
Affective Prosody and Its Impact on the Neurology of Language, Depression, Memory and Emotions

Abstract
1. Introduction
2. Neurology of Language
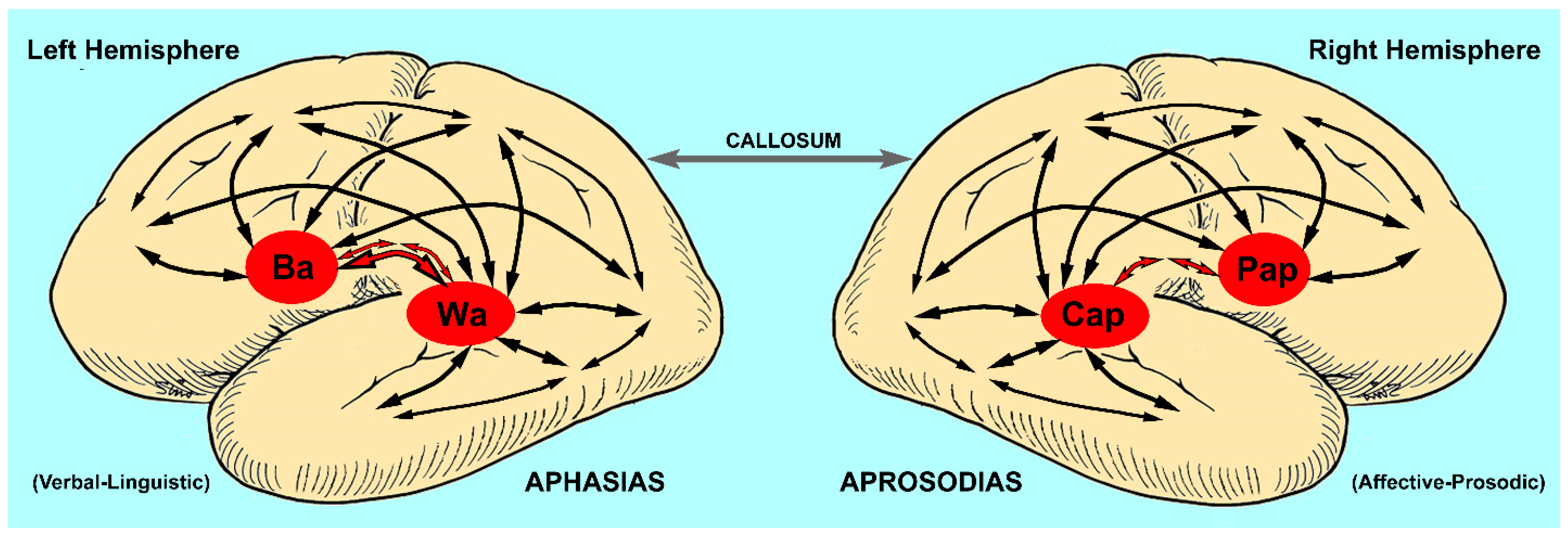
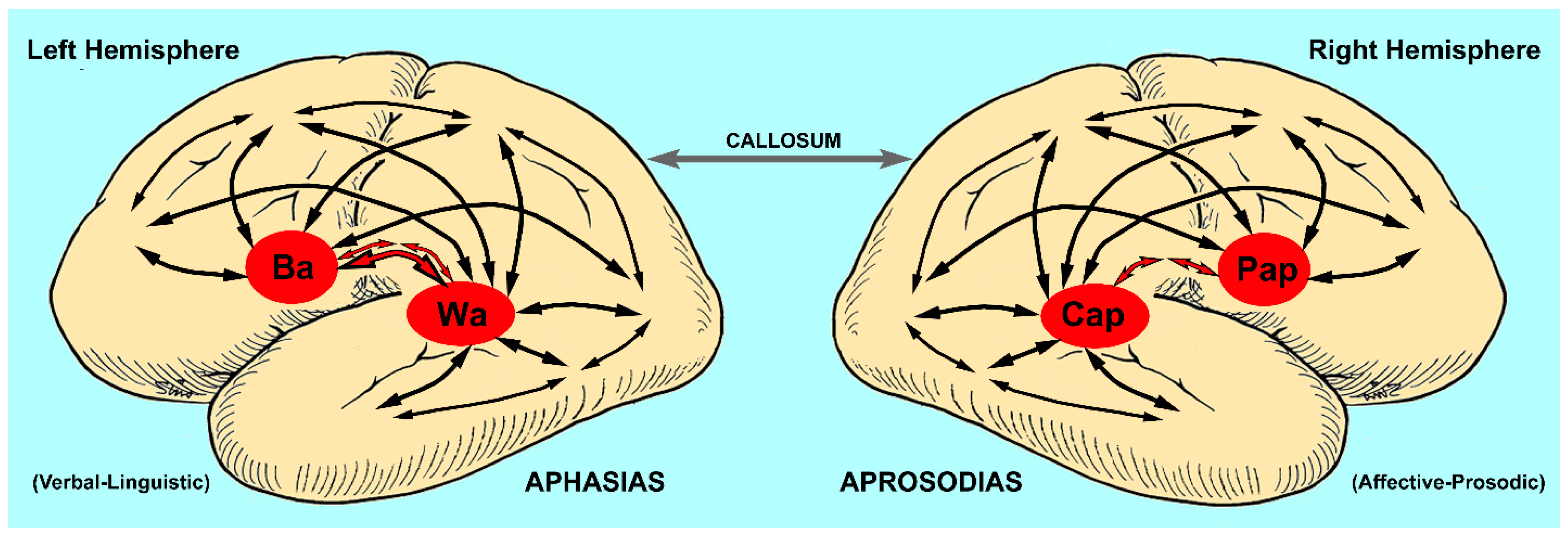
2.1. The Aphasias
2.2. The Aprosodias
2.3. Subcategories of Prosody and Their Acoustic Signatures
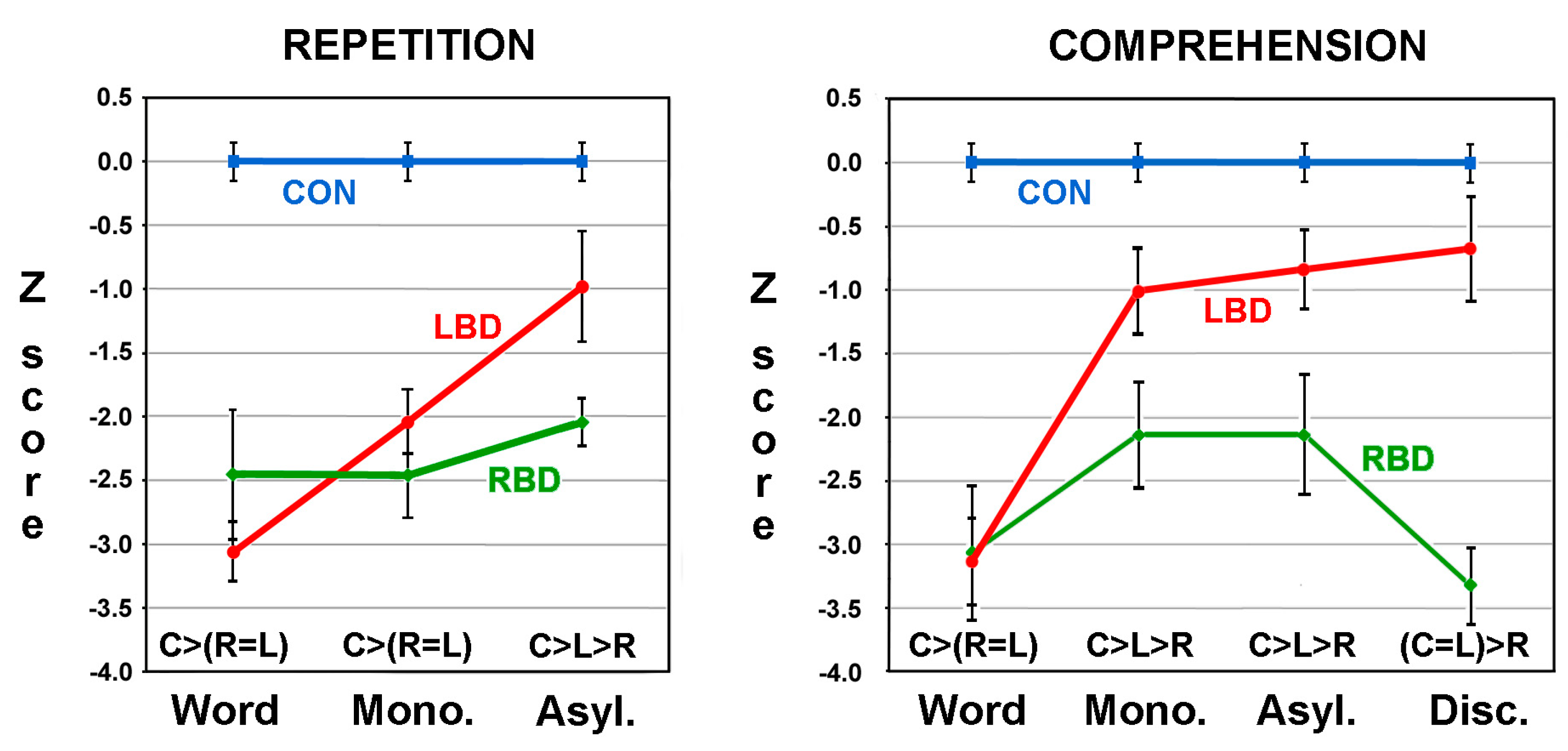
2.4. Acoustic Realization of Affective Prosody in Tone and Non-Tone Languages
2.5. Is Affective Prosody a Dominant and Lateralized Right-Hemisphere Function?
2.6. Are the Aprosodias Functionally and Anatomically Analogous to the Aphasias?
2.7. Neurology of Linguistic Prosody
2.8. Other Right-Hemisphere Contributions to Language and Communication
3. Neurology of Depression and Hemispheric Lateralization of Emotions
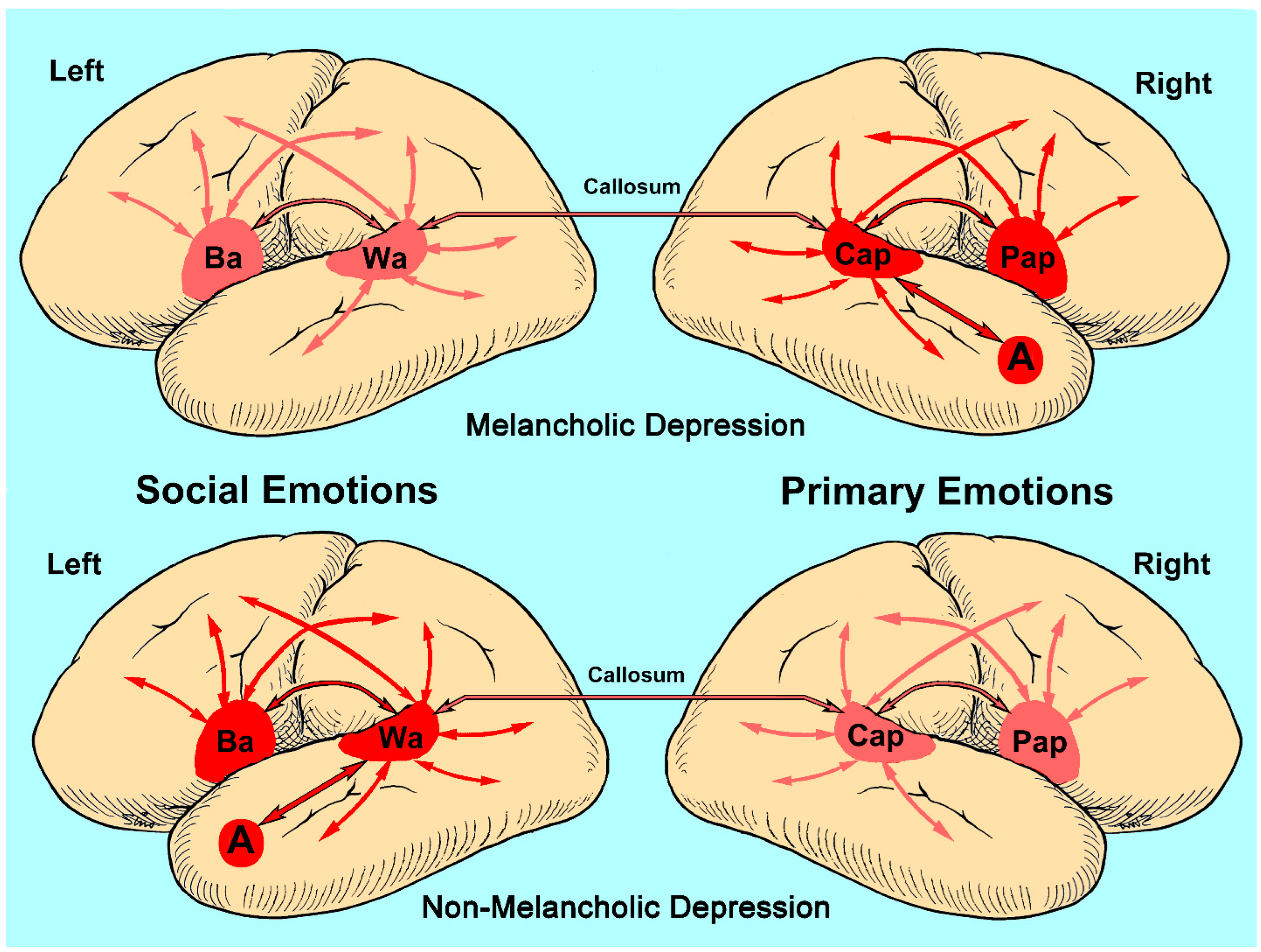
3.1. Academic and Clinical Serendipity
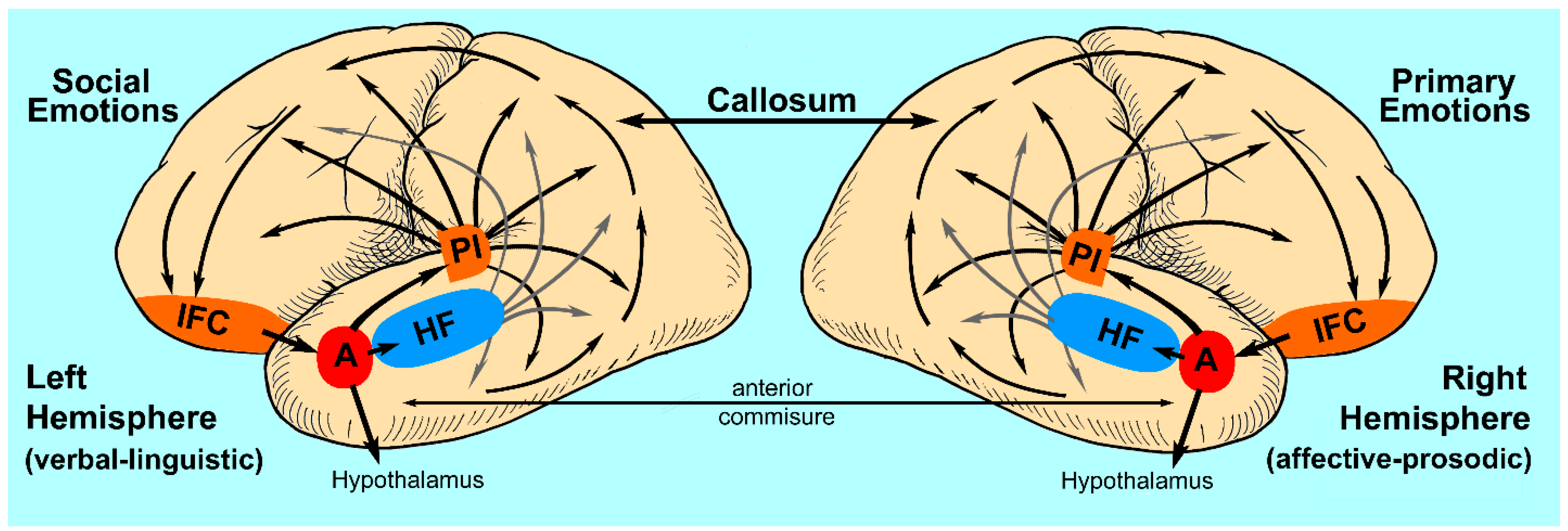
3.2. Social Emotions, Primary Emotions and the Emotion-Type Hypothesis

4. Summary
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Correction Statement
Glossary
| Fundamental frequency | The lowest rate the vocal folds vibrate to produce a complex speech sound. |
| Hertz | An acoustic measure of the frequency of a complex speech sound in cycles per second that is usually the fundamental frequency. The first harmonic of a complex speech sound is usually one musical octave above the fundamental frequency or double the Hertz of the fundamental frequency. The second harmonic is usually one octave above the first harmonic or double the Hertz of the first harmonic. The third harmonic is double the Hertz of the second harmonic, etc. |
| Intonation | Manipulation of pitch over time |
| Lexical tones | Brief pitch contours associated with articulation of words in tone languages that are critical for word meaning. |
| Operculum | Region of the brain that surrounds the Sylvian fissure and overlies the insula. |
| Pitch | A psychoacoustic construct of how high or low a speech sound is perceived by a listener relative to a musical scale. Pitch is produced by vibrations of the vocal folds to produce a sound that has a fundamental frequency and various harmonics that are octave multiples. |
| Semitones | Converts Hertz to an acoustically logarithmic scale in which musical octaves (harmonics) are equally represented as a 12-semitone change in perceived pitch. |
References
- Ross, E.D. Right hemisphere syndromes and the neurology of emotions. In Behavioral Neurology and the Legacy of Norman Geschwind; Schacter, S.C., Devinsky, O., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 1997; Chapter 23; pp. 183–191. [Google Scholar]
- MacPherson, S.; Della Sala, S. Cases of Amnesia: Contributions to Understanding Memory and the Brain; Routledge: New York, NY, USA, 2019. [Google Scholar]
- Ross, E.D. Sensory-specific visual amnesia (Cases 1 and 2): An acquired visual-limbic disconnection syndrome. In Cases of Amnesia: Contributions to Understanding Memory and the Brain; MacPherson, S.E., Della Sala, S.D., Eds.; Routledge: New York, NY, USA, 2019; Chapter 10; pp. 203–219. [Google Scholar]
