
Differential Effects of Light Spectra on Sleep Architecture and Melatonin in Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Designs
- Sleep Behavior Experiment: Twenty-four rats were randomly assigned to blue or orange light groups for light phase (ZT4–ZT12) and dark-phase exposure (ZT16–ZT24) (n = 6 for each group). Another twenty-four rats who received no additional light exposure were used as control to match the light exposure groups (n = 6 for each group). EEG data were collected during 8 h light exposure over 24 h and in the subsequent 24 h, as indicated in Figure 1A.
- Melatonin Experiment: Six rats received dark-phase blue light, with serum collected at four post-exposure time points. Baseline melatonin data were derived from unexposed controls (n = 5). Protocols are detailed in Figure 1B.
2.3. Light Exposure Protocol
2.4. EEG Electrode Implantation and Recording
2.4.1. Surgical Procedure
2.4.2. Recording and Scoring
2.5. Melatonin Quantification
2.6. Statistical Analysis
3. Results
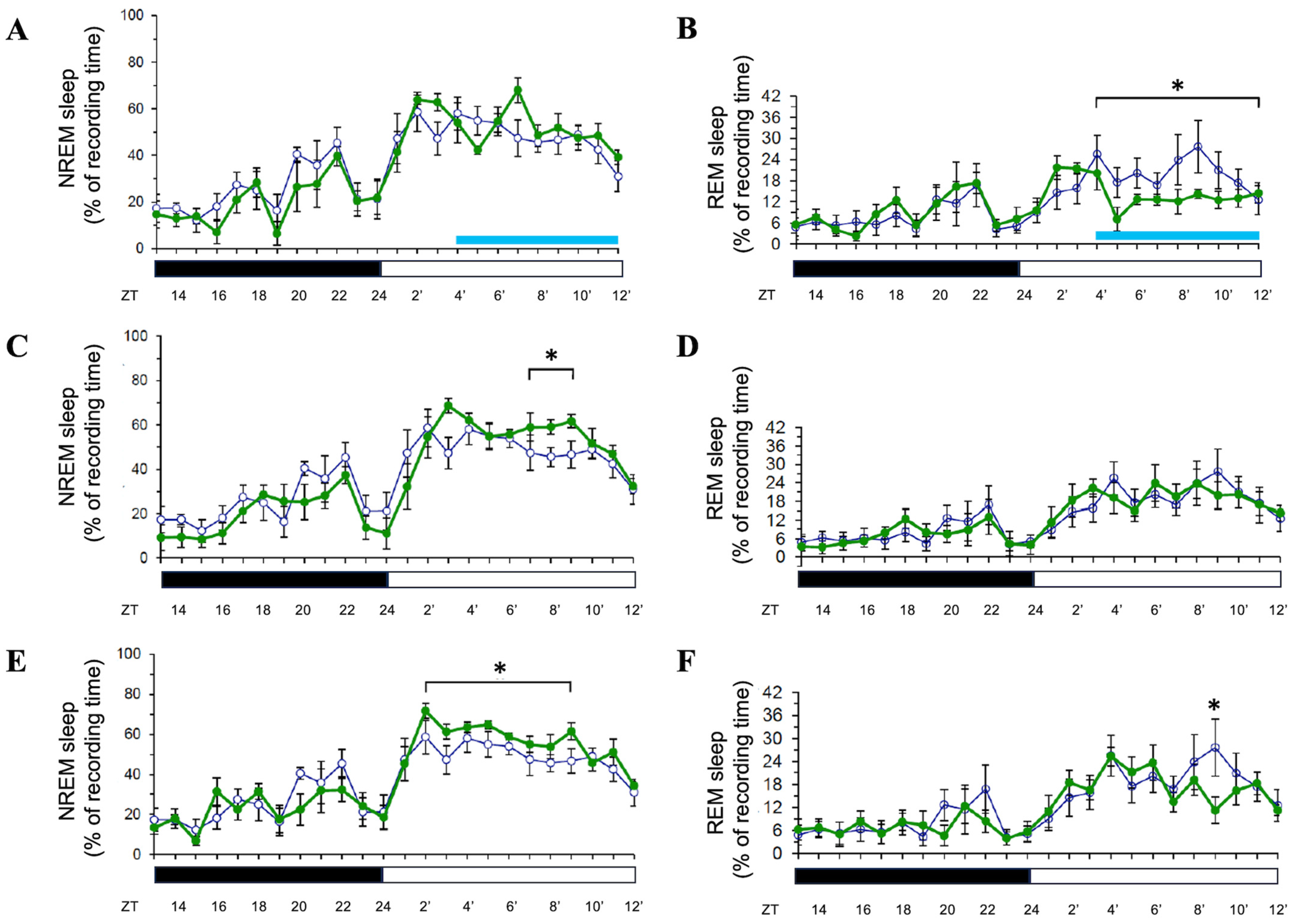
3.1. Effects of 8 h Blue Light Exposure During the Light Period
3.2. Effects of 8 h Orange Light Exposure During the Light Period
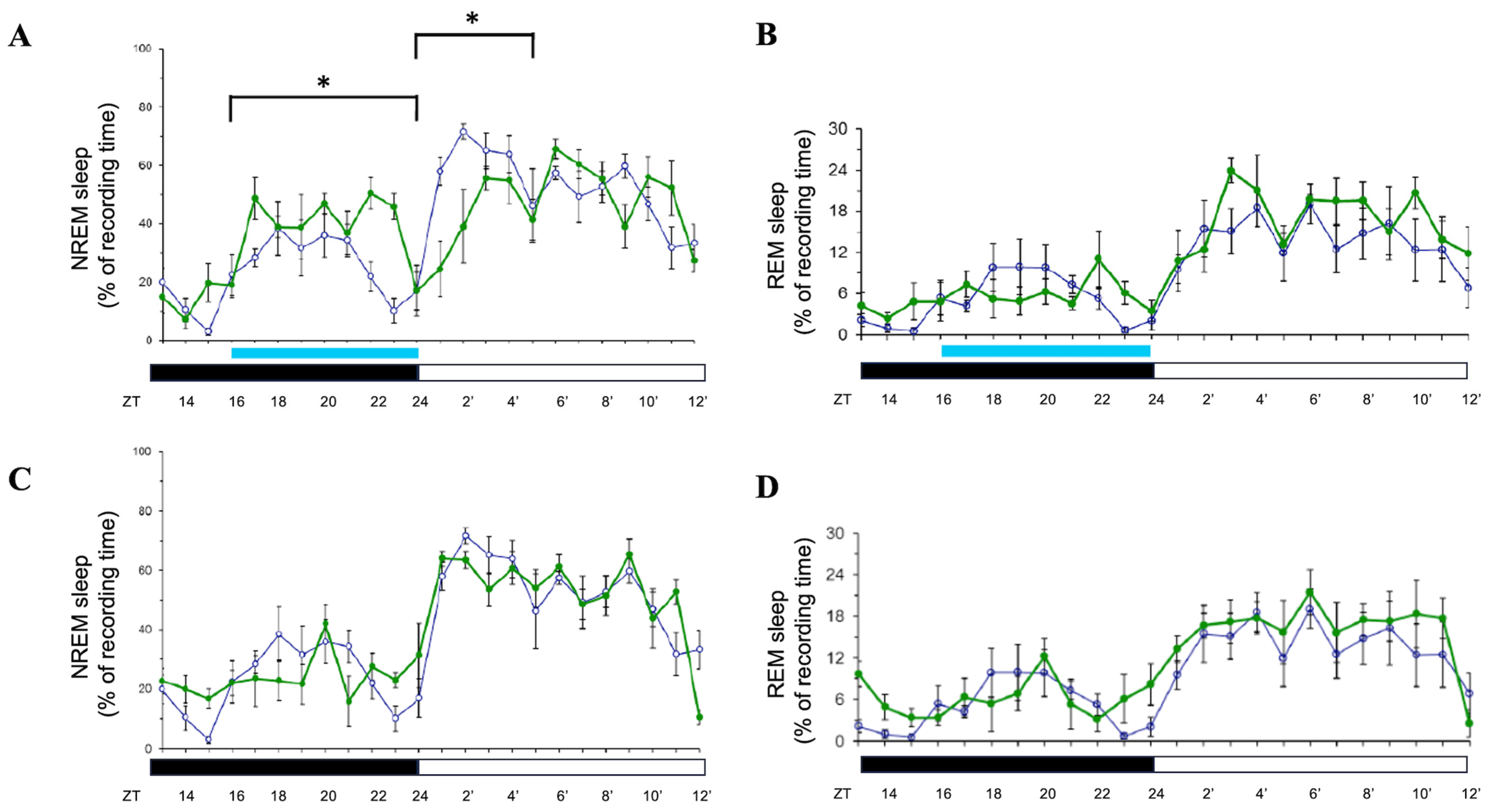
3.3. Effects of 8 h Blue Light or Ornage Light Exposure During the Dark Period
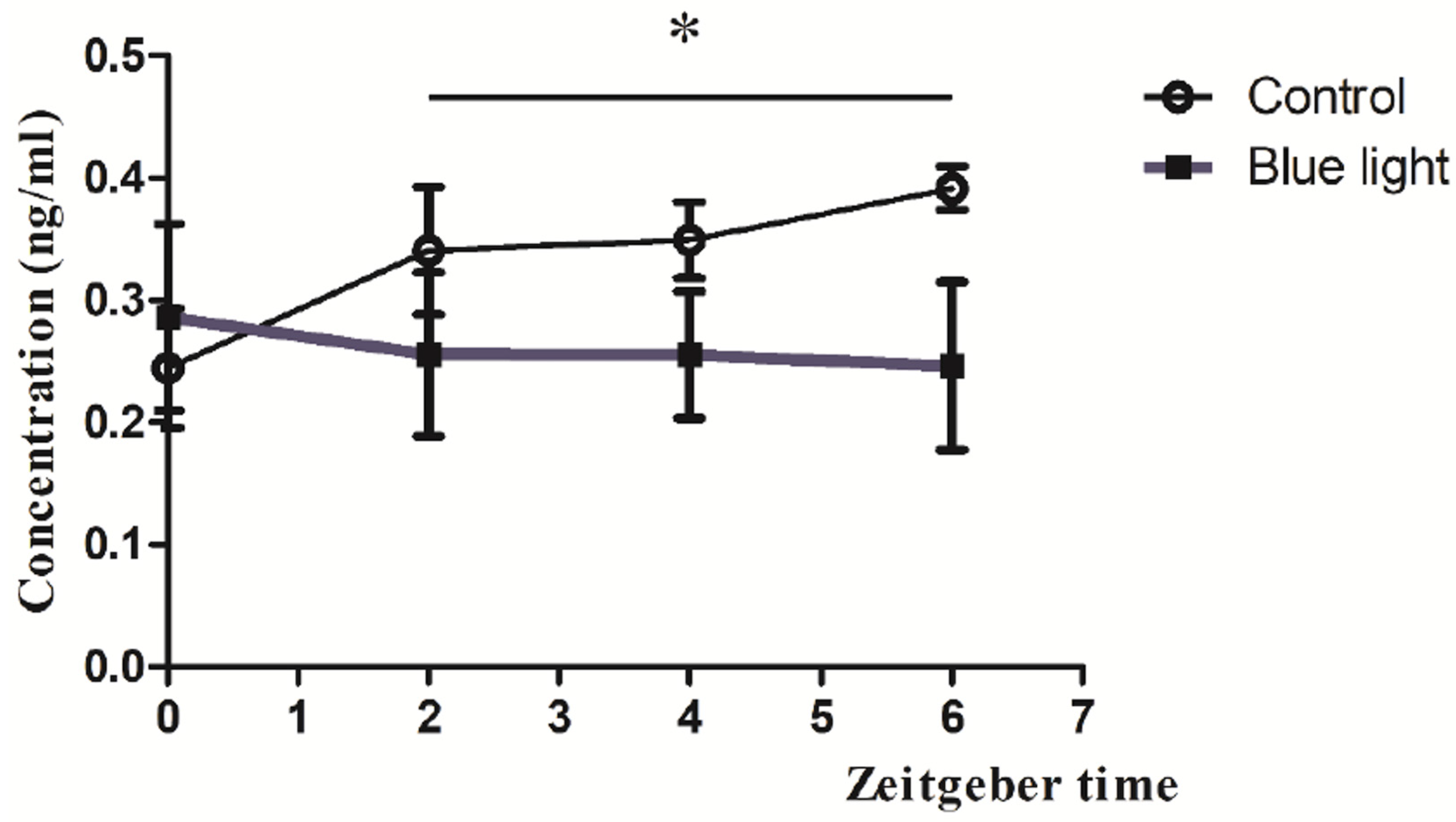
3.4. Melatonin Concentrations Post-Blue Light
4. Discussion
4.1. Blue Light During the Light Period: Selective REM Suppression and Delayed NREM Enhancement
4.2. Orange Light During the Light Period: Biphasic REM and Delayed NREM Effects
4.3. Blue Light During the Dark Period: Transient NREM Boost and Suppression
4.4. Mechanistic Insights and Spectral–Timing Interactions
