
A Transgenic 5xFAD-M Line of Mice for Dendritic Spine Morphology Analysis in Alzheimer’s Disease



Abstract
:
1. Introduction
2. Materials and Methods
3. Results
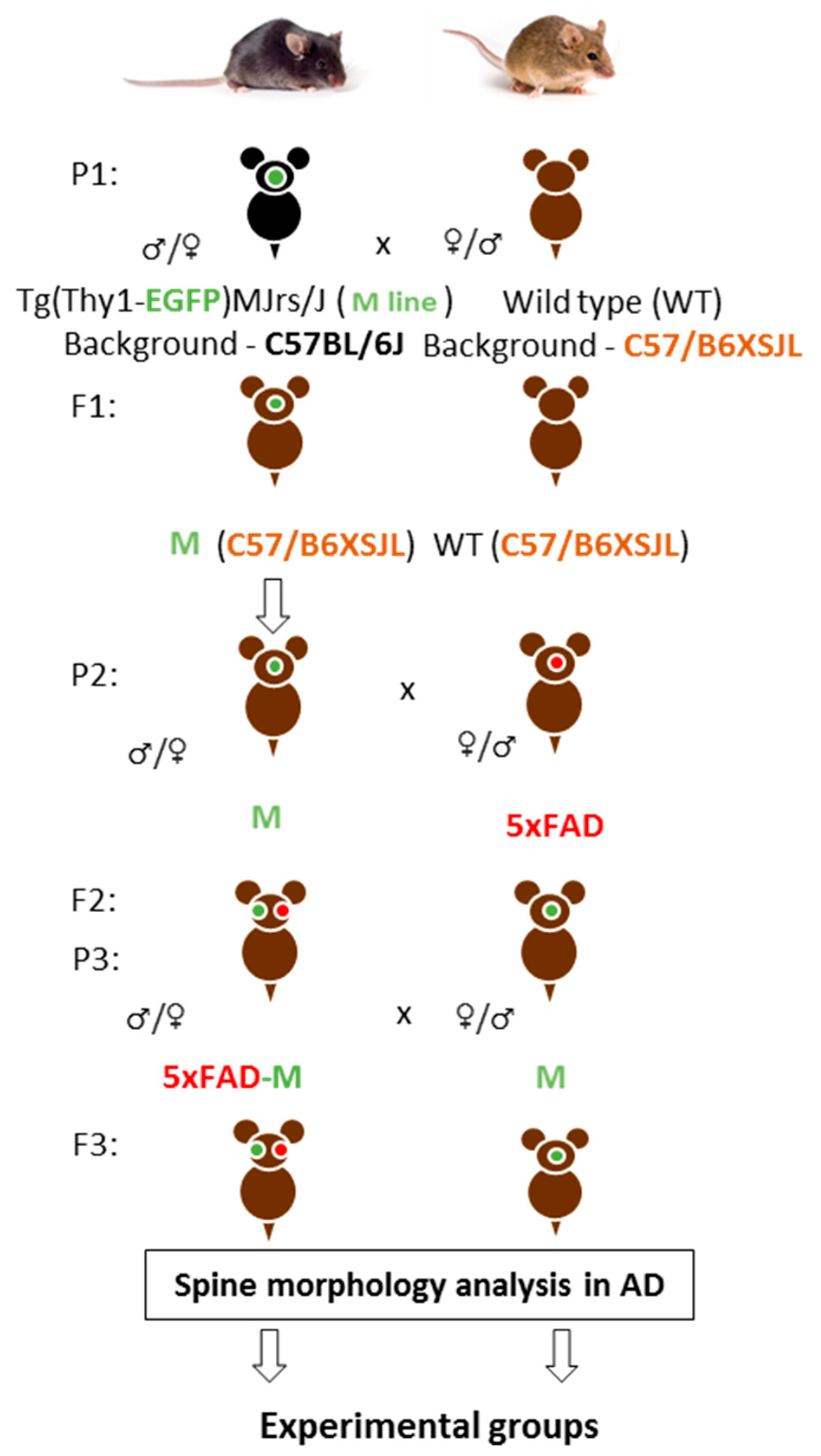
3.1. Creating of a Transgenic Line of 5xFAD-M Mice
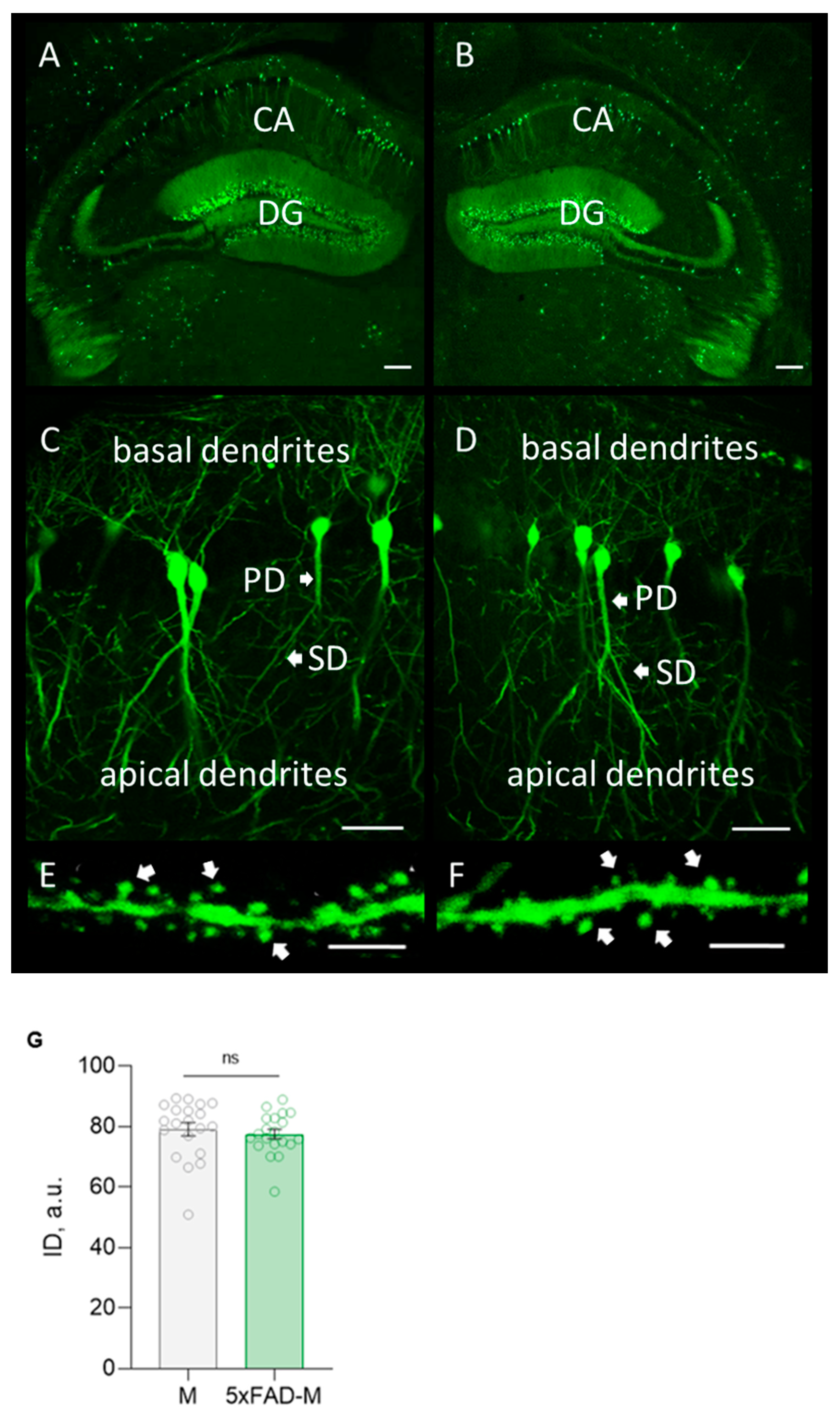
3.2. 5xFAD-M Mice Demonstrate Elimination of Mushroom Spines between 4 and 5 Months of Age
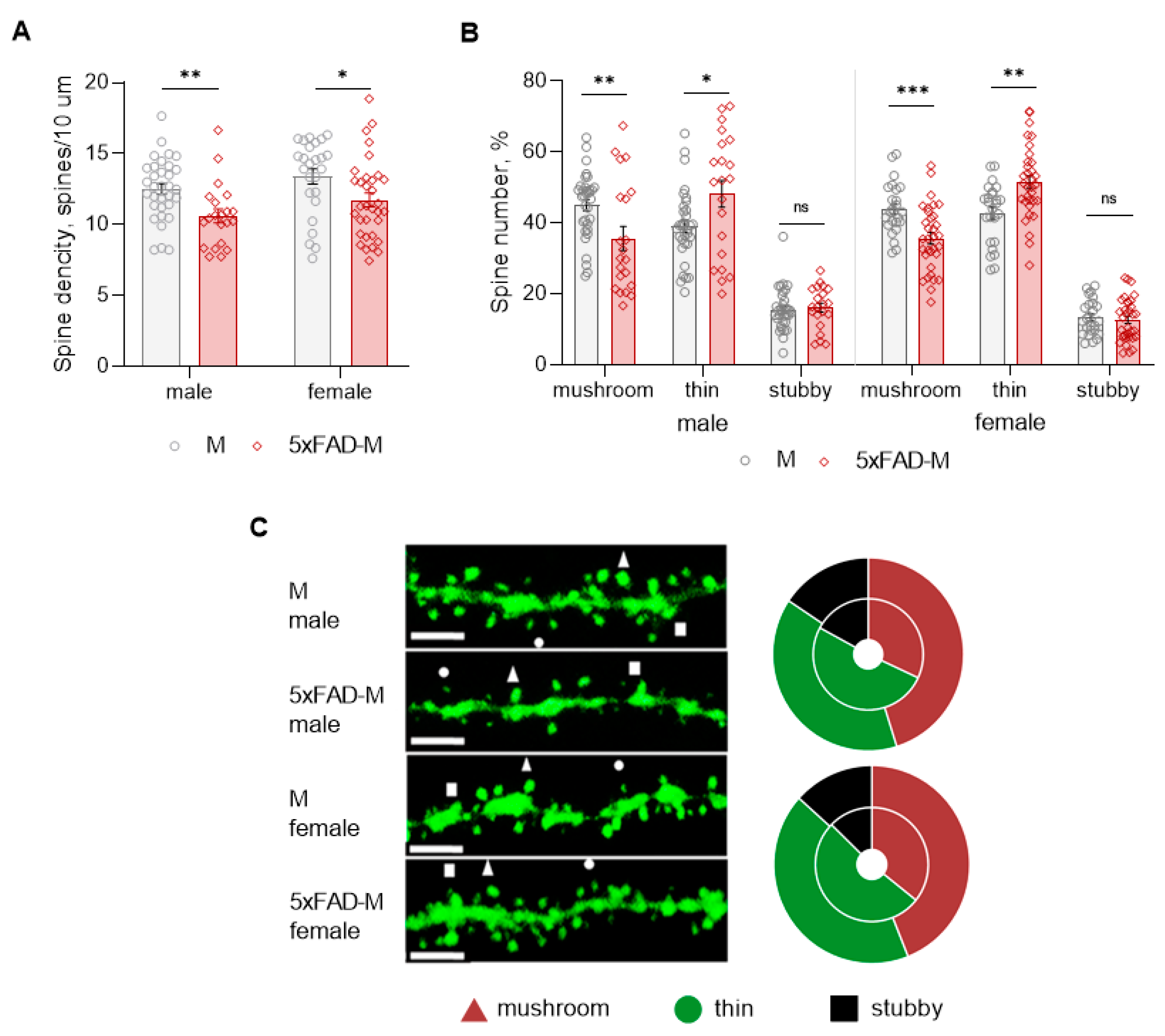
3.3. Dendritic Spine Pathology in Male and Female 5xFAD-M Mice
4. Discussion
5. Conclusions
6. Limitations and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeTure, M.A.; Dickson, D.W. The neuropathological diagnosis of Alzheimer’s disease. Mol. Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef]
- Tarawneh, R.; Holtzman, D.M. The clinical problem of symptomatic Alzheimer disease and mild cognitive impairment. Cold Spring Harb. Perspect. Med. 2012, 2, a006148. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Li, X.; Cheng, J.; Hou, L. Drug Development for Alzheimer’s Disease: Microglia Induced Neuroinflammation as a Target? Int. J. Mol. Sci. 2019, 20, 558. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-Q.; Mobley, W.C. Alzheimer Disease Pathogenesis: Insights From Molecular and Cellular Biology Studies of Oligomeric Aβ and Tau Species. Front. Neurosci. 2019, 13, 659. [Google Scholar] [CrossRef] [PubMed]
- Cronin, T.; Arshad, Q.; Seemungal, B.M. Vestibular Deficits in Neurodegenerative Disorders: Balance, Dizziness, and Spatial Disorientation. Front. Neurol. 2017, 8, 538. [Google Scholar] [CrossRef]
- Vermunt, L.; Sikkes, S.A.M.; van den Hout, A.; Handels, R.; Bos, I.; van der Flier, W.M.; Kern, S.; Ousset, P.-J.; Maruff, P.; Skoog, I.; et al. Duration of preclinical, prodromal, and dementia stages of Alzheimer’s disease in relation to age, sex, and APOE genotype. Alzheimers Dement. 2019, 15, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Palmqvist, S.; Schöll, M.; Strandberg, O.; Mattsson, N.; Stomrud, E.; Zetterberg, H.; Blennow, K.; Landau, S.; Jagust, W.; Hansson, O. Earliest accumulation of β-amyloid occurs within the default-mode network and concurrently affects brain connectivity. Nat. Commun. 2014, 8, 1214. [Google Scholar] [CrossRef]
