
Sex Differences in Excitatory and Inhibitory Function in the Primary Somatosensory Cortex during the Early Follicular Phase: A Preliminary Study

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. SEP and PPI Recording
2.3. Analyses
3. Results
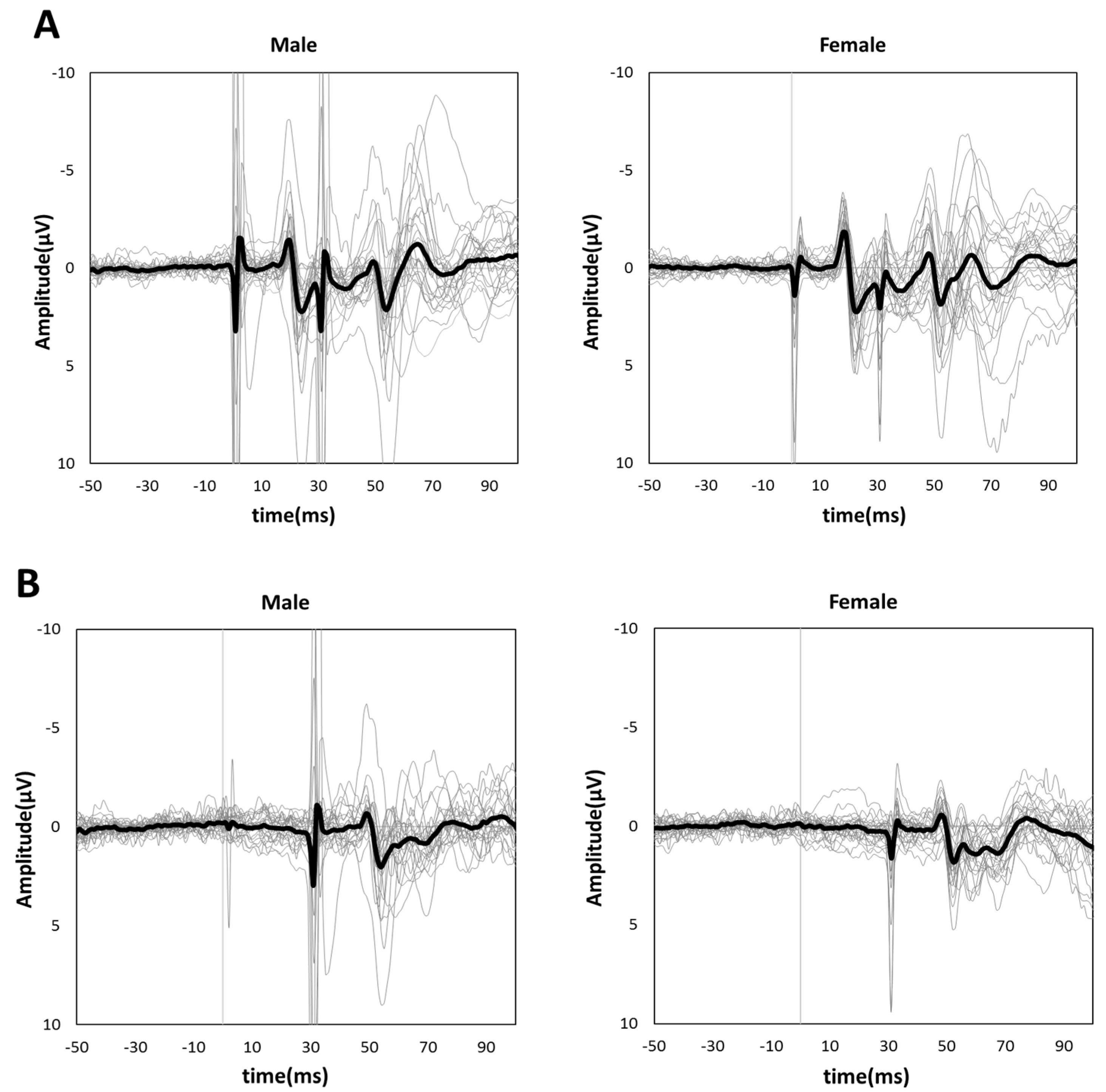
3.1. Single-Pulse SEP Amplitudes of N20 and P25
3.2. PPI-30 ms and 100 ms
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pletzer, B.; Harris, T.; Hidalgo-Lopez, E. Subcortical structural changes along the menstrual cycle: Beyond the hippocampus. Sci. Rep. 2018, 8, 16042. [Google Scholar] [CrossRef] [PubMed]
- al-Dahan, M.I.; Jalilian Tehrani, M.H.; Thalmann, R.H. Regulation of gamma-aminobutyric acidB (GABAB) receptors in cerebral cortex during the estrous cycle. Brain Res. 1994, 640, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, K.; Iguchi, K.; Yamazaki, Y.; Yamashiro, K.; Baba, Y.; Sato, D. Influence of menstrual cycle phases on neural excitability in the primary somatosensory cortex and ankle joint position sense. Womens Health Rep. New Rochelle 2020, 1, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Schloemer, N.; Lenz, M.; Tegenthoff, M.; Dinse, H.R.; Hoffken, O. Parallel modulation of intracortical excitability of somatosensory and visual cortex by the gonadal hormones estradiol and progesterone. Sci. Rep. 2020, 10, 22237. [Google Scholar] [CrossRef] [PubMed]
- Allison, T.; McCarthy, G.; Wood, C.C.; Darcey, T.M.; Spencer, D.D.; Williamson, P.D. Human cortical potentials evoked by stimulation of the median nerve. I. Cytoarchitectonic areas generating short-latency activity. J. Neurophysiol. 1989, 62, 694–710. [Google Scholar] [CrossRef]
- Buchner, H.; Fuchs, M.; Wischmann, H.A.; Dössel, O.; Ludwig, I.; Knepper, A.; Berg, P. Source analysis of median nerve and finger stimulated somatosensory evoked potentials: Multichannel simultaneous recording of electric and magnetic fields combined with 3D-MR tomography. Brain Topogr. 1994, 6, 299–310. [Google Scholar] [CrossRef]
- Desmedt, J.E.; Nguyen, T.H.; Bourguet, M. Bit-mapped color imaging of human evoked potentials with reference to the N20, P22, P27 and N30 somatosensory responses. Electroencephalogr. Clin. Neurophysiol. 1987, 68, 1–19. [Google Scholar] [CrossRef]
- Desmedt, J.E.; Huy, N.T.; Bourguet, M. The cognitive P40, N60 and P100 components of somatosensory evoked potentials and the earliest electrical signs of sensory processing in man. Electroencephalogr. Clin. Neurophysiol. 1983, 56, 272–282. [Google Scholar] [CrossRef]
