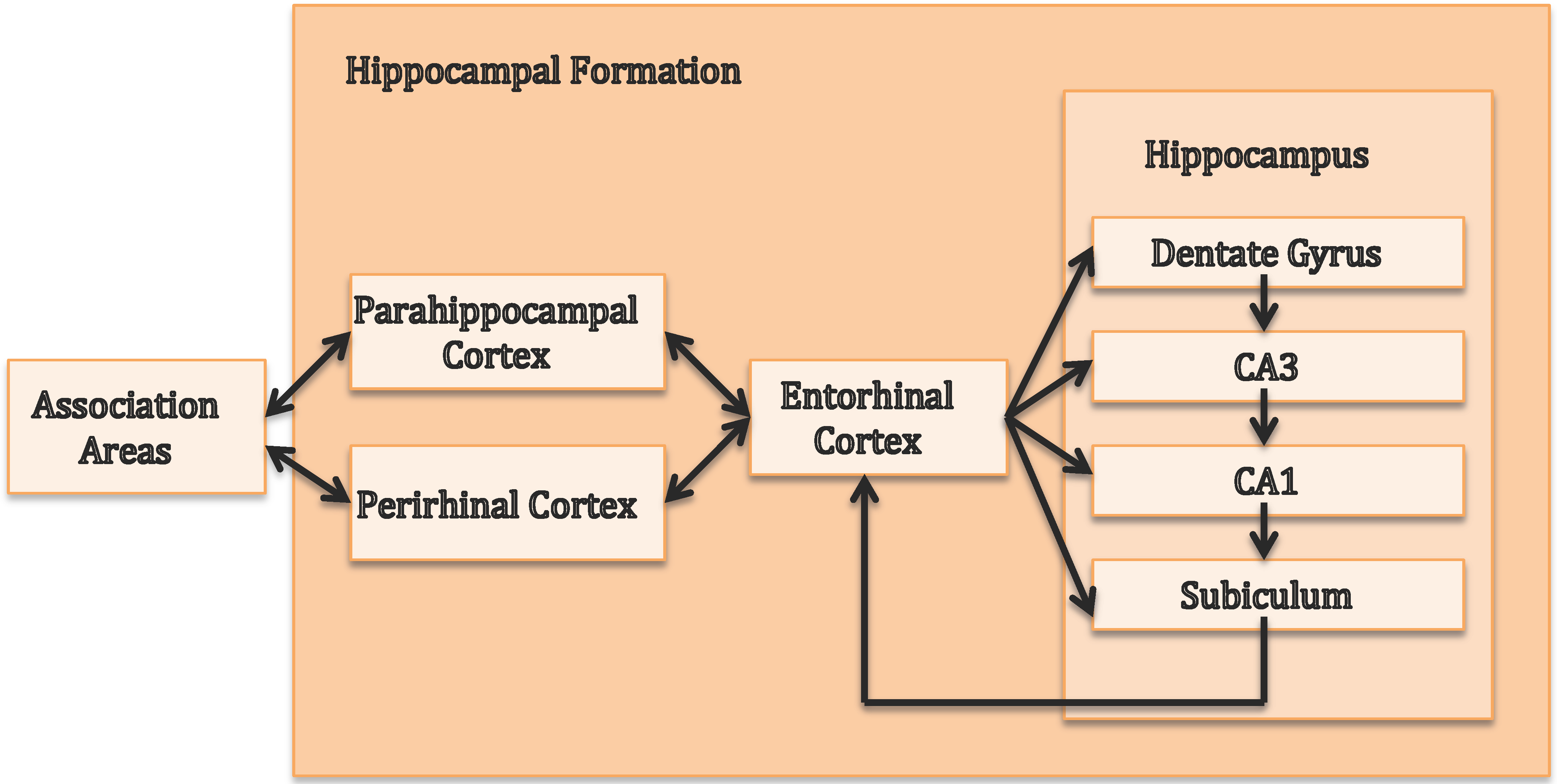
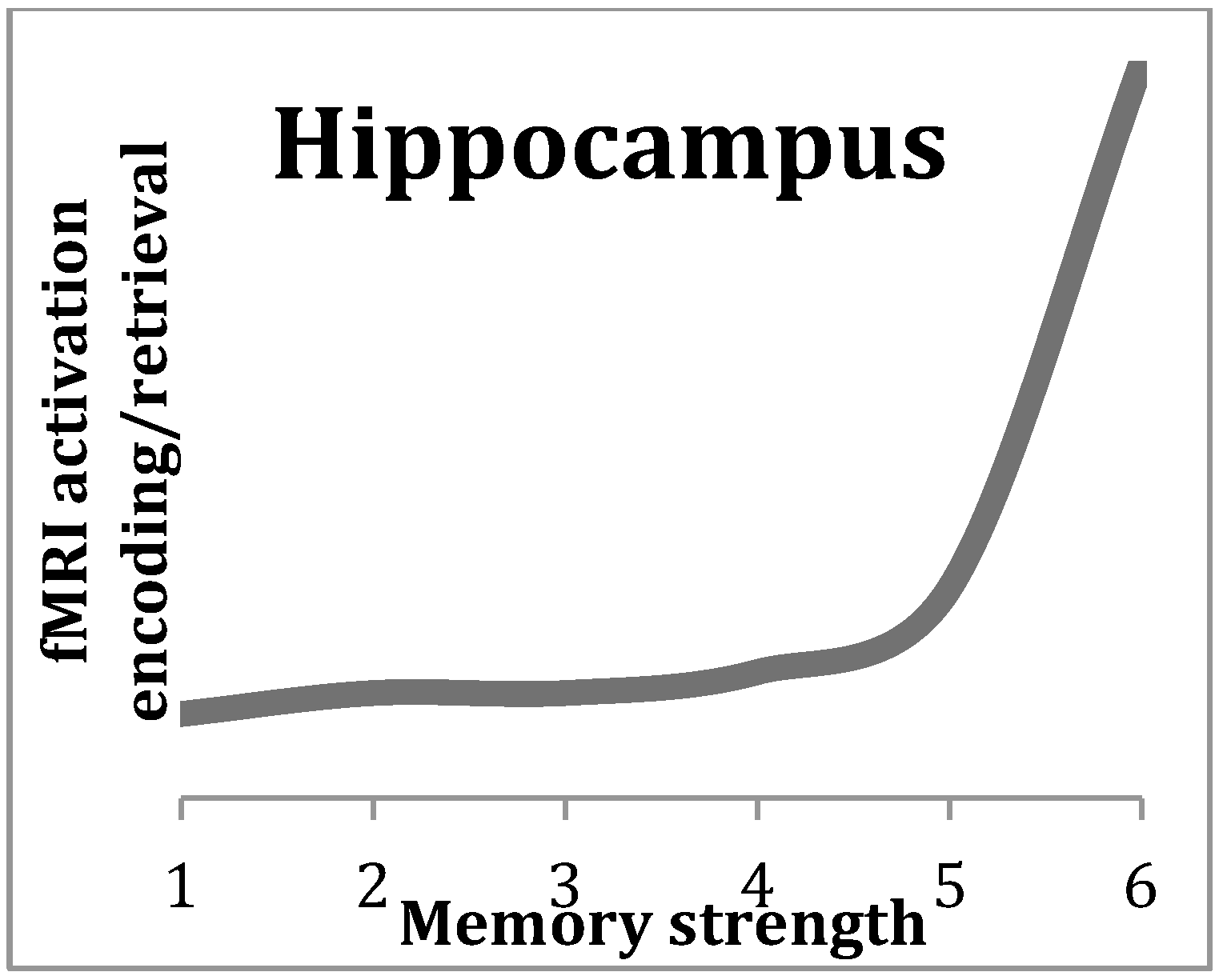
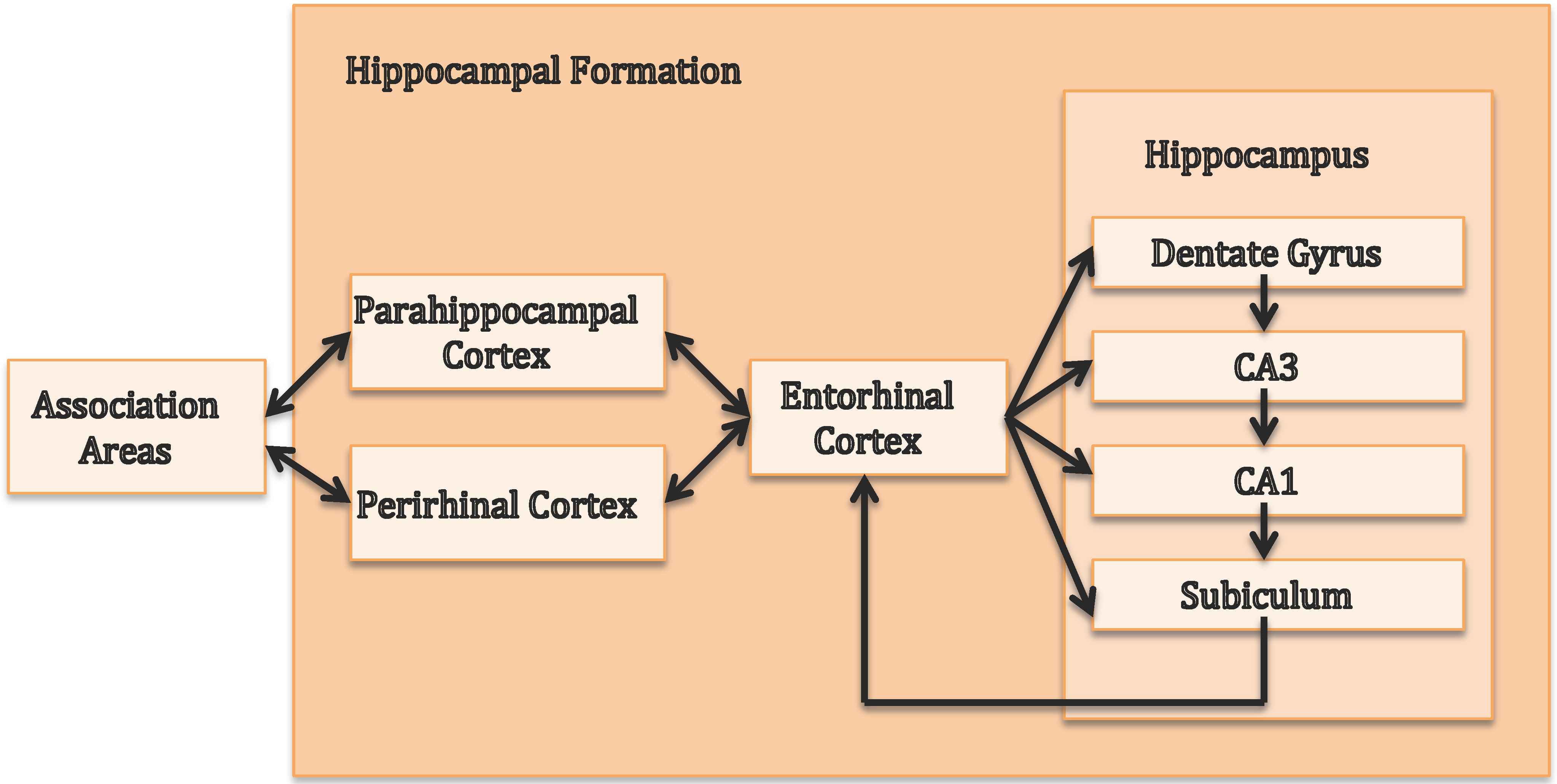
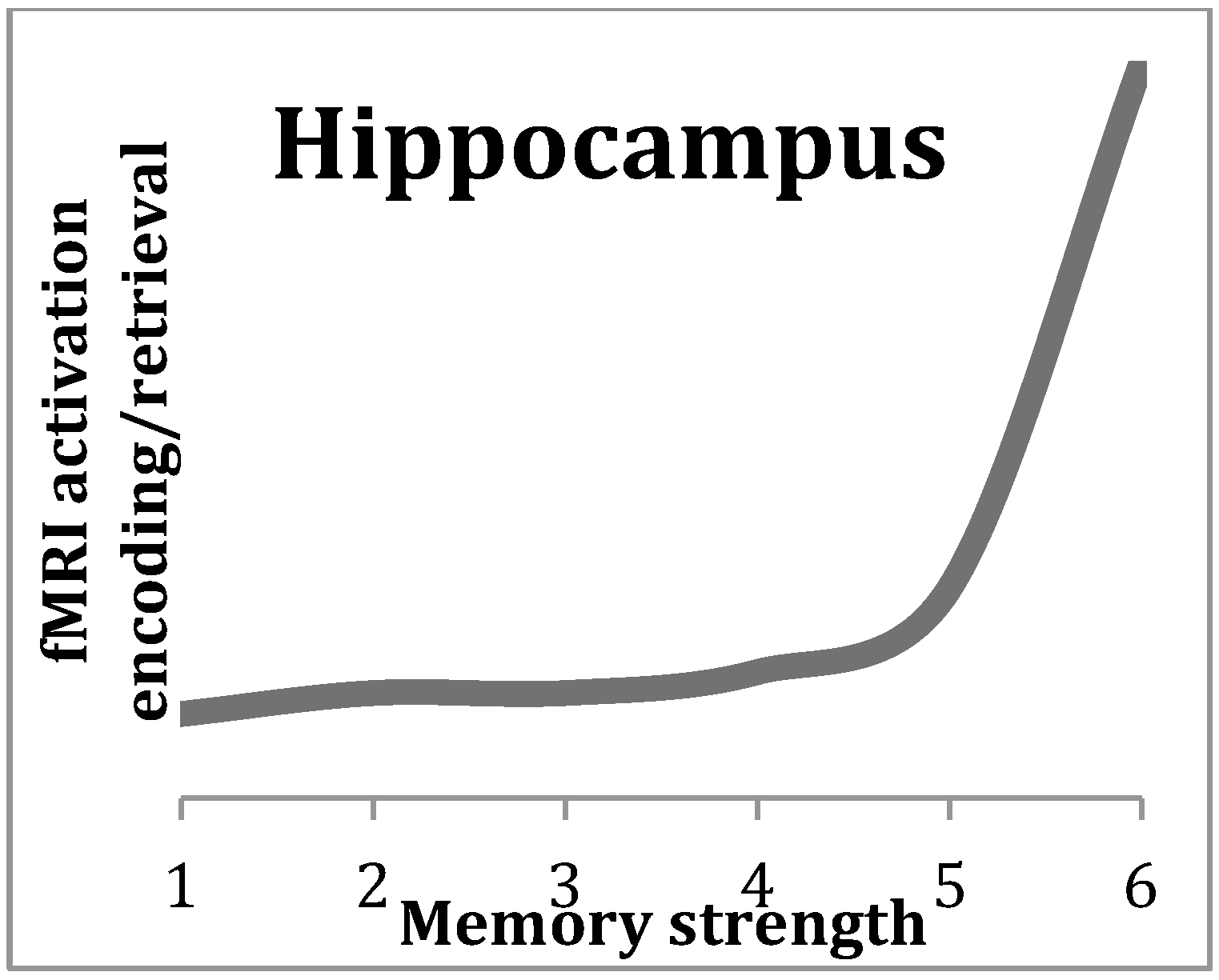
