



Bilingual Spatial Cognition: Spatial Cue Use in Bilinguals and Monolinguals


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
Language Background
2.2. Materials
2.2.1. Spatial Questionnaires
Santa Barbara Sense of Direction (SBSOD) Questionnaire
Wayfinding Strategy Questionnaire
Spatial Anxiety Scale
2.2.2. Mental Rotation Test (MRT)
2.2.3. Eight-Arm Task [43]
2.2.4. Flanker Task
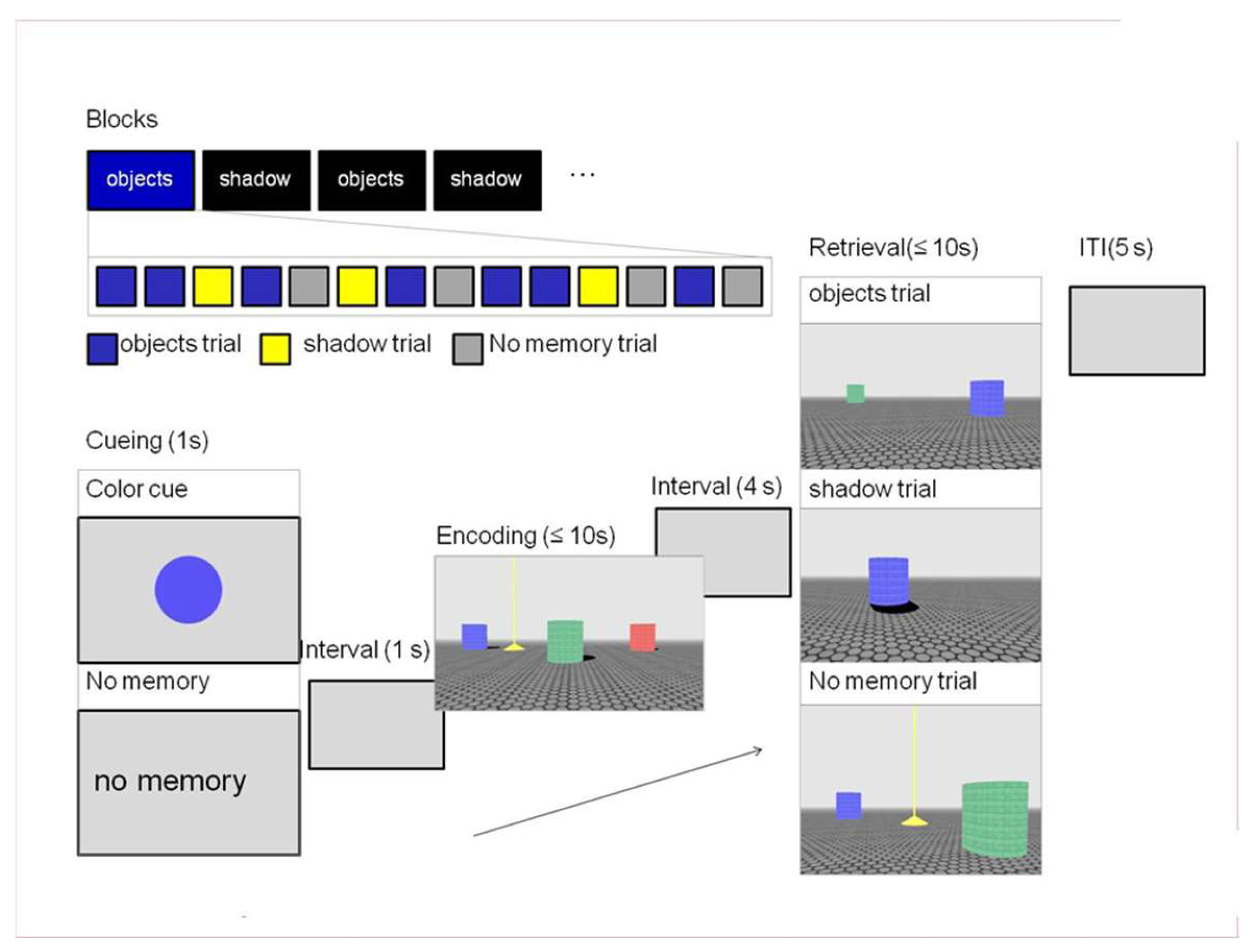
2.2.5. Objects in Space (OiS) Navigation Task (fMRI Task)
2.3. Procedure
2.3.1. Behavioral Session I
2.3.2. Scanning Session
2.3.3. Behavioral Session II
2.4. Functional MRI Data Acquisition and Analysis
3. Results
3.1. Behavioral Experiment
3.1.1. Spatial Questionnaires and Tasks
3.1.2. Flanker Task
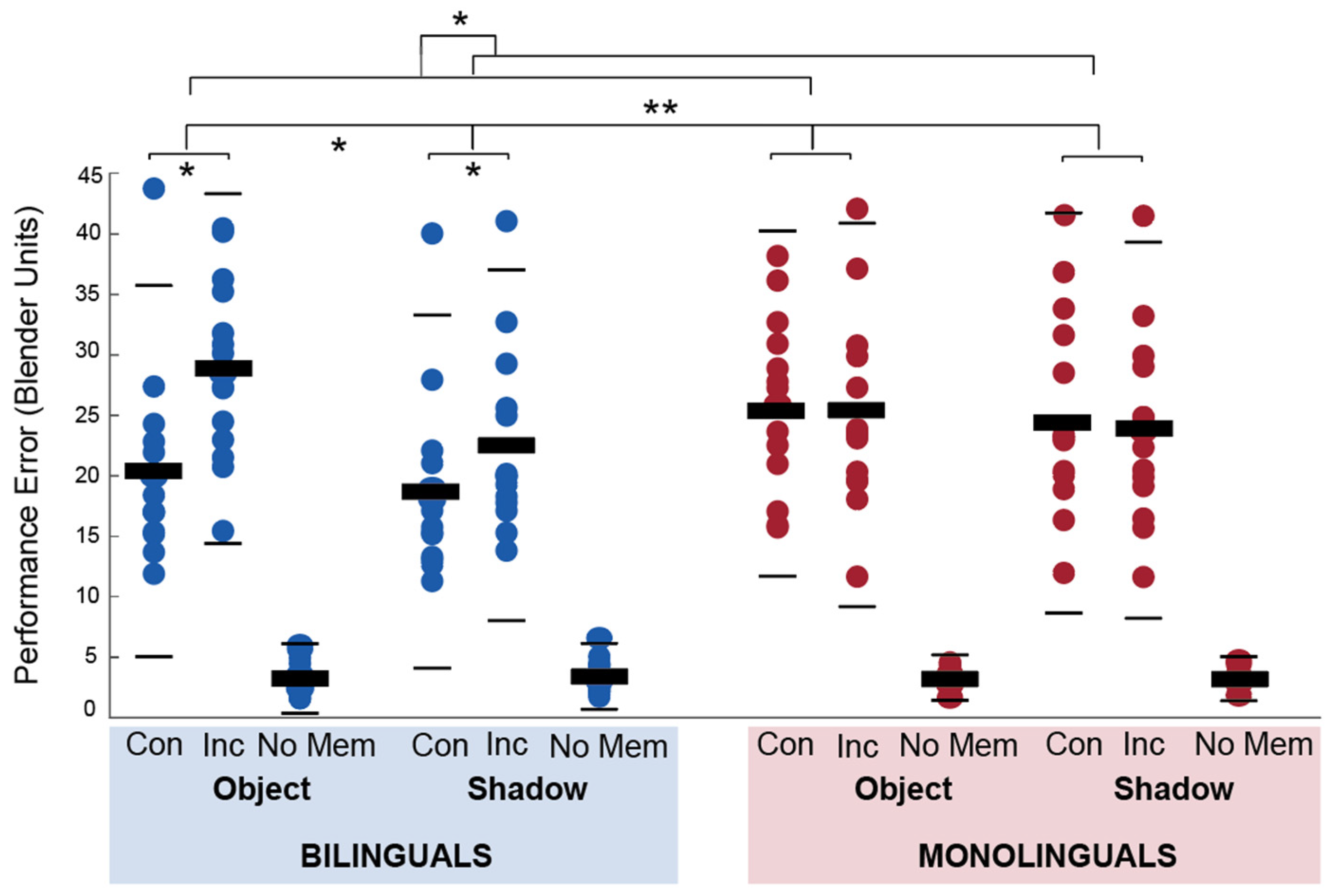
3.1.3. Objects in Space Task
Training
Main Task (fMRI)
3.2. Functional MRI Experiment
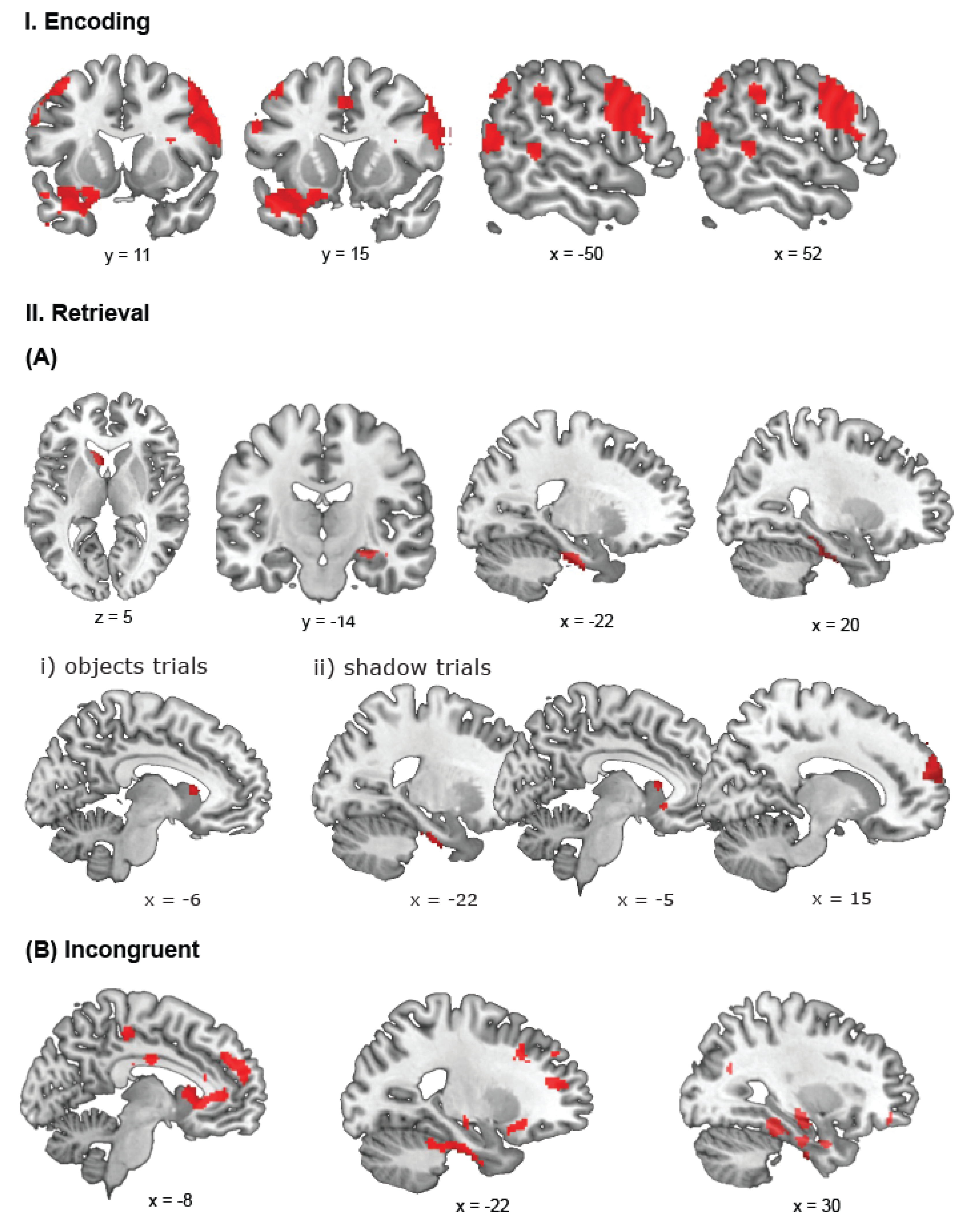
3.2.1. Encoding
3.2.2. Retrieval—Congruent Trials
3.2.3. Retrieval—Incongruent Trials
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bialystok, E. The bilingual adaptation: How minds accommodate experience. Psychol. Bull. 2017, 143, 233–262. [Google Scholar] [CrossRef]
- Carter, F.; DeLuca, V.; Segaert, K.; Mazaheri, A.; Krottet, A. Functional neural architecture of cognitive control mediates the relationship between individual differences in bilingual experience and behaviour. NeuroImage 2023, 273, 120085. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Bak, T.H.; Sorace, A.; Vega-Mendoza, M. Interference suppression in bilingualism: Stimulus-Stimulus vs. Stimulus-Response conflict. Biling. Lang. Cogn. 2022, 25, 256–268. [Google Scholar] [CrossRef]
- Paap, K.R. The bilingual advantage debate: Quantity and quality of the evidence. In The Handbook of Neuroscience of Multilingualism; Schwieter, J.W., Ed.; Wiley-Blackwell: London, UK, 2019; pp. 701–735. [Google Scholar]
- Paap, K.R.; Myuz, H.; Anders-Jefferson, R.; Mason, L.; Zimiga, B. On the ambiguity regarding the relationship between sequential congruency effects, bilingual advantages in cognitive control, and the disengagement of attention. AIMS Neurosci. 2019, 6, 282. [Google Scholar] [CrossRef] [PubMed]
- Paap, K.R.; Mason, L.A.; Zimiga, B.M.; Ayala-Silva, Y.; Frost, M.M.; Gonzalez, M.; Primero, L. Other Language Proficiency Predicts Unique Variance in Verbal Fluency Not Accounted for Directly by Target Language Proficiency: Cross-Language Interference? Brain Sci. 2019, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Paap, K.R.; Anders-Jefferson, R.; Mikulinsky, R.; Masuda, S.; Mason, L. On the encapsulation of bilingual language control. J. Mem. Lang. 2019, 105, 76–92. [Google Scholar] [CrossRef]