- Sun, X.; Chen, W.-D.; Wang, Y.-D. β-Amyloid: The key peptide in the pathogenesis of Alzheimer’s disease. Front. Pharmacol. 2015, 6, 221. [Google Scholar] [CrossRef]
- Thinakaran, G.; Koo, E.H. Amyloid precursor protein trafficking, processing, and function. J. Biol. Chem. 2008, 283, 29615–29619. [Google Scholar] [CrossRef]
- Bhadbhade, A.; Cheng, D.W. Amyloid Precursor Protein Processing in Alzheimer’s Disease. Iran. J. Child Neurol. 2012, 6, 1–4. [Google Scholar]
- Zheng, H.; Koo, E.H. Biology and pathophysiology of the amyloid precursor protein. Mol. Neurodegener. 2011, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Lanoiselée, H.M.; Nicolas, G.; Wallon, D.; Rovelet-Lecrux, A.; Lacour, M.; Rousseau, S.; Richard, A.C.; Pasquier, F.; Rollin-Sillaire, A.; Martinaud, O.; et al. APP, PSEN1, and PSEN2 mutations in early-onset Alzheimer disease: A genetic screening study of familial and sporadic cases. PLoS Med. 2017, 14, e1002270. [Google Scholar] [CrossRef] [PubMed]
- Lue, L.-F.; Kuo, Y.-M.; Roher, A.; Brachova, L.; Shen, Y.; Sue, L.; Beach, T.; Kurth, J.H.; Rydel, R.E.; Rogers, J. Soluble amyloid β peptide concentration as a predictor of synaptic change in Alzheimer’s disease. Am. J. Pathol. 1999, 155, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Terry, R.D.; Masliah, E.; Salmon, D.P.; Butters, N.; DeTeresa, R.; Hill, R.; Hansen, L.A.; Katzman, R. Physical basis of cognitive alterations in Alzheimer’s disease: Synapse loss is the major correlate of cognitive impairment. Ann. Neurol. 1991, 30, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Pchitskaya, E.; Bezprozvanny, I. Dendritic Spines Shape Analysis-Classification or Clusterization? Perspective. Front. Synaptic Neurosci. 2020, 12, 31. [Google Scholar] [CrossRef]