- Ross, E.D. Differential hemispheric lateralization of emotions and related display behaviors: Emotion-type hypothesis. Brain Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Ross, E.D.; Homan, R.W.; Buck, R. Differential hemispheric lateralization of primary and social emotions: Implications for developing a comprehensive neurology for emotion, repression, and the subconscious. Neuropsychiatry Neuropsychol. Behav. Neurol. 1994, 7, 1–19. [Google Scholar]
- Shin, H.S. Reasoning processes in clinical reasoning: From the perspective of cognitive psychology. Korean J. Med. Educ. 2019, 31, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Varpio, L.; Paradis, E.; Uijtdehaage, S.; Young, M. The distinctions between theory, theoretical framework, and conceptual framework. Acad. Med. 2020, 95, 989–994. [Google Scholar] [CrossRef]
- Broca, P. Remarques sur le siege de la faculte du langage articule, suives d’une observation d’aphemie. Bull. Soc. Anatomique 1861, 6, 330–337. [Google Scholar]
- Signoret, J.L.; Castaigne, P.; Lhermitte, F.; Abelanet, R.; Lavorel, P. Rediscovery of Leborgne’s brain: Anatomical description with CT scan. Brain Lang. 1984, 22, 303–319. [Google Scholar] [CrossRef]
- Broca, P. Sur le siege de la faculte du langage articule. Bull Soc d’Anthropologie (Paris) 1865, 6, 337–393, [translated by Berker, E.A.; Berker, A.H.; Smith, A. Arch. Neurol. 1986, 43, 1065–1072]. [Google Scholar]
- Eggert, G.H. Wernicke’s Works on Aphasia: A Sourcebook and Review; Mouton Publishers: The Hague, The Netherlands, 1977. [Google Scholar]
- Margolin, D.I. Cognitive neuropsychology. Resolving enigmas about Wernicke’s aphasia and other higher cortical disorders. Arch. Neurol. 1991, 48, 751–765. [Google Scholar] [CrossRef]
- Ross, E.D. Intellectual origins and theoretical framework of behavioral neurology: A response to Dr. Trimble. Neuropsychiatry Neuropsychol. Behav. Neurol. 1993, 6, 65–67. [Google Scholar]
- Ross, E.D. Sensory-specific amnesia and hypoemotionality in humans and monkeys: Gateway for developing a hodology of memory. Cortex 2008, 44, 1010–1022. [Google Scholar] [CrossRef]
- Ross, E.D. Cerebral Localization of functions and the neurology of language: Fact versus fiction or is it something else? Neuroscientist 2010, 16, 222–243. [Google Scholar] [CrossRef] [PubMed]
- Geschwind, N. Wernicke’s contribution to the study of aphasia. Cortex 1967, 3, 449–463. [Google Scholar] [CrossRef]
- Lichtheim, L. On aphasia. Brain 1885, 7, 433–484. [Google Scholar] [CrossRef]
- Berthier, M. Transcortical Aphasias; Psychology Press: East Sussex, UK, 1999. [Google Scholar]
- Benson, D.F. Aphasia, Alexia, and Agraphia; Churchill Livingstone: New York, NY, USA, 1979. [Google Scholar]
- Kertesz, A. Aphasia and Associated Disorders; Grune & Stratton: New York, NY, USA, 1979. [Google Scholar]
- Tonkonogy, J.M. Vascular Aphasia; MIT Press: Cambridge, MA, USA, 1986. [Google Scholar]
- Damasio, A.R.; Damasio, H.; Rizzo, M.; Varney, N.; Gersch, F. Aphasia with nonhemorrhagic lesions in the basal ganglia and internal capsule. Arch. Neurol. 1982, 39, 15–20. [Google Scholar] [CrossRef]