- Paap, K.R.; Myuz, H.A.; Anders, R.T.; Bockelman, M.F.; Mikulinsky, R.; Sawi, O.M. No compelling evidence for a bilingual advantage in switching or that frequent language switching reduces switch cost. J. Cogn. Psychol. 2016, 29, 89–112. [Google Scholar] [CrossRef]
- Grosjean, F. Studying Bilinguals; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Green, D.W. Mental control of the bilingual lexico-semantic system. Biling. Lang. Cogn. 1998, 1, 67–81. [Google Scholar] [CrossRef]
- Bialystok, E.; Craik, F.I.M. Cognitive and linguistic processing in the bilingual mind. Curr. Dir. Psychol. Sci. 2010, 19, 19–23. [Google Scholar] [CrossRef]
- Bialystok, E.; Craik, F.I.M.; Grady, C.; Chau, W.; Ishii, R.; Gunji, A.; Pantev, C. Effect of bilingualism on cognitive control in the Simon task: Evidence from MEG. NeuroImage 2005, 24, 40–49. [Google Scholar] [CrossRef]
- Costa, A.; Hernandez, M.; Sebastian-Galles, N. Bilingualism aids conflict resolution: Evidence from the ANT task. Cognition 2008, 106, 59–86. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Hernández, M.; Costa-Faidella, J.; Sebastián-Gallés, N. On the bilingual advantage in conflict processing: Now you see it, now you don’t. Cognition 2009, 113, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Prior, A.; Macwhinney, B. Bilingual advantage in task switching. Biling. Lang. Cogn. 2010, 13, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Krizman, J.; Marian, V.; Shook, A.; Skoe, E.; Kraus, N. Subcortical encoding of sound is enhanced in bilinguals and relates to executive function advantages. Proc. Natl. Acad. Sci. USA 2012, 109, 7877–7881. [Google Scholar] [CrossRef]
- Kovelman, I.; Baker, S.A.; Petitto, L.-A. Bilingual and monolingual brains compared: A Functional magnetic resonance imaging investigation of syntactic processing and a possible “neural signature” of bilingualism. J. Cogn. Neurosci. 2008, 20, 153–169. [Google Scholar] [CrossRef]
- Liu, H.; Hu, Z.; Guo, T.; Peng, D. Speaking words in two languages with one brain: Neural overlap and dissociation. Brain Res. 2010, 1316, 75–82. [Google Scholar] [CrossRef]
- Bakker-Marshall, I.; Takashima, A.; Fernandez, C.B.; Janzen, G.; McQueen, J.M.; Van Hell, J.G. Overlapping and distinct neural networks supporting novel word learning in bilinguals and monolinguals. Biling. Lang. Cogn. 2021, 24, 524–536. [Google Scholar] [CrossRef]
