1. Introduction
Advanced age often comes with a decline in sensorimotor control and functioning that affects the ability to perform activities of daily living. Indeed, it has been shown that movements become slower and/or less accurate and more cognition-dependent as we age [
1]. Motor declines overall significantly impact motor independence, which is essential for older adults’ quality of life and interactions with their environment. Additionally, behavioral evidence indicates that aging is frequently linked to slower movements, reduced capacity for learning new motor skills, and diminished ability to adjust a movement plan after initiation [
2]. In fact, Mild Parkinsonian Symptoms (
MPSs), such as rigidity, bradykinesia, and tremor, are commonly diagnosed during clinical examination of older adults who do not have a diagnosed neurological disease [
3].
Advances in neuroimaging techniques have made contributions to a better understanding of the aging brain. For instance, aging impacts brain structure, leading to a decrease in gray and white matter volume, along with an increase in cerebrospinal fluid in ventricles, fissures, and sulci [
4,
5]. These aging-related processes affect almost the entire cortex and underlying white matter, with a steeper decline in the primary motor cortex (
M1) and frontal subcortical white matter [
6]. For instance, aging is also associated with a complex pattern of atrophy [
7], demyelination [
8], free tissue water, and iron reduction within somatosensory and motor areas [
6]. Moreover, age-related atrophy of motor cortical regions and the corpus callosum has been shown to coincide with motor declines such as balance gait deficits and coordination deficits [
9].
Sensorimotor cortex oscillations measured by electroencephalography (
EEG) in the beta band (13–30 Hz) are a predominant feature of movement production and have been shown to be generated by local field potentials within the motor cortex [
10]. Beta oscillations exhibit a robust pattern of movement-related changes, such as pre-movement beta Event-Related Desynchronization (
ERD), Movement-Related Beta Desynchronization (
MRBD), and post-movement beta Event-Related Synchronization (
ERS) or beta rebound [
11,
12]. In terms of how these oscillations relate to motor performance, an association between MRBD and the accuracy with which subjects performed a bimanual task has been demonstrated, where subjects with more negative MRBD values exhibited worse task performance [
13]. Greater MRBDs were also shown to correlate with a longer movement duration to complete a finger-tapping sequence [
14]. In older adults, a greater (i.e. more negative) MRBD in both motor and premotor areas has been observed in subjects performing cued finger button presses [
15,
16] and handgrip tasks [
13]. Additionally, older age has been associated with greater baseline beta power (15–29 Hz) at rest [
17], suggesting that the alterations in brain structure and biochemistry during aging could be the reason behind the observed altered neural activation patterns. Given the association between movement production and beta band features, there is a high interest in modulating these oscillations non-invasively to improve motor ability and performance in older adults.
Transcranial alternating current stimulation (
tACS) is a non-invasive brain stimulation (
NIBS) technique that can alter oscillatory brain rhythms through synchronization of neural networks in a frequency-dependent manner [
18]. This method is believed to entrain endogenous brain oscillations through the synchronization of two oscillatory systems that occurs when a driving external oscillatory force coordinates with another oscillating system [
19,
20,
21,
22]. The effects of tACS depend on key parameters: stimulation location, intensity, and frequency [
23]. In terms of location, the acquisition of motor skills is linked to a number of cortical and subcortical brain regions, but among these, M1 is thought to play a central role [
24,
25,
26,
27], making it a popular target for neurostimulation. Regarding intensity, it is generally set between 1 mA and 2 mA because it is well tolerated and it has been shown to modulate cortex excitability and alter cognitive function [
28]. Additionally, using higher intensities raise concerns about safety and side effects [
29]. Regarding frequency, motor cortex activity during movement predominantly oscillates at 20 Hz (beta band) [
30] and 70 Hz (gamma band) [
31]. Beta band activity within the motor system has been linked to an antikinetic role, as it is associated with slower voluntary movements in both healthy individuals [
21,
32,
33] and those with motor disorders [
34]. A reason for this may be that, in the cortex–basal ganglia circuit, beta activity is associated with promoting tonic rather than voluntary movement [
35,
36]. Also, motor impairments in Parkinson’s disease (
PD) have also been linked to elevated beta band activity in the motor cortex and subthalamic nucleus [
37]. In contrast, gamma band activity is thought to be prokinetic, as it increases in the basal ganglia–cortical motor circuit during voluntary movement [
38]. Behaviorally, the use of 20 Hz tACS has been shown to slow voluntary movement, while 70 Hz tACS enhances motor learning along with an increase in beta power [
32]. However, these results involved participants in the younger range (32.7 ± 6.8 years) and the influence of these tACS frequencies on aging-related brain neural activity has not yet been studied.
