
Spinal Involvement of TRPV1 and PI3K/AKT/mTOR Pathway During Chronic Postoperative Pain in Mice


 ,
, 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
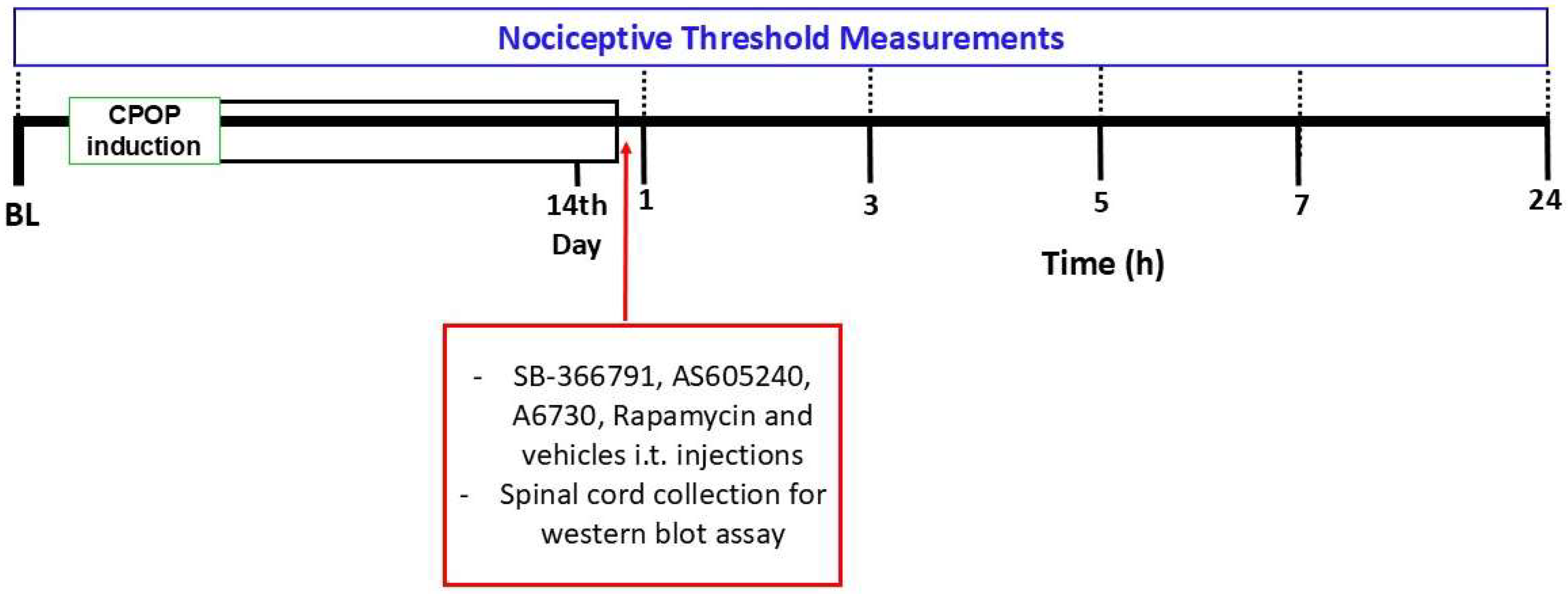
2.2. Experimental Design
2.3. Postoperative Chronic Pain Model
2.4. Nociceptive Threshold Assessment
2.5. Drugs
2.6. Intrathecal Injection
2.7. TRPV1, PI3K, AKT, and mTOR Protein Level Assessment
2.8. Statistical Analysis
3. Results
3.1. Spinal Involvement of TRPV1 and PI3K/AKT/mTOR Pathway in CPOP-Induced Nociception
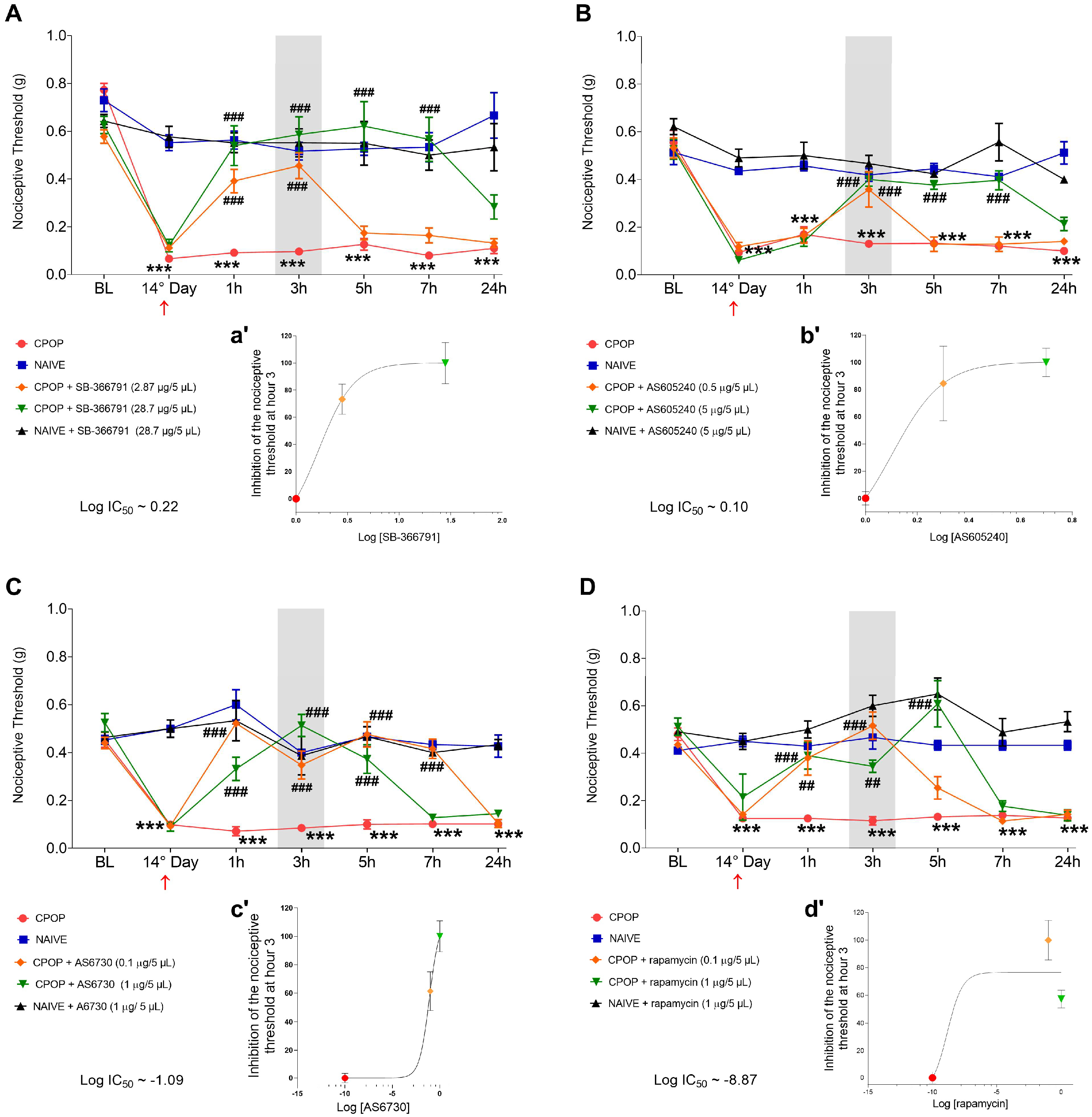
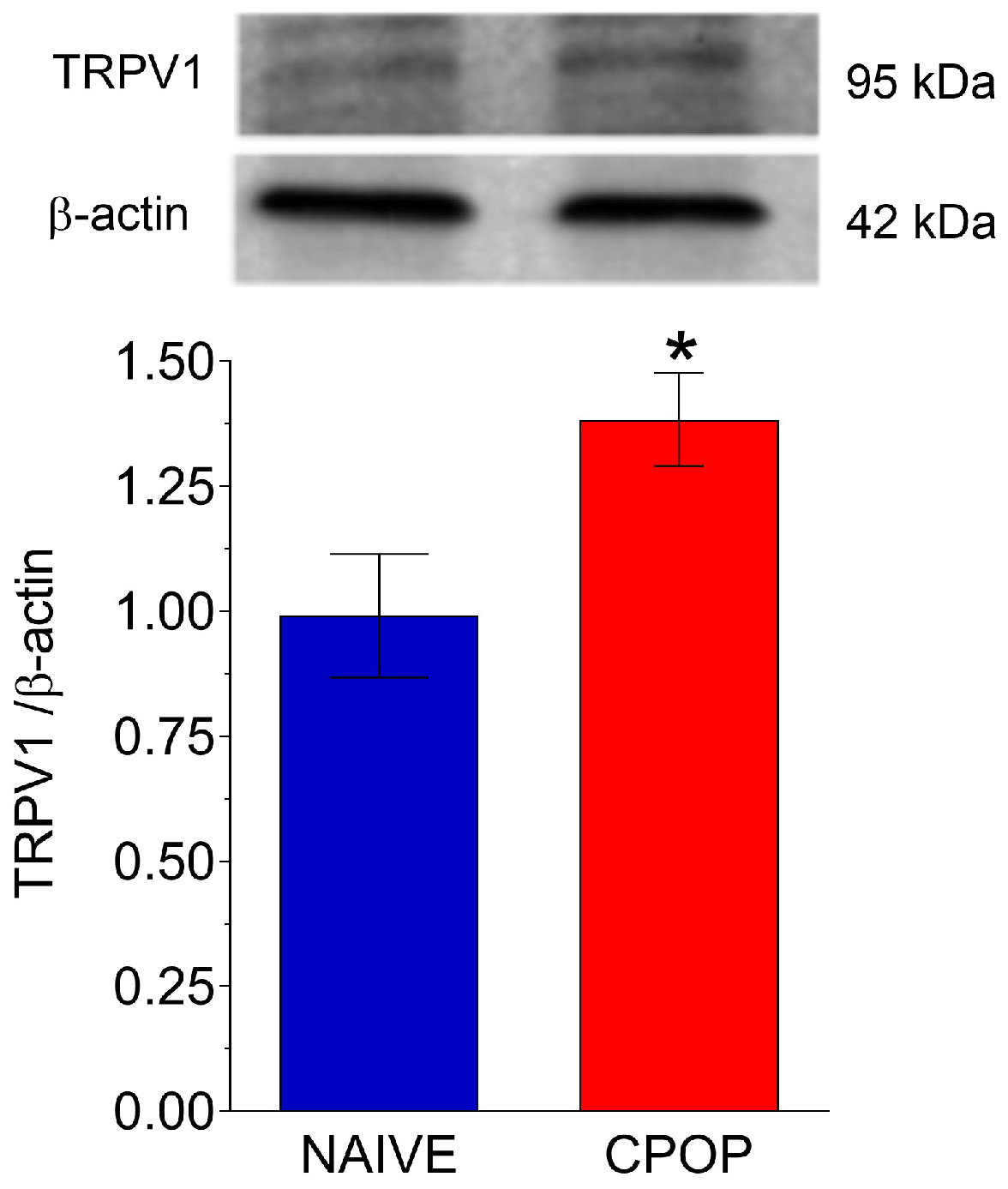
3.2. TRPV1 Protein Levels During CPOP
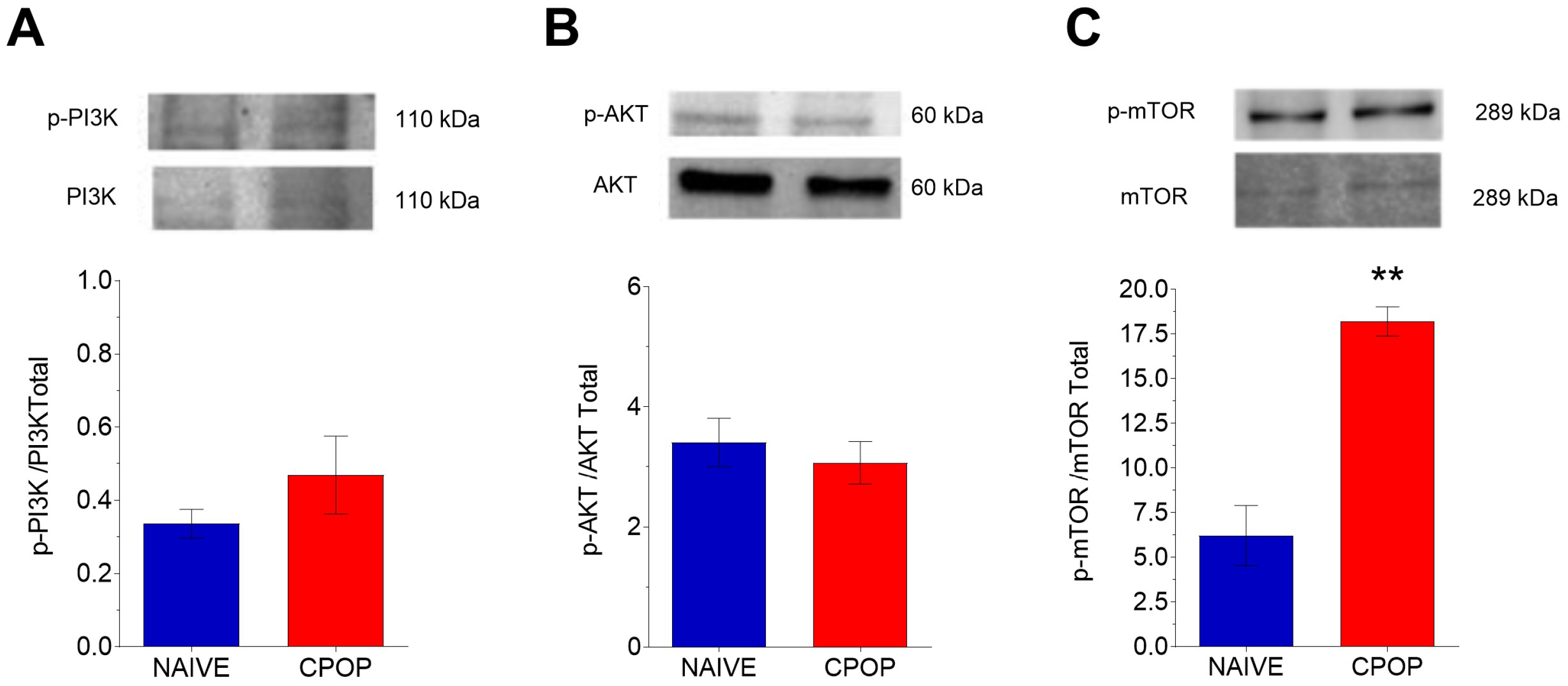
3.3. PI3K/AKT/mTOR Protein Levels During CPOP
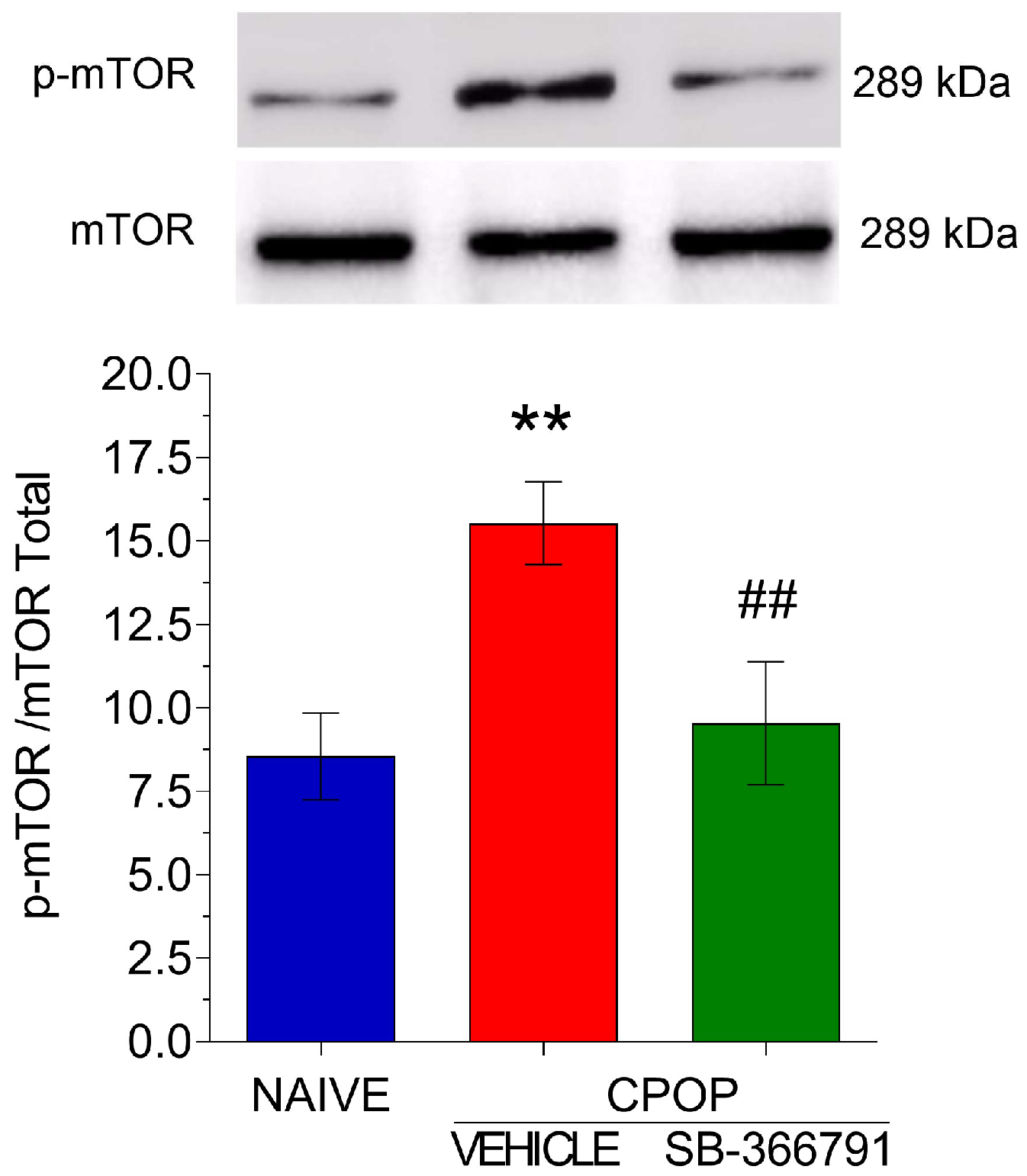
3.4. TRPV1 Involvement in mTOR Protein Activation in the Spinal Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fuller, A.M.; Bharde, S.; Sikandar, S. The Mechanisms and Management of Persistent Postsurgical Pain. Front. Pain Res. 2023, 4, 1154597. [Google Scholar] [CrossRef] [PubMed]
- Brigham, N.C.; Ji, R.-R.; Becker, M.L. Degradable Polymeric Vehicles for Postoperative Pain Management. Nat. Commun. 2021, 12, 1367. [Google Scholar] [CrossRef] [PubMed]
- Health Canada: Working Together to Better Understand, Prevent, and Manage Chronic Pain: What We Heard; MDHealth Canada: Ottawa, ON, Canada; Hagerstown, MD, USA, 2021.
- Chou, R.; Gordon, D.B.; de Leon-Casasola, O.A.; Rosenberg, J.M.; Bickler, S.; Brennan, T.; Carter, T.; Cassidy, C.L.; Chittenden, E.H.; Degenhardt, E.; et al. Management of Postoperative Pain: A Clinical Practice Guideline from the American Pain Society, the American Society of Regional Anesthesia and Pain Medicine, and the American Society of Anesthesiologists’ Committee on Regional Anesthesia, Executive Comm. J. Pain 2016, 17, 131–157. