Closed-Loop tACS Delivered during Slow-Wave Sleep Reduces Retroactive Interference on a Paired-Associates Learning Task

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Sleep and Memory Interference
1.2. Electrical Augmentation of Memory
1.3. Electrical Augmentation of Memory during Sleep
1.4. Slow-Wave Sleep
1.5. Aims and Hypotheses
2. Materials and Methods
2.1. Participants
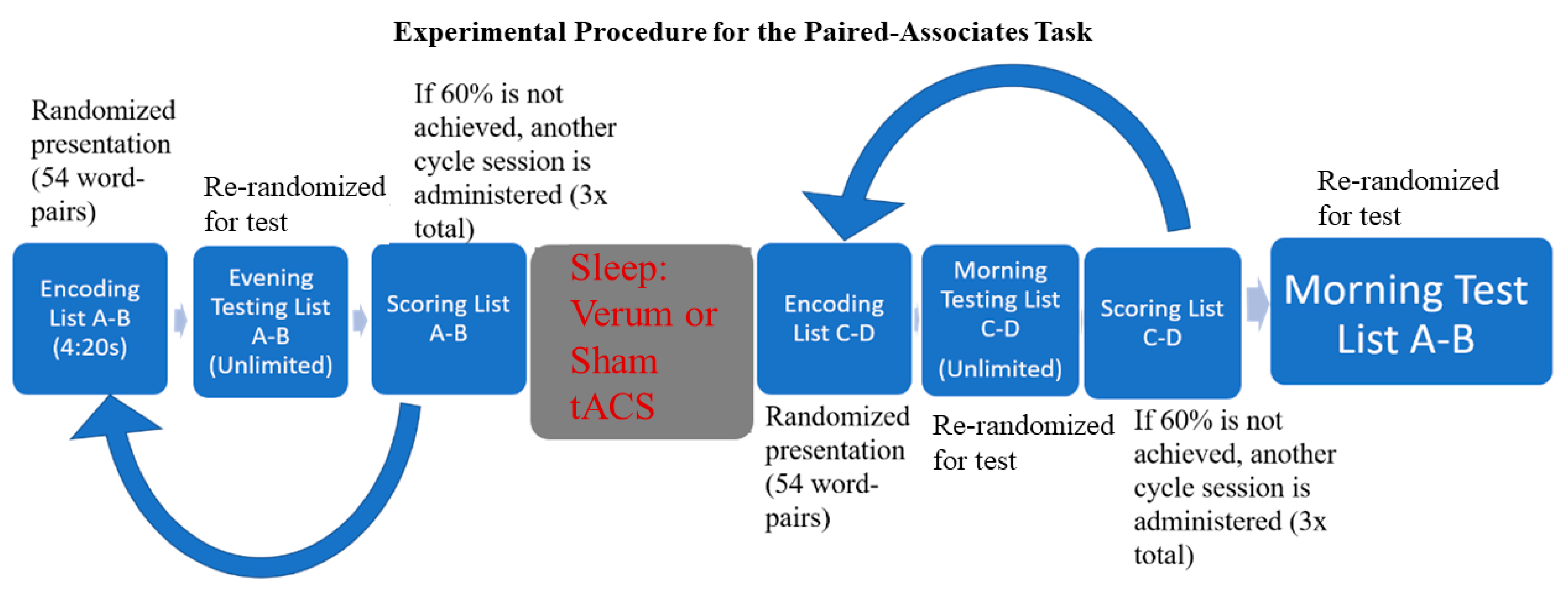
2.2. Procedures
2.3. Tasks/Materials
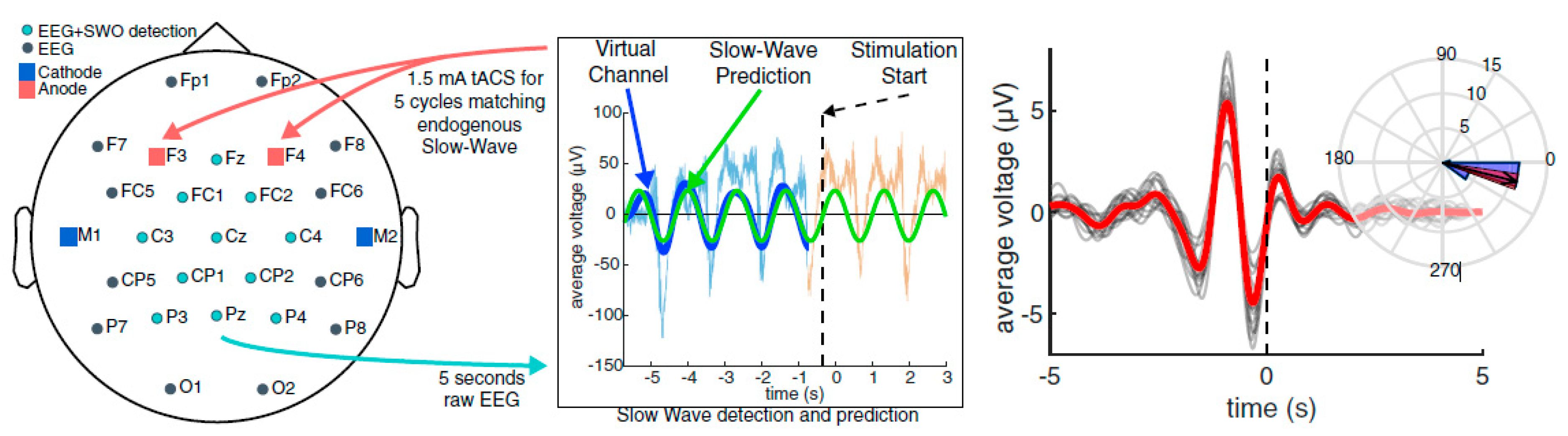
2.4. Polysomnographic (PSG) Data Collection/Closed-Loop Transcranial Alternating Current Stimulation (CL-tACS) during Slow-Wave Oscillations
2.5. Statistical Analysis
3. Results
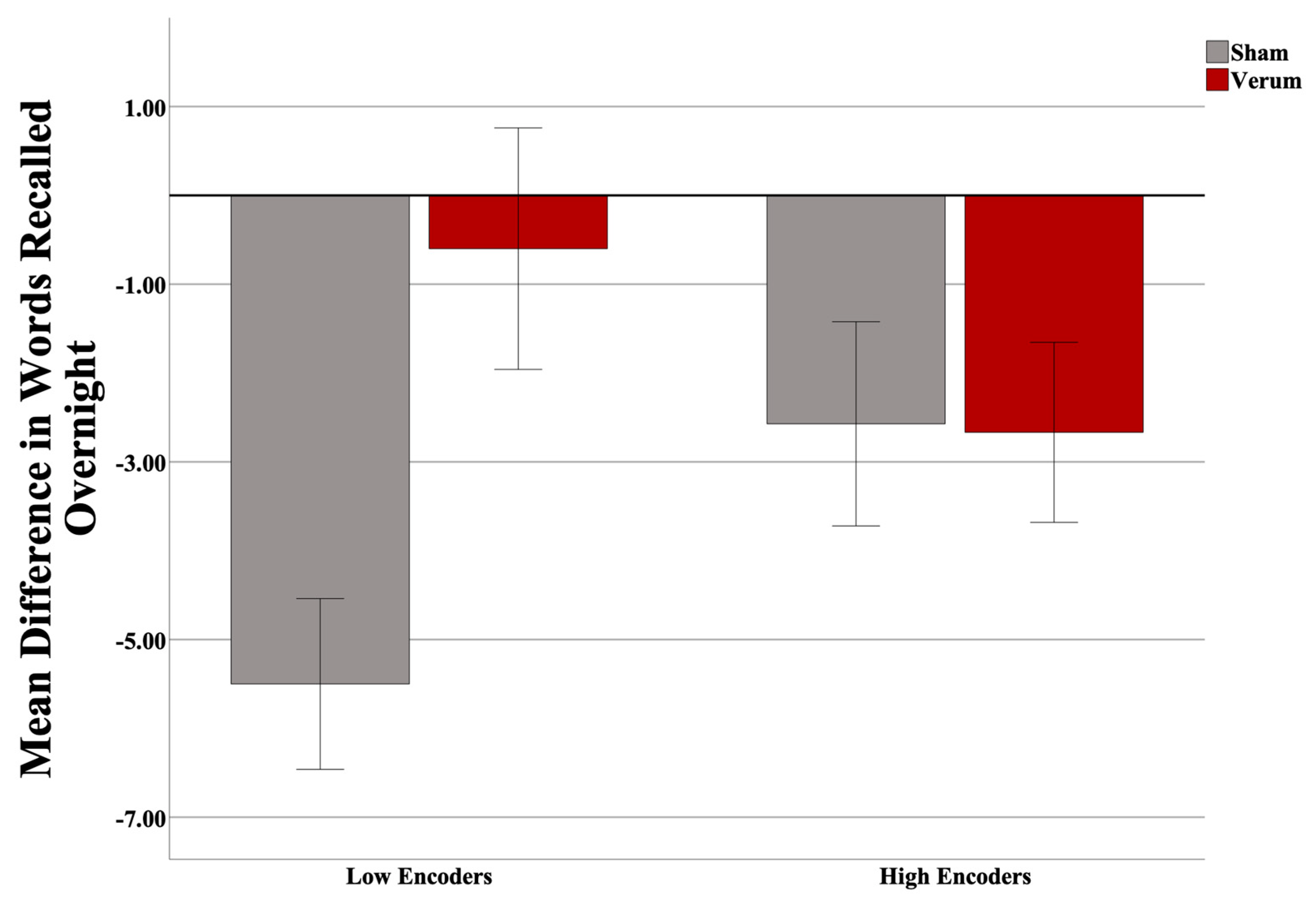
3.1. Effects of CL-tACS and Interactions with Encoding Strength on Learning
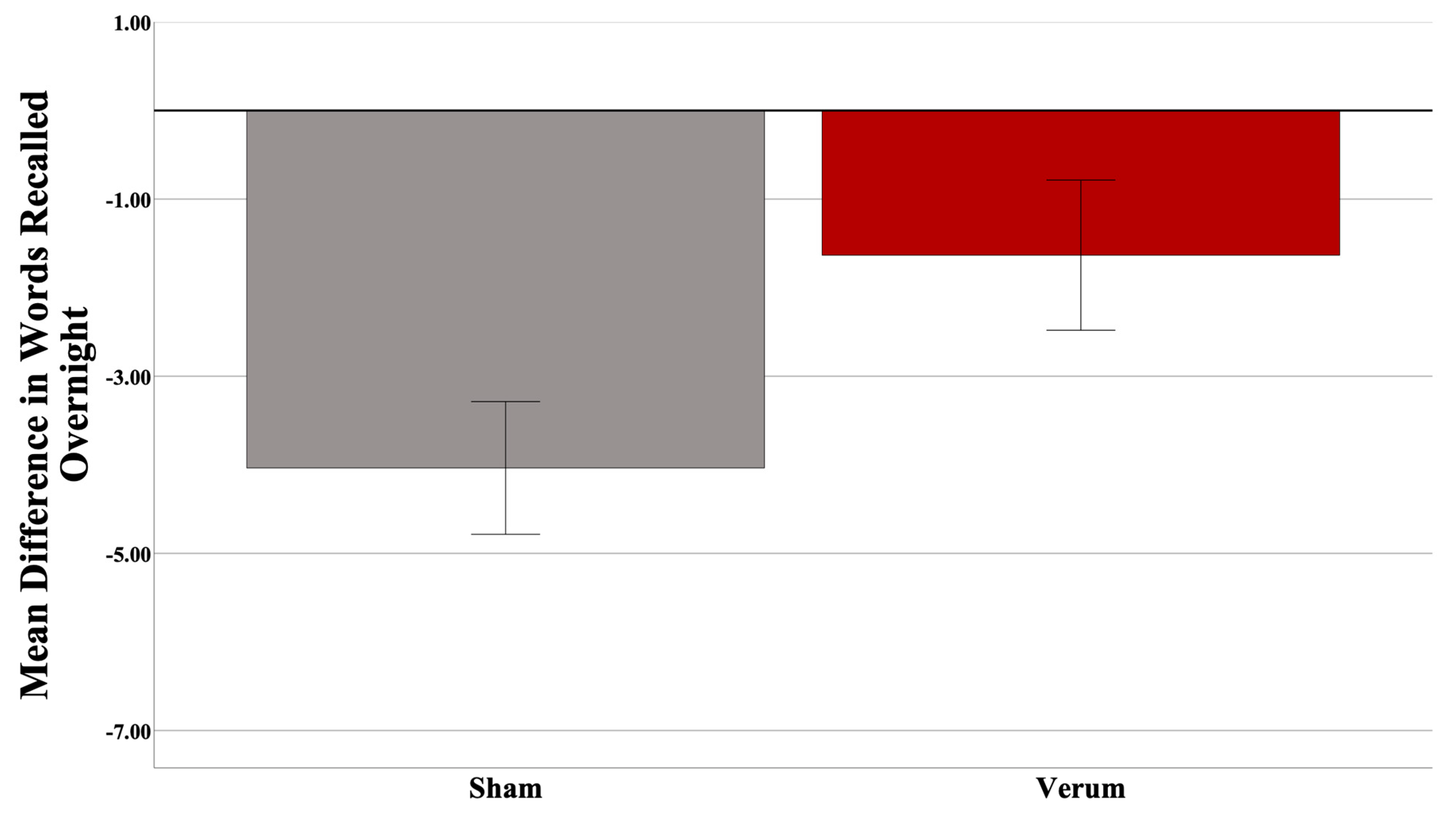
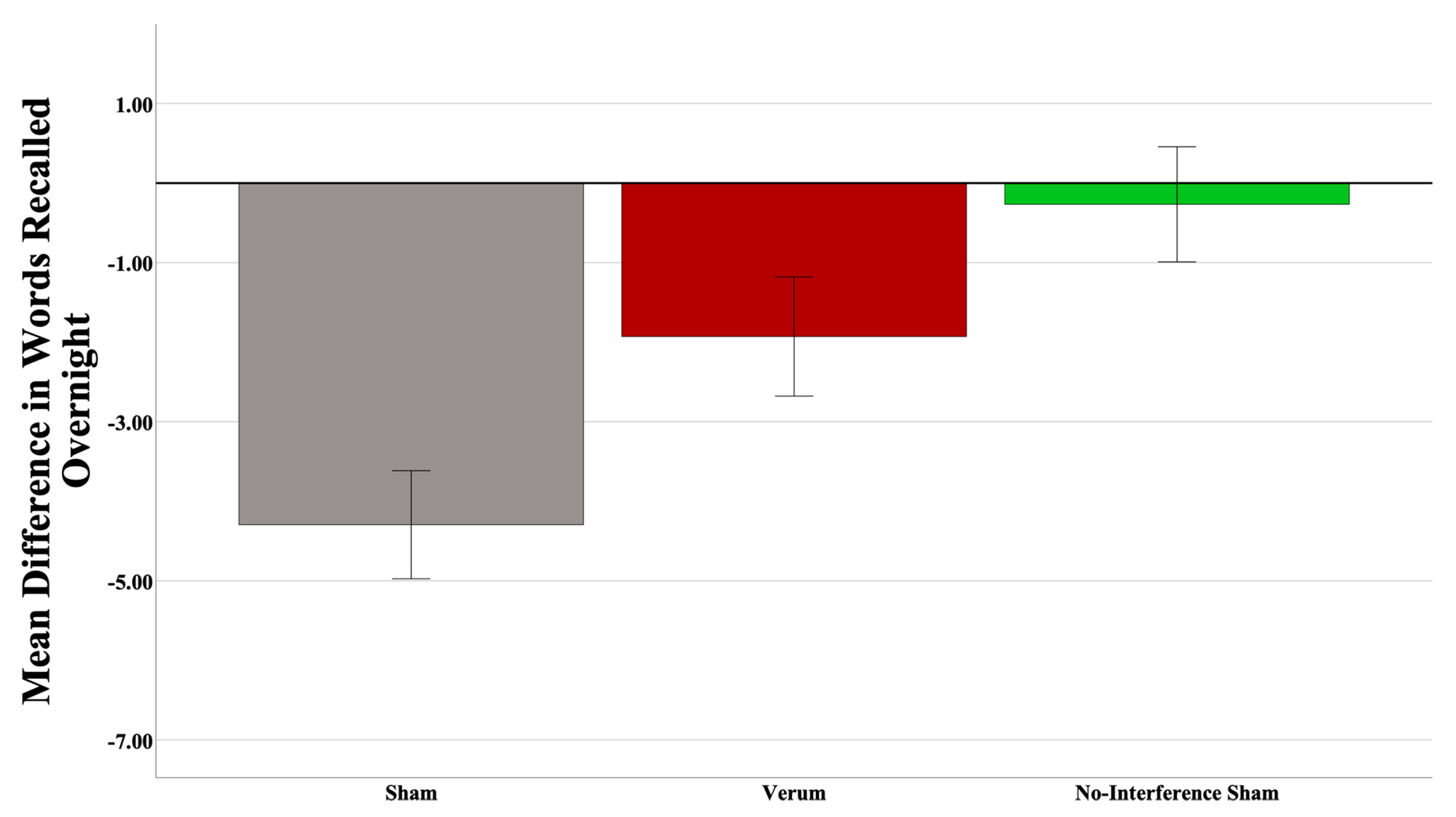
3.2. Effects of CL-tACS on Interference
4. Discussion
4.1. Limitations
4.2. Future Directions
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sejnowski, T.J.; Destexhe, A. Why do we sleep? Brain Res. 2000, 886, 208–223. [Google Scholar] [CrossRef]
- Tononi, G.; Riedner, B.A.; Hulse, B.K.; Ferrarelli, F.; Sarasso, S. Enhancing sleep slow waves with natural stimuli. Medicamundi 2010, 54, 73–79. [Google Scholar]
- Abel, T.; Havekes, R.; Saletin, J.M.; Walker, M.P. Sleep, Plasticity and Memory from Molecules to Whole-Brain Networks. Curr. Biol. 2013, 23, R774–R788. [Google Scholar] [CrossRef]
- Pace-Schott, E.F.; Hobson, J.A. The Neurobiology of Sleep: Genetics, cellular physiology and subcortical networks. Nat. Rev. Neurosci. 2002, 3, 591–605. [Google Scholar] [CrossRef]
- Borbély, A.A.; Daan, S.; Wirz-Justice, A.; Deboer, T. The two-process model of sleep regulation: A reappraisal. J. Sleep Res. 2016, 25, 131–143. [Google Scholar] [CrossRef]
- Marshall, L.; Mölle, M.; Hallschmid, M.; Born, J. Transcranial Direct Current Stimulation during Sleep Improves Declarative Memory. J. Neurosci. 2004, 24, 9985–9992. [Google Scholar] [CrossRef]
- Walker, M.P.; Stickgold, R. Sleep, Memory, and Plasticity. Annu. Rev. Psychol. 2006, 57, 139–166. [Google Scholar] [CrossRef]
- Diekelmann, S.; Born, J. The memory function of sleep. Nat. Rev. Neurosci. 2010, 11, 114–126. [Google Scholar] [CrossRef]
- Rasch, B.; Born, J. About Sleep’s Role in Memory. Physiol. Rev. 2013, 93, 681–766. [Google Scholar] [CrossRef]
