
Dopamine Concentration Changes Associated with the Retrodialysis of Methylone and 3,4-Methylenedioxypyrovalerone (MDPV) into the Caudate Putamen

Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
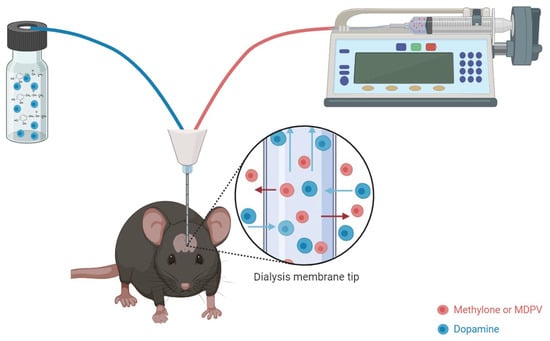
2. Materials and Methods
2.1. Animals
2.2. Drugs and Chemicals
2.3. In Vitro Probe Recovery Evaluation
2.4. Surgical Cannula Placement and Retrodialysis
2.5. High-Performance Liquid Chromatography–Electrochemical Detection (HPLC-EC)
2.6. Statistics
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Banks, M.L.; Worst, T.J.; Rusyniak, D.E.; Sprague, J.E. The synthetic cathinones (the “bath salts”). J. Emerg. Med. 2014, 46, 632–642. [Google Scholar] [CrossRef]
- Prosser, J.; Nelson, L. The toxicology of bath salts: A review of synthetic cathinones. J. Med. Toxicol. 2012, 8, 33–42. [Google Scholar] [CrossRef]
- Calinski, D.M.; Kisor, D.F.; Sprague, J.E. A review of the influence of functional group modifications to the core scaffold of synthetic cathinones on drug pharmacokinetics. Psychopharmacology 2019, 236, 881–890. [Google Scholar] [CrossRef]
- Kolanos, R.; Partilla, J.S.; Baumann, M.H.; Hutsell, B.A.; Banks, M.L.; Negus, S.S.; Glennon, R.A. Stereoselective actions of methylenedioxypyrovalerone (MDPV) to inhibit dopamine and norepinephrine transporters and facilitate intracranial self-stimulation in rats. ACS Chem. Neurosci. 2015, 6, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Gannon, B.M.; Rice, K.C.; Collins, G.T. Reinforcing effects of abused ‘bath salts’ constituents 3,4-methylenedioxypyrovalerone and α-pyrrolidinopentiophenone and their enantiomers. Behav. Pharmacol. 2017, 28, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Mondola, R. Synthetic cathinones: Chemistry, pharmacology and toxicology of a new class of designer drugs of abuse marketed as “bath salts” or “plant food”. Toxicol. Lett. 2012, 211, 144–149. [Google Scholar] [CrossRef]
- Simmler, L.; Buser, T.; Donzelli, M.; Schramm, Y.; Dieu, L.-H.; Huwyler, J.; Liechti, M. Pharmacological characterization of designer cathinones in vitro. Br. J. Pharmacol. 2013, 168, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Baumann, M.H.; Partilla, J.S.; Lehner, K.R.; Thorndike, E.B.; Hoffman, A.F.; Holy, M.; Rothman, R.B.; Goldberg, S.R.; Lupica, C.R.; Sitte, H.H.; et al. Powerful cocaine-like actions of 3,4-methylenedioxypyrovalerone (MDPV), a principal constituent of psychoactive ‘bath salts’products. Neuropsychopharmacology 2012, 38, 552–562. [Google Scholar] [CrossRef]
- Cozzi, N.V.; Sievert, M.K.; Shulgin, A.T.; Jacob, P.; Ruoho, A.E. Inhibition of plasma membrane monoamine transporters by beta-ketoamphetamines. Eur. J. Pharmacol. 1999, 381, 63–69. [Google Scholar] [CrossRef]
- Hadlock, G.C.; Webb, K.M.; McFadden, L.M.; Chu, P.W.; Ellis, J.D.; Allen, S.C.; Andrenyak, D.M.; Vieira-Brock, P.L.; German, C.L.; Conrad, K.M.; et al. 4-Methylmethcathinone (mephedrone): Neuropharmacological effects of a designer stimulant of abuse. J. Pharmacol. Exp. Ther. 2011, 339, 530–536. [Google Scholar] [CrossRef]
- Nagai, F.; Nonaka, R.; Satoh Hisashi Kamimura, K. The effects of non-medically used psychoactive drugs on monoamine neurotransmission in rat brain. Eur. J. Pharmacol. 2007, 559, 132–137. [Google Scholar] [CrossRef]