- Naeser, M.A.; Alexander, M.P.; Helm-Estabrooks, N.; Levine, H.L.; Laughlin, S.A.; Geschwind, N. Aphasia with predominantly subcortical lesion sites: Description of three capsular/putaminal aphasia syndromes. Arch. Neurol. 1982, 39, 2–14. [Google Scholar] [CrossRef]
- Robin, D.A.; Schienberg, S. Subcortical lesions and aphasia. J. Speech Hear. Disord. 1990, 55, 90–100. [Google Scholar] [CrossRef]
- Demonet, J.F. Subcortical aphasia(s): A controversial and promising topic. Brain Lang. 1997, 58, 410–417, discussion 436–458. [Google Scholar] [CrossRef]
- Demonet, J.F.; Puel, M.; Celsi, P.; Cardeba, D. “Subcortical” aphasia: Some proposed pathophysiological mechanisms and their rCBF correlates revealed by SPECT. J. Neurolinguistics 1991, 6, 319–344. [Google Scholar] [CrossRef]
- Mohr, J.P. Rapid amelioration of motor aphasia. Arch. Neurol. 1973, 28, 77–82. [Google Scholar] [CrossRef]
- Naeser, M.A.; Helm-Estabrooks, N.; Haas, G.; Auerbach, S.; Srinivasan, M. Relationship between lesion extent in ‘Wernicke’s area’ on computed tomographic scan and predicting recovery of comprehension in Wernicke’s aphasia. Arch. Neurol. 1987, 44, 73–82. [Google Scholar] [CrossRef]
- Catani, M.; Jones, D.K.; Ffytche, D.H. Perisylvian language networks of the human brain. Ann. Neurol. 2005, 5, 8–16. [Google Scholar] [CrossRef]
- Catani, M.; Allin, M.P.; Husain, M.; Pugliese, L.; Mesulam, M.M.; Murray, R.M.; Jones, D.K. Symmetries in human brain language pathways correlate with verbal recall. Proc. Natl. Acad. Sci. USA 2007, 104, 17163–17168. [Google Scholar] [CrossRef] [PubMed]
- Mesulam, M.M. Behavioral neuroanatomy: Large-scale networks, association cortex, frontal syndromes, the limbic system, and hemispheric specializations. In Principles of Behavioral Neurology; Mesulam, M.M., Ed.; F.A. Davis: Philadelphia, PA, USA, 1985; Chapter 1; pp. 1–120. [Google Scholar]
- Ross, E.D.; Thompson, R.D.; Yenkosky, J.P. Lateralization of affective prosody in brain and the callosal integration of hemispheric language functions. Brain Lang. 1997, 56, 27–54. [Google Scholar] [CrossRef]
- Ross, E.D.; Monnot, M. Neurology of affective prosody and its functional-anatomic organization in right hemisphere. Brain Lang. 2008, 104, 51–74. [Google Scholar] [CrossRef]
- Benton, A.L. The “minor” hemisphere. J. Hist. Allied. Sci. 1972, 27, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.H. On affections of speech from diseases of the brain (parts 1). Brain 1878, 1, 304–330. [Google Scholar] [CrossRef]
- Jackson, J.H. On affections of speech from diseases of the brain (part 2). Brain 1879, 2, 202–222. [Google Scholar]
- Jackson, J.H. On affections of speech from diseases of the brain (part 3). Brain 1879, 2, 323–356. [Google Scholar] [CrossRef]