- Abutalebi, J.; Green, D.W. Control mechanisms in bilingual language production: Neural evidence from language switching studies. Lang. Cogn. Process. 2008, 23, 557–582. [Google Scholar] [CrossRef]
- Posner, M.I.; Sheese, B.E.; Odludas, Y.; Tang, Y. Analyzing and shaping human attentional networks. Neural Netw. 2006, 19, 1422–1429. [Google Scholar] [CrossRef]
- Hedden, T.; Gabrieli, J.D.E. Shared and selective neural correlates of inhibition, facilitation, and shifting processes during executive control. NeuroImage 2010, 51, 421–431. [Google Scholar] [CrossRef]
- Mechelli, A.; Crinion, J.T.; Noppeney, U.; O’Doherty, J.; Ashburner, J.; Franckowiak, R.S.; Price, C.J. Structural plasticity in the bilingual brain. Nature 2004, 431, 757. [Google Scholar] [CrossRef]
- Luk, G.; Bialystok, E.; Craik, F.I.M.; Grady, C.L. Lifelong bilingualism maintains white mater integrity in older adults. J. Neurosci. 2011, 31, 16808–16813. [Google Scholar] [CrossRef]
- Abutalebi, J.; Rosa, P.A.; Green, D.W.; Hernandez, M.; Scifo, P.; Keim, R.; Cappa, S.F.; Costa, A. Bilingualism tunes the anterior cingulated cortex for conflict monitoring. Cereb. Cortex 2012, 22, 2076–2086. [Google Scholar] [CrossRef]
- Luk, G.; Anderson, J.A.E.; Craik, F.I.M.; Grady, C.; Bialystok, E. Distinct neural correlates for two types of inhibition in bilinguals: Response inhibition versus interference suppression. Brain Cogn. 2010, 74, 347–357. [Google Scholar] [CrossRef]
- Garbin, G.; Sanjuan, A.; Forn, C.; Bustamante, J.C.; Rodriguez-Pujadas, A.; Belloch, V.; Hernandez, M.; Costa, A.; Avila, C. Bridging language and attention: Brain basis of the impact of bilingualism on cognitive control. NeuroImage 2010, 53, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Ransdell, S.E.; Fischler, I. Imagery skill and preference in bilinguals. App. Cogn. Psychol. 1991, 5, 97–112. [Google Scholar] [CrossRef]
- McLeay, H. The relationship between bilingualism and the performance of spatial tasks. Int. J. Biling. Educ. Biling. 2003, 6, 423–438. [Google Scholar] [CrossRef]
- Gonzalez-Marquez, M. Language, thought, and … brain? In Cambridge Handbook of Psycholinguistics; Cambridge University: Cambridge, UK, 2012; pp. 674–692. [Google Scholar]
- Morales, J.; Calvo, A.; Bialystok, E. Working memory development in monolingual and bilingual children. J. Exp. Child Psychol. 2013, 114, 187–202. [Google Scholar] [CrossRef]