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.B.S.; Bonilla, P.; Kraychete, D.C.; Flores, F.C.; de Valtolina, E.D.P.; Guerrero, C. Optimizing post-operative pain management in Latin America. Rev. Bras. Anestesiol. 2017, 67, 395–403. [Google Scholar] [CrossRef]
- Elokely, K.; Velisetty, P.; Delemotte, L.; Palovcak, E.; Klein, M.L.; Rohacs, T.; Carnevale, V. Understanding TRPV1 Activation by Ligan.;s: Insights from the Binding Modes of Capsaicin and Resiniferatoxin. Proc. Natl. Acad. Sci. USA 2016, 113, E137–E145. [Google Scholar] [CrossRef]
- Jardín, I.; López, J.J.; Diez, R.; Sánchez-Collado, J.; Cantonero, C.; Albarrán, L.; Woodard, G.E.; Redondo, P.C.; Salido, G.M.; Smani, T.; et al. TRPs in Pain Sensation. Front Physiol. 2017, 8, 392. [Google Scholar] [CrossRef]
- Guo, J.-R.; Wang, H.; Jin, X.-J.; Jia, D.-L.; Zhou, X.; Tao, Q. Effect and Mechanism of Inhibition of PI3K/Akt/MTOR Signal Pathway on Chronic Neuropathic Pain and Spinal Microglia in a Rat Model of Chronic Constriction Injury. Oncotarget 2017, 8, 52923–52934. [Google Scholar] [CrossRef]
- Liu, W.; Lv, Y.; Ren, F. PI3K/Akt Pathway Is Required for Spinal Central Sensitization in Neuropathic Pain. Cell. Mol. Neurobiol. 2018, 38, 747–755. [Google Scholar] [CrossRef]
- Xu, B.; Liu, S.-S.; Wei, J.; Jiao, Z.-Y.; Mo, C.; Lv, C.-M.; Huang, A.-L.; Chen, Q.-B.; Ma, L.; Guan, X.-H. Role of Spinal Cord Akt-MTOR Signaling Pathways in Postoperative Hyperalgesia Induced by Plantar Incision in Mice. Front. Neurosci. 2020, 14, 766. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Wang, H.; Su, Z.; Pang, X. Neuroinflammation and Central PI3K/Akt/MTOR Signal Pathway Contribute to Bone Cancer Pain. Mol. Pain 2019, 15, 1744806919830240. [Google Scholar] [CrossRef]