- Paller, K.A.; Voss, J.L. Memory reactivation and consolidation during sleep. Learn. Mem. 2004, 11, 664–670. [Google Scholar] [CrossRef]
- Plihal, W.; Born, J. Effects of Early and Late Nocturnal Sleep on Declarative and Procedural Memory. J. Cogn. Neurosci. 1997, 9, 534–547. [Google Scholar] [CrossRef]
- Marshall, L.; Born, J. The contribution of sleep to hippocampus-dependent memory consolidation. Trends Cogn. Sci. 2007, 11, 442–450. [Google Scholar] [CrossRef]
- Norman, K.A. Declarative Memory: Sleep Protects New Memories from Interference. Curr. Biol. 2006, 16, R596–R597. [Google Scholar] [CrossRef]
- Alger, S.E.; Lau, H.; Fishbein, W. Slow wave sleep during a daytime nap is necessary for protection from subsequent interference and long-term retention. Neurobiol. Learn. Mem. 2012, 98, 188–196. [Google Scholar] [CrossRef]
- Ekstrand, B.R. Effect of sleep on memory. J. Exp. Psychol. 1967, 75, 64. [Google Scholar] [CrossRef]
- Sheth, B.R.; Varghese, R.; Truong, T. Sleep Shelters Verbal Memory from Different Kinds of Interference. Sleep 2012, 35, 985–996. [Google Scholar] [CrossRef][Green Version]
- Ellenbogen, J.M.; Hulbert, J.C.; Stickgold, R.; Dinges, D.F.; Thompson-Schill, S.L. Interfering with Theories of Sleep and Memory: Sleep, Declarative Memory, and Associative Interference. Curr. Biol. 2006, 16, 1290–1294. [Google Scholar] [CrossRef]
- Deliens, G.; Leproult, R.; Neu, D.; Peigneux, P. Rapid Eye Movement and Non-Rapid Eye Movement Sleep Contributions in Memory Consolidation and Resistance to Retroactive Interference for Verbal Material. Sleep 2013, 36, 1875–1883. [Google Scholar] [CrossRef][Green Version]
- Drosopoulos, S.; Schulze, C.; Fischer, S.; Born, J. Sleep’s function in the spontaneous recovery and consolidation of memories. J. Exp. Psychol. Gen. 2007, 136, 169–183. [Google Scholar] [CrossRef]
- Pöhlchen, D.; Pawlizki, A.; Gais, S.; Schönauer, M. Evidence against a large effect of sleep in protecting verbal memories from interference. J. Sleep Res. 2021, 30, e13042. [Google Scholar] [CrossRef]
- Soetens, E.; Hueting, J.E.; Casaer, S.; D’Hooge, D. Effect of amphetamine on long-term retention of verbal material. Psychopharmacology 1995, 119, 155–162. [Google Scholar] [CrossRef]
- Trumbo, M.; McDaniel, M.A.; Hodge, G.K.; Jones, A.P.; Matzen, L.E.; Kittinger, L.I.; Kittinger, R.S.; Clark, V.P. Is the testing effect ready to be put to work? Evidence from the laboratory to the classroom. Transl. Issues Psychol. Sci. 2021, 7, 332. [Google Scholar] [CrossRef]
