

A Systematic Review of Structural and Functional MRI Studies Investigating Social Networking Site Use

Abstract
1. Introduction
2. Method
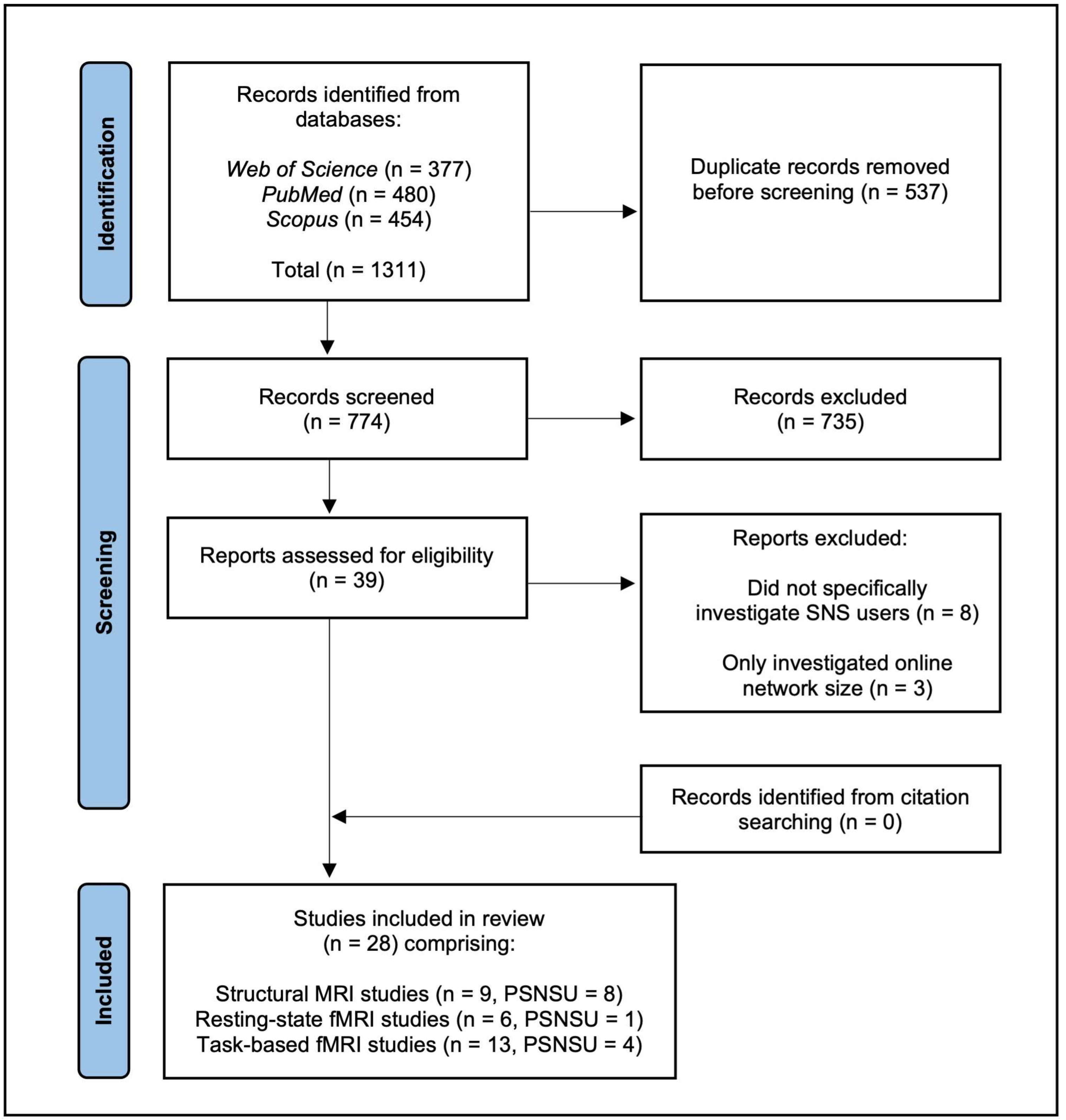
2.1. Search Strategy
2.2. Study Selection
2.3. Data Extraction
2.4. Quality Assessment
3. Results
3.1. Structural MRI Studies

{kind=link}
| Authors (Year) | Sample | Mean Age | PSNSU | SNS Assessment Tool | Design | Main Results | Quality Assessment |
|---|---|---|---|---|---|---|---|
| Achterberg et al. (2022) [56] |
|
|
|
|
|
|
|
| He, Turel and Bechara (2017) [49] a |
|
|
|
|
|
|
|
| He et al. (2017) [47] |
|
|
|
|
|
|
|
| He et al. (2018) [57] a |
|
|
|
|
|
|
|
| Lee et al. (2019) [35] b |
|
|
|
|
|
|
|
| Montag et al. (2017) [46] |
|
|
|
|
|
|
|
| Montag et al. (2018) [48] |
|
|
|
|
|
|
|
| Turel et al. (2018a) [53] |
|
|
|
|
|
|
|
| Turel et al. (2018b) [54] |
|
|
|
|
|
|
|
3.2. Resting-State fMRI Studies
| Authors (Year) | Sample | Mean Age | PSNSU | SNS Assessment Tool | Design | Main Results | Quality Assessment |
|---|---|---|---|---|---|---|---|
| Hu, Cui et al. (2022) [68] |
|
|
|
|
|
|
|
| Hu, Yu et al. (2022) [70] |
|
|
|
|
|
|
|
| Lee et al. (2021) [36] a |
|
|
|
|
|
|
|
| Meshi et al. (2016) [64] |
|
|
|
|
|
|
|
| Wilmer et al. (2019) [37] |
|
|
|
|
|
|
|
| Zhang and Mo (2016) [67] b |
|
|
|
|
|
|
|
3.3. Task-Based fMRI Studies
| Authors (Year) | Sample | Mean Age | PSNSU | SNS Assessment Tool | Design | Main Results | Quality Assessment |
|---|---|---|---|---|---|---|---|
| Dieter et al. (2017) [38] |
|
|
|
|
|
|
|
| Leménager et al. (2016) [82] |
|
|
|
|
|
|
|
| Meshi, Morawetz and Heekeren (2013) [73] |
|
|
|
|
|
|
|
| Nasser et al. (2020) [77] |
|
|
|
|
|
|
|
| Peters et al. (2021) [83] |
|
|
|
|
|
|
|
| Sherman et al. (2016) [74] a |
|
|
|
|
|
|
|
| Sherman, Greenfield et al. (2018) [75] a |
|
|
|
|
|
|
|
| Sherman, Hernandez et al. (2018) [76] a |
|
|
|
|
|
|
|
| Su, Zhou, Gong et al. (2021) [80] b |
|
|
|
|
|
|
|
| Su, Zhou, Wang et al. (2021) [81] b |
|
|
|
|
|
|
|
| Turel et al. (2014) [63] c |
|
|
|
|
|
|
|
| Wikman et al. (2022) [79] |
|
|
|
|
|
|
|
| Zhang and Qu (2020) [72] d |
|
|
|
|
|
|
|
4. Discussion
4.1. Ventral Striatum/Nucleus Accumbens
4.2. Prefrontal Cortex
4.3. Amygdala
4.4. Cingulate Cortex
4.5. Precuneus
4.6. Temporal Parietal Junction
4.7. Insula
4.8. Other Observations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- DataReportal. Digital 2012: Global Digital Overview. Available online: https://datareportal.com/reports/digital-2012-global-digital-overview (accessed on 18 May 2022).
- DataReportal. Digital 2022: Global Overview Report. Available online: https://datareportal.com/reports/digital-2022-global-overview-report (accessed on 18 May 2022).
- Statista. Daily Time Spent on Social Networking by Internet Users Worldwide from 2012 to 2020. Available online: https://www.statista.com/statistics/433871/daily-social-media-usage-worldwide/ (accessed on 18 May 2022).
- Kelly, Y.; Zilanawala, A.; Booker, C.; Sacker, A. Social media use and adolescent mental health: Findings from the UK Millennium Cohort Study. EClinicalMedicine 2018, 6, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Twenge, J.M.; Joiner, T.E.; Rogers, M.L.; Martin, G.N. Increases in depressive symptoms, suicide-related outcomes, and suicide rates among US adolescents after 2010 and links to increased new media screen time. Clin. Psychol. Sci. 2018, 6, 3–17. [Google Scholar] [CrossRef]
- Twenge, J.M.; Martin, G.N.; Campbell, W.K. Decreases in psychological well-being among American adolescents after 2012 and links to screen time during the rise of smartphone technology. Emotion 2018, 18, 765–780. [Google Scholar] [CrossRef]
- Orben, A.; Przybylski, A.K. The association between adolescent well-being and digital technology use. Nat. Hum. Behav. 2019, 3, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Vuorre, M.; Orben, A.; Przybylski, A.K. There is no evidence that associations between adolescents’ digital technology engagement and mental health problems have increased. Clin. Psychol. Sci. 2021, 9, 823–835. [Google Scholar] [CrossRef]
- Cheng, C.; Lau, Y.-C.; Chan, L.; Luk, J.W. Prevalence of social media addiction across 32 nations: Meta-analysis with subgroup analysis of classification schemes and cultural values. Addict. Behav. 2021, 117, 106845. [Google Scholar] [CrossRef] [PubMed]