- Ching, L.-C.; Kou, Y.R.; Shyue, S.-K.; Su, K.-H.; Wei, J.; Cheng, L.-C.; Yu, Y.-B.; Pan, C.-C.; Lee, T.-S. Molecular Mechanisms of Activation of Endothelial Nitric Oxide Synthase Mediated by Transient Receptor Potential Vanilloid Type 1. Cardiovasc. Res. 2011, 91, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M. Ethical Guidelines for Investigations of Experimental Pain in Conscious Animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Elisei, L.M.S.; Moraes, T.R.; Malta, I.H.; Charlie-Silva, I.; Sousa, I.M.O.; Veras, F.P.; Foglio, M.A.; Fraceto, L.F.; Galdino, G. Antinociception Induced by Artemisinin Nanocapsule in a Model of Postoperative Pain via Spinal TLR4 Inhibition. Inflammopharmacology 2020, 28, 1537–1551. [Google Scholar] [CrossRef] [PubMed]
- Santos, G.X.; Barbosa, D.; de-Souza, G.G.; Kosour, C.; Parizotto, N.A.; Dos Reis, L.M. Central Involvement of 5-HT1A Receptors in Antinociception Induced by Photobiomodulation in Animal Model of Neuropathic Pain. Lasers Med. Sci. 2022, 37, 821–829. [Google Scholar] [CrossRef]
- de Oliveira, H.U.; Dos Santos, R.S.; Malta, I.H.S.; Pinho, J.P.; Almeida, A.F.S.; Sorgi, C.A.; Peti, A.P.F.; Xavier, G.S.; Dos Reis, L.M.; Faccioli, L.H.; et al. Investigation of the Involvement of the Endocannabinoid System in TENS-Induced Antinociception. J. Pain 2020, 21, 820–835. [Google Scholar] [CrossRef]
- Fernandes, E.S.; Russell, F.A.; Spina, D.; McDougall, J.J.; Graepel, R.; Gentry, C.; Staniland, A.A.; Mountford, D.M.; Keeble, J.E.; Malcangio, M.; et al. A Distinct Role for Transient Receptor Potential Ankyrin 1, in Addition to Transient Receptor Potential Vanilloid 1, in Tumor Necrosis Factor α-Induced Inflammatory Hyperalgesia and Freund’s Complete Adjuvant-Induced Monarthritis. Arthritis Rheum. 2011, 63, 819–829. [Google Scholar] [CrossRef]