- Kerrigan, L.; Thomas, M.; Bright, P.; Filippi, R. Evidence of an advantage in visuo-spatial memory for bilingual compared to monolingual speakers. Biling. Lang. Cogn. 2017, 20, 602–612. [Google Scholar] [CrossRef]
- Lawton, C.A. Strategies for indoor wayfinding: The role of orientation. J. Environ. Psychol. 1996, 16, 137–145. [Google Scholar] [CrossRef]
- Kato, Y.; Takeuchi, Y. Individual differences in wayfinding strategies. J. Environ. Psychol. 2003, 23, 171–188. [Google Scholar] [CrossRef]
- Meneghetti, C.; Pazzaglia, F.; De Beni, R. Spatial mental representations derived from survey and route descriptions: When individuals prefer extrinsic frame of reference. Learn. Individ. Differ. 2011, 21, 150–157. [Google Scholar] [CrossRef]
- Zhong, J.Y.; Kozhevnikov, M. Relating allocentric and egocentric survey-based representations to the self-reported use of a navigation strategy of egocentric spatial updating. J. Environ. Psychol. 2016, 46, 154–175. [Google Scholar] [CrossRef]
- Burles, F.; Liu, I.; Hart, C.; Murias, K.; Graham, S.A.; Iaria, G. The Emergence of Cognitive Maps for Spatial Navigation in 7- to 10-Year-Old Children. Child Dev. 2019, 91, e733–e744. [Google Scholar] [CrossRef]
- Ruotolo, F.; Ruggiero, G.; Raemaekers, M.; Iachini, T.; van der Ham, I.J.M.; Fracasso, A.; Postma, A. Neural correlates of egocentric and allocentric frames of reference combined with metric and non-metric spatial relations. J. Neurosci. 2019, 409, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Bocchi, A.; Palermo, L.; Boccia, M.; Palmiero, M.; D’Amico, S.; Piccardi, L. Object recognition and location: Which compo-nent of object location memory for landmarks is affected by gender? Evidence from four to ten year-old children. Appl. Neuropsychol. Child 2020, 9, 31–40. [Google Scholar] [CrossRef]
- Van Hoogmoed, A.H.; Wegman, J.; van den Brink, D.; Janzen, G. Development of Landmark Use for Navigation in Children: Effects of Age, Sex, Working Memory and Landmark Type. Brain Sci. 2022, 12, 776. [Google Scholar] [CrossRef]
- Burgess, N. Spatial memory: How egocentric and allocentric combine. Trends Cogn. Sci. 2006, 10, 551–557. [Google Scholar] [CrossRef]
- Shelton, A.L.; Gabrieli, J.D.E. Neural correlates of encoding space from route and survey perspectives. J. Neurosci. 2002, 22, 2711–2717. [Google Scholar] [CrossRef]
- Iaria, G.; Petrides, M.; Dagher, A.; Pike, B.; Bohbot, V.D. Cognitive strategies dependent on the hippocampus and caudate nucleus in human navigation: Variability and change with practice. J. Neurosci. 2003, 23, 5945–5952. [Google Scholar] [CrossRef]
