
Development, Insults and Predisposing Factors of the Brain’s Predictive Coding System to Chronic Perceptual Disorders—A Life-Course Examination


Abstract
1. Introduction
2. Development of the Predictive Coding System through the Life Course
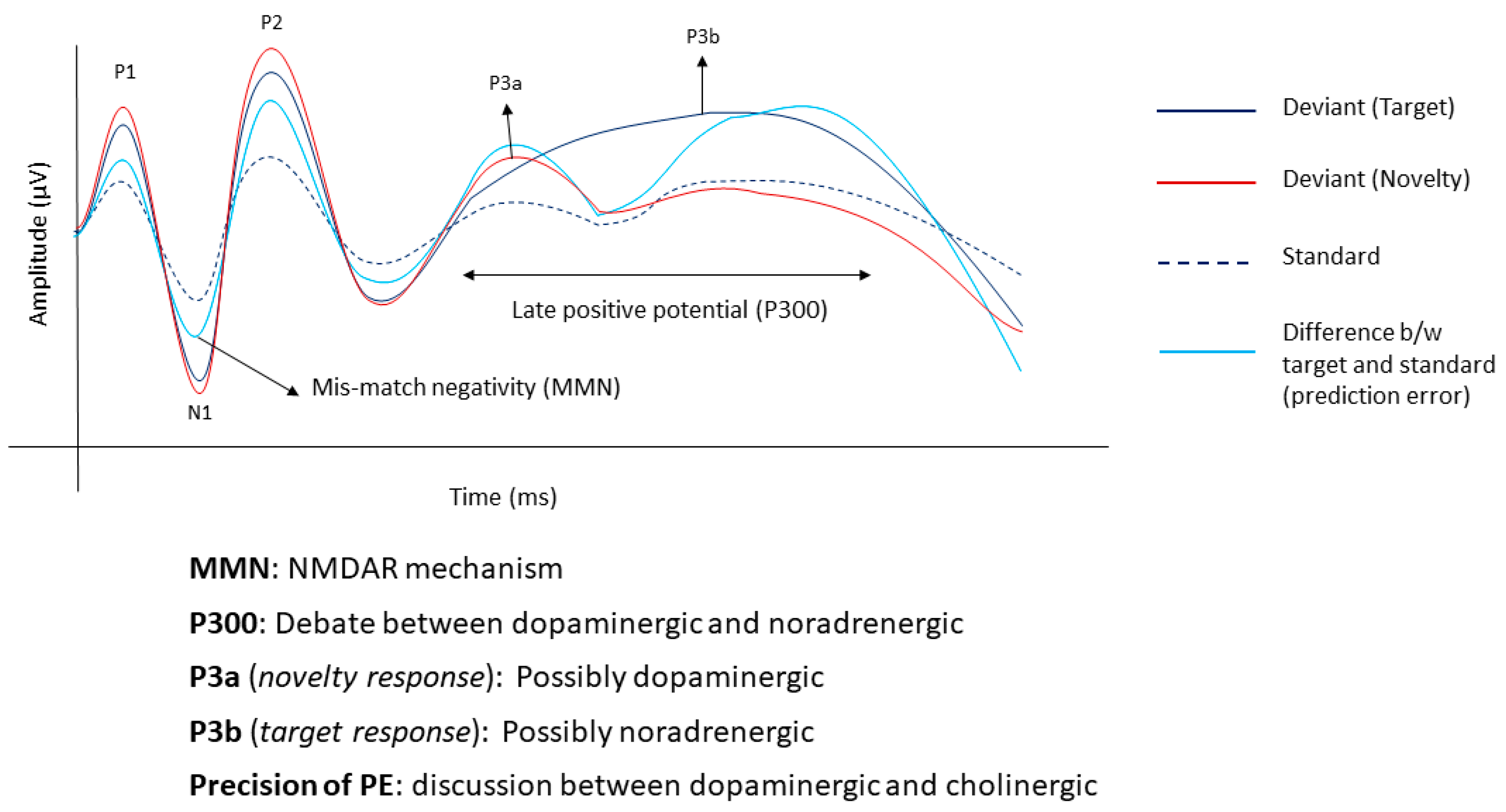
2.1. Neural Markers
2.1.1. Mismatch Negativity
2.1.2. Late Positive Potential
2.2. Molecular Markers
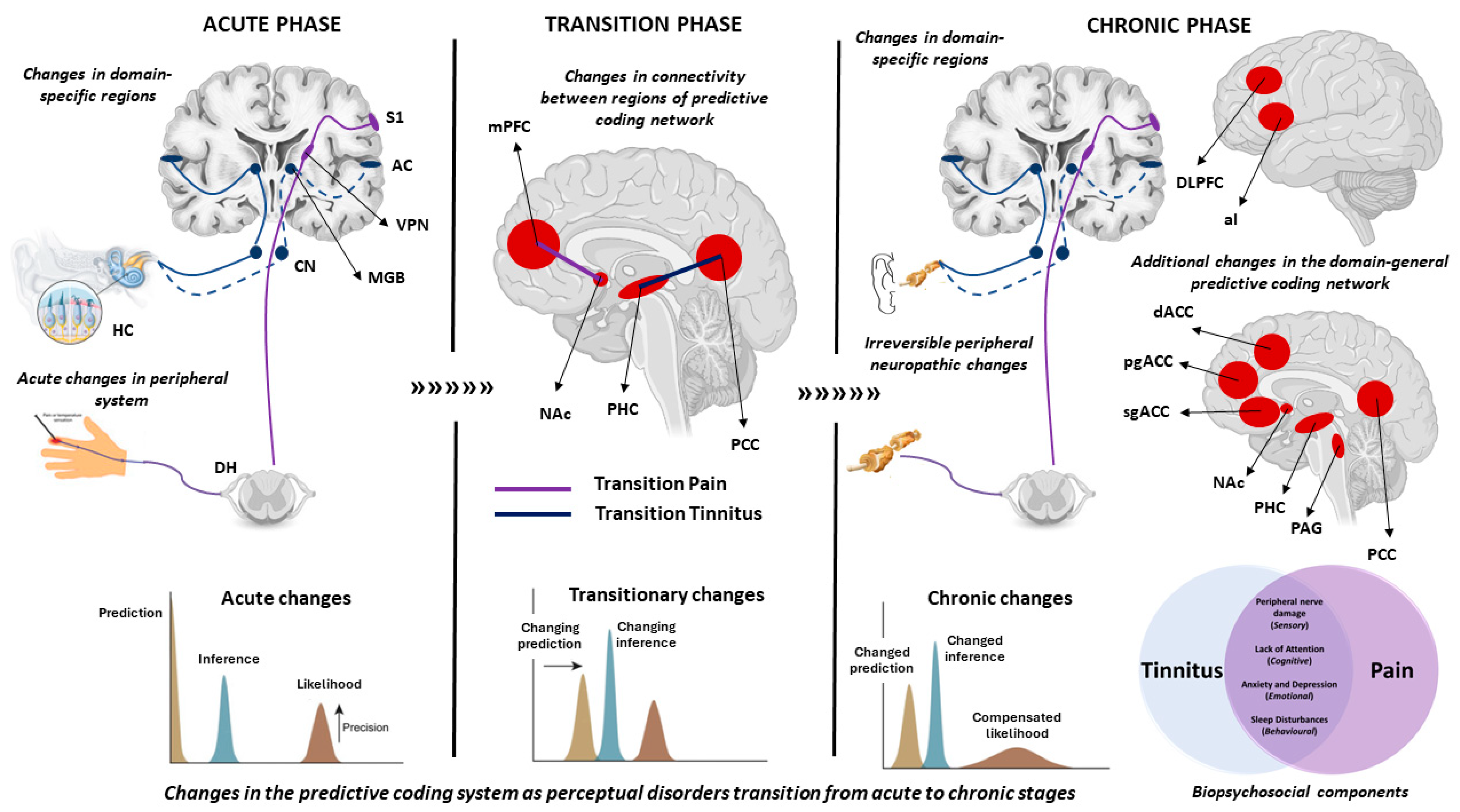
3. Changes in Predictive Coding in the Acute Phase of Phantom Perceptions
3.1. Neural Markers
3.2. Molecular Markers
4. Changes in Predictive Coding in the Chronic Phase of Phantom Perceptions
4.1. Psychosocial Components
4.2. Neural Markers
4.3. Molecular Markers
5. Changes in Predictive Coding when Transitioning from Acute to Chronic Phase
5.1. Psychosocial Components
5.2. Neural Markers
5.3. Molecular Markers
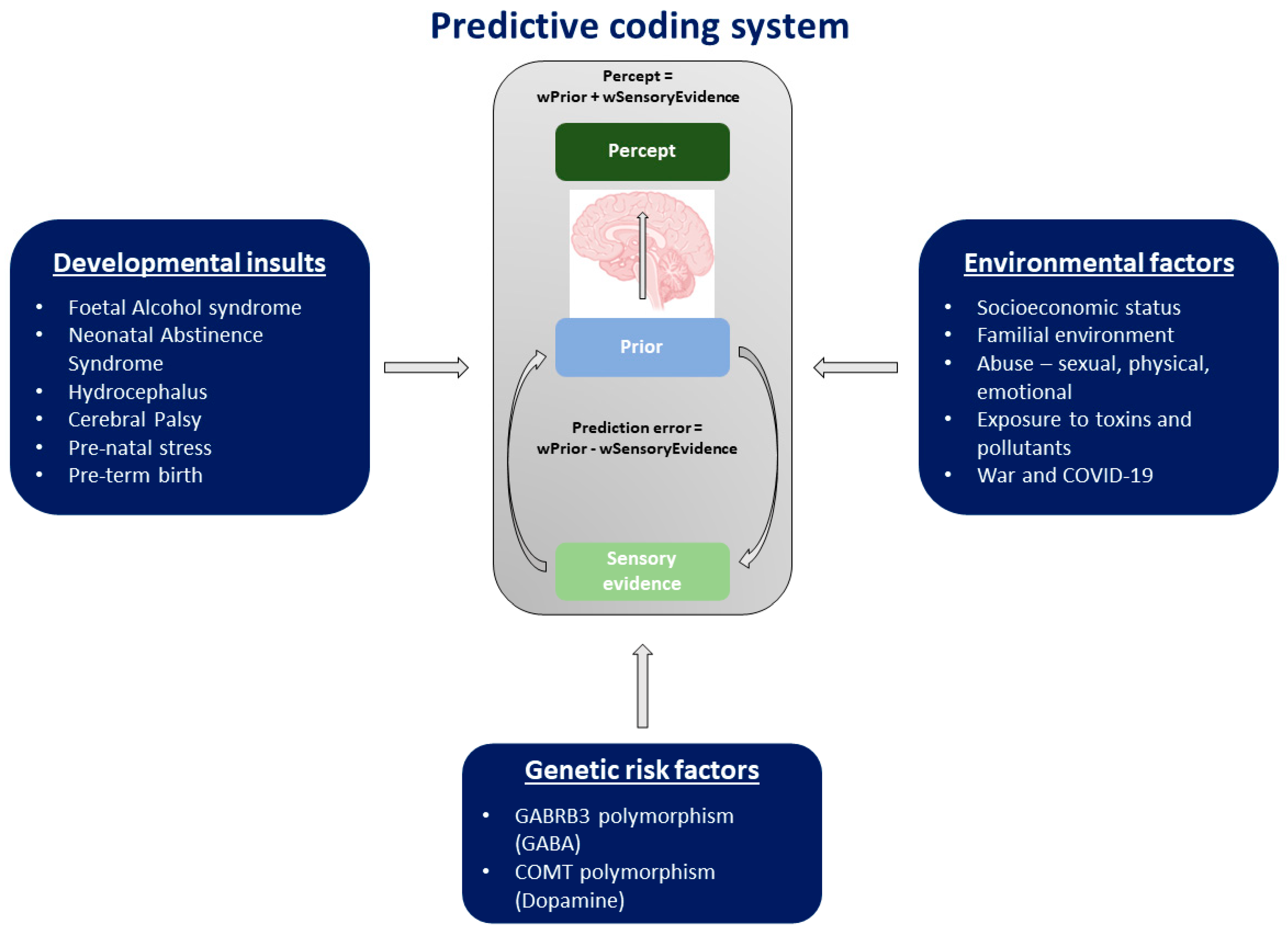
6. Factors Predisposing Children and Adults through the Life-Course to Maladaptive Changes to Predictive Coding and Perceptual Disorders
6.1. Developmental Insults
6.2. Environmental Factors
6.3. Genetic Risk Factors
7. Limitations
8. Future Directions
9. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knill, D.C.; Pouget, A. The Bayesian brain: The role of uncertainty in neural coding and computation. Trends Neurosci. 2004, 27, 712–719. [Google Scholar] [CrossRef]
- Friston, K.; FitzGerald, T.; Rigoli, F.; Schwartenbeck, P.; Doherty, J.O.; Pezzulo, G. Active inference and learning. Neurosci. Biobehav. Rev. 2016, 68, 862–879. [Google Scholar] [CrossRef]
- Powers, A.R.; Mathys, C.; Corlett, P.R. Pavlovian conditioning–induced hallucinations result from overweighting of perceptual priors. Science 2017, 357, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Bhamoo, N.; Riquelme, J.S.; Long, S.; Norena, A.; Vanneste, S. Investigating functional changes in the brain to intermittently induced auditory illusions and its relevance to chronic tinnitus. Hum. Brain Mapp. 2020, 41, 1819–1832. [Google Scholar] [CrossRef]
- Ryan, A.F.; Kujawa, S.G.; Hammill, T.; Le Prell, C.; Kil, J. Temporary and Permanent Noise-induced Threshold Shifts: A Review of Basic and Clinical Observations. Otol. Neurotol. 2016, 37, e271–e275. [Google Scholar] [CrossRef] [PubMed]
- Amaya, F.; Izumi, Y.; Matsuda, M.; Sasaki, M. Tissue injury and related mediators of pain exacerbation. Curr. Neuropharmacol. 2013, 11, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Vanneste, S. Adaptive and maladaptive neural compensatory consequences of sensory deprivation—From a phantom percept perspective. Prog. Neurobiol. 2017, 153, 1–17. [Google Scholar] [CrossRef]
- Corlett, P.R.; Horga, G.; Fletcher, P.C.; Alderson-Day, B.; Schmack, K.; Powers, A.R., III. Hallucinations and strong priors. Trends Cogn. Sci. 2019, 23, 114–127. [Google Scholar] [CrossRef]
- Sedley, W.; Friston, K.J.; Gander, P.E.; Kumar, S.; Griffiths, T.D. An Integrative Tinnitus Model Based on Sensory Precision. Trends Neurosci. 2016, 39, 799–812. [Google Scholar] [CrossRef]
- Rauschecker, J.P.; Leaver, A.M.; Mühlau, M. Tuning out the noise: Limbic-auditory interactions in tinnitus. Neuron 2010, 66, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Rauschecker, J.P.; May, E.S.; Maudoux, A.; Ploner, M. Frontostriatal Gating of Tinnitus and Chronic Pain. Trends Cogn. Sci. 2015, 19, 567–578. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Vanneste, S. Chapter 15—The Bayesian brain in imbalance: Medial, lateral and descending pathways in tinnitus and pain: A perspective. In Progress in Brain Research; Langguth, B., Kleinjung, T., De Ridder, D., Schlee, W., Vanneste, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 262, pp. 309–334. [Google Scholar]
- Vanneste, S.; Alsalman, O.; De Ridder, D. Top-down and Bottom-up Regulated Auditory Phantom Perception. J. Neurosci. 2019, 39, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Luckey, A.; Weisz, N.; Vanneste, S. Predisposition to domain-wide maladaptive changes in predictive coding in auditory phantom perception. NeuroImage 2021, 248, 118813. [Google Scholar] [CrossRef] [PubMed]
- Rubin, J.J. Psychosomatic pain: New insights and management strategies. South Med. J. 2005, 98, 1099–1111. [Google Scholar] [CrossRef]
- Jarach, C.M.; Lugo, A.; Scala, M.; van den Brandt, P.A.; Cederroth, C.R.; Odone, A.; Garavello, W.; Schlee, W.; Langguth, B.; Gallus, S. Global Prevalence and Incidence of Tinnitus: A Systematic Review and Meta-analysis. JAMA Neurol. 2022, 79, 888–900. [Google Scholar] [CrossRef]
- Dahlhamer, J.; Lucas, J.; Zelaya, C.; Nahin, R.; Mackey, S.; DeBar, L.; Kerns, R.; Von Korff, M.; Porter, L.; Helmick, C. Prevalence of chronic pain and high-impact chronic pain among adults—United States, 2016. Morb. Mortal. Wkly. Rep. 2018, 67, 1001. [Google Scholar] [CrossRef]
- Goldstein, E.; Ho, C.-X.; Hanna, R.; Elinger, C.; Yaremchuk, K.L.; Seidman, M.D.; Jesse, M.T. Cost of Care for Subjective Tinnitus in Relation to Patient Satisfaction. Otolaryngol. Head Neck Surg. 2015, 152, 518–523. [Google Scholar] [CrossRef]
- Steglitz, J.; Buscemi, J.; Ferguson, M.J. The future of pain research, education, and treatment: A summary of the IOM report “Relieving pain in America: A blueprint for transforming prevention, care, education, and research”. Transl. Behav. Med. 2012, 2, 6–8. [Google Scholar] [CrossRef]