- Manjavachi, M.N.; Passos, G.F.; Trevisan, G.; Araújo, S.B.; Pontes, J.P.; Fernandes, E.S.; Costa, R.; Calixto, J.B. Spinal Blockage of CXCL1 and Its Receptor CXCR2 Inhibits Paclitaxel-Induced Peripheral Neuropathy in Mice. Neuropharmacology 2019, 151, 136–143. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, X.-F.; Bo, J.-H.; Zhang, J.; Liu, X.-J.; Wu, L.-P.; Ma, Z.-L.; Gu, X.-P. Activation of MTOR in the Spinal Cord Is Required for Pain Hypersensitivity Induced by Chronic Constriction Injury in Mice. Pharmacol. Biochem. Behav. 2013, 111, 64–70. [Google Scholar] [CrossRef]
- Hylden, J.L.; Wilcox, G.L. Intrathecal Morphine in Mice: A New Technique. Eur. J. Pharmacol. 1980, 67, 313–316. [Google Scholar] [CrossRef]
- Izumi, Y.; Sasaki, M.; Hashimoto, S.; Sawa, T.; Amaya, F. MTOR Signaling Controls VGLUT2 Expression to Maintain Pain Hypersensitivity after Tissue Injury. Neuroscience 2015, 308, 169–179. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Jin, H.; Cong, P.; Zhang, Y.; Tong, C.; Shi, X.; Liu, X.; Tong, Z.; Shi, L.; et al. Cold Stress-Induced Brain Injury Regulates TRPV1 Channels and the PI3K/AKT Signaling Pathway. Brain Res. 2017, 1670, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qin, X.; Lu, X.; Jiang, J. Effects of Inhibiting the PI3K/Akt/MTOR Signaling Pathway on the Pain of Sciatic Endometriosis in a Rat Model. Can. J. Physiol. Pharmacol. 