2. The Central Role of the Hippocampus in Episodic Memory
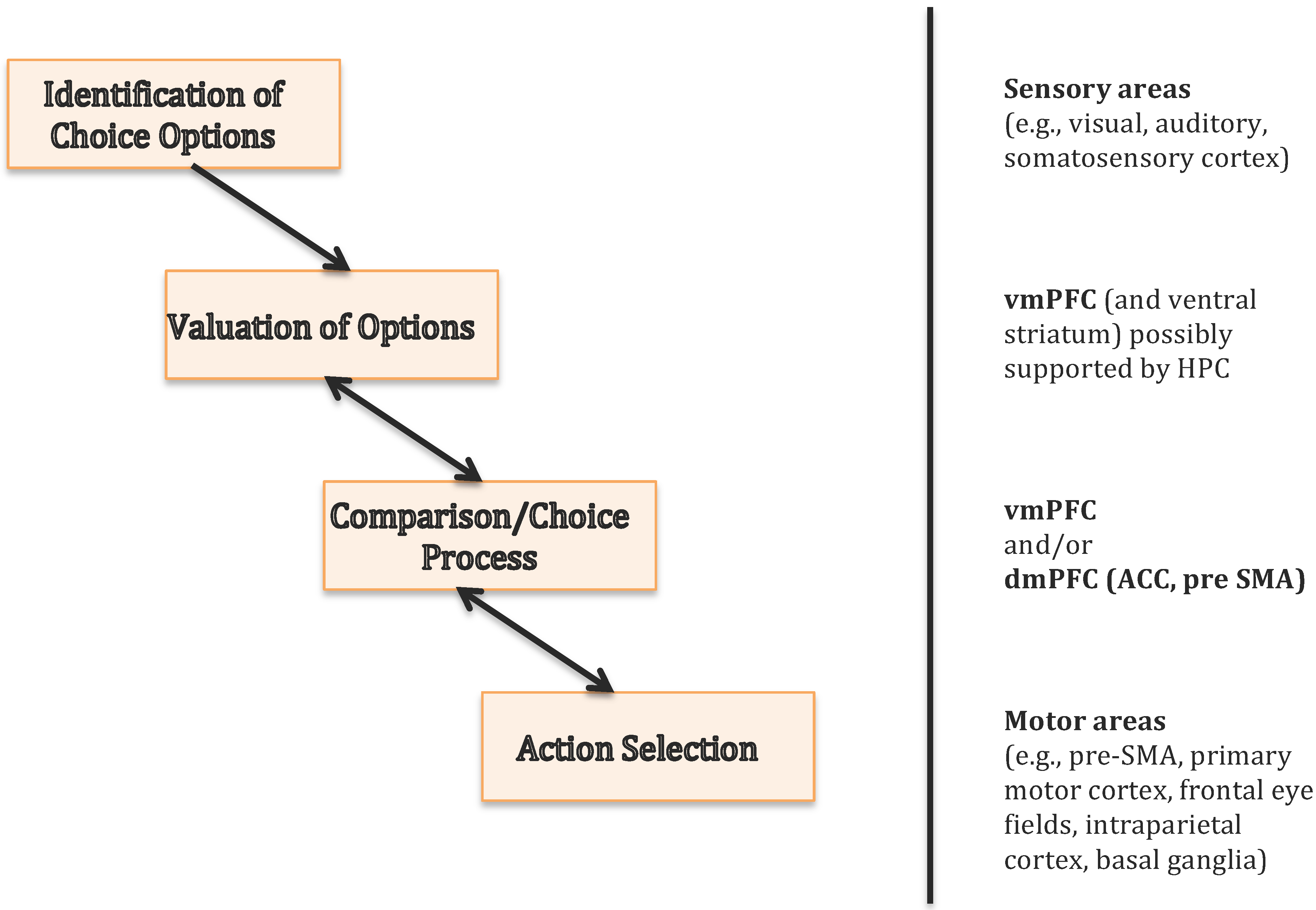
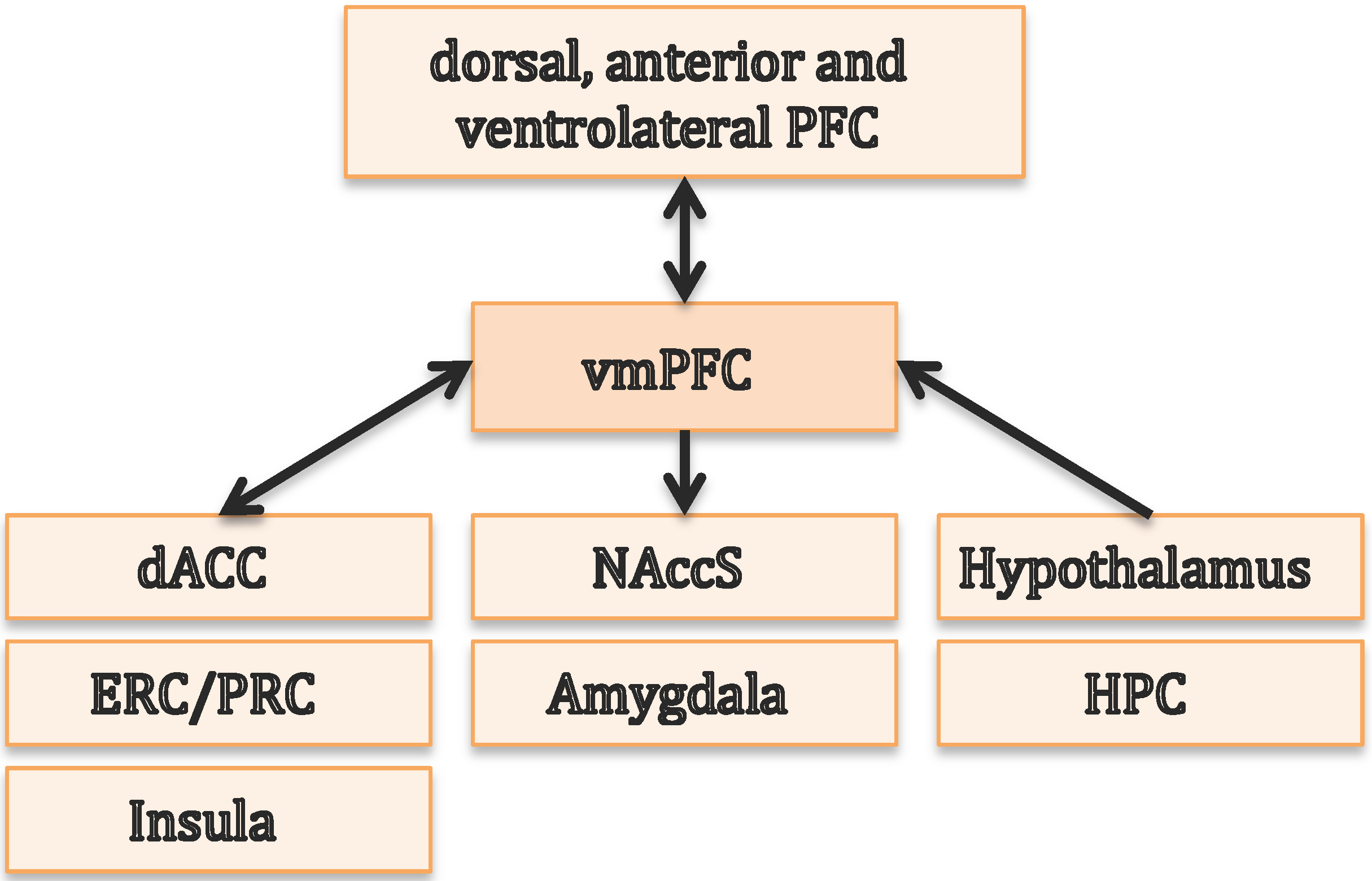
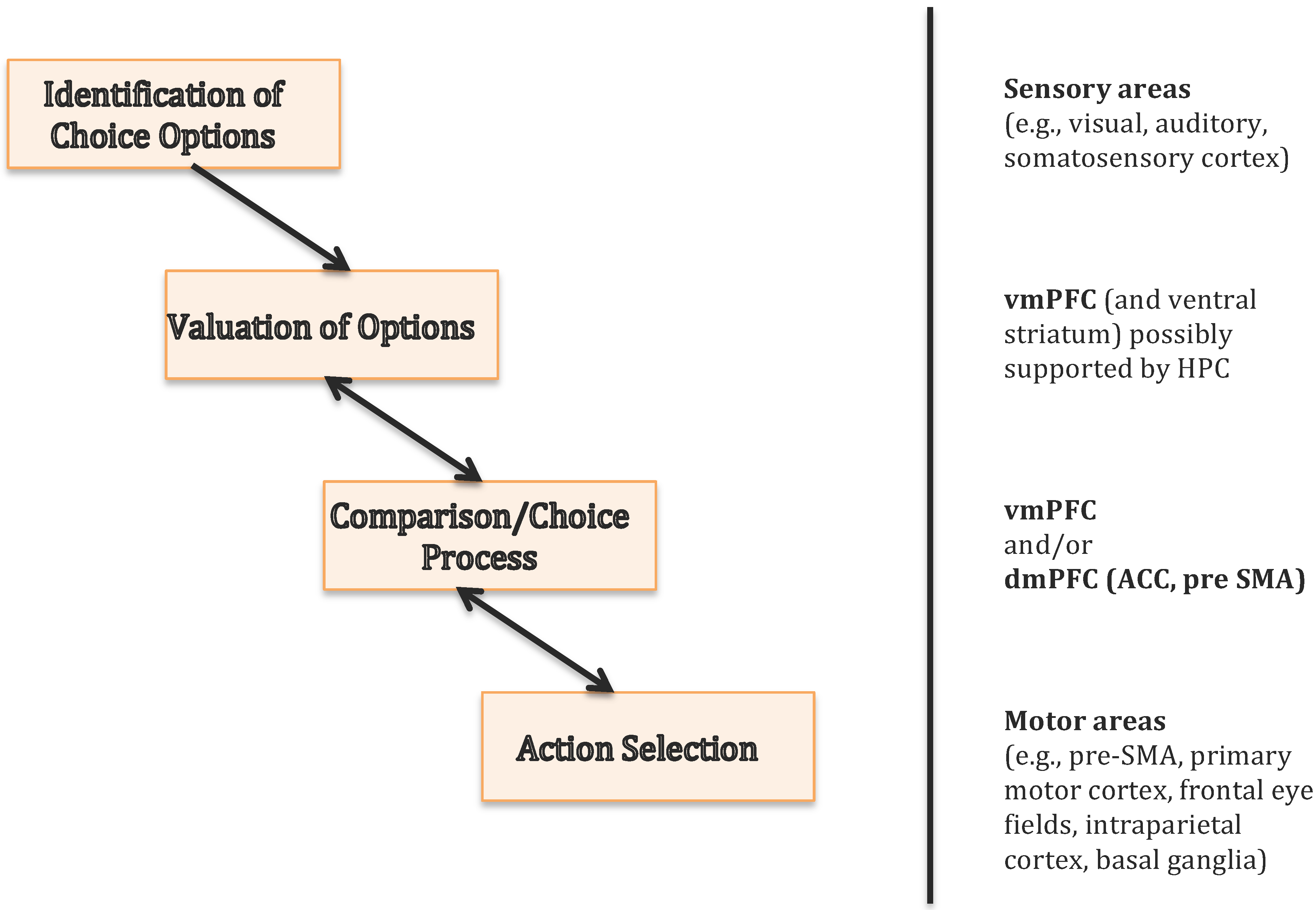