- Heilman, K.M.; Scholes, R.; Watson, R.T. Auditory affective agnosia. Disturbed comprehension of affective speech. J. Neurol. Neurosurg. Psychiatry 1975, 38, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Tucker, D.M.; Watson, R.T.; Heilman, K.M. Discrimination and evocation of affectively intoned speech in patients with right parietal disease. Neurology 1977, 27, 947–950. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.D.; Mesulam, M.M. Dominant language functions of the right hemisphere? Prosody and emotional gesturing. Arch. Neurol. 1979, 36, 144–148. [Google Scholar] [CrossRef]
- Ross, E.D. The aprosodias: Functional-anatomical organization of the affective components of language in the right hemisphere. Arch. Neurol. 1981, 38, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.D. Disorders of vocal emotional expression and comprehension: The aprosodias. In Handbook of Clinical Neurology: Disorders of Emotion in Neurologic Disease; Heilman, K.M., Nadeau, S.E., Eds.; 3rd Series; Elsevier: Amsterdam, The Netherlands, 2021; Volume 183, Chapter 4; pp. 63–108. [Google Scholar]
- Heilman, K.M. Matter of Mind: A Neurologist’s View of Brain Behavior Relationships; Oxford University Press: New York, NY, USA, 2002; pp. 53–63. [Google Scholar]
- Gorelick, P.B.; Ross, E.D. The aprosodias: Further functional-anatomic evidence of the organization of affective language in the right hemisphere. J. Neurol. Neurosurg. Psychiatry 1987, 50, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Bowers, D.; Heilman, K.M. Dissociation between the processing of affective and nonaffective faces: A case study. J. Clin. Neuropsychol. 1984, 6, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Fromkin, V.; Rodman, R. An Introduction to Language; Holt, Reinhart and Winston: New York, NY, USA, 1977. [Google Scholar]
- Monrad-Krohn, G.H. Dysprosody of altered “melody of language”. Brain 1947, 70, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Monrad-Krohn, G.H. The third element of speech: Prosody and its disorders. In Problems in Dynamic Neurology; Halpern, L., Ed.; Hebrew University Press: Jerusalem, Israel, 1963; pp. 101–118. [Google Scholar]
- Ross, E.D. The modulation of affect and non-verbal communication by the right hemisphere. In Principles of Behavioral Neurology; Mesulam, M.M., Ed.; F.A. Davis: Philadelphia, PA, USA, 1985; Chapter 6; pp. 239–257. [Google Scholar]
- Ross, E.D.; Shayya, L.; Rousseau, J.F. Prosodic stress: Acoustic, aphasic, aprosodic and neuroanatomic interactions. J. Neurolinguistics 2013, 26, 526–551. [Google Scholar] [CrossRef]
- Lehiste, I. Phonetic disambiguation of syntactic ambiguity. Glossa 1973, 7, 107–121. [Google Scholar] [CrossRef]
- Mitchell, R.L.; Ross, E.D. Attitudinal prosody: What we know and directions for future study. Neurosci. Biobehavioral Rev. 2013, 37, 471–479. [Google Scholar] [CrossRef]
- Bolinger, D. Language: The Loaded Weapon; Longman Group: London, UK, 1980. [Google Scholar]
- Ackerman, B.P. Form and function in children’s understanding of ironic utterances. J. Exp. Child Psychol. 1983, 35, 487–508. [Google Scholar] [CrossRef]
- Bowers, D.; Coslett, H.B.; Bauer, R.M.; Speedie, L.J.; Heilman, K.M. Comprehension of emotional prosody following unilateral hemispheric lesions: Processing defect vs distraction defect. Neuropsychologia 1987, 25, 317–328. [Google Scholar] [CrossRef]