2019, 97, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Pogatzki-Zahn, E.M.; Shimizu, I.; Caterina, M.; Raja, S.N. Heat Hyperalgesia after Incision Requires TRPV1 and Is Distinct from Pure Inflammatory Pain. Pain 2005, 115, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Gosu, V.; Basith, S.; Hong, S.; Choi, S. Polymodal Transient Receptor Potential Vanilloid Type 1 Nocisensor: Structure, Modulators, and Therapeutic Applications. Adv. Protein Chem. Struct. Biol. 2016, 104, 81–125. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Luongo, L.; de Novellis, V.; Berrino, L.; Rossi, F.; Maione, S. Moving towards Supraspinal TRPV1 Receptors for Chronic Pain Relief. Mol. Pain 2010, 6, 66. [Google Scholar] [CrossRef]
- Nicol, G.D.; Lopshire, J.C.; Pafford, C.M. Tumor Necrosis Factor Enhances the Capsaicin Sensitivity of Rat Sensory Neurons. J. Neurosci. Off. J. Soc. Neurosci. 1997, 17, 975–982. [Google Scholar] [CrossRef]
- Yoo, S.; Lim, J.Y.; Hwang, S.W. Sensory TRP Channel Interactions with Endogenous Lipids and Their Biological Outcomes. Molecules 2014, 19, 4708–4744. [Google Scholar] [CrossRef]
- Kim, Y.H.; Back, S.K.; Davies, A.J.; Jeong, H.; Jo, H.J.; Chung, G.; Na, H.S.; Bae, Y.C.; Kim, S.J.; Kim, J.S.; et al. TRPV1 in GABAergic Interneurons Mediates Neuropathic Mechanical Allodynia and Disinhibition of the Nociceptive Circuitry in the Spinal Cord. Neuron 2012, 74, 640–647. [Google Scholar] [CrossRef]
- Ferrini, F.; Salio, C.; Lossi, L.; Gambino, G.; Merighi, A. Modulation of Inhibitory Neurotransmission by the Vanilloid Receptor Type 1 (TRPV1) in Organotypically Cultured Mouse Substantia Gelatinosa Neurons. Pain 2010, 150, 128–140. [Google Scholar] [CrossRef]
- Moriyama, T.; Higashi, T.; Togashi, K.; Iida, T.; Segi, E.; Sugimoto, Y.; Tominaga, T.; Narumiya, S.; Tominaga, M. Sensitization of TRPV1 by EP1 and IP Reveals Peripheral Nociceptive Mechanism of Prostaglandins. Mol. Pain 2005, 1, 3. [Google Scholar] [CrossRef]