- Litman, L.; Davachi, L. Distributed learning enhances relational memory consolidation. Learn. Mem. 2008, 15, 711–716. [Google Scholar] [CrossRef]
- Wamsley, E.J.; Tucker, M.; Payne, J.D.; Benavides, J.A.; Stickgold, R. Dreaming of a Learning Task Is Associated with Enhanced Sleep-Dependent Memory Consolidation. Curr. Biol. 2010, 20, 850–855. [Google Scholar] [CrossRef]
- Hoscheidt, S.M.; LaBar, K.S.; Ryan, L.; Jacobs, W.J.; Nadel, L. Encoding negative events under stress: High subjective arousal is related to accurate emotional memory despite misinformation exposure. Neurobiol. Learn. Mem. 2014, 112, 237–247. [Google Scholar] [CrossRef]
- Buzsáki, G.; Draguhn, A. Neuronal Oscillations in Cortical Networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef]
- Coffman, B.A.; Clark, V.P.; Parasuraman, R. Battery powered thought: Enhancement of attention, learning, and memory in healthy adults using transcranial direct current stimulation. NeuroImage 2014, 85 Pt 3, 895–908. [Google Scholar] [CrossRef]
- Sarmiento, C.I.; San-Juan, D.; Prasath, V.B.S. Letter to the Editor: Brief history of transcranial direct current stimulation (tDCS): From electric fishes to microcontrollers. Psychol. Med. 2016, 46, 3259–3261. [Google Scholar] [CrossRef]
- Mohammadi, A. Induction of Neuroplasticity by Transcranial Direct Current Stimulation. J. Biomed. Phys. Eng. 2016, 6, 36. Available online: http://www.jbpe.org/Journal_OJS/JBPE/index.php/jbpe/article/view/692 (accessed on 5 March 2023).
- Kuo, M.-F.; Nitsche, M.A. Effects of Transcranial Electrical Stimulation on Cognition. Clin. EEG Neurosci. 2012, 43, 192–199. [Google Scholar] [CrossRef]
- Fröhlich, F. Endogenous and exogenous electric fields as modifiers of brain activity: Rational design of noninvasive brain stimulation with transcranial alternating current stimulation. Dialogues Clin. Neurosci. 2014, 16, 93–102. [Google Scholar] [CrossRef]
- Hsu, W.-Y.; Ku, Y.; Zanto, T.P.; Gazzaley, A. Effects of noninvasive brain stimulation on cognitive function in healthy aging and Alzheimer’s disease: A systematic review and meta-analysis. Neurobiol. Aging 2015, 36, 2348–2359. [Google Scholar] [CrossRef]
- Ali, M.M.; Sellers, K.K.; Fröhlich, F. Transcranial Alternating Current Stimulation Modulates Large-Scale Cortical Network Activity by Network Resonance. J. Neurosci. 2013, 33, 11262–11275. [Google Scholar] [CrossRef]