- Mehrabian, A. Nonverbal Communication; Transaction Publishers: Edison, NJ, USA, 2007. [Google Scholar]
- Bolinger, D. A theory of pitch accent in English. Word 1958, 14, 109–149. [Google Scholar] [CrossRef]
- Cruttenden, A. Intonation, 2nd ed.; University Press: Cambridge, UK, 1977. [Google Scholar]
- McClean, M.D.; Tiffany, W.R. The acoustic parameters of stress in relation to syllable position, speech loudness and rate. Lang. Speech 1973, 16, 283–290. [Google Scholar] [CrossRef]
- Ross, E.D.; Holzapfel, D.; Freeman, F. Assessment of affective behavior in brain damaged patients using quantitative acoustical-phonetic and gestural measurements. Neurology 1983, 33 (Suppl. S2), 219–220. [Google Scholar]
- Shapiro, B.; Danly, M. The role of the right hemisphere in the control of speech prosody in propositional and affective contexts. Brain Lang. 1985, 25, 19–36. [Google Scholar] [CrossRef]
- Ross, E.D.; Edmondson, J.A.; Seibert, G.B.; Homan, R.W. Acoustical analysis of affective prosody during right-sided Wada test: A within subjects verification of the right hemisphere’s role in language. Brain Lang. 1988, 33, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, P. Speech Physiology and Acoustic Phonetics: An Introduction; MacMillan: New York, NY, USA, 1977. [Google Scholar]
- Ladefoged, P. A Course in Phonetics; Harcourt Brace Jovanovich: New York, NY, USA, 1975. [Google Scholar]
- Borden, G.J.; Harris, K.S. Speech Science Primer: Physiology, Acoustics, and Perception of Speech; Williams & Wilkins: Baltimore, MD, USA, 1980. [Google Scholar]
- Ross, E.D.; Edmondson, J.; Seibert, G.B. The effect of affect on various acoustic measures of prosody in tone and non-tone languages: A comparison based on computer analysis of voice. J. Phon. 1986, 14, 283–302. [Google Scholar] [CrossRef]
- Hughes, C.P.; Chan, J.L.; Su, M.S. Aprosodia in Chinese patients with right cerebral hemisphere lesions. Arch. Neurol. 1983, 40, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Naeser, M.A.; Chan, S.W. Case study of a Chinese aphasic with the Boston diagnostic aphasia exam. Neuropsychologia 1983, 18, 389–410. [Google Scholar] [CrossRef]
- Gandour, J. Aphasia in tone languages. In Aphasia in Atypical Populations; Coppens, P., Basso, A., Lebrun, Y., Eds.; Lawrence Erlbaum: Hillsdale, MI, USA, 1998; pp. 170–195. [Google Scholar]
- Edmondson, J.A.; Chan, J.L.; Seibert, G.B.; Ross, E.D. The effect of right-brain damage on acoustical measures of affective prosody in Taiwanese patients. J. Phon. 1987, 15, 219–233. [Google Scholar] [CrossRef]
- Ross, E.D.; Edmondson, J.A.; Seibert, G.B.; Chan, J.L. Affective exploitation of tone in Taiwanese: An acoustical study of “tone latitude”. J. Phon. 1992, 20, 441–456. [Google Scholar] [CrossRef]
- Wada, J.; Rasmussen, T. Intracarotid injection of sodium amytal for the lateralization of cerebral speech dominance. J. Neurosurg. 1960, 17, 226–282. [Google Scholar] [CrossRef]
- Van Lancker, D.; Sidtis, J.J. The identification of affective prosodic stimuli by left- and right- hemisphere-damaged subjects: All errors are not created equal. J. Speech Hear. Res. 1992, 35, 963–970. [Google Scholar] [CrossRef]