- Nagai, Y. Predictive learning: Its key role in early cognitive development. Philos. Trans. R. Soc. B 2019, 374, 20180030. [Google Scholar] [CrossRef]
- Näätänen, R.; Paavilainen, P.; Rinne, T.; Alho, K. The mismatch negativity (MMN) in basic research of central auditory processing: A review. Clin. Neurophysiol. 2007, 118, 2544–2590. [Google Scholar] [CrossRef]
- Wacongne, C.; Labyt, E.; van Wassenhove, V.; Bekinschtein, T.; Naccache, L.; Dehaene, S. Evidence for a hierarchy of predictions and prediction errors in human cortex. Proc. Natl. Acad. Sci. USA 2011, 108, 20754–20759. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Hotson, L.; Trainor, L.J. Mismatch responses to pitch changes in early infancy. J. Cogn. Neurosci. 2007, 19, 878–892. [Google Scholar] [CrossRef]
- He, C.; Trainor, L.J. Finding the pitch of the missing fundamental in infants. J. Neurosci. 2009, 29, 7718–8822. [Google Scholar] [CrossRef] [PubMed]
- Trainor, L.J.; Samuel, S.S.; Desjardins, R.N.; Sonnadara, R.R. Measuring temporal resolution in infants using mismatch negativity. Neuroreport 2001, 12, 2443–2448. [Google Scholar] [CrossRef] [PubMed]
- Trainor, L.; McFadden, M.; Hodgson, L.; Darragh, L.; Barlow, J.; Matsos, L.; Sonnadara, R. Changes in auditory cortex and the development of mismatch negativity between 2 and 6 months of age. Int. J. Psychophysiol. 2003, 51, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Tew, S.; Fujioka, T.; He, C.; Trainor, L. Neural representation of transposed melody in infants at 6 months of age. Ann. N. Y. Acad. Sci. 2009, 1169, 287–290. [Google Scholar] [CrossRef]
- Trainor, L.J.; Lee, K.; Bosnyak, D.J. Cortical plasticity in 4-month-old infants: Specific effects of experience with musical timbres. Brain Topogr. 2011, 24, 192–203. [Google Scholar] [CrossRef]
- Wacongne, C.; Changeux, J.-P.; Dehaene, S. A neuronal model of predictive coding accounting for the mismatch negativity. J. Neurosci. 2012, 32, 3665–3678. [Google Scholar] [CrossRef]
- Chennu, S.; Noreika, V.; Gueorguiev, D.; Blenkmann, A.; Kochen, S.; Ibáñez, A.; Owen, A.M.; Bekinschtein, T.A. Expectation and attention in hierarchical auditory prediction. J. Neurosci. 2013, 33, 11194–11205. [Google Scholar] [CrossRef]
- Emberson, L.L.; Richards, J.E.; Aslin, R.N. Top-down modulation in the infant brain: Learning-induced expectations rapidly affect the sensory cortex at 6 months. Proc. Natl. Acad. Sci. USA 2015, 112, 9585–9590. [Google Scholar] [CrossRef]
- Kouider, S.; Long, B.; Le Stanc, L.; Charron, S.; Fievet, A.-C.; Barbosa, L.S.; Gelskov, S.V. Neural dynamics of prediction and surprise in infants. Nat. Commun. 2015, 6, 8537. [Google Scholar] [CrossRef] [PubMed]
- De Haan, M. Infant EEG and Event-Related Potentials; Psychology Press: London, UK, 2013. [Google Scholar]
- Elsner, B.; Jeschonek, S.; Pauen, S. Event-related potentials for 7-month-olds’ processing of animals and furniture items. Dev. Cogn. Neurosci. 2013, 3, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Marinović, V.; Hoehl, S.; Pauen, S. Neural correlates of human–animal distinction: An ERP-study on early categorical differentiation with 4-and 7-month-old infants and adults. Neuropsychologia 2014, 60, 60–76. [Google Scholar] [CrossRef] [PubMed]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed]
- Harms, L.; Parras, G.G.; Michie, P.T.; Malmierca, M.S. The Role of Glutamate Neurotransmission in Mismatch Negativity (MMN), A Measure of Auditory Synaptic Plasticity and Change-detection. Neuroscience 2021, 456, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Carbajal, G.V.; Malmierca, M.S. The neuronal basis of predictive coding along the auditory pathway: From the subcortical roots to cortical deviance detection. Trends Hear. 2018, 22, 2331216518784822. [Google Scholar] [CrossRef] [PubMed]
- Frodl-Bauch, T.; Bottlender, R.; Hegerl, U. Neurochemical substrates and neuroanatomical generators of the event-related P300. Neuropsychobiology 1999, 40, 86–94. [Google Scholar] [CrossRef]
- Albrecht, M.A.; Martin-Iverson, M.T.; Price, G.; Lee, J.; Iyyalol, R. Dexamphetamine-induced reduction of P3a and P3b in healthy participants. J. Psychopharmacol. 2011, 25, 1623–1631. [Google Scholar] [CrossRef]
- Vanneste, S.; Mohan, A.; Yoo, H.B.; Huang, Y.; Luckey, A.M.; McLeod, S.L.; Tabet, M.N.; Souza, R.R.; McIntyre, C.K.; Chapman, S. The peripheral effect of direct current stimulation on brain circuits involving memory. Sci. Adv. 2020, 6, eaax9538. [Google Scholar] [CrossRef]
- Sales, A.C.; Friston, K.J.; Jones, M.W.; Pickering, A.E.; Moran, R.J. Locus Coeruleus tracking of prediction errors optimises cognitive flexibility: An Active Inference model. PLoS Comput. Biol. 2019, 15, e1006267. [Google Scholar] [CrossRef]
- Murphy, P.R.; Robertson, I.H.; Balsters, J.H.; O’connell, R.G. Pupillometry and P3 index the locus coeruleus–noradrenergic arousal function in humans. Psychophysiology 2011, 48, 1532–1543. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, C.M.; Balsam, P.D.; Weinstein, J.J.; Rosengard, R.J.; Slifstein, M.; Daw, N.D.; Abi-Dargham, A.; Horga, G. A perceptual inference mechanism for hallucinations linked to striatal dopamine. Curr. Biol. 2018, 28, 503–514.e504. [Google Scholar] [CrossRef]