- Merlet, I.; Birot, G.; Salvador, R.; Molaee-Ardekani, B.; Mekonnen, A.; Soria-Frish, A.; Ruffini, G.; Miranda, P.C.; Wendling, F. From Oscillatory Transcranial Current Stimulation to Scalp EEG Changes: A Biophysical and Physiological Modeling Study. PLoS ONE 2013, 8, e57330. [Google Scholar] [CrossRef]
- Herrmann, C.S.; Rach, S.; Neuling, T.; Strüber, D. Transcranial alternating current stimulation: A review of the underlying mechanisms and modulation of cognitive processes. Front. Hum. Neurosci. 2013, 7, 279. [Google Scholar] [CrossRef]
- Helfrich, R.F.; Schneider, T.R.; Rach, S.; Trautmann-Lengsfeld, S.A.; Engel, A.K.; Herrmann, C.S. Entrainment of Brain Oscillations by Transcranial Alternating Current Stimulation. Curr. Biol. 2014, 24, 333–339. [Google Scholar] [CrossRef]
- Živanović, M.; Bjekić, J.; Konstantinović, U.; Filipović, S.R. Effects of online parietal transcranial electric stimulation on associative memory: A direct comparison between tDCS, theta tACS, and theta-oscillatory tDCS. Sci. Rep. 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Wilde, C.; Bruder, R.; Binder, S.; Marshall, L.; Schweikard, A. Closed-loop transcranial alternating current stimulation of slow oscillations. Curr. Dir. Biomed. Eng. 2015, 1, 85–88. [Google Scholar] [CrossRef]
- Gais, S.; Born, J. Low acetylcholine during slow-wave sleep is critical for declarative memory consolidation. Proc. Natl. Acad. Sci. USA 2004, 101, 2140–2144. [Google Scholar] [CrossRef]
- Mednick, S.C.; McDevitt, E.A.; Walsh, J.K.; Wamsley, E.; Paulus, M.; Kanady, J.C.; Drummond, S.P.A. The Critical Role of Sleep Spindles in Hippocampal-Dependent Memory: A Pharmacology Study. J. Neurosci. 2013, 33, 4494–4504. [Google Scholar] [CrossRef]
- Cox, R.; Korjoukov, I.; de Boer, M.; Talamini, L.M. Sound Asleep: Processing and Retention of Slow Oscillation Phase-Targeted Stimuli. PLoS ONE 2014, 9, e101567. [Google Scholar] [CrossRef]
- Rudoy, J.D.; Voss, J.L.; Westerberg, C.E.; Paller, K.A. Strengthening Individual Memories by Reactivating Them During Sleep. Science 2009, 326, 1079. [Google Scholar] [CrossRef]
- Diekelmann, S.; Büchel, C.; Born, J.; Rasch, B. Labile or stable: Opposing consequences for memory when reactivated during waking and sleep. Nat. Neurosci. 2011, 14, 381–386. [Google Scholar] [CrossRef]