- Murakoshi, H.; Yasuda, R. Postsynaptic signaling during plasticity of dendritic spines. Trends Neurosci. 2012, 35, 135–143. [Google Scholar] [CrossRef]
- Holtmaat, A.; Svoboda, K. Experience-dependent structural synaptic plasticity in the mammalian brain. Nat. Rev. Neurosci. 2009, 10, 647–658. [Google Scholar] [CrossRef]
- DeKosky, S.T.; Scheff, S.W. Synapse loss in frontal cortex biopsies in Alzheimer’s disease: Correlation with cognitive severity. Ann. Neurol. 1990, 27, 457–464. [Google Scholar] [CrossRef]
- Jawhar, S.; Trawicka, A.; Jenneckens, C.; Bayer, T.A.; Wirths, O. Motor deficits, neuron loss, and reduced anxiety coinciding with axonal degeneration and intraneuronal Aβ aggregation in the 5XFAD mouse model of Alzheimer’s disease. Neurobiol. Aging 2012, 33, 196.e29–196.e40. [Google Scholar] [CrossRef]
- Oakley, H.; Cole, S.L.; Logan, S.; Maus, E.; Shao, P.; Craft, J.; Guillozet-Bongaarts, A.; Ohno, M.; Disterhoft, J.; Van Eldik, L.; et al. Intraneuronal β-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer’s disease mutations: Potential factors in amyloid plaque formation. J. Neurosci. 2006, 26, 10129–10140. [Google Scholar] [CrossRef]
- O′Leary, T.P.; Brown, R.E. Visuo-spatial learning and memory impairments in the 5xFAD mouse model of Alzheimer′s disease: Effects of age, sex, albinism, and motor impairments. Genes Brain Behav. 2022, 21, e12794. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, H.A.; Han, Y.S.; Jeon, W.K.; Han, J.S. Recognition memory impairments and amyloid-beta deposition of the retrosplenial cortex at the early stage of 5XFAD mice. Physiol. Behav. 2020, 222, 112891. [Google Scholar] [CrossRef] [PubMed]