- Baumann, O.; Chan, E.; Mattingley, J.B. Dissociable neural circuits for encoding and retrieval of object locations during active navigation in humans. NeuroImage 2010, 49, 2816–2825. [Google Scholar] [CrossRef] [PubMed]
- Murias, K.; Slone, E.; Tariq, S.; Iaria, G. Development of spatial orientation skills: An fMRI study. Brain Imaging Behav. 2019, 13, 1590–1601. [Google Scholar] [CrossRef] [PubMed]
- Tansan, M.; Nguyen, K.V.; Newcombe, N.S. Spatial Navigation in Childhood and Aging. Annu. Rev. Dev. Psychol. 2022, 4, 253–272. [Google Scholar] [CrossRef]
- Spiers, H.; Maguire, E. A navigational guidance system in the human brain. Hippocampus 2007, 17, 618–626. [Google Scholar] [CrossRef]
- Janzen, G.; van Turennout, M. Selective neural representation of objects relevant for navigation. Nat. Neurosci. 2004, 7, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Janzen, G.; Wagensveld, B.; van Turennout, M. Neural representation of navigational relevance is rapidly induced and long-lasting. Cereb. Cortex 2007, 17, 975–981. [Google Scholar] [CrossRef]
- Janzen, G.; Jansen, C.; van Turennout, M. Memory consolidation of landmarks in good navigators. Hippocampus 2008, 18, 40–47. [Google Scholar] [CrossRef]
- Wegman, J.; Janzen, G. The neural processing of spatial information during learning and rest. J. Cogn. Neurosci. 2011, 23, 3841–3854. [Google Scholar] [CrossRef][Green Version]
- Bohbot, V.D.; Lerch, J.; Thorndycraft, B.; Iaria, G.; Zijdenbos, A.P. Gray matter differences correlate with spontaneous strategies in a human virtual navigation task. J. Neurosci. 2007, 27, 10078–10083. [Google Scholar] [CrossRef] [PubMed]
- Iaria, G.; Lanyon, L.J.; Fox, C.J.; Giaschi, D.; Barton, J.J. Navigational skills correlate with hippocampal fractional anisotropy in humans. Hippocampus 2008, 18, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Doeller, C.; King, J.A.; Burgess, N. Parallel striatal and hippocampal systems for landmarks and boundaries in spatial memory. Proc. Natl. Acad. Sci. USA 2008, 105, 5915–5920. [Google Scholar] [CrossRef]
- Igloi, K.; Doeller, C.F.; Berthoz, A.; Rondi-Reig, L.; Burgess, N. Lateralized human hippocampal activity predicts navigation based on sequence or place memory. Proc. Natl. Acad. Sci. USA 2010, 107, 14466–14471. [Google Scholar] [CrossRef] [PubMed]