Other NIBS techniques, such as transcranial direct current stimulation (
tDCS), have been shown to have greater effects on motor performance when applied during a motor task, a technique often called
online stimulation, compared to before the motor task [
39]. There is also evidence that applying tDCS during practice triggers effects that outlast the stimulation period and facilitate neuroplasticity [
28]. Previous studies have reported mixed results regarding the effects of NIBS on young adults, and these results cannot be easily transferred to older adults. The stimulation sites and frequencies that modulate brain oscillatory activity in young adults may not result in the same effect in older adults, and functional reorganization of the aging brain may be an explanation [
40].
In recent years, standard double-electrode tACS has shown limitations in controlling the stimulation focus and intensity. The use of different electrode montages, such as High-Definition tACS (
HD-tACS), has allowed more precise stimulation control. Prior research has indicated that HD-tACS yields a more pronounced focalization of its effects through multiple smaller electrodes, possibly resulting from reduced distribution of the electrical field compared to conventional tACS [
41,
42]. Notably, online HD-tACS, applied during a motor task, induces phase- and frequency-dependent effects on cortical excitability [
43,
44].
After-effects on brain oscillations are a common outcome following tACS [
45]. For instance, 10 Hz tACS stimulation of the parieto-occipital area resulted in an enhancement of the EEG-recorded alpha amplitude during the stimulation and this effect was seen to last at least 30 min after a 10 min stimulation period [
46,
47]. Other NIBS techniques, such as tDCS, have induced long-lasting excitability elevations in the human motor cortex [
48], and in animals, a stimulation period of 5 to 30 min causes an effect lasting for hours after the end of stimulation [
49].
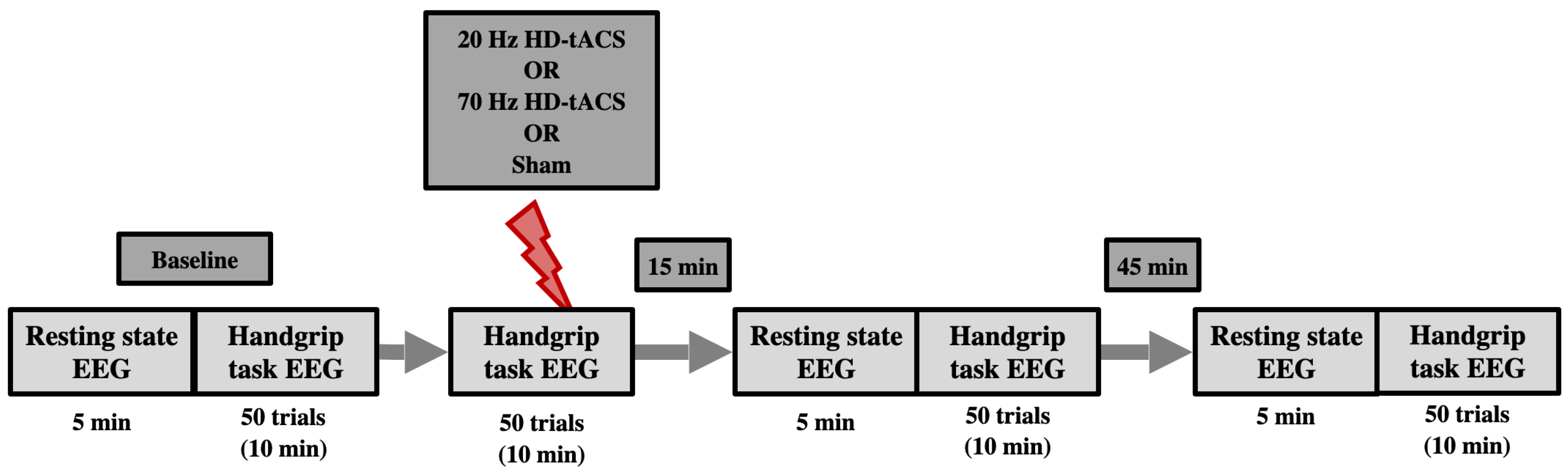
This study aims to explore the after-effects of 70 Hz and 20 Hz HD-tACS on beta brain oscillatory patterns in healthy older adults. Based on the previously mentioned effect of tACS on motor performance on younger cohorts and how their beta brain oscillations differ from older ones, we hypothesized that 70 Hz HD-tACS would decrease resting-state beta power and promote a more positive MRBD (lower desynchronization). Conversely, we hypothesized that 20 Hz HD-tACS would increase beta power at rest and induce more negative MRBD (higher desynchronization).