- Iglesias, S.; Mathys, C.; Brodersen, K.H.; Kasper, L.; Piccirelli, M.; den Ouden, H.E.; Stephan, K.E. Hierarchical prediction errors in midbrain and basal forebrain during sensory learning. Neuron 2013, 80, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, S.; Kasper, L.; Harrison, S.J.; Manka, R.; Mathys, C.; Stephan, K.E. Cholinergic and dopaminergic effects on prediction error and uncertainty responses during sensory associative learning. NeuroImage 2020, 226, 117590. [Google Scholar] [CrossRef] [PubMed]
- Moran, R.J.; Campo, P.; Symmonds, M.; Stephan, K.E.; Dolan, R.J.; Friston, K.J. Free energy, precision and learning: The role of cholinergic neuromodulation. J. Neurosci. 2013, 33, 8227–8236. [Google Scholar] [CrossRef]
- Conn, P.M. Sourcebook of Models for Biomedical Research; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Martucci, K.T.; Mackey, S.C. Imaging Pain. Anesth. Clin. 2016, 34, 255–269. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Vanneste, S.; Smith, M.; Adhia, D. Pain and the Triple Network Model. Front. Neurol. 2022, 13, 757241. [Google Scholar] [CrossRef] [PubMed]
- Leaver, A.M.; Renier, L.; Chevillet, M.A.; Morgan, S.; Kim, H.J.; Rauschecker, J.P. Dysregulation of limbic and auditory networks in tinnitus. Neuron 2011, 69, 33–43. [Google Scholar] [CrossRef]
- Sedley, W.; Alter, K.; Gander, P.E.; Berger, J.; Griffiths, T.D. Exposing Pathological Sensory Predictions in Tinnitus Using Auditory Intensity Deviant Evoked Responses. J. Neurosci. 2019, 39, 10096–10103. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Y.; Sun, Y.; Wang, J.Y. A Brain Signature to Differentiate Acute and Chronic Pain in Rats. Front. Comput. Neurosci. 2016, 10, 41. [Google Scholar] [CrossRef]
- Legrain, V.; Guérit, J.-M.; Bruyer, R.; Plaghki, L. Electrophysiological correlates of attentional orientation in humans to strong intensity deviant nociceptive stimuli, inside and outside the focus of spatial attention. Neurosci. Lett. 2003, 339, 107–110. [Google Scholar] [CrossRef]
- Leske, S.; Tse, A.; Oosterhof, N.N.; Hartmann, T.; Müller, N.; Keil, J.; Weisz, N. The strength of alpha and beta oscillations parametrically scale with the strength of an illusory auditory percept. NeuroImage 2014, 88, 69–78. [Google Scholar] [CrossRef]
- Norena, A.; Micheyl, C.; Chery-Croze, S. An auditory negative after-image as a human model of tinnitus. Hear. Res. 2000, 149, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and molecular mechanisms of pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Knipper, M.; Zimmermann, U.; Müller, M. Molecular aspects of tinnitus. Hear. Res. 2010, 266, 60–69. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Schlee, W.; Vanneste, S.; Londero, A.; Weisz, N.; Kleinjung, T.; Shekhawat, G.S.; Elgoyhen, A.B.; Song, J.J.; Andersson, G.; et al. Tinnitus and tinnitus disorder: Theoretical and operational definitions (an international multidisciplinary proposal). Prog. Brain Res. 2021, 260, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised International Association for the Study of Pain definition of pain: Concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef]
- Baguley, D.; McFerran, D.; Hall, D. Tinnitus. Lancet 2013, 382, 1600–1607. [Google Scholar] [CrossRef]
- Cohen, S.P.; Vase, L.; Hooten, W.M. Chronic pain: An update on burden, best practices, and new advances. Lancet 2021, 397, 2082–2097. [Google Scholar] [CrossRef]
- Hearing Loss Association of America. Hearing Loss Facts and Statistics; Hearing Loss Association of America: Bethesda, MD, USA, 2019. [Google Scholar]
- Treede, R.D.; Rief, W.; Barke, A.; Aziz, Q.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Evers, S.; Finnerup, N.B.; First, M.B.; et al. A classification of chronic pain for ICD-11. Pain 2015, 156, 1003–1007. [Google Scholar] [CrossRef]
- Volcheck, M.M.; Graham, S.M.; Fleming, K.C.; Mohabbat, A.B.; Luedtke, C.A. Central sensitization, chronic pain, and other symptoms: Better understanding, better management. Cleve. Clin. J. Med. 2023, 90, 245–254. [Google Scholar] [CrossRef] [PubMed]
- De Meulemeester, K.; Meeus, M.; De Pauw, R.; Cagnie, B.; Keppler, H.; Lenoir, D. Suffering from chronic tinnitus, chronic neck pain, or both: Does it impact the presence of signs and symptoms of central sensitization? PLoS ONE 2023, 18, e0290116. [Google Scholar] [CrossRef] [PubMed]
- Noreña, A.J. An integrative model of tinnitus based on a central gain controlling neural sensitivity. Neurosci. Biobehav. Rev. 2011, 35, 1089–1109. [Google Scholar] [CrossRef] [PubMed]
- Harte, S.E.; Harris, R.E.; Clauw, D.J. The neurobiology of central sensitization. J. Appl. Biobehav. Res. 2018, 23, e12137. [Google Scholar] [CrossRef]
- Zeng, F.G. Tinnitus and hyperacusis: Central noise, gain and variance. Curr. Opin. Physiol. 2020, 18, 123–129. [Google Scholar] [CrossRef]
- Latremoliere, A.; Woolf, C.J. Central sensitization: A generator of pain hypersensitivity by central neural plasticity. J. Pain 2009, 10, 895–926. [Google Scholar] [CrossRef]