- Johannes, N.; Dora, J.; Rusz, D. Social smartphone apps do not capture attention despite their perceived high reward value. Collabra Psychol. 2019, 5, 14. [Google Scholar] [CrossRef]
- Thomson, K.; Hunter, S.C.; Butler, S.H.; Robertson, D.J. Social media ‘addiction’: The absence of an attentional bias to social media stimuli. J. Behav. Addict. 2021, 10, 302–313. [Google Scholar] [CrossRef]
- Wilcockson, T.D.; Osborne, A.M.; Ellis, D.A. Digital detox: The effect of smartphone abstinence on mood, anxiety, and craving. Addict. Behav. 2019, 99, 106013. [Google Scholar] [CrossRef]
- Wadsley, M.; Ihssen, N. The roles of implicit approach motivation and explicit reward in excessive and problematic use of social networking sites. PLoS ONE 2022, 17, e0264738. [Google Scholar] [CrossRef] [PubMed]
- Billieux, J.; Schimmenti, A.; Khazaal, Y.; Maurage, P.; Heeren, A. Are we overpathologizing everyday life? A tenable blueprint for behavioral addiction research. J. Behav. Addict. 2015, 4, 119–123. [Google Scholar] [CrossRef]
- Kardefelt-Winther, D. Commentary on: Are we overpathologizing everyday life? A tenable blueprint for behavioral addiction research: Problems with atheoretical and confirmatory research approaches in the study of behavioral addictions. J. Behav. Addict. 2015, 4, 126–129. [Google Scholar] [CrossRef] [PubMed]
- Maraz, A.; Király, O.; Demetrovics, Z. Commentary on: Are we overpathologizing everyday life? A tenable blueprint for behavioral addiction research: The diagnostic pitfalls of surveys: If you score positive on a test of addiction, you still have a good chance not to be addicted. J. Behav. Addict. 2015, 4, 151–154. [Google Scholar] [CrossRef]
- Koob, G.F.; Le Moal, M. Drug abuse: Hedonic homeostatic dysregulation. Science 1997, 278, 52–58. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. The neural basis of drug craving: An incentive-sensitization theory of addiction. Brain Res. Rev. 1993, 18, 247–291. [Google Scholar] [CrossRef] [PubMed]
- Fowler, J.S.; Volkow, N.D.; Kassed, C.A.; Chang, L. Imaging the addicted human brain. Sci. Pract. Perspect. 2007, 3, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Goudriaan, A.E.; De Ruiter, M.B.; Van Den Brink, W.; Oosterlaan, J.; Veltman, D.J. Brain activation patterns associated with cue reactivity and craving in abstinent problem gamblers, heavy smokers and healthy controls: An fMRI study. Addict. Biol. 2010, 15, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Limbrick-Oldfield, E.H.; Mick, I.; Cocks, R.; McGonigle, J.; Sharman, S.; Goldstone, A.P.; Stokes, P.; Waldman, A.; Erritzoe, D.; Bowden-Jones, H. Neural substrates of cue reactivity and craving in gambling disorder. Transl. Psychiatry 2017, 7, e992. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Fellows, L.; Small, D.; Dagher, A. Food and drug cues activate similar brain regions: A meta-analysis of functional MRI studies. Physiol. Behav. 2012, 106, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, J.M.; Versace, F.; Robinson, J.D.; Minnix, J.A.; Lam, C.Y.; Cui, Y.; Brown, V.L.; Cinciripini, P.M. Neural substrates of smoking cue reactivity: A meta-analysis of fMRI studies. Neuroimage 2012, 60, 252–262. [Google Scholar] [CrossRef]
- Quaglieri, A.; Mari, E.; Boccia, M.; Piccardi, L.; Guariglia, C.; Giannini, A.M. Brain network underlying executive functions in gambling and alcohol use disorders: An activation likelihood estimation meta-analysis of fMRI studies. Brain Sci. 2020, 10, 353. [Google Scholar] [CrossRef]
- van Holst, R.J.; de Ruiter, M.B.; van den Brink, W.; Veltman, D.J.; Goudriaan, A.E. A voxel-based morphometry study comparing problem gamblers, alcohol abusers, and healthy controls. Drug Alcohol Depend. 2012, 124, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Meshi, D.; Tamir, D.I.; Heekeren, H.R. The emerging neuroscience of social media. Trends Cogn. Sci. 2015, 19, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Ihssen, N.; Wadsley, M. A Reward and Incentive-Sensitization Perspective on Compulsive Use of Social Networking Sites–Wanting but not Liking Predicts Checking Frequency and Problematic Use Behavior. Addict. Behav. 2021, 116, 106808. [Google Scholar] [CrossRef]
- Berridge, K.C.; Robinson, T.E. Liking, wanting, and the incentive-sensitization theory of addiction. Am. Psychol. 2016, 71, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Huang, J.; Du, X. Enhanced reward sensitivity and decreased loss sensitivity in Internet addicts: An fMRI study during a guessing task. J. Psychiatr. Res. 2011, 45, 1525–1529. [Google Scholar] [CrossRef] [PubMed]