4. Discussion
This study aimed to quantitatively examine the after-effects of HD-tACS on electrophysiological features in healthy older adults. Our rmANOVA anaysis for beta power revealed a noteworthy main effect of Stimulation for C3, as well as a significant effect of Time for CP3. Conversely, no other electrodes displayed any statistically significant main effects. Upon post hoc analysis of t-tests, a notable reduction in resting-state beta power was observed post-45 min for 20 Hz HD-tACS for both electrodes. In contrast, in the case of 70 Hz HD-tACS and Sham, no significant changes were observed in either electrode.
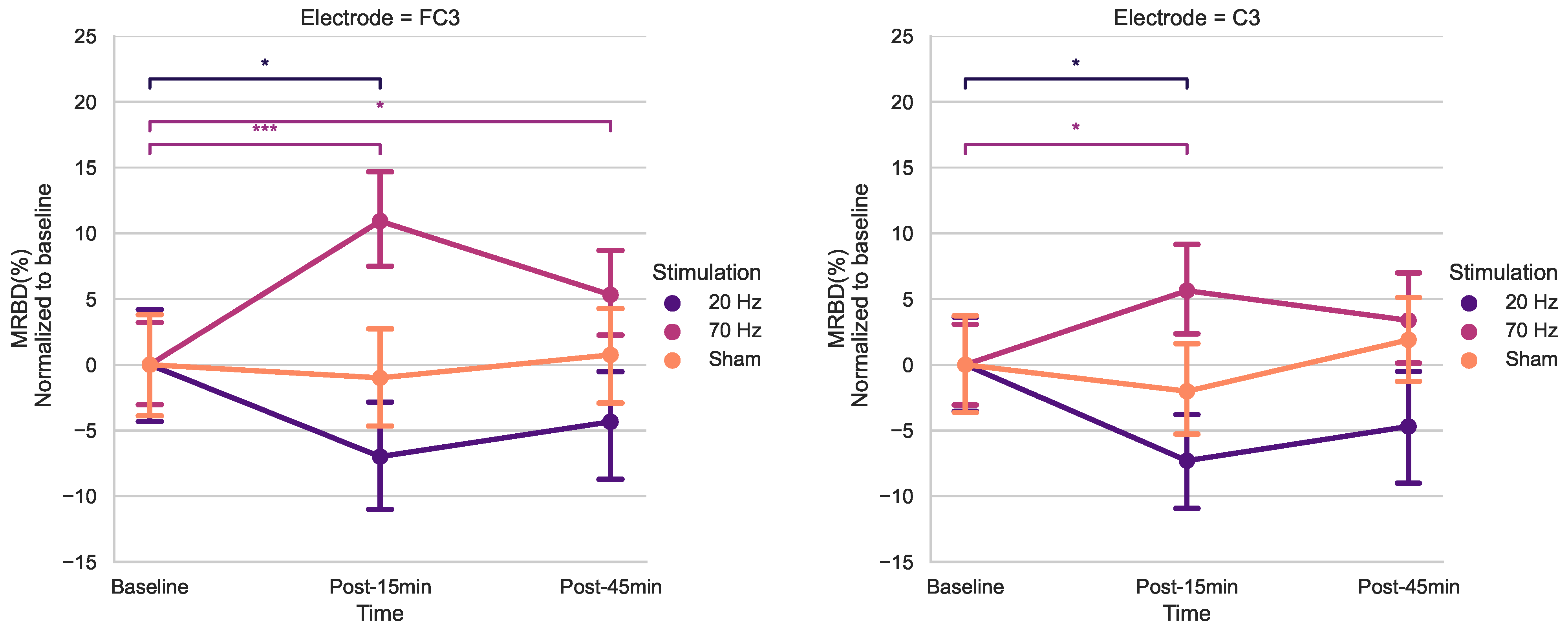
When analyzing MRBD effects, there was a significant Time × Stimulation interaction for FC3 and C3. The results of the post hoc t-tests showed higher MRBD values (more negative) 15 min after 20 Hz HD-tACS in both electrodes. Applying 70 Hz HD-tACS resulted in significant reductions in MRBD values after 15 min in FC3 and C3 and after 45 min only in FC3. Notably, no significant changes were observed during Sham stimulation. In the subsequent subsections, the implications of these results on beta oscillatory patterns are discussed in greater detail.
4.1. Population Behavioral Scores and Baseline Features
To ensure the representativeness of our participants concerning motor performance, we employed the Desrosiers et al. [
59] model to predict the PPT score for each participant based on their age when performing the motor tests during the eligibility session. The PPT quantifies fingertip dexterity and gross movement of the hand, fingers, and arm. The predicted and actual scores of the PPT were similar in all the subtests, which suggests that the motor performance of our participants fell within the range of normal scores for the older population.
4.2. Modulation of Resting-State Beta Power
After administering 20 Hz HD-tACS, a delayed reduction in beta power was observed for electrodes C3 and CP3. This modulation appeared only 45 min post-stimulation, with no change detected at 15 min, suggesting that the impact on beta power is gradual rather than immediate. This finding aligns with previous studies showing no after-effects on beta power within 5 to 20 min after 20 Hz tACS [