- Leong, S.L.; Tchen, S.; Robertson, I.H.; Alsalman, O.; To, W.T.; Vanneste, S. The potential interruptive effect of tinnitus-related distress on attention. Sci. Rep. 2020, 10, 11911. [Google Scholar] [CrossRef]
- Attridge, N.; Keogh, E.; Eccleston, C. The effect of pain on task switching: Pain reduces accuracy and increases reaction times across multiple switching paradigms. Pain 2016, 157, 2179–2193. [Google Scholar] [CrossRef]
- Moore, D.J.; Meints, S.M.; Lazaridou, A.; Johnson, D.; Franceschelli, O.; Cornelius, M.; Schreiber, K.; Edwards, R.R. The Effect of Induced and Chronic Pain on Attention. J. Pain 2019, 20, 1353–1361. [Google Scholar] [CrossRef]
- Amaro-Díaz, L.; Montoro, C.I.; Fischer-Jbali, L.R.; Galvez-Sánchez, C.M. Chronic Pain and Emotional Stroop: A Systematic Review. J. Clin. Med. 2022, 11, 3259. [Google Scholar] [CrossRef]
- Bhatt, J.M.; Bhattacharyya, N.; Lin, H.W. Relationships between tinnitus and the prevalence of anxiety and depression. Laryngoscope 2017, 127, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Mullins, P.M.; Yong, R.J.; Bhattacharyya, N. Associations between chronic pain, anxiety, and depression among adults in the United States. Pain Pract. 2023, 23, 589–594. [Google Scholar] [CrossRef] [PubMed]
- McKenna, L.; Baguley, D.; McFerran, D. Living with Tinnitus and Hyperacusis: New Edition; Sheldon Press: London, UK, 2021. [Google Scholar]
- Tang, N.K. Insomnia Co-Occurring with Chronic Pain: Clinical Features, Interaction, Assessments and Possible Interventions. Rev. Pain 2008, 2, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Alster, J.; Shemesh, Z.; Ornan, M.; Attias, J. Sleep disturbance associated with chronic tinnitus. Biol. Psychiatry 1993, 34, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Milinski, L.; Nodal, F.R.; Vyazovskiy, V.V.; Bajo, V.M. Tinnitus: At a crossroad between phantom perception and sleep. Brain Commun. 2022, 4, fcac089. [Google Scholar] [CrossRef] [PubMed]
- Mathias, J.L.; Cant, M.L.; Burke, A.L.J. Sleep disturbances and sleep disorders in adults living with chronic pain: A meta-analysis. Sleep Med. 2018, 52, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; De Ridder, D. The auditory and non-auditory brain areas involved in tinnitus. An emergent property of multiple parallel overlapping subnetworks. Front. Syst. Neurosci. 2012, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Chang, M.C. Chronic Pain: Structural and Functional Changes in Brain Structures and Associated Negative Affective States. Int. J. Mol. Sci. 2019, 20, 3130. [Google Scholar] [CrossRef]
- Hullfish, J.; Sedley, W.; Vanneste, S. Prediction and perception: Insights for (and from) tinnitus. Neurosci. Biobehav. Rev. 2019, 102, 1–12. [Google Scholar] [CrossRef]
- Chen, Z.S. Hierarchical predictive coding in distributed pain circuits. Front. Neural Circuits 2023, 17, 1073537. [Google Scholar] [CrossRef]
- Song, Y.; Yao, M.; Kemprecos, H.; Byrne, A.; Xiao, Z.; Zhang, Q.; Singh, A.; Wang, J.; Chen, Z.S. Predictive coding models for pain perception. J. Comput. Neurosci. 2021, 49, 107–127. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.A.; Akrofi, K.; Carpenter-Thompson, J.R.; Husain, F.T. Default Mode, Dorsal Attention and Auditory Resting State Networks Exhibit Differential Functional Connectivity in Tinnitus and Hearing Loss. PLoS ONE 2013, 8, e76488. [Google Scholar] [CrossRef] [PubMed]
- Shahsavarani, S.; Schmidt, S.A.; Khan, R.A.; Tai, Y.; Husain, F.T. Salience, emotion, and attention: The neural networks underlying tinnitus distress revealed using music and rest. Brain Res. 2021, 1755, 147277. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; Plazier, M.; der Loo, E.v.; de Heyning, P.V.; Congedo, M.; De Ridder, D. The neural correlates of tinnitus-related distress. NeuroImage 2010, 52, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Davidson, C.; De Ridder, D.; Vanneste, S. Effective connectivity analysis of inter- and intramodular hubs in phantom sound perception–identifying the core distress network. Brain Imaging Behav. 2020, 14, 289–307. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Vanneste, S.; Song, J.J.; Adhia, D. Tinnitus and the Triple Network Model: A Perspective. Clin. Exp. Otorhinolaryngol. 2022, 15, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Dick, B.D.; Connolly, J.F.; McGrath, P.J.; Finley, G.A.; Stroink, G.; Houlihan, M.E.; Clark, A.J. The disruptive effect of chronic pain on mismatch negativity. Clin. Neurophysiol. 2003, 114, 1497–1506. [Google Scholar] [CrossRef]
- Fan, L.; Sun, Y.-B.; Sun, Z.-K.; Wang, N.; Luo, F.; Yu, F.; Wang, J.-Y. Modulation of auditory sensory memory by chronic clinical pain and acute experimental pain: A mismatch negativity study. Sci. Rep. 2018, 8, 15673. [Google Scholar] [CrossRef]
- Yao, S.; Liu, X.; Yang, W.; Wang, X. Preattentive processing abnormalities in chronic pain: Neurophysiological evidence from mismatch negativity. Pain Med. 2011, 12, 773–781. [Google Scholar] [CrossRef]
- Choi, W.; Lim, M.; Kim, J.S.; Kim, D.J.; Chung, C.K. Impaired pre-attentive auditory processing in fibromyalgia: A mismatch negativity (MMN) study. Clin. Neurophysiol. 2015, 126, 1310–1318. [Google Scholar] [CrossRef]