- Yuan, K.; Qin, W.; Wang, G.; Zeng, F.; Zhao, L.; Yang, X.; Liu, P.; Liu, J.; Sun, J.; von Deneen, K.M. Microstructure abnormalities in adolescents with internet addiction disorder. PLoS ONE 2011, 6, e20708. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lin, F.-C.; Du, Y.-S.; Zhao, Z.-M.; Xu, J.-R.; Lei, H. Gray matter abnormalities in Internet addiction: A voxel-based morphometry study. Eur. J. Radiol. 2011, 79, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Fernandez, O. How has internet addiction research evolved since the advent of internet gaming disorder? An overview of cyberaddictions from a psychological perspective. Curr. Addict. Rep. 2015, 2, 263–271. [Google Scholar] [CrossRef]
- Tashjian, S.M.; Galván, A. Dorsolateral prefrontal cortex response to negative tweets relates to executive functioning. Soc. Cogn. Affect. Neurosci. 2020, 15, 775–787. [Google Scholar] [CrossRef]
- Radke, S.; Jankowiak, K.; Tops, S.; Abel, T.; Habel, U.; Derntl, B. Neurobiobehavioral responses to virtual social rejection in females—Exploring the influence of oxytocin. Soc. Cogn. Affect. Neurosci. 2021, 16, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Namkoong, K.; Lee, J.; Lee, B.O.; Jung, Y.C. Lateral orbitofrontal gray matter abnormalities in subjects with problematic smartphone use. J. Behav. Addict. 2019, 8, 404–411. [Google Scholar] [CrossRef]
- Lee, D.; Lee, J.; Namkoong, K.; Jung, Y.-C. Altered functional connectivity of the dorsal attention network among problematic social network users. Addict. Behav. 2021, 116, 106823. [Google Scholar] [CrossRef] [PubMed]
- Wilmer, H.H.; Hampton, W.H.; Olino, T.M.; Olson, I.R.; Chein, J.M. Wired to be connected? Links between mobile technology engagement, intertemporal preference and frontostriatal white matter connectivity. Soc. Cogn. Affect. Neurosci. 2019, 14, 367–379. [Google Scholar] [CrossRef]
- Dieter, J.; Hoffmann, S.; Mier, D.; Reinhar, I.; Beutel, M.; Vollstadt-Klein, S.; Kiefer, F.; Mann, K.; Lemenager, T. The role of emotional inhibitory control in specific internet addiction—An fMRI study. Behav. Brain Res. 2017, 324, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kuss, D.J.; Griffiths, M.D. Internet and gaming addiction: A systematic literature review of neuroimaging studies. Brain Sci. 2012, 2, 347–374. [Google Scholar] [CrossRef] [PubMed]
- Sharifat, H.; Rashid, A.A.; Suppiah, S. Systematic review of the utility of functional MRI to investigate internet addiction disorder: Recent updates on resting state and task-based fMRI. Malays. J. Med. Health Sci. 2018, 14, 21–33. [Google Scholar]
- Sepede, G.; Tavino, M.; Santacroce, R.; Fiori, F.; Salerno, R.M.; Di Giannantonio, M. Functional magnetic resonance imaging of internet addiction in young adults. World J. Radiol. 2016, 8, 210. [Google Scholar] [CrossRef]
- Yin, T.; Li, Z.; Xiong, J.; Lan, L.; Sun, R.; Ren, F.; Zhang, P. Neuroimaging biomarkers of psychogenic erectile dysfunction: Protocol for a systematic review. BMJ Open 2019, 9, e030061. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.; Ciliska, D.; Dobbins, M.; Micucci, S. A process for systematically reviewing the literature: Providing the research evidence for public health nursing interventions. Worldviews Evid.-Based Nurs. 2004, 1, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Hengstschläger, A.; Sommerlad, A.; Huntley, J. What are the neural correlates of impaired awareness of social cognition and function in dementia? A systematic review. Brain Sci. 2022, 12, 1136. [Google Scholar] [CrossRef]
- McLachlan, E.; Rai, S.; Al-Shihabi, A.; Huntley, J.; Burgess, N.; Howard, R.; Reeves, S. Neuroimaging correlates of false memory in’Alzheimer’s disease: A preliminary systematic review. Psychiatry Res. Neuroimaging 2020, 296, 111021. [Google Scholar] [CrossRef] [PubMed]
- Montag, C.; Markowetz, A.; Blaszkiewicz, K.; Andone, I.; Lachmann, B.; Sariyska, R.; Trendafilov, B.; Eibes, M.; Kolb, J.; Reuter, M.; et al. Facebook usage on smartphones and gray matter volume of the nucleus accumbens. Behav. Brain Res. 2017, 329, 221–228. [Google Scholar] [CrossRef] [PubMed]