- Kimura, R.; Ohno, M. Impairments in remote memory stabilization precede hippocampal synaptic and cognitive failures in 5XFAD Alzheimer mouse model. Neurobiol. Dis. 2009, 33, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Spires, T.L.; Meyer-Luehmann, M.; Stern, E.A.; McLean, P.J.; Skoch, J.; Nguyen, P.T.; Bacskai, B.J.; Hyman, B.T. Hyman Dendritic Spine Abnormalities in Amyloid Precursor Protein Transgenic Mice Demonstrated by Gene Transfer and Intravital Multiphoton Microscopy. J. Neurosci. 2005, 25, 7278–7287. [Google Scholar] [CrossRef]
- Feng, G.; Mellor, R.H.; Bernstein, M.; Keller-Peck, C.; Nguyen, Q.T.; Wallace, M.; Nerbonne, J.M.; Lichtman, J.W.; Sanes, J.R. Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP. Neuron 2000, 28, 41–51. [Google Scholar] [CrossRef]
- Rodriguez, A.; Ehlenberger, D.B.; Dickstein, D.L.; Hof, P.R.; Wearne, S.L. Automated three-dimensional detection and shape classification of dendritic spines from fluorescence microscopy images. PLoS ONE 2008, 3, e1997. [Google Scholar] [CrossRef]
- Gatto, R.G.; Chu, Y.; Ye, A.Q.; Price, S.D.; Tavassoli, E.; Buenaventura, A.; Brady, S.T.; Magin, R.L.; Kordower, J.H.; Morfini, G.A. Analysis of YFP(J16)-R6/2 reporter mice and postmortem brains reveals early pathology and increased vulnerability of callosal axons in Huntington’s disease. Hum. Mol. Genet. 2015, 24, 5285–5298. [Google Scholar] [CrossRef]
- Ryskamp, D.; Wu, L.; Wu, J.; Kim, D.; Rammes, G.; Geva, M.; Hayden, M.; Bezprozvanny, I. Pridopidine stabilizes mushroom spines in mouse models of Alzheimer′s disease by acting on the sigma-1 receptor. Neurobiol. Dis. 2019, 124, 489–504. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, S.; Wu, L.; Pchitskaya, E.; Zakharova, O.; Fon Tacer, K.; Bezprozvanny, I. Store-Operated Calcium Channel Complex in Postsynaptic Spines: A New Therapeutic Target for Alzheimer′s Disease Treatment. J. Neurosci. 2016, 36, 11837–11850. [Google Scholar] [CrossRef]
- Crowe, S.E.; Ellis-Davies, G.C. Spine pruning in 5xFAD mice starts on basal dendrites of layer 5 pyramidal neurons. Brain Struct. Funct. 2014, 219, 571–580. [Google Scholar] [CrossRef]