- Ivry, R.; Robertson, L.C. The Two Sides of Perception; MIT Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Zatorre, R.J.; Belin, P. Spectral and temporal processing in human auditory cortex. Cereb. Cortex 2001, 11, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Poeppel, D. The analysis of speech in different temporal integration windows: Cerebral lateralization as ‘asymmetric sampling in time’. Speech Comm. 2003, 41, 245–255. [Google Scholar] [CrossRef]
- Dehaene-Lambertz, G. Cerebral specialization for speech and non-speech stimuli in infants. J. Cogn. Neurosci. 2000, 12, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Dehaene-Lambertz, G.; Pena, M.; Christophe, A.; Landrieu, P. Phoneme perception in a neonate with a left sylvian infarct. Brain Lang. 2004, 88, 26–38. [Google Scholar] [CrossRef]
- Schlanger, B.B.; Schlanger, P.; Gerstmann, L.J. The perception of emotionally toned sentences by right hemisphere-damaged and aphasic subjects. Brain Lang. 1976, 3, 396–403. [Google Scholar] [CrossRef]
- Seron, X.; Van der Kaa, M.A.; Vanderlinden, M.; Remits, A.; Feyereisen, P. Decoding paralinguistic signals: Effect of semantic and prosodic cues on aphasics’ comprehension. J. Commun. Disord. 1982, 5, 223–231. [Google Scholar] [CrossRef]
- de Bleser, R.; Poeck, K. Analysis of prosody in the spontaneous speech of patients with CV-recurring utterances. Cortex 1985, 21, 405–415. [Google Scholar] [CrossRef]
- Cancelliere, A.E.B.; Kertesz, A. Lesion localization in acquired deficits of emotional expression and comprehension. Brain Cogn. 1990, 13, 133–147. [Google Scholar] [CrossRef]
- Bowers, D.; Blonder, L.X.; Heilman, K.M. The Florida Affect Battery (Revised); The Center for Neuropsychological Studies, University of Florida: Gainsville, FL, USA, 1989. [Google Scholar]
- Ross, E.D. Prosodic lateralization: Fact vs fancy or is it all just semantics. Arch. Neurol. 1988, 45, 338–339. [Google Scholar] [CrossRef] [PubMed]
- Blonder, L.X.; Bowers, D.; Heilman, K.M. The role of the right hemisphere in emotional communication. Brain 1991, 114, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.D.; Harney, J.H.; de Lacoste, C.; Purdy, P. How the brain, integrates affective and propositional language into a unified brain function. Hypotheses based on clinicopathological correlations. Arch. Neurol. 1981, 38, 745–748. [Google Scholar] [CrossRef] [PubMed]
- Speedie, L.J.; Coslett, B.; Heilman, K.M. Repetition of affective prosody in mixed transcortical aphasia. Arch. Neurol. 1984, 41, 268–270. [Google Scholar] [CrossRef]
- Mitchell, R.L.; Ross, E.D. fMRI evidence for the effect of verbal complexity on lateralisation of the neural response associated with decoding prosodic emotion. Neuropsychologia 2008, 46, 2880–2887. [Google Scholar] [CrossRef]
- Boersma, P.; Weenink, D. Praat: Doing Phonetics by Computer; Phonetic Sciences, University of Amsterdam: Amsterdam, The Netherlands, 2008; Available online: https://www.fon.hum.uva.nl/praat/ (accessed on 31 October 2023).