3. The Ventromedial Prefrontal Cortex Is Essential for Value-Based Decisions
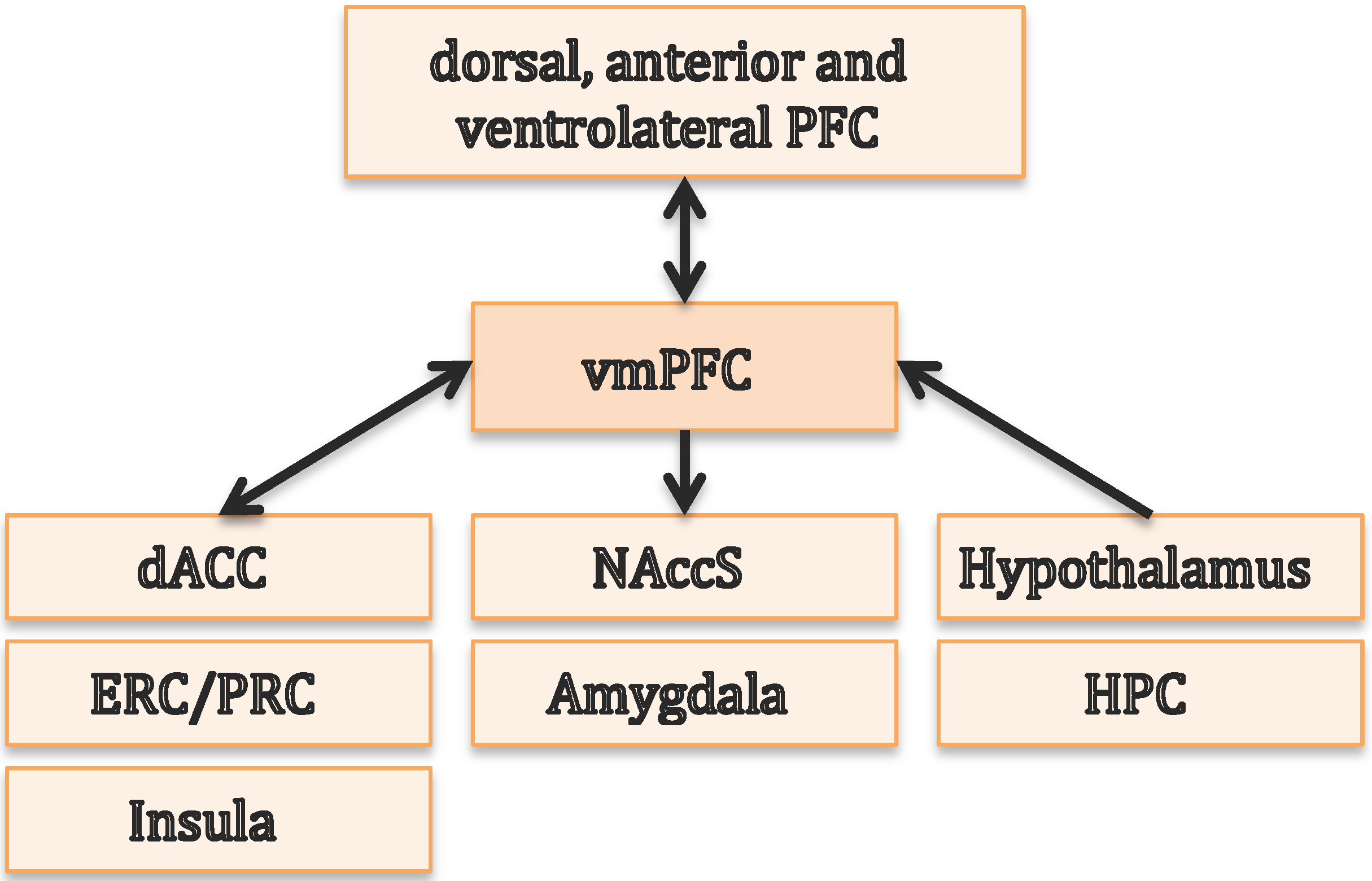
4. Hippocampal-Prefrontal Interactions in Episodic Memory and Decision Making
5. Open Questions and Tentative Predictions for Future Studies on the Interplay of Value-Based Decisions Making and Episodic Memory