4.5. Possible Mechanisms of Melatonin Suppression by Blue Light Exposure and Implications for Health and Circadian Regulation
4.6. Translational Relevance and Implications for Human Sleep Hygiene
4.7. Implications and Limitations
4.8. Future Directions
4.9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AP | anterior–posterior |
| EEG | electroencephalogram |
| ip | intraperitoneal |
| ipRGC | intrinsically photosensitive retrinal ganglion cell |
| LED | light-emitting diodes |
| ML | medial–lateral |
| NREM | non-rapid eye movement |
| REM | rapid eye movement |
| SCN | suprachiasmatic nucleus |
| ZT | zeitgeber time |
References
- Gringras, P.; Middleton, B.; Skene, D.J.; Revell, V.L. Bigger, brighter, bluer-better? Current light-emitting devices—Adverse sleep properties and preventative strategies. Front. Public Health 2015, 3, 233. [Google Scholar] [CrossRef] [PubMed]
- Chellappa, S.L.; Steiner, R.; Blattner, P.; Oelhafen, P.; Götz, T.; Cajochen, C. Non-visual effects of light on melatonin, alertness and cognitive performance: Can blue-enriched light keep us alert? PLoS ONE 2011, 6, e16429. [Google Scholar] [CrossRef] [PubMed]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef] [PubMed]
- Hattar, S.; Lucas, R.J.; Mrosovsky, N.; Thompson, S.; Douglas, R.H.; Hankins, M.W.; Yau, K.W. Melanopsin and rod-cone photoreceptive systems account for all major accessory visual functions in mice. Nature 2003, 424, 76–81. [Google Scholar] [CrossRef]
- Lockley, S.W.; Brainard, G.C.; Czeisler, C.A. High sensitivity of the human circadian melatonin rhythm to resetting by short wavelength light. J. Clin. Endocrinol. Metab. 2003, 88, 4502–4505. [Google Scholar] [CrossRef]