- Zhang, J.; Long, B.; Li, A.; Sun, Q.; Tian, J.; Luo, T.; Ding, Z.; Gong, H.; Li, X. Whole-Brain Three-Dimensional Profiling Reveals Brain Region Specific Axon Vulnerability in 5xFAD Mouse Model. Front. Neuroanat. 2020, 14, 608177. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Pozo, A.; Frosch, M.P.; Masliah, E.; Hyman, B.T. Neuropathological alterations in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef] [PubMed]
- Berry, K.P.; Nedivi, E. Spine Dynamics: Are They All the Same? Neuron 2017, 96, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Bourne, J.; Harris, K.M. Do thin spines learn to be mushroom spines that remember? Curr. Opin. Neurobiol. 2007, 17, 381–386. [Google Scholar] [CrossRef]
- Tackenberg, C.; Ghori, A.; Brandt, R. Thin, stubby or mushroom: Spine pathology in Alzheimer′s disease. Curr. Alzheimer Res. 2009, 6, 261–268. [Google Scholar] [CrossRef]
- Kasai, H.; Ziv, N.E.; Okazaki, H.; Yagishita, S.; Toyoizumi, T. Spine dynamics in the brain, mental disorders and artificial neural networks. Nat. Rev. Neurosci. 2021, 22, 407–422. [Google Scholar] [CrossRef]
- Runge, K.; Cardoso, C.; de Chevigny, A. Dendritic Spine Plasticity: Function and Mechanisms. Front. Synaptic Neurosci. 2020, 12, 36. [Google Scholar] [CrossRef]
- Baltaci, S.B.; Mogulkoc, R.; Baltaci, A.K. Molecular Mechanisms of Early and Late LTP. Neurochem. Res. 2019, 44, 281–296. [Google Scholar] [CrossRef]
- Halpain, S.; Spencer, K.; Graber, S. Dynamics and pathology of dendritic spines. Prog Brain Res. 2005, 147, 29–37. [Google Scholar]
- Lee, S.; Zhang, H.; Webb, D. Dendritic spine morphology and dynamics in health and disease. Cell Health Cytoskelet. 2015, 7, 121–131. [Google Scholar]