- Guo, Y.; Tian, Q.; Xu, S.; Han, M.; Sun, Y.; Hong, Y.; Hou, X.; Liu, X. The impact of attack frequency and duration on neurocognitive processing in migraine sufferers: Evidence from event-related potentials using a modified oddball paradigm. BMC Neurol. 2019, 19, 73. [Google Scholar] [CrossRef] [PubMed]
- Tomasevic-Todorovic, S.; Boskovic, K.; Filipovic, D.; Milekic, B.; Grajic, M.; Hanna, F. Auditory Event-Related P300 Potentials in Rheumatoid Arthritis Patients. Neurophysiology 2015, 47, 138–143. [Google Scholar] [CrossRef]
- Niso, G.; Tjepkema-Cloostermans, M.C.; Lenders, M.W.P.M.; de Vos, C.C. Modulation of the Somatosensory Evoked Potential by Attention and Spinal Cord Stimulation. Front. Neurol. 2021, 12, 4310. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, P.; Liossi, C.; Schoth, D.E. Attentional bias to somatosensory stimuli in chronic pain patients: A systematic review and meta-analysis. Pain 2021, 162, 332–352. [Google Scholar] [CrossRef] [PubMed]
- Sendesen, E.; Erbil, N.; Türkyılmaz, M.D. The mismatch negativity responses of individuals with tinnitus with normal extended high-frequency hearing-is it possible to use mismatch negativity in the evaluation of tinnitus? Eur. Arch. Otorhinolaryngol. 2022, 279, 3425–3434. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Lu, X.; Sun, S. Application of auditory mismatch negativity in tinnitus patients based on high-resolution electroencephalogram signals. Transl. Neurosci. 2022, 13, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Mohebbi, M.; Daneshi, A.; Asadpour, A.; Mohsen, S.; Farhadi, M.; Mahmoudian, S. The potential role of auditory prediction error in decompensated tinnitus: An auditory mismatch negativity study. Brain Behav. 2019, 9, e01242. [Google Scholar] [CrossRef] [PubMed]
- Weisz, N.; Voss, S.; Berg, P.; Elbert, T. Abnormal auditory mismatch response in tinnitus sufferers with high-frequency hearing loss is associated with subjective distress level. BMC Neurosci. 2004, 5, 8. [Google Scholar] [CrossRef]
- Cardon, E.; Joossen, I.; Vermeersch, H.; Jacquemin, L.; Mertens, G.; Vanderveken, O.M.; Topsakal, V.; Van de Heyning, P.; Van Rompaey, V.; Gilles, A. Systematic review and meta-analysis of late auditory evoked potentials as a candidate biomarker in the assessment of tinnitus. PLoS ONE 2020, 15, e0243785. [Google Scholar] [CrossRef]
- Møller, A.R. Tinnitus and pain. Prog. Brain Res. 2007, 166, 47–53. [Google Scholar] [CrossRef]
- Isler, B.; von Burg, N.; Kleinjung, T.; Meyer, M.; Stämpfli, P.; Zölch, N.; Neff, P. Lower glutamate and GABA levels in auditory cortex of tinnitus patients: A 2D-JPRESS MR spectroscopy study. Sci. Rep. 2022, 12, 4068. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lei, Y.; Tian, Y.; Xu, S.; Shen, X.; Wu, H.; Bao, S.; Wang, F. The etiological contribution of GABAergic plasticity to the pathogenesis of neuropathic pain. Mol. Pain 2019, 15, 1744806919847366. [Google Scholar] [CrossRef] [PubMed]
- Stolzberg, D.; Salvi, R.J.; Allman, B.L. Salicylate toxicity model of tinnitus. Front. Syst. Neurosci. 2012, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Goljanian Tabrizi, A.; Safavi Naini, A.; Baradaran, N. Short-Term Effect of Gabapentin on Subjective Tinnitus in Acoustic Trauma Patients. Iran. J. Otorhinolaryngol. 2017, 29, 95–100. [Google Scholar] [PubMed]
- Peek, A.L.; Rebbeck, T.; Puts, N.A.J.; Watson, J.; Aguila, M.-E.R.; Leaver, A.M. Brain GABA and glutamate levels across pain conditions: A systematic literature review and meta-analysis of 1H-MRS studies using the MRS-Q quality assessment tool. NeuroImage 2020, 210, 116532. [Google Scholar] [CrossRef]
- Barr, M.S.; Farzan, F.; Davis, K.D.; Fitzgerald, P.B.; Daskalakis, Z.J. Measuring GABAergic Inhibitory Activity with TMS-EEG and Its Potential Clinical Application for Chronic Pain. J. Neuroimmune Pharmacol. 2013, 8, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Gonzalez, M.A.; Esteban-Ortega, F. Tinnitus dopaminergic pathway. Ear noises treatment by dopamine modulation. Med. Hypotheses 2005, 65, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, S.; Lu, X.; Tao, F. Role of Descending Dopaminergic Pathways in Pain Modulation. Curr. Neuropharmacol. 2019, 17, 1176–1182. [Google Scholar] [CrossRef]
- Hruschak, V.; Cochran, G. Psychosocial predictors in the transition from acute to chronic pain: A systematic review. Psychol. Health Med. 2018, 23, 1151–1167. [Google Scholar] [CrossRef]
- Wallhäusser-Franke, E.; D’Amelio, R.; Glauner, A.; Delb, W.; Servais, J.J.; Hörmann, K.; Repik, I. Transition from Acute to Chronic Tinnitus: Predictors for the Development of Chronic Distressing Tinnitus. Front. Neurol. 2017, 8, 605. [Google Scholar] [CrossRef]
- Vielsmeier, V.; Santiago Stiel, R.; Kwok, P.; Langguth, B.; Schecklmann, M. From Acute to Chronic Tinnitus: Pilot Data on Predictors and Progression. Front. Neurol. 2020, 11, 997. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ji, L.; Wang, L.; Yin, Z.; Cen, J.; Guo, Y. Clinical characteristics and psychoacoustic analysis of acute and chronic subjective tinnitus. Laryngoscope Investig. Otolaryngol. 2023, 8, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Lan, L.; Li, J.; Chen, Y.; Chen, W.; Li, W.; Zhao, F.; Chen, G.; Liu, J.; Chen, Y.; Li, Y.; et al. Alterations of brain activity and functional connectivity in transition from acute to chronic tinnitus. Hum. Brain Mapp. 2021, 42, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Baliki, M.N.; Petre, B.; Torbey, S.; Herrmann, K.M.; Huang, L.; Schnitzer, T.J.; Fields, H.L.; Apkarian, A.V. Corticostriatal functional connectivity predicts transition to chronic back pain. Nat. Neurosci. 2012, 15, 1117–1119. [Google Scholar] [CrossRef] [PubMed]