5.1. Question 1: What Is the Neural Code That Underlies Memory-Based Decisions?
5.2. Question 2: How Can We Bring Schema-Related and Choice-Related HPC–vmPFC Interactions Together?
5.3. Question 3: What Is the Direction of HPC–vmPFC Connectivity?
5.4. Question 4: Does the HPC Represent Subjective Value?
6. Conclusions
Author Contributions
Conflicts of Interest
References
- US National Library of Medicine National Institutes of Health. Available online: https://www.ncbi.nlm.nih.gov/pubmed (accessed on 13 September 2016).
- Shadlen, M.N.; Shohamy, D. Decision making and sequential sampling from memory. Neuron 2016, 90, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Simons, J.S.; Spiers, H.J. Prefrontal and medial temporal lobe interactions in long-term memory. Nat. Rev. Neurosci. 2003, 4, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Milner, B.; Squire, L.R.; Kandel, E.R. Cognitive neuroscience review and the study of memory. Neuron 1998, 20, 445–468. [Google Scholar] [CrossRef]
- Scoville, W.B.; Milner, B. Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry 1957, 20, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Zola-Morgan, S.; Squire, L.R.; Amaral, D.G. Human amnesia and the medial temporal region: Enduring memory impairment following a bilateral lesion limited to field CA1 of the hippocampus. J. Neurosci. 1986, 6, 2950–2967. [Google Scholar] [CrossRef] [PubMed]
- Vargha-Khadem, F.; Gadian, D.G.; Watkins, K.E. Differential effects of early hippocampal pathology on episodic and semantic memory. Science 1997, 277, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Eichenbaum, H.; Yonelinas, A.P.; Ranganath, C. The medial temporal lobe and recognition memory. Annu. Rev. Neurosci. 2007, 30, 123–152. [Google Scholar] [CrossRef] [PubMed]
- Bernard, F.A.; Bullmore, E.T.; Graham, K.S.; Thompson, S.A.; Hodges, J.R.; Fletcher, P.C. The hippocampal region is involved in successful recognition of both remote and recent famous faces. Neuroimage 2004, 22, 1704–1714. [Google Scholar] [CrossRef] [PubMed]
- Gold, J.J.; Smith, C.N.; Bayley, P.J.; Shrager, Y.; Brewer, J.B.; Stark, C.E.L.; Hopkins, R.O.; Squire, L.R. Item memory, source memory, and the medial temporal lobe: Concordant findings from fMRI and memory-impaired patients. Proc. Natl. Acad. Sci. USA 2006, 103, 9351–9356. [Google Scholar] [CrossRef] [PubMed]
- Squire, L.R.; Wixted, J.T.; Clark, R.E. Recognition memory and the medial temporal lobe: A new perspective. Nat. Rev. Neurosci. 2007, 8, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Bor, D.; Duncan, J.; Wiseman, R.J.; Owen, A.M. Encoding strategies dissociate prefrontal activity from working memory demand. Neuron 2003, 37, 361–367. [Google Scholar] [CrossRef]
- Unsworth, N.; Brewer, G.A.; Spillers, G.J. Working memory capacity and retrieval from long-term memory: The role of controlled search. Mem. Cognit. 2013, 41, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Nadel, L.; Moscovitch, M. Memory consolidation, retrograde amnesia and the hippocampal complex. Curr. Opin. Neurobiol. 1997, 7, 217–227. [Google Scholar] [CrossRef]
- Moscovitch, M.; Nadel, L.; Winocur, G.; Gilboa, A.; Rosenbaum, R.S. The cognitive neuroscience of remote episodic, semantic and spatial memory. Curr. Opin. Neurobiol. 2006, 16, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Winocur, G.; Moscovitch, M. Memory transformation and systems consolidation. J. Int. Neuropsychol. Soc. 2011, 17, 766–780. [Google Scholar] [CrossRef] [PubMed]
- O’keefe, J.; Nadel, L. The Hippocampus as a Cognitive Map; Oxford University Press: Oxford, MS, USA, 1978. [Google Scholar]
- Dudai, Y. The neurobiology of consolidations, or, how stable is the engram? Annu. Rev. Psychol. 2004, 55, 51–86. [Google Scholar] [CrossRef] [PubMed]
- Binder, S.; Dere, E.; Zlomuzica, A. A critical appraisal of the what-where-when episodic-like memory test in rodents: Achievements, caveats and future directions. Prog. Neurobiol. 2015, 130, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Dere, E.; Huston, J.P.; De Souza Silva, M.A. Integrated memory for objects, places, and temporal order: Evidence for episodic-like memory in mice. Neurobiol. Learn. Mem. 2005, 84, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Dere, E.; Huston, J.P.; De Souza Silva, M.A. Episodic-like memory in mice: Simultaneous assessment of object, place and temporal order memory. Brain Res. Protoc. 2005, 16, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, R.E.; Pfeiffer, B.E.; Foster, D.J. Reverse replay of hippocampal place cells is uniquely modulated by changing reward. Neuron 2016, 91, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Hok, V.; Chah, E.; Save, E.; Poucet, B. Prefrontal cortex focally modulates hippocampal place cell firing patterns. J. Neurosci. 2013, 33, 3443–3451. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J. Place units in the hippocampus of the freely moving rat. Exp. Neurol. 1976, 51, 78–109. [Google Scholar] [CrossRef]