63]. However, prior research demonstrated that tACS could have effects on endogenous EEG power in the range of the stimulation frequency up to 70 min after the stimulation [
64], which guided our decision to monitor changes up to 45 min post-stimulation.
The delayed response may reflect plasticity mechanisms, long-lasting modifications in neural connections and activity, with dynamic brain changes emerging over time. These results are consistent with reports of late plasticity changes in corticospinal excitability following 20 Hz tACS [
65]. A relationship between beta oscillations and corticospinal excitability has been observed, with increased spontaneous beta oscillatory activity linked to smaller motor-evoked potential (
MEP) amplitudes [
66]. Furthermore, beta tACS has been shown to elevate cortical excitability in the M1 during stimulation, as evidenced by increased MEP amplitudes [
67,
68]. Heise et al. [
41] also reported that more focal HD stimulation is significantly more effective in modulating MEPs post-stimulation. Additionally, delayed effects induced by tACS may be attributed to spike-timing-dependent plasticity (
STDP), a process where the precise timing of neural activity determines whether synaptic connections are strengthened or weakened [
69]. According to STDP principles, synapses in circuits resonating at frequencies similar to repetitive inputs are strengthened during stimulation. After stimulation ends, these synaptic modifications persist, leading to enhanced neural activity at the circuits’ resonant frequencies. This aligns with findings that beta tACS can sustain elevated beta oscillations and cortical excitability for at least an hour post-stimulation [
64,
65,
70]. Currently, evidence on STDP and its specific manifestations in aging populations is limited. However, studies have shown that synaptic plasticity mechanisms, including STDP, may be less efficient in older adults, potentially due to age-related changes in synaptic connectivity, neurotransmitter levels, and cortical excitability [
71]. This could influence the extent and timing of tACS-induced plasticity effects. While our findings do not directly investigate these mechanisms, they provide a basis for future research to explore how aging might modulate tACS effects through STDP-related processes.