- Monnot, M.; Orbelo, D.; Riccardo, L.; Sikka, S.; Pleasant, H.; Ross, E. Acoustic analyses support subjective judgements of vocal emotion. Ann. N. Y. Acad. Sci. 2003, 1000, 288–292. [Google Scholar] [CrossRef]
- Geigenberger, A.; Ziegler, W. Receptive prosodic processing in aphasia. Aphasiology 2001, 15, 1169–1188. [Google Scholar] [CrossRef]
- Selnes, O.A.; Knopman, D.S.; Niccum, N.; Rubens, A.B.; Larson, D. Computed tomographic scan correlates of auditory comprehension deficits in aphasia: A prospective recovery study. Ann. Neurol. 1983, 13, 558–566. [Google Scholar] [CrossRef]
- Parker, G.J.; Luzzi, S.; Alexander, D.C.; Wheeler-Kingshott, C.A.; Ciccarelli, O.; Lambon Ralph, M.A. Lateralization of ventral and dorsal auditory-language pathways in the human brain. Neuroimage 2005, 24, 656–666. [Google Scholar] [CrossRef]
- Starkstein, S.E.; Federoff, J.P.; Price, T.R.; Leiguarda, R.C.; Robinson, R.G. Neuropsychological and neuroradiological correlates of emotional prosody comprehension. Neurology 1994, 44, 515–522. [Google Scholar] [CrossRef]
- Ermine, C.M.; Bivard, A.; Parsons, M.W.; Baron, J.C. The ischemic penumbra: From concept to reality. Int. J. Stroke 2021, 16, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Hillis, A.E.; Barker, P.B.; Beauchamp, N.J.; Gordon, B.; Wityk, R.J. MR perfusion imaging reveals regions of hypoperfusion associated with aphasia and neglect. Neurology 2000, 55, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Hillis, A.E. Magnetic resonance perfusion imaging in the study of language. Brain Lang. 2007, 102, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.M.; Cao, Y.; Nagesh, V. Magnetic resonance assessment of acute and chronic stroke. Prog. Cardiovasc. Dis. 2000, 43, 113–134. [Google Scholar] [CrossRef] [PubMed]
- Vymazal, J.; Rulseh, A.M.; Keller, J.; Janouskova, L. Comparison of CT and MR imaging in ischemic stroke. Insights Imaging 2012, 3, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Becker, H.; Desch, H.; Hacker, H.; Pencz, A. CT fogging effect with ischemic cerebral infarcts. Neuroradiology 1979, 31, 185–192. [Google Scholar] [CrossRef]
- Skriver, E.B.; Olsen, T.S. Transient disappearance of cerebral infarcts on CT scan, the so-called fogging effect. Neuroradiology 1981, 22, 61–65. [Google Scholar] [CrossRef]
- Patel, S.; Oishi, K.; Wright, A.; Sutherland-Foggio, H.; Saxena, S.; Sheppard, S.M.; Hillis, A.E. Right hemisphere regions critical for expression of emotion through prosody. Front. Neurol. 2018, 9, 224. [Google Scholar] [CrossRef]
- Cohen, M.J.; Riccio, C.A.; Flannery, A.M. Expressive aprosodia following stroke to the right basal ganglia: A case report. Neuropsychology 1994, 8, 242–245. [Google Scholar] [CrossRef]
- Denes, G.; Caldognetto, E.M.; Semenza, C.; Vagges, K.; Zettin, M. Discrimination and identification of emotions in human voice by brain damaged subjects. Acta Neurol. Scand. 1984, 69, 154–162. [Google Scholar] [CrossRef]
- Darby, D.G. Sensory aprosodia: A clinical clue to lesions of the inferior division of the right middle cerebral artery? Neurology 1993, 34, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, S.M.; Keator, L.M.; Breining, B.L.; Wright, A.E.; Saxena, S.; Tippett, D.C.; Hillis, A.E. Right hemisphere ventral stream for emotional prosody identification: Evidence from acute stroke. Neurology 2020, 94, e1013–e1020. [Google Scholar] [CrossRef]
- Wolfe, G.I.; Ross, E.D. Sensory aprosodia with left hemiparesis from subcortical infarction. Right hemisphere analogue of sensory-type aphasia with right hemiparesis? Arch. Neurol. 1987, 44, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Bell, W.L.; Davis, D.L.; Morgan-Fisher, A.; Ross, E.D. Acquired aprosodia in children. J. Child Neurol. 1990, 5, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Stringer, A.Y.; Hodnett, C. Transcortical motor aprosodia: Functional and anatomical correlates. Arch. Clin. Neuropsychol. 1991, 6, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Heilman, K.M.; Leon, S.A.; Rosenbek, J.C. Affective aprosodia from a medial frontal stroke. Brain Lang. 2004, 89, 411–416. [Google Scholar] [CrossRef]
- Hornak, J.; Rolls, E.T.; Wade, D. Face and voice expression identification in patients with emotional and behavioural changes following ventral frontal lobe damage. Neuropsychologia 1996, 34, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Hornak, J.; Bramham, J.; Rolls, E.T.; Morris, R.G.; O’Doherty, J.; Bullock, P.R.; Polkey, C.E. Changes in emotion after circumscribed surgical lesions of the orbitofrontal and cingulate cortices. Brain 2003, 126, 1691–1712. [Google Scholar] [CrossRef]
- Ross, E.D.; Monnot, M. Affective prosody: What do comprehension errors tell us about hemispheric lateralization of emotions, sex and aging effects. Neuropsychologia 2011, 49, 866–877. [Google Scholar] [CrossRef]