- He, Q.H.; Turel, O.; Brevers, D.; Bechara, A. Excess social media use in normal populations is associated with amygdala-striatal but not with prefrontal morphology. Psychiatry Res.-Neuroimaging 2017, 269, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Montag, C.; Zhao, Z.; Sindermann, C.; Xu, L.; Fu, M.; Li, J.; Zheng, X.; Li, K.; Kendrick, K.M.; Dai, J. Internet communication disorder and the structure of the human brain: Initial insights on WeChat addiction. Sci. Rep. 2018, 8, 2155. [Google Scholar] [CrossRef] [PubMed]
- He, Q.H.; Turel, O.; Bechara, A. Brain anatomy alterations associated with Social Networking Site (SNS) addiction. Sci. Rep. 2017, 7, 45064. [Google Scholar] [CrossRef]
- MacDonald, A.W.; Cohen, J.D.; Stenger, V.A.; Carter, C.S. Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science 2000, 288, 1835–1838. [Google Scholar] [CrossRef]
- Jentsch, J.D.; Pennington, Z.T. Reward, interrupted: Inhibitory control and its relevance to addictions. Neuropharmacology 2014, 76, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Makris, N.; Gasic, G.P.; Seidman, L.J.; Goldstein, J.M.; Gastfriend, D.R.; Elman, I.; Albaugh, M.D.; Hodge, S.M.; Ziegler, D.A.; Sheahan, F.S. Decreased absolute amygdala volume in cocaine addicts. Neuron 2004, 44, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Turel, O.; He, Q.; Brevers, D.; Bechara, A. Social networking sites use and the morphology of a social-semantic brain network. Soc. Neurosci. 2018, 13, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Turel, O.; He, Q.H.; Brevers, D.; Bechara, A. Delay discounting mediates the association between posterior insular cortex volume and social media addiction symptoms. Cogn. Affect. Behav. Neurosci. 2018, 18, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, N.H.; Bechara, A. The insula and drug addiction: An interoceptive view of pleasure, urges, and decision-making. Brain Struct. Funct. 2010, 214, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Achterberg, M.; Becht, A.; van der Cruijsen, R.; van de Groep, I.H.; Spaans, J.P.; Klapwijk, E.; Crone, E.A. Longitudinal associations between social media use, mental well-being and structural brain development across adolescence. Dev. Cogn. Neurosci. 2022, 54, 101088. [Google Scholar] [CrossRef] [PubMed]
- He, Q.H.; Turel, O.; Bechara, A. Association of excessive social media use with abnormal white matter integrity of the corpus callosum. Psychiatry Res.-Neuroimaging 2018, 278, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Meerkerk, G.-J.; Van Den Eijnden, R.J.; Vermulst, A.A.; Garretsen, H.F. The compulsive internet use scale (CIUS): Some psychometric properties. Cyberpsychol. Behav. 2009, 12, 1–6. [Google Scholar] [CrossRef]
- Kim, D.; Lee, Y.; Lee, J.; Nam, J.K.; Chung, Y. Development of Korean smartphone addiction proneness scale for youth. PLoS ONE 2014, 9, e97920. [Google Scholar] [CrossRef]
- Montag, C.; Bey, K.; Sha, P.; Li, M.; Chen, Y.F.; Liu, W.Y.; Zhu, Y.K.; Li, C.B.; Markett, S.; Keiper, J. Is it meaningful to distinguish between generalized and specific Internet addiction? Evidence from a cross-cultural study from Germany, Sweden, Taiwan and China. Asia-Pac. Psychiatry 2015, 7, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowski, M.; Altstötter-Gleich, C.; Brand, M. Validation and psychometric properties of a short version of Young’s Internet Addiction Test. Comput. Hum. Behav. 2013, 29, 1212–1223. [Google Scholar] [CrossRef]
- Van Rooij, A.J.; Schoenmakers, T.M.; Vermulst, A.A.; Van Den Eijnden, R.J.; Van De Mheen, D. Online video game addiction: Identification of addicted adolescent gamers. Addiction 2011, 106, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Turel, O.; He, Q.H.; Xue, G.; Xiao, L.; Bechara, A. Examination of neural systems sub-serving facebook “addiction”. Psychol. Rep. 2014, 115, 675–695. [Google Scholar] [CrossRef] [PubMed]
- Meshi, D.; Mamerow, L.; Kirilina, E.; Morawetz, C.; Margulies, D.S.; Heekeren, H.R. Sharing self-related information is associated with intrinsic functional connectivity of cortical midline brain regions. Sci. Rep. 2016, 6, 22491. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, C.J. Narcissism on Facebook: Self-promotional and anti-social behavior. Personal. Individ. Differ. 2012, 52, 482–486. [Google Scholar] [CrossRef]
- Northoff, G.; Heinzel, A.; De Greck, M.; Bermpohl, F.; Dobrowolny, H.; Panksepp, J. Self-referential processing in our brain—A meta-analysis of imaging studies on the self. Neuroimage 2006, 31, 440–457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Mo, L. Mentalizing and Information Propagation through Social Network: Evidence from a Resting-State-fMRI Study. Front. Psychol. 2016, 7, 1716. [Google Scholar] [CrossRef]