Our results disagree with recent research showing that 20 Hz HD-tACS increased beta power following stimulation of the visual [
72] and parietal cortexes [
73]. Also, a study reported no significant effects when applying tACS at 20 Hz, also in M1 during rest, on the beta power in younger participants using the standard double-electrode tACS montage [
74]. We can identify three possible reasons why our results differ from the aforementioned studies. First, the difference in age of our participants is a possible reason, as it has been shown before that older groups demonstrated a decrease in tACS-induced neuroplasticity compared to a younger cohort [
75]. We chose an older population, since these HD-tACS frequencies have not been studied before in aging, and we specifically examined its effects on beta oscillations. Secondly, in our study, HD-tACS was applied while participants were performing a handgrip task (online) as opposed to HD-tACS during rest (offline). Differences have been observed across studies comparing tACS-induced changes in online and offline protocols [
46]. Additionally, a recent study comparing online and offline HD-tDCS showed that only the online stimulation reduced the power of the alpha rhythm during motor skill execution [
76]. Thirdly, we used a different electrode montage and it has been shown that an HD-tACS electrode montage delivers a more focal current to M1 than the standard double-electrode tACS montage [
77,
78]. It is important to note that beta-tACS has been shown to have mixed results on other outcomes, such as corticospinal excitability and motor function [
79]. Therefore, more research should focus on applying HD-tACS at this frequency, while keeping similar parameters to the ones used in studies showing significant effects.
For 70 Hz, there were no significant changes in resting beta power, which does not align with our initial hypothesis. This null finding underscores a critical limitation in current HD-tACS research: the application of fixed-frequency stimulation across diverse neural populations. Sugata et al. [
32] found frequency-specific increases in beta power, while our results are more consistent with Mastakouri et al. [
80], who demonstrated that gamma-tACS effects are heterogeneous and subject-specific. The recent literature increasingly emphasizes the importance of individualized stimulation protocols. Multiple studies [
69,
81,
82,
83,
84] have shown that tACS delivered at a subject-specific frequency can produce more robust effects on cortical oscillations compared to fixed-frequency approaches. Moreover, age-related neuroplasticity differences may further complicate our findings. Guerra et al. [
85] demonstrated that gamma stimulation mechanisms differ between younger and older adults, with older participants showing enhanced motor skill consolidation despite potentially reduced long-term potentiation (LTP)-like plasticity.
One promising solution emerges in the form of closed-loop brain–computer interfaces (BCIs). As highlighted by Xu et al., these systems provide a dynamic approach to neural modulation by continuously monitoring outcomes and adaptively modifying stimulation parameters [
86]. By identifying specific neural biomarkers in real time, such as oscillatory patterns in gamma or beta frequency ranges, closed-loop systems can adjust stimulation accordingly. This approach addresses the current limitations of fixed-frequency stimulation by enabling real-time adaptation to an individual’s unique neural dynamics. These nuanced findings suggest that future research should achieve the following:
Develop individualized stimulation protocols based on baseline neural characteristics;
Consider age-related neuroplasticity differences;
Employ a more comprehensive assessment of stimulation effects;
Explore closed-loop systems for real-time neural modulation.