- Iaria, G.; Chen, J.-K.; Guariglia, C.; Ptito, A.; Petrides, M. Retrosplenial and hippocampal brain regions in human navigation: Complementary functional contributions to the formation and use of cognitive maps. Eur. J. Neurosci. 2007, 25, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Seubert, J.; Humphreys, G.W.; Muller, H.J.; Klaus, G. Straight after the turn: The role of the parietal lobes in egocentric space processing. Neurocase: Neural Basis Cogn. 2008, 14, 204–219. [Google Scholar] [CrossRef]
- Ino, T.; Inoue, Y.; Kage, M.; Hirose, S.; Kimura, T.; Fukuyama, H. Mental navigation in humans is processed in the anterior bank of the parietal-occipital sulcus. Neurosci. Lett. 2002, 322, 182–186. [Google Scholar] [CrossRef]
- Baumann, O.; Mattingley, J.B. Medial parietal cortex encodes perceived heading direction in humans. J. Neurosci. 2010, 30, 12897–12901. [Google Scholar] [CrossRef]
- Sack, A.T. Parietal cortex and spatial cognition. Behav. Brain Res. 2009, 202, 153–161. [Google Scholar] [CrossRef]
- Miyake, A.; Friedman, N.P.; Rettinger, D.A.; Shah, P.; Hegarty, M. How are visuospatial working memory, executive functioning, and spatial abilities related? A latent-variable analysis. J. Exp. Psychol. 2001, 130, 621–640. [Google Scholar] [CrossRef] [PubMed]
- Moffat, S.D.; Kennedy, K.M.; Rodrigue, K.M.; Raz, N. Extrahippocampal contributions to age differences in human spatial navigation. Cereb. Cortex 2007, 17, 1274–1282. [Google Scholar] [CrossRef]
- Ciaramelli, E. The role of ventromedial prefrontal cortex in navigation: A case of impaired wayfinding and rehabilitation. Neuropsychologia 2008, 46, 2099–2105. [Google Scholar] [CrossRef]
- Li, P.; Sepanski, S.; Zhao, X. Language history questionnaire: A web-based interface for bilingual research. Behav. Res. Methods 2006, 38, 202–210. [Google Scholar] [CrossRef]
- Council of Europe. A Common European Framework of Reference for Languages: Learning, Teaching, Assessment; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Hegarty, M.; Montello, D.; Richardson, A.; Ishikawa, T.; Lovelace, K. Spatial abilities at different scales: Individual differences in aptitude-test performance and spatial-layout learning. Intelligence 2006, 34, 151–176. [Google Scholar] [CrossRef]
- Hegarty, M.; Richardson, A.; Montello, D.; Lovelace, K.; Subbiah, I. Development of a self-report measure of environmental spatial ability. Intelligence 2002, 30, 425–447. [Google Scholar] [CrossRef]
- Ishikawa, T.; Montello, D.R. Spatial knowledge acquisition from direct experience in the environment: Individual differences in the development of metric knowledge and the integration of separately learned places. Cogn. Psychol. 2006, 52, 93–129. [Google Scholar] [CrossRef]