- Squire, L.R.; Genzel, L.; Wisted, J.T.; Morris, R.G.M. Memory consolidation. Cold Spring Harb. Perspect. Biol. 2015, 7, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.; McNaughton, B. Reactivation of hippocampal ensemble memories during sleep. Science 1994, 265, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Peigneux, P.; Laureys, S.; Fuchs, S.; Collette, F.; Perrin, F.; Reggers, J.; Phillips, C.; Degueldre, C.; Del Fiore, G.; Aerts, J.; et al. Are spatial memories strengthened in the human hippocampus during slow wave sleep? Neuron 2004, 44, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Davidson, T.J.; Kloosterman, F.; Wilson, M.A. Hippocampal replay of extended experience. Neuron 2009, 63, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.P.; Frank, L.M. Awake replay of remote experiences in the hippocampus. Nat. Neurosci. 2009, 12, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Diba, K.; Buzsáki, G. Forward and reverse hippocampal place-cell sequences during ripples. Nat. Neurosci. 2007, 10, 1241–1242. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G.; Moser, E.I. Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat. Neurosci. 2013, 16, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Haber, S.N.; Behrens, T.E.J. The neural network underlying incentive-based learning: Implications for interpreting circuit disruptions in psychiatric disorders. Neuron 2014, 83, 1019–1039. [Google Scholar] [CrossRef] [PubMed]
- Haber, S.N.; Knutson, B. The reward circuit: Linking primate anatomy and human imaging. Neuropsychopharmacology 2009, 35, 4–26. [Google Scholar] [CrossRef] [PubMed]
- Rushworth, M.F.S.; Noonan, M.P.; Boorman, E.D.; Walton, M.E.; Behrens, T.E. Frontal cortex and reward-guided learning and decision-making. Neuron 2011, 70, 1054–1069. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.R.; Reasor, J.E.; Truckenbrod, L.M.; Lubke, K.N.; Johnson, S.A.; Bizon, J.L.; Maurer, A.P.; Burke, S.N. Medial prefrontal-perirhinal cortical communication is necessary for flexible response selection. Neurobiol. Learn. Mem. 2016, 137, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Kahn, I.; Andrews-Hanna, J.R.; Vincent, J.L.; Snyder, A.Z.; Buckner, R.L. Distinct cortical anatomy linked to subregions of the medial temporal lobe revealed by intrinsic functional connectivity. J. Neurophysiol. 2008, 100, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Kinnavane, L.; Amin, E.; Olarte-Sánchez, C.M.; Aggleton, J.P. Detecting and discriminating novel objects: The impact of perirhinal cortex disconnection on hippocampal activity patterns. Hippocampus 2016, 26, 1393–1413. [Google Scholar] [CrossRef] [PubMed]
- Takehara-Nishiuchi, K. Entorhinal cortex and consolidated memory. Neurosci. Res. 2014, 84, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Damasio, H.; Grabowski, T.; Frank, R.; Galaburda, A.; Damasio, A. The return of Phineas Gage: Clues about the brain from the skull of a famous patient. Science 1994, 264, 1102–1105. [Google Scholar] [CrossRef] [PubMed]
- Rangel, A.; Camerer, C.; Montague, P.R. A framework for studying the neurobiology of value-based decision making. Nat. Rev. Neurosci. 2008, 9, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Bartra, O.; McGuire, J.T.; Kable, J.W. The valuation system: A coordinate-based meta-analysis of BOLD fMRI experiments examining neural correlates of subjective value. Neuroimage 2013, 76, 412–427. [Google Scholar] [CrossRef] [PubMed]
- Bechara, A.; Damasio, A.R.; Damasio, H.; Anderson, S.W. Insensitivity to future consequences following damage to human prefrontal cortex. Cognition 1994, 50, 7–15. [Google Scholar] [CrossRef]
- Fellows, L.K.; Farah, M.J. The role of ventromedial prefrontal cortex in decision making: Judgment under uncertainty or judgment per se? Cereb. Cortex 2007, 17, 2669–2674. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.; D’Esposito, M. Effects of medial orbitofrontal cortex lesions on self-control in intertemporal choice. Curr. Biol. 2016, 26, 2625–2628. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, A.R.; Fellows, L.K. Ventromedial frontal cortex is critical for guiding attention to reward-predictive visual features in humans. J. Neurosci. 2015, 35, 12813–12823. [Google Scholar] [CrossRef] [PubMed]
- Chib, V.S.; Rangel, A.; Shimojo, S.; O’Doherty, J.P. Evidence for a common representation of decision values for dissimilar goods in human ventromedial prefrontal cortex. J. Neurosci. 2009, 29, 12315–12320. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, M.; Jorge, S.; Michel, V.; Thirion, B.; Pessiglione, M. An automatic valuation system in the human brain: Evidence from functional neuroimaging. Neuron 2009, 64, 431–439. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, T.H.B.; Seymour, B.; Dolan, R.J. The role of human orbitofrontal cortex in value comparison for incommensurable objects. J. Neurosci. 2009, 29, 8388–8395. [Google Scholar] [CrossRef] [PubMed]