- Hu, B.; Cui, Y.-L.; Yu, Y.; Li, Y.-T.; Yan, L.-F.; Sun, J.-T.; Sun, Q.; Zhang, J.; Wang, W.; Cui, G.-B. Combining Dynamic Network Analysis and Cerebral Carryover Effect to Evaluate the Impacts of Reading Social Media Posts and Science Fiction in the Natural State on the Human Brain. Front. Neurosci. 2022, 16, 66. [Google Scholar] [CrossRef]
- Brewer, J.A.; Worhunsky, P.D.; Gray, J.R.; Tang, Y.-Y.; Weber, J.; Kober, H. Meditation experience is associated with differences in default mode network activity and connectivity. Proc. Natl. Acad. Sci. USA 2011, 108, 20254–20259. [Google Scholar] [CrossRef]
- Hu, B.; Yu, Y.; Yan, L.F.; Qi, G.Q.; Wu, D.; Li, Y.T.; Shi, A.P.; Liu, C.X.; Shang, Y.X.; Li, Z.Y. Intersubject correlation analysis reveals the plasticity of cerebral functional connectivity in the long-term use of social media. Hum. Brain Mapp. 2022, 43, 2262–2275. [Google Scholar] [CrossRef]
- Wilmer, H.H.; Chein, J.M. Mobile technology habits: Patterns of association among device usage, intertemporal preference, impulse control, and reward sensitivity. Psychon. Bull. Rev. 2016, 23, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Qu, C. Emotional, especially negative microblogs are more popular on the web: Evidence from an fMRI study. Brain Imaging Behav. 2020, 14, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Meshi, D.; Morawetz, C.; Heekeren, H.R. Nucleus accumbens response to gains in reputation for the self relative to gains for others predicts social media use. Front. Hum. Neurosci. 2013, 7, 439. [Google Scholar] [CrossRef]
- Sherman, L.E.; Payton, A.A.; Hernandez, L.M.; Greenfield, P.M.; Dapretto, M. The power of the like in adolescence: Effects of peer influence on neural and behavioral responses to social media. Psychol. Sci. 2016, 27, 1027–1035. [Google Scholar] [CrossRef]
- Sherman, L.E.; Greenfield, P.M.; Hernandez, L.M.; Dapretto, M. Peer Influence Via Instagram: Effects on Brain and Behavior in Adolescence and Young Adulthood. Child Dev. 2018, 89, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Sherman, L.E.; Hernandez, L.M.; Greenfield, P.M.; Dapretto, M. What the brain ‘Likes’: Neural correlates of providing feedback on social media. Soc. Cogn. Affect. Neurosci. 2018, 13, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Nasser, N.S.; Sharifat, H.; Rashid, A.A.; Ab Hamid, S.; Rahim, E.A.; Loh, J.L.; Ching, S.M.; Hoo, F.K.; Ismail, S.I.F.; Tyagi, R.; et al. Cue-Reactivity Among Young Adults With Problematic Instagram Use in Response to Instagram-Themed Risky Behavior Cues: A Pilot fMRI Study. Front. Psychol. 2020, 11, 556060. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, S.J.; Ketcherside, A.; McQueeny, T.M.; Dunlop, J.P.; Filbey, F.M. The hyper-sentient addict: An exteroception model of addiction. Am. J. Drug Alcohol Abus. 2015, 41, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Wikman, P.; Moisala, M.; Ylinen, A.; Lindblom, J.; Leikas, S.; Salmela-Aro, K.; Lonka, K.; Güroğlu, B.; Alho, K. Brain Responses to Peer Feedback in Social Media Are Modulated by Valence in Late Adolescence. Front. Behav. Neurosci. 2022, 16, 790478. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Zhou, H.; Gong, L.Y.; Teng, B.Y.; Geng, F.J.; Hu, Y.Z. Viewing personalized video clips recommended by TikTok activates default mode network and ventral tegmental area. Neuroimage 2021, 237, 118136. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Zhou, H.; Wang, C.; Geng, F.; Hu, Y. Individualized video recommendation modulates functional connectivity between large scale networks. Hum. Brain Mapp. 2021, 42, 5288–5299. [Google Scholar] [CrossRef] [PubMed]
- Leménager, T.; Dieter, J.; Hill, H.; Hoffmann, S.; Reinhard, I.; Beutel, M.; Vollstädt-Klein, S.; Kiefer, F.; Mann, K. Exploring the Neural Basis of Avatar Identification in Pathological Internet Gamers and of Self-Reflection in Pathological Social Network Users. J. Behav. Addict. 2016, 5, 485–499. [Google Scholar] [CrossRef]
- Peters, S.; Van der Cruijsen, R.; van der Aar, L.; Spaans, J.; Becht, A.; Crone, E. Social media use and the not-so-imaginary audience: Behavioral and neural mechanisms underlying the influence on self-concept. Dev. Cogn. Neurosci. 2021, 48, 100921. [Google Scholar] [CrossRef] [PubMed]