- Ammassari-Teule, M. Early-Occurring Dendritic Spines Alterations in Mouse Models of Alzheimer′s Disease Inform on Primary Causes of Neurodegeneration. Front. Synaptic Neurosci. 2020, 12, 566615. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, Y.; Li, L.J.; Zhu, K.; Zheng, Y.; Wang, X.M. Glutamate receptor delocalization in postsynaptic membrane and reduced hippocampal synaptic plasticity in the early stage of Alzheimer’s disease. Neural Regen. Res. 2019, 14, 1037–1045. [Google Scholar] [PubMed]
- Zhang, M.; Bao, Y.; Yang, Y.; Kimber, M.; Levine, M.; Xie, F. Identifying Attributes for a Value Assessment Framework in China: A Qualitative Study. Pharmacoeconomics 2023, 1–17. [Google Scholar] [CrossRef]
- Seo, H.J.; Park, J.E.; Choi, S.M.; Kim, T.; Cho, S.H.; Lee, K.H.; Song, W.K.; Song, J.; Jeong, H.S.; Kim, D.H.; et al. Inhibitory Neural Network′s Impairments at Hippocampal CA1 LTP in an Aged Transgenic Mouse Model of Alzheimer′s Disease. Int. J. Mol. Sci. 2021, 22, 698. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Chen, L.; Baglietto-Vargas, D.; Kamalipour, P.; Ye, Q.; LaFerla, F.M.; Nitz, D.A.; Holmes, T.C.; Xu, X. Spatial coding defects of hippocampal neural ensemble calcium activities in the triple-transgenic Alzheimer’s disease mouse model. Neurobiol. Dis. 2022, 162, 105562. [Google Scholar] [CrossRef]
- Yuste, R.; Bonhoeffer, T. Morphological changes in dendritic spines associated with long-term synaptic plasticity. Annu. Rev. Neurosci. 2001, 24, 1071–1089. [Google Scholar] [CrossRef]
- Khavinson, V.; Ilina, A.; Kraskovskaya, N.; Linkova, N.; Kolchina, N.; Mironova, E.; Erofeev, A.; Petukhov, M. Neuroprotective effects of tripeptides—Epigenetic regulators in mouse model of Alzheimer’s disease. Pharmaceuticals 2021, 14, 515. [Google Scholar] [CrossRef]
- Bishop, H.I.; Guan, D.; Bocksteins, E.; Parajuli, L.K.; Murray, K.D.; Cobb, M.M.; Misonou, H.; Zito, K.; Foehring, R.C.; Trimmer, J.S. Distinct Cell- and Layer-Specific Expression Patterns and Independent Regulation of Kv2 Channel Subtypes in Cortical Pyramidal Neurons. J. Neurosci. 2015, 35, 14922–14942. [Google Scholar] [CrossRef]