- Kable, J.W.; Glimcher, P.W. The neural correlates of subjective value during intertemporal choice. Nat. Neurosci. 2007, 10, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Clithero, J.A.; Rangel, A. Informatic parcellation of the network involved in the computation of subjective value. Soc. Cogn. Affect. Neurosci. 2014, 9, 1289–1302. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.J.; Glimcher, P.W. The root of all value: A neural common currency for choice. Curr. Opin. Neurobiol. 2012, 22, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Hunt, L.T.; Kolling, N.; Soltani, A.; Woolrich, M.W.; Rushworth, M.F.S.; Behrens, T.E.J. Mechanisms underlying cortical activity during value-guided choice. Nat. Neurosci. 2012, 15, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Strait, C.E.; Blanchard, T.C.; Hayden, B.Y. Reward value comparison via mutual inhibition in ventromedial prefrontal cortex. Neuron 2014, 82, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Boorman, E.D.; Behrens, T.E.J.; Woolrich, M.W.; Rushworth, M.F.S. How green is the grass on the other side? Frontopolar cortex and the evidence in favor of alternative courses of action. Neuron 2009, 62, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Hare, T.A.; Schultz, W.; Camerer, C.F.; O’Doherty, J.P.; Rangel, A. Transformation of stimulus value signals into motor commands during simple choice. Proc. Natl. Acad. Sci. USA 2011, 108, 18120–18125. [Google Scholar] [CrossRef] [PubMed]
- Gluth, S.; Rieskamp, J.; Büchel, C. Deciding when to decide: Time-variant sequential sampling models explain the emergence of value-based decisions in the human brain. J. Neurosci. 2012, 32, 10686–10698. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, K.; Rangel, A.; O’Doherty, J.P. Neural computations underlying action-based decision making in the human brain. Proc. Natl. Acad. Sci. USA 2009, 106, 17199–17204. [Google Scholar] [CrossRef] [PubMed]
- Klein-Flügge, M.C.; Bestmann, S. Time-dependent changes in human corticospinal excitability reveal value-based competition for action during decision processing. J. Neurosci. 2012, 32, 8373–8382. [Google Scholar] [CrossRef] [PubMed]
- Gluth, S.; Rieskamp, J.; Büchel, C. Classic EEG motor potentials track the emergence of value-based decisions. Neuroimage 2013, 79, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Cisek, P.; Kalaska, J.F. Neural correlates of reaching decisions in dorsal premotor cortex: Specification of multiple direction choices and final selection of action. Neuron 2005, 45, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Heath, T.B.; Chatterjee, S. Asymmetric decoy effects on lower-quality versus higher-quality brands: Meta-analytic and experimental evidence. J. Consum. Res. 1995, 22, 268–284. [Google Scholar] [CrossRef]
- Rieskamp, J.; Busemeyer, J.R.; Mellers, B.A. Extending the bounds of rationality: Evidence and theories of preferential choice. J. Econ. Lit. 2006, 44, 631–661. [Google Scholar] [CrossRef]
- Vlaev, I.; Chater, N.; Stewart, N.; Brown, G.D.A. Does the brain calculate value? Trends Cogn. Sci. 2011, 15, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Hedgcock, W.; Rao, A.R. Trade-off aversion as an explanation for the attraction effect: A functional magnetic resonance imaging study. J. Mark. Res. 2009, 46, 1–13. [Google Scholar] [CrossRef]
- Hu, J.; Yu, R. The neural correlates of the decoy effect in decisions. Front. Behav. Neurosci. 2014, 8, 271. [Google Scholar] [CrossRef] [PubMed]
- Hunt, L.T.; Dolan, R.J.; Behrens, T.E.J. Hierarchical competitions subserving multi-attribute choice. Nat. Neurosci. 2014, 17, 1613–1622. [Google Scholar] [CrossRef] [PubMed]
- Gluth, S.; Hotaling, J.M.; Rieskamp, J. The Attraction Effect Modulates Reward Prediction Errors and Intertemporal Choices. J. Neurosci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Zeithamova, D.; Dominick, A.L.; Preston, A.R. Hippocampal and ventral medial prefrontal activation during retrieval-mediated learning supports novel inference. Neuron 2012, 75, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Kumaran, D.; Summerfield, J.J.; Hassabis, D.; Maguire, E.A. Tracking the emergence of conceptual knowledge during human decision making. Neuron 2009, 63, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.Y.; Frank, L.M. Hippocampal–cortical interaction in decision making. Neurobiol. Learn. Mem. 2015, 117, 34–41. [Google Scholar] [CrossRef] [PubMed]
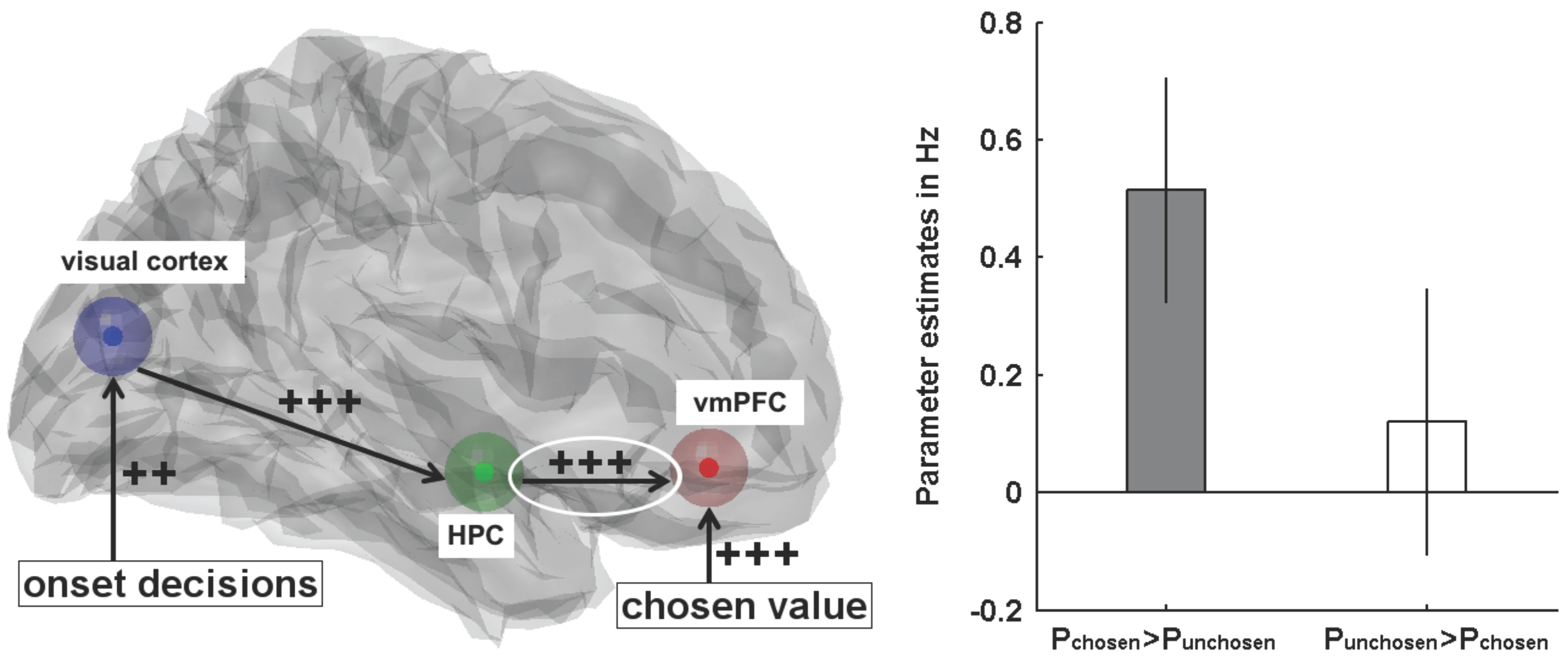
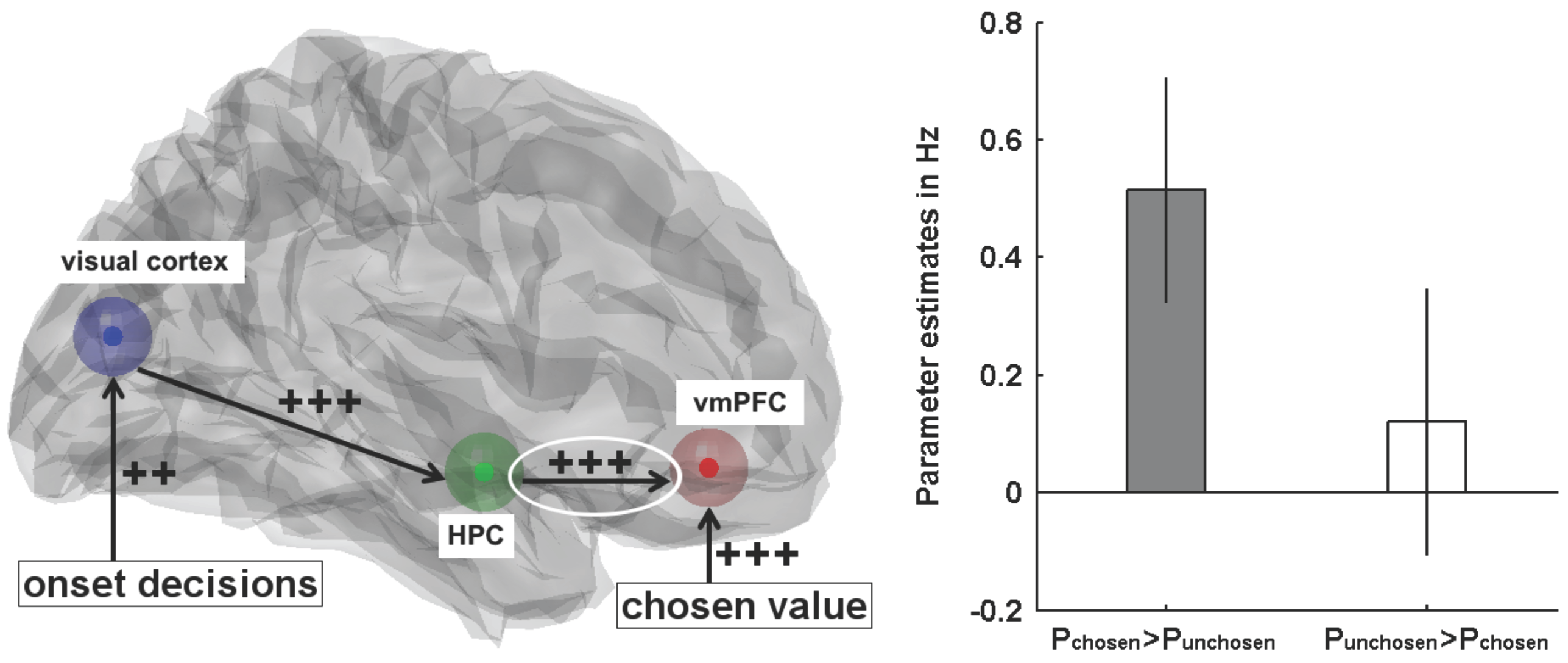
- Gluth, S.; Sommer, T.; Rieskamp, J.; Büchel, C. Effective connectivity between hippocampus and ventromedial prefrontal cortex controls preferential choices from memory. Neuron 2015, 86, 1078–1090. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.D.; Jadhav, S.P. Multiple modes of hippocampal–prefrontal interactions in memory-guided behavior. Curr. Opin. Neurobiol. 2016, 40, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Benchenane, K.; Peyrache, A.; Khamassi, M.; Tierney, P.L.; Gioanni, Y.; Battaglia, F.P.; Wiener, S.I. Coherent theta oscillations and reorganization of spike timing in the hippocampal-prefrontal network upon learning. Neuron 2010, 66, 921–936. [Google Scholar] [CrossRef] [PubMed]
- Guitart-Masip, M.; Barnes, G.R.; Horner, A.; Bauer, M.; Dolan, R.J.; Duzel, E. Synchronization of medial temporal lobe and prefrontal rhythms in human decision making. J. Neurosci. 2013, 33, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G. Theta oscillations in the hippocampus. Neuron 2002, 33, 325–340. [Google Scholar] [CrossRef]