- Maiese, K. Warming Up to New Possibilities with the Capsaicin Receptor TRPV1: MTOR, AMPK, and Erythropoietin. Curr. Neurovasc. Res. 2017, 14, 184–189. [Google Scholar] [CrossRef]
- Shang, Y.C.; Chong, Z.Z.; Wang, S.; Maiese, K. Tuberous Sclerosis Protein 2 (TSC2) Modulates CCN4 Cytoprotection during Apoptotic Amyloid Toxicity in Microglia. Curr. Neurovasc. Res. 2013, 10, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.-L.; Tee, A.R.; Short, J.D.; Bergeron, J.M.; Kim, J.; Shen, J.; Guo, R.; Johnson, C.L.; Kiguchi, K.; Walker, C.L. Activity of TSC2 Is Inhibited by AKT-Mediated Phosphorylation and Membrane Partitioning. J. Cell Biol. 2006, 173, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Su, Z.; Wang, H.; Pang, X. Involvement of Pro-Inflammation Signal Pathway in Inhibitory Effects of Rapamycin on Oxaliplatin-Induced Neuropathic Pain. Mol. Pain 2018, 14, 1744806918769426. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves Dos Santos, G.; Delay, L.; Yaksh, T.L.; Corr, M. Neuraxial Cytokines in Pain States. Front. Immunol. 2019, 10, 3061. [Google Scholar] [CrossRef] [PubMed]
- Barabas, M.E.; Stucky, C.L. TRPV1, but not TRPA1, in primary sensory neurons contributes to cutaneous incision-mediated hypersensitivity. Mol Pain. 2013, 9, 9. [Google Scholar] [CrossRef]
- Khan, M.I.; Khan, A.; Zafar, S.; Aslam, S.; Khanm, A.U.; Shal, B.; Haider, R.; Din, F.U.; Khan, S. Anti-nociceptive effects of magnolol via inhibition of TRPV1/P2Y and TLR4/NF-κB signaling in a postoperative pain model. Life Sci. 2023, 312, 121202. [Google Scholar] [CrossRef]
- Song, C.; Liu, P.; Zhao, Q.; Guo, S.; Wang, G. TRPV1 channel contributes to remifentanil-induced postoperative hyperalgesia via regulation of NMDA receptor trafficking in dorsal root ganglion. J. Pain Res. 2019, 12, 667–677. [Google Scholar] [CrossRef]