- Cajochen, C.; Münch, M.; Kobialka, S.; Kräuchi, K.; Steiner, R.; Oelhafen, P.; Orgül, S.; Wirz-Justice, A. High sensitivity of human melatonin, alertness, thermoregulation, and heart rate to short wavelength light. J. Clin. Endocrinol. Metab. 2005, 90, 1311–1316. [Google Scholar] [CrossRef]
- Chang, A.M.; Aeschbach, D.; Duffy, J.F.; Czeisler, C.A. Evening use of light-emitting eReaders negatively affects sleep, circadian timing, and next-morning alertness. Proc. Natl. Acad. Sci. USA 2015, 112, 1232–1237. [Google Scholar] [CrossRef]
- Borbély, A.A.; Daan, S.; Wirz-Justice, A.; Deboer, T. The two-process model of sleep regulation: A reappraisal. J. Sleep Res. 2016, 25, 131–143. [Google Scholar] [CrossRef]
- Lucas, R.J.; Lall, G.S.; Allen, A.E.; Brown, T.M. How rod, cone, and melanopsin photoreceptors come together to enlighten the mammalian circadian clock. Prog. Brain Res. 2012, 199, 1–18. [Google Scholar] [CrossRef]
- Golombek, D.A.; Rosenstein, R.E. Physiology of circadian entrainment. Physiol. Rev. 2010, 90, 1063–1102. [Google Scholar] [CrossRef]
- Siegel, J.M. Do all animals sleep? Trends Neurosci. 2008, 31, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Muindi, F.; Zeitzer, J.M.; Colas, D.; Heller, H.C. The acute effects of light on murine sleep during the dark phase: Importance of melanopsin for maintenance of light-induced sleep. Eur. J. Neurosci. 2013, 37, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Pilorz, V.; Tam, S.K.; Hughes, S.; Pothecary, C.A.; Jagannath, A.; Hankins, M.W.; Bannerman, D.M.; Lightman, S.L.; Vyazovskiy, V.V.; Nolan, P.M.; et al. Melanopsin regulates both sleep-promoting and arousal-promoting responses to light. PLoS Biol. 2016, 14, e1002482. [Google Scholar] [CrossRef]