- Pilly, P.K.; Skorheim, S.W.; Hubbard, R.J.; Ketz, N.A.; Roach, S.M.; Lerner, I.; Jones, A.P.; Robert, B.; Bryant, N.B.; Hartholt, A.; et al. One-Shot Tagging During Wake and Cueing During Sleep With Spatiotemporal Patterns of Transcranial Electrical Stimulation Can Boost Long-Term Metamemory of Individual Episodes in Humans. Front. Neurosci. 2020, 13, 1416. [Google Scholar] [CrossRef]
- Hubbard, R.J.; Zadeh, I.; Jones, A.P.; Robert, B.; Bryant, N.B.; Clark, V.P.; Pilly, P.K. Brain connectivity alterations during sleep by closed-loop transcranial neurostimulation predict metamemory sensitivity. Netw. Neurosci. 2021, 5, 734–756. [Google Scholar] [CrossRef]
- Bayer, L.; Constantinescu, I.; Perrig, S.; Vienne, J.; Vidal, P.-P.; Mühlethaler, M.; Schwartz, S. Rocking synchronizes brain waves during a short nap. Curr. Biol. 2011, 21, R461–R462. [Google Scholar] [CrossRef]
- Marshall, L.; Helgadóttir, H.; Mölle, M.; Born, J. Boosting slow oscillations during sleep potentiates memory. Nature 2006, 444, 610–613. [Google Scholar] [CrossRef]
- Kirov, R.; Weiss, C.; Siebner, H.R.; Born, J.; Marshall, L. Slow oscillation electrical brain stimulation during waking promotes EEG theta activity and memory encoding. Proc. Natl. Acad. Sci. USA 2009, 106, 15460–15465. [Google Scholar] [CrossRef]
- Saebipour, M.R.; Joghataei, M.T.; Yoonessi, A.; Sadeghniiat-Haghighi, K.; Khalighinejad, N.; Khademi, S. Slow oscillating transcranial direct current stimulation during sleep has a sleep-stabilizing effect in chronic insomnia: A pilot study. J. Sleep Res. 2015, 24, 518–525. [Google Scholar] [CrossRef]
- Berényi, A.; Belluscio, M.; Mao, D.; Buzsáki, G. Closed-Loop Control of Epilepsy by Transcranial Electrical Stimulation. Science 2012, 337, 735–737. [Google Scholar] [CrossRef]
- Del Felice, A.; Magalini, A.; Masiero, S. Slow-oscillatory Transcranial Direct Current Stimulation Modulates Memory in Temporal Lobe Epilepsy by Altering Sleep Spindle Generators: A Possible Rehabilitation Tool. Brain Stimul. 2015, 8, 567–573. [Google Scholar] [CrossRef]
- Del Felice, A.; Magalini, A.; Masiero, S.; Manganotti, P. P65. Memory rehabilitation in temporal lobe epilepsy: Slow-oscillatory transcranial direct current stimulation modulates memory by altering sleep spindle generators. Clin. Neurophysiol. 2015, 126, e126–e127. [Google Scholar] [CrossRef]