- Uchytilova, E.; Spicarova, D.; Palecek, J. TRPV1 antagonist attenuates postoperative hypersensitivity by central and peripheral mechanisms. Mol. Pain. 2014, 10, 67. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Z.; Liu, Y.; Xiao, S.; Liu, G. PKC regulates postoperative pain via ERK and mTOR pathways. Int. J. Clin. Exp. Med. 2019, 12, 632–638. [Google Scholar]
- Yang, J.; Yu, H.M.; Zhou, X.D.; Kolosov, V.P.; Perelman, J.M. Study on TRPV1-Mediated Mechanism for the Hypersecretion of Mucus in Respiratory Inflammation. Mol. Immunol. 2013, 53, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Melick, C.H.; Jewell, J.L. Regulation of MTORC1 by Upstream Stimuli. Genes 2020, 11, 989. [Google Scholar] [CrossRef] [PubMed]
- Szwed, A.; Kim, E.; Jacinto, E. Regulation and metabolic functions of mTORC1 and mTORC2. Physiol. Rev. 2021, 101, 1371–1426. [Google Scholar] [CrossRef]
- Xu, Q.; Fitzsimmons, B.; Steinauer, J.; O’Neill, A.; Newton, A.C.; Hua, X.-Y.; Yaksh, T.L. Spinal Phosphinositide 3-Kinase-Akt-Mammalian Target of Rapamycin Signaling Cascades in Inflammation-Induced Hyperalgesia. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 2113–2124. [Google Scholar] [CrossRef]
- Tighe, P.J.; Riley, J.L., 3rd; Fillingim, R.B. Sex differences in the incidence of severe pain events following surgery: A review of 333,000 pain scores. Pain Med. 2014, 15, 1390–1404. [Google Scholar] [CrossRef]
- Gunthorpe, M.J.; Rami, H.K.; Jerman, J.C.; Smart, D.; Gill, C.H.; Soffin, E.M.; Luis Hannan, S.; Lappin, S.C.; Egerton, J.; Smith, G.D.; et al. Identification and characterisation of SB-366791, a potent and selective vanilloid receptor (VR1/TRPV1) antagonist. Neuropharmacology 2004, 46, 133–149. [Google Scholar] [CrossRef]