- Mutso, A.A.; Petre, B.; Huang, L.; Baliki, M.N.; Torbey, S.; Herrmann, K.M.; Schnitzer, T.J.; Apkarian, A.V. Reorganization of hippocampal functional connectivity with transition to chronic back pain. J. Neurophysiol. 2014, 111, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Apkarian, A.V.; Baliki, M.N.; Farmer, M.A. Predicting transition to chronic pain. Curr. Opin. Neurol. 2013, 26, 360. [Google Scholar] [CrossRef]
- Glare, P.; Overton, S.; Aubrey, K. Transition from acute to chronic pain: Where cells, systems and society meet. Pain Manag. 2020, 10, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Mifflin, K.A.; Kerr, B.J. The transition from acute to chronic pain: Understanding how different biological systems interact. Can. J. Anesth. 2014, 61, 112–122. [Google Scholar] [CrossRef]
- Murawski, N.J.; Moore, E.M.; Thomas, J.D.; Riley, E.P. Advances in diagnosis and treatment of fetal alcohol spectrum disorders: From animal models to human studies. Alcohol Res. Curr. Rev. 2015, 37, 97. [Google Scholar]
- O’Leary-Moore, S.K.; Parnell, S.E.; Lipinski, R.J.; Sulik, K.K. Magnetic resonance-based imaging in animal models of fetal alcohol spectrum disorder. Neuropsychol. Rev. 2011, 21, 167–185. [Google Scholar] [CrossRef]
- Godin, E.A.; O’Leary-Moore, S.K.; Khan, A.A.; Parnell, S.E.; Ament, J.J.; Dehart, D.B.; Johnson, B.W.; Allan Johnson, G.; Styner, M.A.; Sulik, K.K. Magnetic resonance microscopy defines ethanol-induced brain abnormalities in prenatal mice: Effects of acute insult on gestational day 7. Alcohol. Clin. Exp. Res. 2010, 34, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Parnell, S.E.; Holloway, H.T.; O’Leary-Moore, S.K.; Dehart, D.B.; Paniaqua, B.; Oguz, I.; Budin, F.; Styner, M.A.; Johnson, G.A.; Sulik, K.K. Magnetic resonance microscopy-based analyses of the neuroanatomical effects of gestational day 9 ethanol exposure in mice. Neurotoxicol. Teratol. 2013, 39, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.M.; Migliorini, R.; Infante, M.A.; Riley, E.P. Fetal alcohol spectrum disorders: Recent neuroimaging findings. Curr. Dev. Disord. Rep. 2014, 1, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Hudak, M.L.; Tan, R.C. The Committee on Drugs and the Committee on Fetus and Newborn. Neonatal Drug Withdrawal (vol 129, pg e540, 2012). Pediatrics 2014, 133, 937–938. [Google Scholar]
- Palla, M.R.; Khan, G.; Haghighat, Z.M.; Bada, H. EEG findings in infants with neonatal abstinence syndrome presenting with clinical seizures. Front. Pediatr. 2019, 7, 111. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Rubic, M.; Seah, J.; Rae, C.; Wright, I.M.; Kaltenbach, K.; Feller, J.; Abdel-Latif, M.E.; Chu, C.; Oei, J.L. Do maternal opioids reduce neonatal regional brain volumes? A pilot study. J. Perinatol. 2014, 34, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.L.; Moore, C.F.; Gajewski, L.L.; Larson, J.A.; Roberts, A.D.; Converse, A.K.; DeJesus, O.T. Sensory processing disorder in a primate model: Evidence from a longitudinal study of prenatal alcohol and prenatal stress effects. Child Dev. 2008, 79, 100–113. [Google Scholar] [CrossRef]
- Ryckman, J.; Hilton, C.; Rogers, C.; Pineda, R. Sensory processing disorder in preterm infants during early childhood and relationships to early neurobehavior. Early Hum. Dev. 2017, 113, 18–22. [Google Scholar] [CrossRef]
- Mitchell, A.W.; Moore, E.M.; Roberts, E.J.; Hachtel, K.W.; Brown, M.S. Sensory processing disorder in children ages birth–3 years born prematurely: A systematic review. Am. J. Occup. Ther. 2015, 69, 6901220030. [Google Scholar] [CrossRef]
- Ahn, R.R.; Miller, L.J.; Milberger, S.; McIntosh, D.N. Prevalence of parents’ perceptions of sensory processing disorders among kindergarten children. Am. J. Occup. Ther. 2004, 58, 287–293. [Google Scholar] [CrossRef]
- McMahon, K.; Anand, D.; Morris-Jones, M.; Rosenthal, M.Z. A path from childhood sensory processing disorder to anxiety disorders: The mediating role of emotion dysregulation and adult sensory processing disorder symptoms. Front. Integr. Neurosci. 2019, 13, 22. [Google Scholar] [CrossRef]
- Koechlin, H.; Coakley, R.; Schechter, N.; Werner, C.; Kossowsky, J. The role of emotion regulation in chronic pain: A systematic literature review. J. Psychosom. Res. 2018, 107, 38–45. [Google Scholar] [CrossRef]
- Bar-Shalita, T.; Vatine, J.-J.; Seltzer, Z.e.; Parush, S. Psychophysical correlates in children with sensory modulation disorder (SMD). Physiol. Behav. 2009, 98, 631–639. [Google Scholar] [CrossRef]
- Prego-Domínguez, J.; Khazaeipour, Z.; Mallah, N.; Takkouche, B. Socioeconomic status and occurrence of chronic pain: A meta-analysis. Rheumatology 2021, 60, 1091–1105. [Google Scholar] [CrossRef]