- Prehn-Kristensen, A.; Munz, M.; Göder, R.; Wilhelm, I.; Korr, K.; Vahl, W.; Wiesner, C.D.; Baving, L. Transcranial Oscillatory Direct Current Stimulation During Sleep Improves Declarative Memory Consolidation in Children with Attention-deficit/hyperactivity Disorder to a Level Comparable to Healthy Controls. Brain Stimul. 2014, 7, 793–799. [Google Scholar] [CrossRef]
- Barham, M.P.; Enticott, P.G.; Conduit, R.; Lum, J.A.G. Transcranial electrical stimulation during sleep enhances declarative (but not procedural) memory consolidation: Evidence from a meta-analysis. Neurosci. Biobehav. Rev. 2016, 63, 65–77. [Google Scholar] [CrossRef]
- Massimini, M.; Ferrarelli, F.; Esser, S.K.; Riedner, B.A.; Huber, R.; Murphy, M.; Peterson, M.J.; Tononi, G. Triggering sleep slow waves by transcranial magnetic stimulation. Proc. Natl. Acad. Sci. USA 2007, 104, 8496–8501. [Google Scholar] [CrossRef]
- Marshall, L.; Kirov, R.; Brade, J.; Mölle, M.; Born, J. Transcranial Electrical Currents to Probe EEG Brain Rhythms and Memory Consolidation during Sleep in Humans. PLoS ONE 2011, 6, e16905. [Google Scholar] [CrossRef]
- Jones, A.P.; Choe, J.; Bryant, N.B.; Robinsons, C.S.H.; Ketz, N.A.; Skorheim, S.W.; Combs, A.; Lamphere, M.L.; Robert, B.; Gill, H.A.; et al. Dose-dependent effects of closed-loop tACS delivered during slow-wave oscillations on memory consolidation. Front. Neurosci. 2018, 12, 867. [Google Scholar] [CrossRef]
- Ketz, N.; Jones, A.P.; Bryant, N.B.; Clark, V.P.; Pilly, P.K. Closed-loop slow-wave tACS improves sleep-dependent long-term memory generalization by modulating endogenous oscillations. J. Neurosci. 2018, 38, 7314–7326. [Google Scholar] [CrossRef]
- Robinson, C.S.; Bryant, N.B.; Maxwell, J.W.; Jones, A.P.; Robert, B.; Lamphere, M.; Combs, A.; Al Azzawi, H.M.; Gibson, B.C.; Sanguinetti, J.L.; et al. The benefits of closed-loop transcranial alternating current stimulation on subjective sleep quality. Brain Sci. 2018, 8, 204. [Google Scholar] [CrossRef]
- Antonenko, D.; Diekelmann, S.; Olsen, C.; Born, J.; Mölle, M. Napping to renew learning capacity: Enhanced encoding after stimulation of sleep slow oscillations. Eur. J. Neurosci. 2013, 37, 1142–1151. [Google Scholar] [CrossRef]
- Westerberg, C.E.; Florczak, S.M.; Weintraub, S.; Mesulam, M.-M.; Marshall, L.; Zee, P.C.; Paller, K.A. Memory improvement via slow-oscillatory stimulation during sleep in older adults. Neurobiol. Aging 2015, 36, 2577–2586. [Google Scholar] [CrossRef]