- Rangel, L.M.; Rueckemann, J.W.; Riviere, P.D.; Keefe, K.R.; Porter, B.S.; Heimbuch, I.S.; Budlong, C.H.; Eichenbaum, H. Rhythmic coordination of hippocampal neurons during associative memory processing. eLife 2016, 5, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Colgin, L.L. Oscillations and hippocampal–prefrontal synchrony. Curr. Opin. Neurobiol. 2011, 21, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Spellman, T.; Rigotti, M.; Ahmari, S.E.; Fusi, S.; Gogos, J.A.; Gordon, J.A. Hippocampal–prefrontal input supports spatial encoding in working memory. Nature 2015, 522, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Sirota, A.; Montgomery, S.; Fujisawa, S.; Isomura, Y.; Zugaro, M.; Buzsáki, G. Entrainment of neocortical neurons and gamma oscillations by the hippocampal theta rhythm. Neuron 2008, 60, 683–697. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Harrison, L.; Penny, W. Dynamic causal modelling. Neuroimage 2003, 19, 1273–1302. [Google Scholar] [CrossRef]
- Wimmer, G.E.; Büchel, C. Reactivation of reward-related patterns from single past episodes supports memory-based decision making. J. Neurosci. 2016, 36, 2868–2880. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Morris, R.G.M. Hippocampal-neocortical interactions in memory formation, consolidation, and reconsolidation. Annu. Rev. Psychol. 2010, 61, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Euston, D.R.; Gruber, A.J.; McNaughton, B.L. The role of medial prefrontal cortex in memory and decision making. Neuron 2012, 76, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuis, I.L.C.; Takashima, A. The role of the ventromedial prefrontal cortex in memory consolidation. Behav. Brain Res. 2011, 218, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Preston, A.R.; Eichenbaum, H. Interplay of hippocampus and prefrontal cortex in memory. Curr. Biol. 2013, 23, R764–R773. [Google Scholar] [CrossRef] [PubMed]
- Piaget, J. The Child’s Conception of the World; Routldge & Kegan Paul Ltd.: London, UK, 1929. [Google Scholar]
- Wang, J.X.; Cohen, N.J.; Voss, J.L. Covert rapid action-memory simulation (CRAMS): A hypothesis of hippocampal–prefrontal interactions for adaptive behavior. Neurobiol. Learn. Mem. 2015, 117, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Addis, D.R.; Schacter, D.L. The hippocampus and imagining the future: Where do we stand? Front. Hum. Neurosci. 2011, 5, 173. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.C.; Schacter, D.L.; Corballis, M.C.; Addis, D.R. A role for the hippocampus in encoding simulations of future events. Proc. Natl. Acad. Sci. USA 2011, 108, 13858–13863. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, B.E.; Foster, D.J. Hippocampal place-cell sequences depict future paths to remembered goals. Nature 2013, 497, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Schacter, D.L.; Addis, D.R.; Hassabis, D.; Martin, V.C.; Spreng, R.N.; Szpunar, K.K. The Future of Memory: Remembering, Imagining, and the Brain. Neuron 2012, 76, 677–694. [Google Scholar] [CrossRef] [PubMed]
- Mack, M.L.; Preston, A.R. Decisions about the past are guided by reinstatement of specific memories in the hippocampus and perirhinal cortex. Neuroimage 2016, 127, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.J.; Bradley, M.M.; Cuthbert, B.N. International Affective Picture System (IAPS): Technical Manual and Affective Ratings. Available online: http://www2.unifesp.br/dpsicobio/adap/instructions.pdf (accessed on 12 December 2016).
- Palombo, D.J.; Keane, M.M.; Verfaellie, M. How does the hippocampus shape decisions? Neurobiol. Learn. Mem. 2015, 125, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ghim, J.-W.; Kim, H.; Lee, D.; Jung, M. Hippocampal neural correlates for values of experienced events. J. Neurosci. 2012, 32, 15053–15065. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, G.E.; Shohamy, D. Preference by association: How memory mechanisms in the hippocampus bias decisions. Science 2012, 338, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Hölscher, C.; Jacob, W.; Mallot, H.A. Reward modulates neuronal activity in the hippocampus of the rat. Behav. Brain Res. 2003, 142, 181–191. [Google Scholar] [CrossRef]
- Rolls, E.T.; Xiang, J.-Z. Reward-spatial view representations and learning in the primate hippocampus. J. Neurosci. 2005, 25, 6167–6174. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weilbächer, R.A.; Gluth, S. The Interplay of Hippocampus and Ventromedial Prefrontal Cortex in Memory-Based Decision Making. Brain Sci. 2017, 7, 4. https://doi.org/10.3390/brainsci7010004
Weilbächer RA, Gluth S. The Interplay of Hippocampus and Ventromedial Prefrontal Cortex in Memory-Based Decision Making. Brain Sciences. 2017; 7(1):4. https://doi.org/10.3390/brainsci7010004
Chicago/Turabian StyleWeilbächer, Regina A., and Sebastian Gluth. 2017. "The Interplay of Hippocampus and Ventromedial Prefrontal Cortex in Memory-Based Decision Making" Brain Sciences 7, no. 1: 4. https://doi.org/10.3390/brainsci7010004
APA StyleWeilbächer, R. A., & Gluth, S. (2017). The Interplay of Hippocampus and Ventromedial Prefrontal Cortex in Memory-Based Decision Making. Brain Sciences, 7(1), 4. https://doi.org/10.3390/brainsci7010004