- Somogyi, P.; Takagi, H. A note on the use of picric acid-paraformaldehyde-glutaraldehyde fixative for correlated light and electron microscopic immunocytochemistry. Neuroscience 1982, 7, 1779–1783. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Indicators | M Mice | 5xFAD-M Mice |
|---|---|---|---|
| 4-month | DSD | 13.82 ± 0.62 | 11.64 ± 0.74 |
| MS | 34.83 ± 2.02 | 36.44 ± 2.48 | |
| TS | 47.68 ± 2.48 | 43.29 ± 3.51 | |
| SS | 17.86 ± 1.88 | 20.26 ± 2.35 | |
| 5-month | DSD | 12.40 ± 0.38 | 10.62 ± 0.49 * |
| MS | 45.06 ± 1.64 | 35.64 ± 3.41 ** | |
| TS | 39.07 ± 1.78 | 48.21 ± 3.78 * | |
| SS | 15.61 ± 1.02 | 16.15 ± 1.30 |
| Sex | Indicators | M Mice | 5xFAD-M Mice |
|---|---|---|---|
| Males | DSD | 12.52 ± 0.38 | 10.62 ± 0.49 ** |
| MS | 45.06 ± 1.64 | 35.64 ± 3.41 ** | |
| TS | 39.07 ± 1.78 | 48.21 ± 3.78 * | |
| SS | 15.61 ± 1.02 | 16.15 ± 1.30 | |
| Females | DSD | 13.39 ± 0.53 | 11.74 ± 0.49 * |
| MS | 43.94 ± 1.44 | 35.60 ± 1.62 *** | |
| TS | 42.65 ± 1.69 | 51.38 ± 1.76 ** | |
| SS | 13.31 ± 0.98 | 12.62 ± 1.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilina, A.; Linkova, N. A Transgenic 5xFAD-M Line of Mice for Dendritic Spine Morphology Analysis in Alzheimer’s Disease. Brain Sci. 2023, 13, 307. https://doi.org/10.3390/brainsci13020307
Ilina A, Linkova N. A Transgenic 5xFAD-M Line of Mice for Dendritic Spine Morphology Analysis in Alzheimer’s Disease. Brain Sciences. 2023; 13(2):307. https://doi.org/10.3390/brainsci13020307
Chicago/Turabian StyleIlina, Anastasiia, and Natalia Linkova. 2023. "A Transgenic 5xFAD-M Line of Mice for Dendritic Spine Morphology Analysis in Alzheimer’s Disease" Brain Sciences 13, no. 2: 307. https://doi.org/10.3390/brainsci13020307
APA StyleIlina, A., & Linkova, N. (2023). A Transgenic 5xFAD-M Line of Mice for Dendritic Spine Morphology Analysis in Alzheimer’s Disease. Brain Sciences, 13(2), 307. https://doi.org/10.3390/brainsci13020307