- Ladenbauer, J.; Külzow, N.; Passmann, S.; Antonenko, D.; Grittner, U.; Tamm, S.; Flöel, A. Brain stimulation during an afternoon nap boosts slow oscillatory activity and memory consolidation in older adults. NeuroImage 2016, 142, 311–323. [Google Scholar] [CrossRef]
- Sahlem, G.L.; Badran, B.W.; Halford, J.J.; Williams, N.R.; Korte, J.E.; Leslie, K.; Strachan, M.; Breedlove, J.L.; Runion, J.; Bachman, D.L.; et al. Oscillating Square Wave Transcranial Direct Current Stimulation (tDCS) Delivered During Slow Wave Sleep Does Not Improve Declarative Memory More Than Sham: A Randomized Sham Controlled Crossover Study. Brain Stimul. 2015, 8, 528–534. [Google Scholar] [CrossRef]
- Eggert, T.; Dorn, H.; Sauter, C.; Nitsche, M.A.; Bajbouj, M.; Danker-Hopfe, H. No Effects of Slow Oscillatory Transcranial Direct Current Stimulation (tDCS) on Sleep-Dependent Memory Consolidation in Healthy Elderly Subjects. Brain Stimul. 2013, 6, 938–945. [Google Scholar] [CrossRef]
- Paßmann, S.; Külzow, N.; Ladenbauer, J.; Antonenko, D.; Grittner, U.; Tamm, S.; Flöel, A. Boosting Slow Oscillatory Activity Using tDCS during Early Nocturnal Slow Wave Sleep Does Not Improve Memory Consolidation in Healthy Older Adults. Brain Stimul. 2016, 9, 730–739. [Google Scholar] [CrossRef]
- Frase, L.; Piosczyk, H.; Jahn, F.; Zittel, S.; Krone, L.; Selhausen, P.; Feige, B.; Maier, J.G.; Riemann, D.; Nitsche, M.A.; et al. P173 Bi-frontal transcranial direct current stimulation does modulate sleep, but does not affect sleep-related memory consolidation. Clin. Neurophysiol. 2017, 128, e101–e102. [Google Scholar] [CrossRef]
- Lafon, B.; Henin, S.; Huang, Y.; Friedman, D.; Melloni, L.; Thesen, T.; Doyle, W.; Buzsáki, G.; Devinsky, O.; Parra, L.C.; et al. Low frequency transcranial electrical stimulation does not entrain sleep rhythms measured by human intracranial recordings. Nat. Commun. 2017, 8, 1199. [Google Scholar] [CrossRef]
- Fehér, K.D.; Wunderlin, M.; Maier, J.G.; Hertenstein, E.; Schneider, C.L.; Mikutta, C.; Züst, M.A.; Klöppel, S.; Nissen, C. Shaping the slow waves of sleep: A systematic and integrative review of sleep slow wave modulation in humans using non-invasive brain stimulation. Sleep Med. Rev. 2021, 58, 101438. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- MATLAB, Version 9.3 (R2017b); The MathWorks Inc.: Natick, MA, USA, 2017.