- Wölfling, K.; Beutel, M.; Müller, K. Construction of a standardized clinical interview to assess internet addiction: First findings regarding the usefulness of AICA-C. J. Addict. Res. Ther. 2012, 6, 003. [Google Scholar]
- Ellison, N.B.; Steinfield, C.; Lampe, C. The benefits of Facebook “friends:” Social capital and college students’ use of online social network sites. J. Comput.-Mediat. Commun. 2007, 12, 1143–1168. [Google Scholar] [CrossRef]
- Young, K.S. Caught in the Net: How to Recognize the Signs of Internet Addiction—And a Winning Strategy for Recovery; John Wiley & Sons: Hoboken, NJ, USA, 1998. [Google Scholar]
- Gao, X.; Zhang, M.; Yang, Z.; Wen, M.; Huang, H.; Zheng, R.; Wang, W.; Wei, Y.; Cheng, J.; Han, S.; et al. Structural and Functional Brain Abnormalities in Internet Gaming Disorder and Attention-Deficit/Hyperactivity Disorder: A Comparative Meta-Analysis. Front. Psychiatry 2021, 12, 679437. [Google Scholar] [CrossRef]
- Salgado, S.; Kaplitt, M.G. The nucleus accumbens: A comprehensive review. Stereotact. Funct. Neurosurg. 2015, 93, 75–93. [Google Scholar] [CrossRef] [PubMed]
- David, S.P.; Munafò, M.R.; Johansen-Berg, H.; Smith, S.M.; Rogers, R.D.; Matthews, P.M.; Walton, R.T. Ventral striatum/nucleus accumbens activation to smoking-related pictorial cues in smokers and nonsmokers: A functional magnetic resonance imaging study. Biol. Psychiatry 2005, 58, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Kühn, S.; Gallinat, J. Common biology of craving across legal and illegal drugs—A quantitative meta-analysis of cue-reactivity brain response. Eur. J. Neurosci. 2011, 33, 1318–1326. [Google Scholar] [CrossRef]
- Makris, N.; Oscar-Berman, M.; Jaffin, S.K.; Hodge, S.M.; Kennedy, D.N.; Caviness, V.S.; Marinkovic, K.; Breiter, H.C.; Gasic, G.P.; Harris, G.J. Decreased volume of the brain reward system in alcoholism. Biol. Psychiatry 2008, 64, 192–202. [Google Scholar] [CrossRef]
- Seifert, C.L.; Magon, S.; Sprenger, T.; Lang, U.E.; Huber, C.G.; Denier, N.; Vogel, M.; Schmidt, A.; Radue, E.-W.; Borgwardt, S. Reduced volume of the nucleus accumbens in heroin addiction. Eur. Arch. Psychiatry Clin. Neurosci. 2015, 265, 637–645. [Google Scholar] [CrossRef]
- Siddiqui, S.V.; Chatterjee, U.; Kumar, D.; Siddiqui, A.; Goyal, N. Neuropsychology of prefrontal cortex. Indian J. Psychiatry 2008, 50, 202. [Google Scholar]
- Kringelbach, M.L. The human orbitofrontal cortex: Linking reward to hedonic experience. Nat. Rev. Neurosci. 2005, 6, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, R.Z.; Volkow, N.D. Dysfunction of the prefrontal cortex in addiction: Neuroimaging findings and clinical implications. Nat. Rev. Neurosci. 2011, 12, 652–669. [Google Scholar] [CrossRef] [PubMed]
- Crunelle, C.L.; Kaag, A.M.; Van Wingen, G.; van den Munkhof, H.E.; Homberg, J.R.; Reneman, L.; Van Den Brink, W. Reduced frontal brain volume in non-treatment-seeking cocaine-dependent individuals: Exploring the role of impulsivity, depression, and smoking. Front. Hum. Neurosci. 2014, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Matochik, J.A.; Cadet, J.-L.; London, E.D. Smaller volume of prefrontal lobe in polysubstance abusers: A magnetic resonance imaging study. Neuropsychopharmacology 1998, 18, 243–252. [Google Scholar] [CrossRef]
- Pfefferbaum, A.; Sullivan, E.V.; Rosenbloom, M.J.; Mathalon, D.H.; Lim, K.O. A controlled study of cortical gray matter and ventricular changes in alcoholic men over a 5-year interval. Arch. Gen. Psychiatry 1998, 55, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.E.; Odlaug, B.L.; Chamberlain, S.R. Reduced cortical thickness in gambling disorder: A morphometric MRI study. Eur. Arch. Psychiatry Clin. Neurosci. 2015, 265, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Koehler, S.; Hasselmann, E.; Wüstenberg, T.; Heinz, A.; Romanczuk-Seiferth, N. Higher volume of ventral striatum and right prefrontal cortex in pathological gambling. Brain Struct. Funct. 2015, 220, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F. Brain stress systems in the amygdala and addiction. Brain Res. 2009, 1293, 61–75. [Google Scholar] [CrossRef]
- Chase, H.W.; Eickhoff, S.B.; Laird, A.R.; Hogarth, L. The neural basis of drug stimulus processing and craving: An activation likelihood estimation meta-analysis. Biol. Psychiatry 2011, 70, 785–793. [Google Scholar] [CrossRef]
- Warlow, S.M.; Robinson, M.J.; Berridge, K.C. Optogenetic central amygdala stimulation intensifies and narrows motivation for cocaine. J. Neurosci. 2017, 37, 8330–8348. [Google Scholar] [CrossRef] [PubMed]