- Lawton, C.A. Gender differences in way-finding strategies: Relationship to spatial ability and spatial anxiety. Sex Roles 1994, 30, 765–779. [Google Scholar] [CrossRef]
- Hund, A.M.; Minarik, L.M. Getting from here to there: Spatial anxiety, wayfinding strategies, direction type, and wayfinding efficiency. Spat. Cogn. Comput. 2006, 6, 179–201. [Google Scholar] [CrossRef]
- Vandenberg, S.G.; Kuse, A.R. Mental rotations, a group test of three-dimensional spatial visualization. Percept. Mot. Ski. 1978, 47, 599–601. [Google Scholar] [CrossRef]
- Shepard, R.; Metzler, J. Mental rotation of three dimensional objects. Science 1971, 171, 701–703. [Google Scholar] [CrossRef]
- Morris, R.G.M. Spatial localization does not require the presence of local cues. Learn. Motiv. 1981, 12, 239–260. [Google Scholar] [CrossRef]
- Wegman, J.; Tyborowska, A.; Janzen, G. Encoding and retrieval of landmark-related spatial cues during navigation: An fMRI study. Hippocampus 2014, 24, 853–868. [Google Scholar] [CrossRef]
- Poser, B.A.; Versluis, M.J.; Hoogduin, J.M.; Norris, D.G. BOLD contrast sensitivity enhancement and artifact reduction with multiecho EPI: Parallel-acquired inhomogeneity desensitized fMRI. Magn. Reson. Med. 2006, 55, 1227–1235. [Google Scholar] [CrossRef]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyar, B.; Joliot, M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Emmorey, K.; Luk, G.; Pyers, J.E.; Bialystok, E. The source of enhanced cognitive control in bilinguals: Evidence from bimodal bilinguals. Psychol. Sci. 2008, 19, 1201–1206. [Google Scholar] [CrossRef]
- Lehtonen, M.; Soveri, A.; Laine, A.; Järvenpää, J.; de Bruin, A.; Antfolk, J. Is bilingualism associated with enhanced exec-utive functioning in adults? A meta-analytic review. Psychol. Bull. 2018, 144, 394–425. [Google Scholar] [CrossRef]
- Nichols, E.S.; Wild, C.J.; Stojanoski, B.; Battista, M.E.; Owen, A.M. Bilingualism Affords No General Cognitive Advantages: A Population Study of Executive Function in 11,000 People. Psychol. Sci. 2020, 31, 548–567. [Google Scholar] [CrossRef] [PubMed]
- Grundy, J.G. The effects of bilingualism on executive functions: An updated quantitative analysis. J. Cult. Cogn. Sci. 2020, 4, 177–199. [Google Scholar] [CrossRef]