- Garside, P.; Arizpe, J.; Lau, C.-I.; Goh, C.; Walsh, V. Cross-hemispheric Alternating Current Stimulation During a Nap Disrupts Slow Wave Activity and Associated Memory Consolidation. Brain Stimul. 2015, 8, 520–527. [Google Scholar] [CrossRef]
- Fenn, K.M.; Hambrick, D.Z. General intelligence predicts memory change across sleep. Psychon. Bull. Rev. 2015, 22, 791–799. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows, Version 24.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical power analyzes using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef]
- Tseng, P.; Hsu, T.Y.; Chang, C.F.; Tzeng, O.J.; Hung, D.L.; Muggleton, N.G.; Walsh, V.; Liang, W.K.; Cheng, S.K.; Juan, C.H. Unleashing potential: Transcranial direct current stimulation over the right posterior parietal cortex improves change detection in low-performing individuals. J. Neurosci. 2012, 32, 10554–10561. [Google Scholar] [CrossRef]
- Hsu, T.Y.; Juan, C.H.; Tseng, P. Individual differences and state-dependent responses in transcranial direct current stimulation. Front. Hum. Neurosci. 2016, 10, 643. [Google Scholar] [CrossRef]
- Katz, B.; Au, J.; Buschkuehl, M.; Abagis, T.; Zabel, C.; Jaeggi, S.M.; Jonides, J. Individual differences and long-term consequences of tDCS-augmented cognitive training. J. Cogn. Neurosci. 2017, 29, 1498–1508. [Google Scholar] [CrossRef]
- Toussaint, M.; Luthringer, R.; Schaltenbrand, N.; Carelli, G.; Lainey, E.; Jacqmin, A.; Muzet, A.; Macher, J.-P. First-Night Effect in Normal Subjects and Psychiatric Inpatients. Sleep 1995, 18, 463–469. [Google Scholar] [CrossRef]
- Ngo, H.-V.V.; Martinetz, T.; Born, J.; Mölle, M. Auditory Closed-Loop Stimulation of the Sleep Slow Oscillation Enhances Memory. Neuron 2013, 78, 545–553. [Google Scholar] [CrossRef]
- Oudiette, D.; Antony, J.W.; Paller, K.A. Fear not: Manipulating sleep might help you forget. Trends Cogn. Sci. 2014, 18, 3–4. [Google Scholar] [CrossRef][Green Version]
- Clark, V.P.; Coffman, B.A.; Mayer, A.R.; Weisend, M.P.; Lane, T.D.R.; Calhoun, V.D.; Raybourn, E.M.; Garcia, C.M.; Wassermann, E.M. TDCS guided using fMRI significantly accelerates learning to identify concealed objects. NeuroImage 2012, 59, 117–128. [Google Scholar] [CrossRef]
- Coffman, B.A.; Trumbo, M.C.; Flores, R.A.; Garcia, C.M.; Van Der Merwe, A.J.; Wassermann, E.M.; Weisend, M.P.; Clark, V.P. Impact of tDCS on performance and learning of target detection: Interaction with stimulus characteristics and experimental design. Neuropsychologia 2012, 50, 1594–1602. [Google Scholar] [CrossRef][Green Version]
- Coffman, B.A.; Trumbo, M.C.; Clark, V.P. Enhancement of object detection with transcranial direct current stimulation is associated with increased attention. BMC Neurosci. 2012, 13, 108. [Google Scholar] [CrossRef]
- Clark, V.P.; Coffman, B.A.; Trumbo, M.C.S.; Wegele, A.R. An Evolutionary Perspective on Attentional Processes. In Cognitive Electrophysiology of Attention: Signals of the Mind; Mangun, G.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 207–215. ISBN 9780123984517. [Google Scholar]
- Falcone, B.; Coffman, B.A.; Clark, V.P.; Parasuraman, R. Transcranial Direct Current Stimulation Augments Perceptual Sensitivity and 24-Hour Retention in a Complex Threat Detection Task. PLoS ONE 2012, 7, e34993. [Google Scholar] [CrossRef]
- Trumbo, M.C.; Matzen, L.E.; Coffman, B.A.; Hunter, M.A.; Jones, A.P.; Robinson, C.S.; Clark, V.P. Enhanced working memory performance via transcranial direct current stimulation: The possibility of near and far transfer. Neuropsychologia 2016, 93, 85–96. [Google Scholar] [CrossRef]
- Hunter, M.A.; Lieberman, G.; Coffman, B.A.; Trumbo, M.C.; Armenta, M.L.; Robinson, C.S.; Clark, V.P. Mindfulness-based training with transcranial direct current stimulation modulates neuronal resource allocation in working memory: A randomized pilot study with a nonequivalent control group. Heliyon 2018, 4, e00685. [Google Scholar] [CrossRef]
- Lerner, I.; Ketz, N.A.; Jones, A.P.; Bryant, N.B.; Robert, B.; Skorheim, S.W.; Hartholt, A.; Rizzo, A.S.; Gluck, M.A.; Clark, V.P.; et al. Transcranial current stimulation during sleep facilitates insight into temporal rules, but does not consolidate memories of individual sequential experiences. Sci. Rep. 2019, 9, 1516. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, A.P.; Bryant, N.B.; Robert, B.M.; Mullins, T.S.; Trumbo, M.C.S.; Ketz, N.A.; Howard, M.D.; Pilly, P.K.; Clark, V.P. Closed-Loop tACS Delivered during Slow-Wave Sleep Reduces Retroactive Interference on a Paired-Associates Learning Task. Brain Sci. 2023, 13, 468. https://doi.org/10.3390/brainsci13030468
Jones AP, Bryant NB, Robert BM, Mullins TS, Trumbo MCS, Ketz NA, Howard MD, Pilly PK, Clark VP. Closed-Loop tACS Delivered during Slow-Wave Sleep Reduces Retroactive Interference on a Paired-Associates Learning Task. Brain Sciences. 2023; 13(3):468. https://doi.org/10.3390/brainsci13030468
Chicago/Turabian StyleJones, Aaron P., Natalie B. Bryant, Bradley M. Robert, Teagan S. Mullins, Michael C. S. Trumbo, Nicholas A. Ketz, Michael D. Howard, Praveen K. Pilly, and Vincent P. Clark. 2023. "Closed-Loop tACS Delivered during Slow-Wave Sleep Reduces Retroactive Interference on a Paired-Associates Learning Task" Brain Sciences 13, no. 3: 468. https://doi.org/10.3390/brainsci13030468
APA StyleJones, A. P., Bryant, N. B., Robert, B. M., Mullins, T. S., Trumbo, M. C. S., Ketz, N. A., Howard, M. D., Pilly, P. K., & Clark, V. P. (2023). Closed-Loop tACS Delivered during Slow-Wave Sleep Reduces Retroactive Interference on a Paired-Associates Learning Task. Brain Sciences, 13(3), 468. https://doi.org/10.3390/brainsci13030468