- Gill, S.V.; May-Benson, T.A.; Teasdale, A.; Munsell, E.G. Birth and developmental correlates of birth weight in a sample of children with potential sensory processing disorder. BMC Pediatr. 2013, 13, 29. [Google Scholar] [CrossRef]
- Bailey, B.E.; Freedenfeld, R.N.; Kiser, R.S.; Gatchel, R.J. Lifetime physical and sexual abuse in chronic pain patients: Psychosocial correlates and treatment outcomes. Disabil. Rehabil. 2003, 25, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Linton, S.J. A population-based study of the relationship between sexual abuse and back pain: Establishing a link. Pain 1997, 73, 47–53. [Google Scholar] [CrossRef]
- Davis, D.A.; Luecken, L.J.; Zautra, A.J. Are Reports of Childhood Abuse Related to the Experience of Chronic Pain in Adulthood?: A Meta-analytic Review of the Literature. Clin. J. Pain 2005, 21, 398–405. [Google Scholar] [CrossRef]
- Paras, M.L.; Murad, M.H.; Chen, L.P.; Goranson, E.N.; Sattler, A.L.; Colbenson, K.M.; Elamin, M.B.; Seime, R.J.; Prokop, L.J.; Zirakzadeh, A. Sexual abuse and lifetime diagnosis of somatic disorders: A systematic review and meta-analysis. JAMA 2009, 302, 550–561. [Google Scholar] [CrossRef]
- Behere, A.P.; Basnet, P.; Campbell, P. Effects of family structure on mental health of children: A preliminary study. Indian J. Psychol. Med. 2017, 39, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.R.H.; Lynch, A.K.; Call, C.D.; Cross, D.R. Sensory processing in children with a history of maltreatment: An occupational therapy perspective. Vulnerable Child. Youth Stud. 2020, 15, 60–67. [Google Scholar] [CrossRef]
- Rosati, R.; Jamesdaniel, S. Environmental exposures and hearing loss. Int. J. Environ. Res. Public Health 2020, 17, 4879. [Google Scholar] [CrossRef]
- Morata, T.C.; Dunn, D.E.; Sieber, W.K. Occupational exposure to noise and ototoxic organic solvents. Arch. Environ. Health Int. J. 1994, 49, 359–365. [Google Scholar] [CrossRef]
- Sliwinska-Kowalska, M.; Zamyslowska-Szmytke, E.; Szymczak, W.; Kotylo, P.; Fiszer, M.; Wesolowski, W.; Pawlaczyk-Luszczynska, M. Ototoxic effects of occupational exposure to styrene and co-exposure to styrene and noise. J. Occup. Environ. Med. 2003, 45, 15–24. [Google Scholar] [CrossRef]
- Staudt, A.M.; Whitworth, K.W.; Chien, L.-C.; Whitehead, L.W.; de Porras, D.G.R. Association of organic solvents and occupational noise on hearing loss and tinnitus among adults in the US, 1999–2004. Int. Arch. Occup. Environ. Health 2019, 92, 403–413. [Google Scholar] [CrossRef]
- Roggia, S.M.; de França, A.G.; Morata, T.C.; Krieg, E.; Earl, B.R. Auditory system dysfunction in Brazilian gasoline station workers. Int. J. Audiol. 2019, 58, 484–496. [Google Scholar] [CrossRef]
- Bolden, A.L.; Kwiatkowski, C.F.; Colborn, T. New look at BTEX: Are ambient levels a problem? Environ. Sci. Technol. 2015, 49, 5261–5276. [Google Scholar] [CrossRef]
- Danioth, L.; Brotschi, G.; Croy, I.; Friedrich, H.; Caversaccio, M.-D.; Negoias, S. Multisensory environmental sensitivity in patients with chronic tinnitus. J. Psychosom. Res. 2020, 135, 110155. [Google Scholar] [CrossRef] [PubMed]
- Beukes, E.W.; Baguley, D.M.; Jacquemin, L.; Lourenco, M.P.; Allen, P.M.; Onozuka, J.; Stockdale, D.; Kaldo, V.; Andersson, G.; Manchaiah, V. Changes in tinnitus experiences during the COVID-19 pandemic. Front. Public Health 2020, 8, 681. [Google Scholar] [CrossRef] [PubMed]
- Ceban, F.; Ling, S.; Lui, L.M.W.; Lee, Y.; Gill, H.; Teopiz, K.M.; Rodrigues, N.B.; Subramaniapillai, M.; Di Vincenzo, J.D.; Cao, B.; et al. Fatigue and cognitive impairment in Post-COVID-19 Syndrome: A systematic review and meta-analysis. Brain Behav. Immun. 2022, 101, 93–135. [Google Scholar] [CrossRef] [PubMed]
- Morris, P.; Ali, K.; Merritt, M.; Pelletier, J.; Macedo, L.G. A systematic review of the role of inflammatory biomarkers in acute, subacute and chronic non-specific low back pain. BMC Musculoskelet. Disord. 2020, 21, 142. [Google Scholar] [CrossRef]
- Eisen, S.A.; Kang, H.K.; Murphy, F.M.; Blanchard, M.S.; Reda, D.J.; Henderson, W.G.; Toomey, R.; Jackson, L.W.; Alpern, R.; Parks, B.J. Gulf War veterans’ health: Medical evaluation of a US cohort. Ann. Intern. Med. 2005, 142, 881–890. [Google Scholar] [CrossRef]
- Henry, J.A.; Griest, S.E.; Blankenship, C.; Thielman, E.J.; Theodoroff, S.M.; Hammill, T.; Carlson, K.F. Impact of Tinnitus on Military Service Members. Mil. Med. 2019, 184, 604–614. [Google Scholar] [CrossRef]
- Kühn, S.; Butler, O.; Willmund, G.; Wesemann, U.; Zimmermann, P.; Gallinat, J. The brain at war: Effects of stress on brain structure in soldiers deployed to a war zone. Transl. Psychiatry 2021, 11, 247. [Google Scholar] [CrossRef]
- Kadir, A.; Shenoda, S.; Goldhagen, J. Effects of armed conflict on child health and development: A systematic review. PLoS ONE 2019, 14, e0210071. [Google Scholar] [CrossRef]
- Constantino, J.N.; Todd, R.D. Autistic traits in the general population: A twin study. Arch. Gen. Psychiatry 2003, 60, 524–530. [Google Scholar] [CrossRef]
- Maas, I.L.; Brüggemann, P.; Requena, T.; Bulla, J.; Edvall, N.K.; vB Hjelmborg, J.; Szczepek, A.J.; Canlon, B.; Mazurek, B.; Lopez-Escamez, J.A. Genetic susceptibility to bilateral tinnitus in a Swedish twin cohort. Genet. Med. 2017, 19, 1007–1012. [Google Scholar] [CrossRef]