- Sogawa, C.; Sogawa, N.; Ohyama, K.; Kikura-Hanajiri, R.; Goda, Y.; Sora, I.; Kitayama, S. Methylone and monoamine transporters: Correlation with toxicity. Curr. Neuropharmacol. 2011, 9, 58–62. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Baumann, M.H.; Ayestas, M.A.; Partilla, J.S.; Sink, J.R.; Shulgin, A.T.; Daley, P.F.; Brandt, S.D.; Rothman, R.B.; Ruoho, A.E.; Cozzi, N.V. The designer methcathinone analogs, mephedrone and methylone, are substrates for monoamine transporters in brain tissue. Neuropsychopharmacology 2011, 37, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, K.G. Presynaptic regulation of dopamine release: Role of the DAT and VMAT2 transporters. Neurochem. Int. 2019, 122, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Suyama, J.A.; Sakloth, F.; Kolanos, R.; Glennon, R.A.; Lazenka, M.F.; Negus, S.S.; Banks, M.L. Abuse-Related Neurochemical Effects of Para-Substituted Methcathinone Analogs in Rats: Microdialysis Studies of Nucleus Accumbens Dopamine and Serotonin. J. Pharmacol. Exp. Ther. 2016, 356, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Schindler, C.W.; Thorndike, E.B.; Goldberg, S.R.; Lehner, K.R.; Cozzi, N.V.; Brandt, S.D.; Baumann, M.H. Reinforcing and neurochemical effects of the “bath salts” constituents 3,4-methylenedioxypyrovalerone (MDPV) and 3,4-methylenedioxy-N-methylcathinone (methylone) in male rats. Psychopharmacology 2015, 233, 1981–1990. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, N.V.; Brandt, S.D.; Daley, P.F.; Partilla, J.S.; Rothman, R.B.; Tulzer, A.; Sitte, H.H.; Baumann, M.H. Pharmacological examination of trifluoromethyl ring-substituted methcathione analogs. Eur. J. Pharmacol. 2013, 699, 180–187. [Google Scholar] [CrossRef]
- Johnson, A.R.; Banks, M.L.; Selley, D.E.; Negus, S.S. Amphetamine maintenance differentially modulates effects of cocaine, methylenedioxypyrovalerone (MDPV), and methamphetamine on intracranial self-stimulation and nucleus accumbens dopamine in rats. Neuropsychopharmacology 2018, 43, 1753–1762. [Google Scholar] [CrossRef]
- Zwartsen, A.; Olijhoek, M.E.; Westerink, R.H.S.; Hondebrink, L. Hazard Characterization of Synthetic Cathinones Using Viability, Monoamine Reuptake, and Neuronal Activity Assays. Front. Neurosci. 2020, 14, 9. [Google Scholar] [CrossRef]
- Bourne, J.A. Intracerebral microdialysis: 30 years as a tool for the neuroscientist. Clin. Exp. Pharmacol. Physiol. 2003, 30, 16–24. [Google Scholar] [CrossRef]
- Proctor, C.M.; Chan, C.Y.; Porcarelli, L.; Udabe, E.; Sanchez-Sanchez, A.; Del Agua, I.; Mecerreyes, D.; Malliaras, G.G. Ionic Hydrogel for Accelerated Dopamine Delivery via Retrodialysis. Chem. Mater. Publ. Am. Chem. Soc. 2019, 31, 7080–7084. [Google Scholar] [CrossRef]
- Matto, V.; Terasmaa, A.; Vasar, E.; Koks, S. Impaired striatal dopamine output of homozygous Wfs1 mutant mice in response to [K+] challenge. J. Physiol. Biochem. 2011, 67, 53–60. [Google Scholar] [CrossRef]
- Baumann, K.; Falkencrone, S.; Knudsen, N.P.H.; Woetmann, A.; Dabelsteen, S.; Skov, P.S. The skin reservoir model: A tool for evaluating microdialysis sampling of large biomarkers from human skin. Acta Derm.-Venereol. 2020, 100, adv00008. [Google Scholar] [CrossRef]
- Cheng, F.C.; Kuo, J.S.; Huang, H.M.; Yang, D.Y.; Wu, T.F.; Tsai, T.H. Determination of catecholamines in pheochromocytoma cell (PC-12) culture medium by microdialysis-microbore liquid chromatography. J. Chromatogr. A 2000, 870, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: San Diego, CA, USA, 2007. [Google Scholar]
- Grecco, G.G.; Kisor, D.F.; Magura, J.S.; Sprague, J.E. Impact of common clandestine structural modifications on synthetic cathinone “bath salt” pharmacokinetics. Toxicol. Appl. Pharmacol. 2017, 328, 18–24. [Google Scholar] [CrossRef]
- Hitchcock, S.A.; Pennington, L.D. Structure-brain exposure relationships. J. Med. Chem. 2006, 49, 7559–7583. [Google Scholar] [CrossRef] [PubMed]