- Iftinca, M.; Defaye, M.; Altier, C. TRPV1-Targeted Drugs in Development for Human Pain Conditions. Drugs 2021, 81, 7–27. [Google Scholar] [CrossRef]
- Lisi, L.; Aceto, P.; Navarra, P.; Dello Russo, C. mTOR kinase: A possible pharmacological target in the management of chronic pain. BioMed Res. Int. 2015, 2015, 394257. [Google Scholar] [CrossRef]
- Ali, E.S.; Mitra, K.; Akter, S.; Ramproshad, S.; Mondal, B.; Khan, I.N.; Islam, M.T.; Sharifi-Rad, J.; Calina, D.; Cho, W.C. Recent advances and limitations of mTOR inhibitors in the treatment of cancer. Cancer Cell Int. 2022, 22, 284. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, G.X.; dos Anjos-Garcia, T.; Vieira, A.C.d.J.; Galdino, G. Spinal Involvement of TRPV1 and PI3K/AKT/mTOR Pathway During Chronic Postoperative Pain in Mice. Brain Sci. 2025, 15, 53. https://doi.org/10.3390/brainsci15010053
Santos GX, dos Anjos-Garcia T, Vieira ACdJ, Galdino G. Spinal Involvement of TRPV1 and PI3K/AKT/mTOR Pathway During Chronic Postoperative Pain in Mice. Brain Sciences. 2025; 15(1):53. https://doi.org/10.3390/brainsci15010053
Chicago/Turabian StyleSantos, Gabriela Xavier, Tayllon dos Anjos-Garcia, Ana Carolina de Jesus Vieira, and Giovane Galdino. 2025. "Spinal Involvement of TRPV1 and PI3K/AKT/mTOR Pathway During Chronic Postoperative Pain in Mice" Brain Sciences 15, no. 1: 53. https://doi.org/10.3390/brainsci15010053
APA StyleSantos, G. X., dos Anjos-Garcia, T., Vieira, A. C. d. J., & Galdino, G. (2025). Spinal Involvement of TRPV1 and PI3K/AKT/mTOR Pathway During Chronic Postoperative Pain in Mice. Brain Sciences, 15(1), 53. https://doi.org/10.3390/brainsci15010053