Our null findings for 70 Hz stimulation should not be interpreted as a complete absence of effect, but rather as an invitation to develop more sophisticated, personalized neuromodulation approaches [
87].
4.3. Modulation of MRBD
We found that after 15 min of 20 Hz HD-tACS, MRBD values became more negative in FC3 and C3. Conversely, after 15 min of 70 Hz HD-tACS, MRBD values became more positive in both FC3 and C3, and the effect persisted after 45 min only in FC3. These results align closely with our initial hypothesis. To the best of our knowledge, there are no other studies investigating the specific effects of HD-tACS on MRBD values in older adults. A recent study demonstrated that tACS at 10 Hz enhanced MRBD during a motor imagery task compared to pseudo-stimulation, indicating the capability of tACS to modify movement-related brain oscillations [
88]. Additionally, other forms of NIBS, such as tDCS, have been shown to induce more negative MRBDs during motor imagery after 15 min of stimulation.
Xifra-Porxas et al. [
13] previously mentioned that the motor performance decline observed in healthy aging may not be due to an impairment in the capacity to modulate beta oscillations. In fact, they observed a larger modulation in older compared to younger adults. On the other hand, beta oscillations at rest are greater in older adults [
17], which suggests that increased desynchronization is needed to reach a threshold to initiate a movement. This would mean that modulating this desynchronization could later translate into a change in motor performance. While our study primarily focused on brain oscillations as a potential mechanism of HD-tACS-induced effects, we also evaluated motor task performance through measures of accuracy and reaction time. However, no significant changes were observed, potentially due to the simplicity of the motor task employed. Future research aiming to validate this hypothesis should utilize more complex motor tasks, such as the sequential visual isometric pinch task (
SVIPT), which better resembles the level of difficulty encountered in everyday life skills [
89]. Tasks like the SVIPT have been shown to challenge motor learning and performance to a degree comparable to real-world activities [
90], making them ideal for studying the interplay between MRBD, beta modulation, and motor performance.
Finally, the fact that electrode C3, the anode of our HD-tACS montage, showed significant changes in both resting-state beta power and MRBD values suggests a focalization of the current right on the electrode that delivers the current. This focalization possibly results from a reduced distribution of the electrical field compared to the conventional tACS montage, which utilizes two distant patch electrodes [
41].
Taken together, the results of this study are partly aligned with our initial hypothesis. In terms of beta power, only 20 Hz HD-tACS showed a decrease in beta power, but only 45 min after the end of stimulation, while 70 Hz HD-tACS did not show any significant changes, contrary to our hypothesis that it would decrease beta power. Regarding MRBD, both 20 Hz and 70 Hz showed the expected results: the former resulted in a more negative MRBD and the latter in a more positive. However, these changes in beta oscillations may not necessarily translate to improvements in motor performance. We do not report effects on motor tasks, since the complexity of our task is not enough to show impacts. Future research should focus on assessing the impact of HD-tACS on motor performance using tasks that are more complex and reflective of real-world daily activities, particularly for older adults.
There are certain limitations in our study that may explain some of variability in our HD-tACS outcomes and previously reported outcomes, such as interindividual differences, including skull thickness and the actual amount of current that reaches the cortex [
91,
92]. Because of these differences, individualized stimulation frequencies and current amplitudes, validated by studies such as Yamaguchi et al. [
93], emphasize the parameter-dependent nature of tACS effects. In addition, our limited stimulation duration (10 min) contrasts with longer-lasting effects seen in extended gamma-tACS in mice, suggesting prolonged sessions or multiple-day approaches akin to tDCS studies for in-depth exploration [
49,
94]. Finally, the analysis of ongoing brain signals during concurrent stimulation is of primary interest and would also shed light on the immediate effects and mechanisms of HD-tACS. Recent advances in artifact removal algorithms will enable this type of analysis in our data [
95].

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}