- Gilman, J.M.; Kuster, J.K.; Lee, S.; Lee, M.J.; Kim, B.W.; Makris, N.; van der Kouwe, A.; Blood, A.J.; Breiter, H.C. Cannabis use is quantitatively associated with nucleus accumbens and amygdala abnormalities in young adult recreational users. J. Neurosci. 2014, 34, 5529–5538. [Google Scholar] [CrossRef] [PubMed]
- Padula, C.B.; McQueeny, T.; Lisdahl, K.M.; Price, J.S.; Tapert, S.F. Craving is associated with amygdala volumes in adolescent marijuana users during abstinence. Am. J. Drug Alcohol Abus. 2015, 41, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Wrase, J.; Makris, N.; Braus, D.F.; Mann, K.; Smolka, M.N.; Kennedy, D.N.; Caviness, V.S.; Hodge, S.M.; Tang, L.; Albaugh, M. Amygdala volume associated with alcohol abuse relapse and craving. Am. J. Psychiatry 2008, 165, 1179–1184. [Google Scholar] [CrossRef]
- Zakiniaeiz, Y.; Scheinost, D.; Seo, D.; Sinha, R.; Constable, R.T. Cingulate cortex functional connectivity predicts future relapse in alcohol dependent individuals. NeuroImage Clin. 2017, 13, 181–187. [Google Scholar] [CrossRef]
- Zhao, Y.; Sallie, S.N.; Cui, H.; Zeng, N.; Du, J.; Yuan, T.; Li, D.; De Ridder, D.; Zhang, C. Anterior cingulate cortex in addiction: New insights for neuromodulation. Neuromodul. Technol. Neural Interface 2021, 24, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Fritz, H.-C.; Wittfeld, K.; Schmidt, C.O.; Domin, M.; Grabe, H.J.; Hegenscheid, K.; Hosten, N.; Lotze, M. Current smoking and reduced gray matter volume—A voxel-based morphometry study. Neuropsychopharmacology 2014, 39, 2594–2600. [Google Scholar] [CrossRef]
- Yang, X.; Tian, F.; Zhang, H.; Zeng, J.; Chen, T.; Wang, S.; Jia, Z.; Gong, Q. Cortical and subcortical gray matter shrinkage in alcohol-use disorders: A voxel-based meta-analysis. Neurosci. Biobehav. Rev. 2016, 66, 92–103. [Google Scholar] [CrossRef]
- Wang, H.; Jin, C.; Yuan, K.; Shakir, T.M.; Mao, C.; Niu, X.; Niu, C.; Guo, L.; Zhang, M. The alteration of gray matter volume and cognitive control in adolescents with internet gaming disorder. Front. Behav. Neurosci. 2015, 9, 64. [Google Scholar] [CrossRef]
- Luijten, M.; Machielsen, M.W.; Veltman, D.J.; Hester, R.; de Haan, L.; Franken, I.H. Systematic review of ERP and fMRI studies investigating inhibitory control and error processing in people with substance dependence and behavioural addictions. J. Psychiatry Neurosci. 2014, 39, 149–169. [Google Scholar] [CrossRef]
- Leong, S.L.; Glue, P.; Manning, P.; Vanneste, S.; Lim, L.J.; Mohan, A.; De Ridder, D. Anterior cingulate cortex implants for alcohol addiction: A feasibility study. Neurotherapeutics 2020, 17, 1287–1299. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. The cingulate cortex and limbic systems for emotion, action, and memory. Brain Struct. Funct. 2019, 224, 3001–3018. [Google Scholar] [CrossRef] [PubMed]
- Ames, S.L.; Wong, S.W.; Bechara, A.; Cappelli, C.; Dust, M.; Grenard, J.L.; Stacy, A.W. Neural correlates of a Go/NoGo task with alcohol stimuli in light and heavy young drinkers. Behav. Brain Res. 2014, 274, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Van Holst, R.J.; van Holstein, M.; van Den Brink, W.; Veltman, D.J.; Goudriaan, A.E. Response inhibition during cue reactivity in problem gamblers: An fMRI study. PLoS ONE 2012, 7, e30909. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E. The brain’s default mode network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef]
- Zhang, R.; Volkow, N.D. Brain default-mode network dysfunction in addiction. Neuroimage 2019, 200, 313–331. [Google Scholar] [CrossRef]
- Cavanna, A.E.; Trimble, M.R. The precuneus: A review of its functional anatomy and behavioural correlates. Brain 2006, 129, 564–583. [Google Scholar] [CrossRef]
- Schurz, M.; Radua, J.; Aichhorn, M.; Richlan, F.; Perner, J. Fractionating theory of mind: A meta-analysis of functional brain imaging studies. Neurosci. Biobehav. Rev. 2014, 42, 9–34. [Google Scholar] [CrossRef]
- Schacht, J.P.; Anton, R.F.; Myrick, H. Functional neuroimaging studies of alcohol cue reactivity: A quantitative meta-analysis and systematic review. Addict. Biol. 2013, 18, 121–133. [Google Scholar] [CrossRef]
- Starcke, K.; Antons, S.; Trotzke, P.; Brand, M. Cue-reactivity in behavioral addictions: A meta-analysis and methodological considerations. J. Behav. Addict. 2018, 7, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Frith, C.D.; Frith, U. The neural basis of mentalizing. Neuron 2006, 50, 531–534. [Google Scholar] [CrossRef]