- Saper, C.B.; Scammell, T.E.; Lu, J. Hypothalamic regulation of sleep and circadian rhythms. Nature 2005, 437, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gan, L.; Luo, D.; Sun, C. Melatonin promotes circadian rhythm-induced proliferation through Clock/histone deacetylase 3/c-Myc interaction in mouse adipose tissue. J. Pineal Res. 2017, 62, e12383. [Google Scholar] [CrossRef]
- Chang, F.C.; Opp, M.R. Blockade of corticotropin-releasing hormone receptors reduces spontaneous waking in the rat. Am. J. Physiol. 1998, 275, R793–R802. [Google Scholar] [CrossRef]
- West, K.E.; Jablonski, M.R.; Warfield, B.; Cecil, K.S.; James, M.; Ayers, M.A.; Maida, J.; Bowen, C.; Sliney, D.H.; Rollag, M.D.; et al. Blue light from light-emitting diodes elicits a dose-dependent suppression of melatonin in humans. J. Appl. Physiol. 2011, 110, 619–626. [Google Scholar] [CrossRef]
- Gooley, J.J.; Chamberlain, K.; Smith, K.A.; Khalsa, S.B.; Rajaratnam, S.M.; Van Reen, E.; Zeitzer, J.M.; Czeisler, C.A.; Lockley, S.W. Exposure to room light before bedtime suppresses melatonin onset and shortens melatonin duration in humans. J. Clin. Endocrinol. Metab. 2011, 96, E463–E472. [Google Scholar] [CrossRef]
- Lee, S.I.; Matsumori, K.; Nishimura, K.; Nishimura, Y.; Ikeda, Y.; Eto, T.; Higuchi, S. Melatonin suppression and sleepiness in children exposed to blue-enriched white LED lighting at night. Physiol. Rep. 2018, 6, e13942. [Google Scholar] [CrossRef]
- Hanifin, J.P.; Lockley, S.W.; Cecil, K.; West, K.; Jablonski, M.; Warfield, B.; James, M.; Ayers, M.; Byrne, B.; Gerner, E.; et al. Randomized trial of polychromatic blue-enriched light for circadian phase shifting, melatonin suppression, and alerting responses. Physiol. Behav. 2019, 198, 57–66. [Google Scholar] [CrossRef]