- Hilchey, M.D.; Klein, R.M. Are there bilingual advantages on nonlinguistic interference tasks? Implications for the plasticity of executive control processes. Psychon. Bull. Rev. 2011, 18, 625–658. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Federspiel, A.; Koenig, T.; Wirth, M.; Strik, W.; Wiest, R.; Dierks, T. Structural plasticity in the language system related to increased second language proficiency. Cortex 2012, 48, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Seghier, M.L. The angular gyrus: Multiple functions and multiple subdivisions. Neuroscientist 2012, 19, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Voermans, N.C.; Petersson, K.M.; Daudey, L.; Weber, B.; Van Spaendonck, K.P.; Kremer, H.P.H.; Fernandez, G. Interaction between the human hippocampus and the caudate nucleus during route recognition. Neuron 2004, 43, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Crinion, J.; Turner, R.; Grogan, A.; Hanakawa, T.; Noppeney, U.; Devlin, J.T.; Price, C.J. Language control in the bilingual brain. Science 2006, 312, 1537–1540. [Google Scholar] [CrossRef]
- Bush, G.; Luu, P.; Posner, M.I. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn. Sci. 2000, 4, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Rosselli, M.; Loewenstein, D.A.; Curiel, R.E.; Penate, A.; Torres, V.L.; Lang, M.; Greig, M.T.; Barker, W.W.; Duara, R. Effects of Bilingualism on Verbal and Nonverbal Memory Measures in Mild Cognitive Impairment. J. Int. Neuropsychol. Soc. 2019, 25, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Naeem, K.; Filippi, R.; Periche-Tomas, E.; Papageorgiou, A.; Bright, P. The Importance of Socioeconomic Status as a Modu-lator of the Bilingual Advantage in Cognitive Ability. Front. Psychol. 2018, 9, 1818. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyborowska, A.; Wegman, J.; Janzen, G. Bilingual Spatial Cognition: Spatial Cue Use in Bilinguals and Monolinguals. Brain Sci. 2024, 14, 134. https://doi.org/10.3390/brainsci14020134
Tyborowska A, Wegman J, Janzen G. Bilingual Spatial Cognition: Spatial Cue Use in Bilinguals and Monolinguals. Brain Sciences. 2024; 14(2):134. https://doi.org/10.3390/brainsci14020134
Chicago/Turabian StyleTyborowska, Anna, Joost Wegman, and Gabriele Janzen. 2024. "Bilingual Spatial Cognition: Spatial Cue Use in Bilinguals and Monolinguals" Brain Sciences 14, no. 2: 134. https://doi.org/10.3390/brainsci14020134
APA StyleTyborowska, A., Wegman, J., & Janzen, G. (2024). Bilingual Spatial Cognition: Spatial Cue Use in Bilinguals and Monolinguals. Brain Sciences, 14(2), 134. https://doi.org/10.3390/brainsci14020134