- Dean, S.F.; Fede, S.J.; Diazgranados, N.; Momenan, R. Addiction neurocircuitry and negative affect: A role for neuroticism in understanding amygdala connectivity and alcohol use disorder. Neurosci. Lett. 2020, 722, 134773. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Ma, Y.; Zhuang, K.; Chen, Q.; Shi, B.; Qiu, J. Linking temporal-parietal junction to internet addiction tendency: Moderating effect of critical thinking. J. Behav. Addict. 2021, 10, 759–766. [Google Scholar] [CrossRef]
- Gennatas, E.D.; Avants, B.B.; Wolf, D.H.; Satterthwaite, T.D.; Ruparel, K.; Ciric, R.; Hakonarson, H.; Gur, R.E.; Gur, R.C. Age-related effects and sex differences in gray matter density, volume, mass, and cortical thickness from childhood to young adulthood. J. Neurosci. 2017, 37, 5065–5073. [Google Scholar] [CrossRef]
- Pehlivanova, M.; Wolf, D.H.; Sotiras, A.; Kaczkurkin, A.N.; Moore, T.M.; Ciric, R.; Cook, P.A.; de La Garza, A.G.; Rosen, A.F.; Ruparel, K. Diminished cortical thickness is associated with impulsive choice in adolescence. J. Neurosci. 2018, 38, 2471–2481. [Google Scholar] [CrossRef] [PubMed]
- Gardini, S.; Venneri, A. Reduced grey matter in the posterior insula as a structural vulnerability or diathesis to addiction. Brain Res. Bull. 2012, 87, 205–211. [Google Scholar] [CrossRef]
- Naqvi, N.H.; Rudrauf, D.; Damasio, H.; Bechara, A. Damage to the insula disrupts addiction to cigarette smoking. Science 2007, 315, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Janes, A.; Gilman, J.; Radoman, M.; Pachas, G.; Fava, M.; Evins, A. Revisiting the role of the insula and smoking cue-reactivity in relapse: A replication and extension of neuroimaging findings. Drug Alcohol Depend. 2017, 179, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef]
- Brand, M.; Rumpf, H.-J.; King, D.L.; Potenza, M.N.; Wegmann, E. Clarifying terminologies in research on gaming disorder and other addictive behaviors: Distinctions between core symptoms and underlying psychological processes. Curr. Opin. Psychol. 2020, 36, 49–54. [Google Scholar] [CrossRef]
- Kuss, D.J.; Griffiths, M.D.; Pontes, H.M. Chaos and confusion in DSM-5 diagnosis of Internet Gaming Disorder: Issues, concerns, and recommendations for clarity in the field. J. Behav. Addict. 2017, 6, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Burnell, K.; George, M.J.; Kurup, A.R.; Underwood, M.K.; Ackerman, R.A. Associations between self-reports and device-reports of social networking site use: An application of the truth and bias model. Commun. Methods Meas. 2021, 15, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Ernala, S.K.; Burke, M.; Leavitt, A.; Ellison, N.B. How well do people report time spent on Facebook? An evaluation of established survey questions with recommendations. In Proceedings of the 2020 CHI Conference on Human Factors in Computing Systems, Honolulu, HI, USA, 25–30 April 2020; pp. 1–14. [Google Scholar]
- Shannon, H.; Bush, K.; Villeneuve, P.J.; Hellemans, K.G.; Guimond, S. Problematic Social Media Use in Adolescents and Young Adults: Systematic Review and Meta-analysis. JMIR Ment. Health 2022, 9, e33450. [Google Scholar] [CrossRef] [PubMed]
- Moccia, L.; Pettorruso, M.; De Crescenzo, F.; De Risio, L.; Di Nuzzo, L.; Martinotti, G.; Bifone, A.; Janiri, L.; Di Nicola, M. Neural correlates of cognitive control in gambling disorder: A systematic review of fMRI studies. Neurosci. Biobehav. Rev. 2017, 78, 104–116. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wadsley, M.; Ihssen, N. A Systematic Review of Structural and Functional MRI Studies Investigating Social Networking Site Use. Brain Sci. 2023, 13, 787. https://doi.org/10.3390/brainsci13050787
Wadsley M, Ihssen N. A Systematic Review of Structural and Functional MRI Studies Investigating Social Networking Site Use. Brain Sciences. 2023; 13(5):787. https://doi.org/10.3390/brainsci13050787
Chicago/Turabian StyleWadsley, Michael, and Niklas Ihssen. 2023. "A Systematic Review of Structural and Functional MRI Studies Investigating Social Networking Site Use" Brain Sciences 13, no. 5: 787. https://doi.org/10.3390/brainsci13050787
APA StyleWadsley, M., & Ihssen, N. (2023). A Systematic Review of Structural and Functional MRI Studies Investigating Social Networking Site Use. Brain Sciences, 13(5), 787. https://doi.org/10.3390/brainsci13050787