- Tomberg, C.; Desmedt, J.E.; Ozaki, I.; Nguyen, T.H.; Chalklin, V. Mapping somatosensory evoked potentials to finger stimulation at intervals of 450 to 4000 msec and the issue of habituation when assessing early cognitive components. Electroencephalogr. Clin. Neurophysiol. 1989, 74, 347–358. [Google Scholar] [CrossRef]
- Lenz, M.; Tegenthoff, M.; Kohlhaas, K.; Stude, P.; Hoffken, O.; Gatica Tossi, M.A.; Kalisch, T.; Kowalewski, R.; Dinse, H.R. Increased excitability of somatosensory cortex in aged humans is associated with impaired tactile acuity. J. Neurosci. 2012, 32, 1811–1816. [Google Scholar] [CrossRef]
- Rocchi, L.; Casula, E.; Tocco, P.; Berardelli, A.; Rothwell, J. Somatosensory temporal discrimination threshold involves inhibitory mechanisms in the primary somatosensory area. J. Neurosci. 2016, 36, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Yamashiro, K.; Onishi, H.; Otsuru, N.; Kojima, S.; Saito, K.; Sato, D. Modulation of inhibitory function in the primary somatosensory cortex and temporal discrimination threshold induced by acute aerobic exercise. Behav. Brain Res. 2020, 377, 112253. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Lin, Y.Y. Aging-related decline in somatosensory inhibition of the human cerebral cortex. Exp. Brain Res. 2013, 226, 145–152. [Google Scholar] [CrossRef]
- Vidoni, E.D.; Acerra, N.E.; Dao, E.; Meehan, S.K.; Boyd, L.A. Role of the primary somatosensory cortex in motor learning: An rTMS study. Neurobiol. Learn. Mem. 2010, 93, 532–539. [Google Scholar] [CrossRef]
- Ikuta, T.; Furuta, N. Sex differences in the human group mean SEP. Electroencephalogr. Clin. Neurophysiol. 1982, 54, 449–457. [Google Scholar] [CrossRef]
- Kakigi, R.; Shibasaki, H. Effects of age, gender, and stimulus side on scalp topography of somatosensory evoked potentials following median nerve stimulation. J. Clin. Neurophysiol. 1991, 8, 320–330. [Google Scholar] [CrossRef]
- Huttunen, J.; Wikström, H.; Salonen, O.; Ilmoniemi, R.J. Human somatosensory cortical activation strengths: Comparison between males and females and age-related changes. Brain Res. 1999, 818, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Edama, M.; Ohya, T.; Maruyama, S.; Shagawa, M.; Sekine, C.; Hirabayashi, R.; Yokota, H.; Ishigaki, T.; Akuzawa, H.; Togashi, R.; et al. Relationship between changes in foot arch and sex differences during the menstrual cycle. Int. J. Environ. Res. Public Health 2022, 20, 509. [Google Scholar] [CrossRef] [PubMed]