- Bryk, A.A.; Blagonravov, M.L.; Goryachev, V.A.; Chibisov, S.M.; Azova, M.M.; Syatkin, S.P. Daytime Exposure to Blue Light Alters Cardiovascular Circadian Rhythms, Electrolyte Excretion and Melatonin Production. Pathophysiology 2022, 29, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Tobeiha, M.; Jafari, A.; Fadaei, S.; Mirazimi, S.M.A.; Dashti, F.; Amiri, A.; Khan, H.; Asemi, Z.; Reiter, R.J.; Hamblin, M.R.; et al. Evidence for the Benefits of Melatonin in Cardiovascular Disease. Front. Cardiovasc. Med. 2022, 9, 888319. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, M.; Uchiumi, T.; Takahata, M.; Yamaki, M.; Sato, T. Effects of pre-bedtime blue-light exposure on ratio of deep sleep in healthy young men. Sleep Med. 2021, 84, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Hester, L.; Dang, D.; Barker, C.J.; Heath, M.; Mesiya, S.; Tienabeso, T.; Watson, K. Evening wear of blue-blocking glasses for sleep and mood disorders: A systematic review. Chronobiol. Int. 2021, 38, 1375–1383. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, Y.-C.; Yi, P.-L.; Chang, F.-C. Differential Effects of Light Spectra on Sleep Architecture and Melatonin in Rats. Brain Sci. 2025, 15, 445. https://doi.org/10.3390/brainsci15050445
Chiu Y-C, Yi P-L, Chang F-C. Differential Effects of Light Spectra on Sleep Architecture and Melatonin in Rats. Brain Sciences. 2025; 15(5):445. https://doi.org/10.3390/brainsci15050445
Chicago/Turabian StyleChiu, Yuan-Chun, Pei-Lu Yi, and Fang-Chia Chang. 2025. "Differential Effects of Light Spectra on Sleep Architecture and Melatonin in Rats" Brain Sciences 15, no. 5: 445. https://doi.org/10.3390/brainsci15050445
APA StyleChiu, Y.-C., Yi, P.-L., & Chang, F.-C. (2025). Differential Effects of Light Spectra on Sleep Architecture and Melatonin in Rats. Brain Sciences, 15(5), 445. https://doi.org/10.3390/brainsci15050445