- Gejman, P.V.; Sanders, A.R.; Duan, J. The role of genetics in the etiology of schizophrenia. Psychiatr. Clin. 2010, 33, 35–66. [Google Scholar] [CrossRef]
- Vanneste, S.; Alsalman, O.; De Ridder, D. COMT and the neurogenetic architecture of hearing loss induced tinnitus. Hear. Res. 2018, 365, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, I.S.; Wilson, N.; Dias, R.; Torkamani, A. A genome-wide association study of tinnitus reveals shared genetic links to neuropsychiatric disorders. Sci. Rep. 2022, 12, 22511. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.J.A.; Adams, M.J.; Nicholl, B.I.; Ward, J.; Strawbridge, R.J.; Ferguson, A.; McIntosh, A.M.; Bailey, M.E.S.; Smith, D.J. Genome-wide association study of multisite chronic pain in UK Biobank. PLoS Genet. 2019, 15, e1008164. [Google Scholar] [CrossRef]
- Schaaf, C.P. Nicotinic acetylcholine receptors in human genetic disease. Genet. Med. 2014, 16, 649–656. [Google Scholar] [CrossRef]
- Nagatsu, T. The catecholamine system in health and disease -Relation to tyrosine 3-monooxygenase and other catecholamine-synthesizing enzymes. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2007, 82, 388–415. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Phase | Neural Markers | Molecular Markers |
|---|---|---|
| Acute phase | Changes in auditory MMN in both tinnitus and pain. Larger P300 to deviant/novel noxious stimuli compared to standard. P300 amplitudes to novel stimuli enhanced with attention. Acute pain impacted performance on cognitive tasks Similar patterns of activation as chronic tinnitus seen in generation of acute intermittent ringing. | Increase in excitatory post-synaptic potential in acute tinnitus and pain. Pain: inflammatory “soup” that regulates pain perception. Tinnitus: changes in conductance of outer hair cells by decreasing GABA or increasing NMDA. |
| Chronic Phase: has a biopsychosocial component where the disorder is accompanied by a myriad of symptoms including reduced concentration, attention on external tasks and increased focus on the percept (pain/tinnitus), mood disorders and reduced sleep. | Pain: Changes in MMN and P300 to auditory and visual oddballs. Reduction in MMN correlates with McGill Pain Questionnaire. Increase in somatosensory ERP, possibly explained by an attentional bias towards somatosensory stimuli. Tinnitus: Mixed results in auditory oddball in the MMN timeframe. Meta-analysis showed a reduction in P300 amplitude. | Low glutamate and GABA levels in both pain and tinnitus. Pain: level of glutamate and GABA changed with different types of pain. Tinnitus: NMDA receptor activation increases tinnitus, NMDA receptor antagonist improves tinnitus. |
| Transitionary phase: psychosocial components start to play a role in this transition: Pain: systematic review shows that depression, fear avoidance, pain catastrophising, high traumatic exposure, emotional distress, feeling of helplessness and hopelessness play a role in transition from acute to chronic state. Tinnitus: Preliminary evidence suggests that depression, anxiety levels, hyperacusis, severity and frequency of hearing loss may play a role in transition from acute to chronic state. | Pain: decrease in grey matter volume in insula, nucleus accumbens, striatum and sensorimotor cortex; decreased connectivity between medial prefrontal cortex and hippocampus; connectivity between medial prefrontal cortex and nucleus accumbens determined transition accuracy by >80%. Tinnitus: Increased activity in regions of predictive coding with increased connectivity between parahippocampus and posterior cingulate cortex. Theoretical evidence suggests a change in the prediction from silence to perceiving a ringing in the absence of external stimuli. | Pain: recruitment of NMDA receptors in the dorsal horn, release of BDNF leading to a depolarising shift in dorsal horn causing GABA-related signals to become excitatory ending in hyperalgesia. Tinnitus: not a lot is known about molecular markers of transition from acute to chronic phase of tinnitus, to our knowledge. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasoda-Mohan, A.; Vanneste, S. Development, Insults and Predisposing Factors of the Brain’s Predictive Coding System to Chronic Perceptual Disorders—A Life-Course Examination. Brain Sci. 2024, 14, 86. https://doi.org/10.3390/brainsci14010086
Yasoda-Mohan A, Vanneste S. Development, Insults and Predisposing Factors of the Brain’s Predictive Coding System to Chronic Perceptual Disorders—A Life-Course Examination. Brain Sciences. 2024; 14(1):86. https://doi.org/10.3390/brainsci14010086
Chicago/Turabian StyleYasoda-Mohan, Anusha, and Sven Vanneste. 2024. "Development, Insults and Predisposing Factors of the Brain’s Predictive Coding System to Chronic Perceptual Disorders—A Life-Course Examination" Brain Sciences 14, no. 1: 86. https://doi.org/10.3390/brainsci14010086
APA StyleYasoda-Mohan, A., & Vanneste, S. (2024). Development, Insults and Predisposing Factors of the Brain’s Predictive Coding System to Chronic Perceptual Disorders—A Life-Course Examination. Brain Sciences, 14(1), 86. https://doi.org/10.3390/brainsci14010086