- Huttunen, J.; Pekkonen, E.; Kivisaari, R.; Autti, T.; Kahkonen, S. Modulation of somatosensory evoked fields from SI and SII by acute GABA A-agonism and paired-pulse stimulation. Neuroimage 2008, 40, 427–434. [Google Scholar] [CrossRef]
- Tamura, Y.; Matsuhashi, M.; Lin, P.; Ou, B.; Vorbach, S.; Kakigi, R.; Hallett, M. Impaired intracortical inhibition in the primary somatosensory cortex in focal hand dystonia. Mov. Disord. 2008, 23, 558–565. [Google Scholar] [CrossRef]
- Allison, T.; Wood, C.C.; Goff, W.R. Brain stem auditory, pattern-reversal visual, and short-latency somatosensory evoked potentials: Latencies in relation to age, sex, and brain and body size. Electroencephalogr. Clin. Neurophysiol. 1983, 55, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.D.; Cole, N.B.; Greenberger, V.; Segal, M. Estradiol increases dendritic spine density by reducing GABA neurotransmission in hippocampal neurons. J. Neurosci. 1998, 18, 2550–2559. [Google Scholar] [CrossRef] [PubMed]
- Hoffken, O.; Veit, M.; Knossalla, F.; Lissek, S.; Bliem, B.; Ragert, P.; Dinse, H.R.; Tegenthoff, M. Sustained increase of somatosensory cortex excitability by tactile coactivation studied by paired median nerve stimulation in humans correlates with perceptual gain. J. Physiol. 2007, 584, 463–471. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anazawa, S.; Yamashiro, K.; Makibuchi, T.; Ikarashi, K.; Fujimoto, T.; Ochi, G.; Sato, D. Sex Differences in Excitatory and Inhibitory Function in the Primary Somatosensory Cortex during the Early Follicular Phase: A Preliminary Study. Brain Sci. 2023, 13, 761. https://doi.org/10.3390/brainsci13050761
Anazawa S, Yamashiro K, Makibuchi T, Ikarashi K, Fujimoto T, Ochi G, Sato D. Sex Differences in Excitatory and Inhibitory Function in the Primary Somatosensory Cortex during the Early Follicular Phase: A Preliminary Study. Brain Sciences. 2023; 13(5):761. https://doi.org/10.3390/brainsci13050761
Chicago/Turabian StyleAnazawa, Sayaka, Koya Yamashiro, Taiki Makibuchi, Koyuki Ikarashi, Tomomi Fujimoto, Genta Ochi, and Daisuke Sato. 2023. "Sex Differences in Excitatory and Inhibitory Function in the Primary Somatosensory Cortex during the Early Follicular Phase: A Preliminary Study" Brain Sciences 13, no. 5: 761. https://doi.org/10.3390/brainsci13050761
APA StyleAnazawa, S., Yamashiro, K., Makibuchi, T., Ikarashi, K., Fujimoto, T., Ochi, G., & Sato, D. (2023). Sex Differences in Excitatory and Inhibitory Function in the Primary Somatosensory Cortex during the Early Follicular Phase: A Preliminary Study. Brain Sciences, 13(5), 761. https://doi.org/10.3390/brainsci13050761