- Elmore, J.S.; Dillon-Carter, O.; Partilla, J.S.; Ellefsen, K.N.; Concheiro, M.; Suzuki, M.; Rice, K.C.; Huestis, M.A.; Baumann, M.H. Pharmacokinetic Profiles and Pharmacodynamic Effects for Methylone and Its Metabolites in Rats. Neuropsychopharmacology 2016, 42, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Bonano, J.S.; Glennon, R.A.; De Felice, L.J.; Banks, M.L.; Negus, S.S. Abuse-related and abuse-limiting effects of methcathinone and the synthetic “bath salts” cathinone analogs methylenedioxypyrovalerone (MDPV), methylone and mephedrone on intracranial self-stimulation in rats. Psychopharmacology 2013, 231, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Ferry, B.; Parrot, S.; Marien, M.; Lazarus, C.; Cassel, J.C.; McGaugh, J.L. Noradrenergic influences in the basolateral amygdala on inhibitory avoidance memory are mediated by an action on α2-adrenoceptors. Psychoneuroendocrinology 2015, 51, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Kozai, T.D.Y.; Jaquins-Gerstl, A.S.; Vazquez, A.L.; Michael, A.C.; Cui, X.T. Dexamethasone retrodialysis attenuates microglial response to implanted probes in vivo. Biomaterials 2016, 87, 157–169. [Google Scholar] [CrossRef]
- Nesbitt, K.M.; Varner, E.L.; Jaquins-Gerstl, A.; Michael, A.C. Microdialysis in the Rat Striatum: Effects of 24 h Dexamethasone Retrodialysis on Evoked Dopamine Release and Penetration Injury. ACS Chem. Neurosci. 2015, 6, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-DeRose, J.; Stauber, G.; Khroyan, T.V.; Xie, X.S.; Zaveri, N.T.; Toll, L. Retrodialysis of N/OFQ into the nucleus accumbens shell blocks cocaine-induced increases in extracellular dopamine and locomotor activity. Eur. J. Pharmacol. 2013, 699, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Hanberg, P.; Bue, M.; Öbrink-Hansen, K.; Kabel, J.; Thomassen, M.; Tøttrup, M.; Søballe, K.; Stilling, M. Simultaneous Retrodialysis by Drug for Cefuroxime Using Meropenem as an Internal Standard-A Microdialysis Validation Study. J. Pharm. Sci. 2020, 109, 1373–1379. [Google Scholar] [CrossRef]
- Yu, Y.; Chandasana, H.; Sangari, T.; Seubert, C.; Derendorf, H. Simultaneous Retrodialysis by Calibrator for Rapid In Vivo Recovery Determination in Target Site Microdialysis. J. Pharm. Sci. 2018, 107, 2259–2265. [Google Scholar] [CrossRef]
- Can, A.; Zanos, P.; Moaddel, R.; Kang, H.J.; Dossou, K.S.S.; Wainer, I.W.; Cheer, J.F.; Frost, D.O.; Huang, X.P.; Gould, T.D. Effects of ketamine and ketamine metabolites on evoked striatal dopamine release, dopamine receptors, and monoamine transporters. J. Pharmacol. Exp. Ther. 2016, 359, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Aburahma, A. Phenylethylamine Derivatives: Pharmacological and Toxicological Studies. Ph.D. Thesis, Bowling Green State University, Bowling Green, OH, USA, 2021. Available online: http://rave.ohiolink.edu/etdc/view?acc_num=bgsu1636041464110327 (accessed on 27 February 2024).



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldsmith, R.; Aburahma, A.; Sprague, J.E. Dopamine Concentration Changes Associated with the Retrodialysis of Methylone and 3,4-Methylenedioxypyrovalerone (MDPV) into the Caudate Putamen. Brain Sci. 2024, 14, 265. https://doi.org/10.3390/brainsci14030265
Goldsmith R, Aburahma A, Sprague JE. Dopamine Concentration Changes Associated with the Retrodialysis of Methylone and 3,4-Methylenedioxypyrovalerone (MDPV) into the Caudate Putamen. Brain Sciences. 2024; 14(3):265. https://doi.org/10.3390/brainsci14030265
Chicago/Turabian StyleGoldsmith, Robert, Amal Aburahma, and Jon E. Sprague. 2024. "Dopamine Concentration Changes Associated with the Retrodialysis of Methylone and 3,4-Methylenedioxypyrovalerone (MDPV) into the Caudate Putamen" Brain Sciences 14, no. 3: 265. https://doi.org/10.3390/brainsci14030265
APA StyleGoldsmith, R., Aburahma, A., & Sprague, J. E. (2024). Dopamine Concentration Changes Associated with the Retrodialysis of Methylone and 3,4-Methylenedioxypyrovalerone (MDPV) into the Caudate Putamen. Brain Sciences, 14(3), 265. https://doi.org/10.3390/brainsci14030265