- George, M.S.; Parekh, P.I.; Rosinsky, N.; Ketter, T.A.; Kimbrell, T.A.; Heilman, K.M.; Herscovitch, P.; Post, R.M. Understanding emotional prosody activates right hemisphere regions. Arch. Neurol. 1996, 53, 665–670. [Google Scholar] [CrossRef]
- Buchanan, T.W.; Lutz, K.; Mirzazade, S.; Specht, K.; Shah, N.J.; Zilles, K.; Jäncke, L. Recognition of emotional prosody and verbal components of spoken language: An fMRI study. Cogn. Brain Res. 2000, 9, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Van Lancker Sidtis, D. Does functional neuroimaging solve the questions of neurolinguistics? Brain Lang. 2006, 98, 276–290. [Google Scholar] [CrossRef]
- Wildgruber, D.; Pihan, H.; Ackermann, H.; Erb, M.; Grodd, W. Dynamic brain activation during processing of emotional intonation: Influence of acoustic parameters, emotional valence, and sex. NeuroImage 2002, 15, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Sidtis, J.J. Some problems for representations of brain organization based on activation in functional imaging. Brain Lang. 2007, 102, 130–140. [Google Scholar] [CrossRef]
- Mariën, P.; Keulen, S.; Verhoeven, J. Neurological aspects of foreign accent syndrome in stroke patients. J. Commun. Disord. 2019, 77, 94–113. [Google Scholar] [CrossRef]
- Weintraub, S.; Mesulam, M.M.; Kramer, L. Disturbances in prosody. Arch. Neurol. 1981, 38, 742–744. [Google Scholar] [CrossRef]
- Ouellette, G.P.; Baum, S.R. Acoustic analysis of prosodic cues in left- and right-hemisphere-damaged patients. Aphasiology 1994, 8, 257–283. [Google Scholar] [CrossRef]
- Hollen, C.; Ross, E.D.; Ray, B. Prosodic stress: Acoustic changes in spoken English in patients with left and right hemisphere lesions. Neuorolgy 2019, 92 (Suppl. S15), P3.1-012. Available online: https://n.neurology.org/content/92/15_Supplement/P3.1-012 (accessed on 31 October 2023).
- Winner, E.; Gardner, H. The comprehension of metaphor in brain-damaged patients. Brain 1997, 100, 717–729. [Google Scholar] [CrossRef]
- Cicone, M.; Wapner, W.; Gardner, H. Sensitivity to emotional expressions and situations in organic patients. Cortex 1980, 16, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Delis, D.; Wapner, W.; Gardner, H.; Moses, J.A. The contribution of the right hemisphere to the organization of paragraphs. Cortex 1983, 19, 43–50. [Google Scholar] [CrossRef]
- Brownell, H.H.; Potter, H.H.; Michelow, D.; Gardner, H. Sensitivity to lexical denotation and connotation in brain damaged patients: A double dissociation? Brain Lang. 1984, 22, 253–265. [Google Scholar] [CrossRef]
- Brownell, H.H.; Potter, H.H.; Bihrle, A. Inference deficits in right brain-damaged patients. Brain Lang. 1986, 29, 310–321. [Google Scholar] [CrossRef]
- Foldi, N.S. Appreciation of pragmatic interpretations of indirect commands: Comparison of right and left brain damaged patients. Brain Lang. 1987, 31, 88–108. [Google Scholar] [CrossRef]
- Van Lancker, D.; Kempler, D. Comprehension of familiar phrases by left- but not right-hemisphere damaged patients. Brain Lang. 1987, 32, 256–277. [Google Scholar] [CrossRef]
- Van Lancker, D. The neurology of proverbs. Behav. Neurol. 1990, 3, 169–187. [Google Scholar] [CrossRef]
- Hough, M.S. Narrative comprehension in adults with right and left hemisphere brain-damage: Theme organization. Brain Lang. 1990, 38, 253–277. [Google Scholar] [CrossRef]
- Borod, J.C.; Andelman, F.; Obler, L.K.; Tweedy, J.R.; Welkowitz, J. Right hemisphere specialization for the identification of emotional words and sentences: Evidence from stroke patients. Neuropsychologia 1992, 30, 827–844. [Google Scholar] [CrossRef] [PubMed]
- Rehak, A.; Kaplan, J.; Weylman, S.; Kelly, B.; Brownell, H.H.; Gardner, H. Story processing in right hemisphere brain damaged patients. Brain Lang. 1992, 42, 320–336. [Google Scholar] [CrossRef] [PubMed]
- Schneiderman, E.; Murasugi, K.; Saddy, D. Story arrangement ability in right-brain damaged patients. Brain Lang. 1992, 43, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Marini, A.; Carlomagno, S.; Caltagirone, C.; Nocentini, U. The role played by the right hemisphere in the organization of complex textual structures. Brain Lang. 2005, 93, 46–54. [Google Scholar] [CrossRef]
- Mashal, N.; Faust, M.; Hendler, T.; Jung-Beeman, M. Hemispheric differences in processing the literal interpretation of idioms: Converging evidence from behavioral and fMRI studies. Cortex 2008, 44, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Rush, A.J.; Beck, A.T. Cognitive therapy of depression and suicide. Am. J. Psychother. 1978, 32, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, M.; Rush, A.J.; Beck, A.T.; Hollon, S.D. Depressed outpatients treated with cognitive therapy or pharmacotherapy. A one-year follow-up. Arch. Gen. Psychiatry 1981, 38, 33–39. [Google Scholar] [CrossRef]
- Beck, A.T.; Rush, A.J.; Shaw, B.F.; Emery, G. Cognitive Therapy for Depresssion; Guilford Press: New York, NY, USA, 1987. [Google Scholar]
- Carroll, B.J.; Martin, F.I.; Davies, B. Resistance to suppression by dexamethasone of plasma 11-O.H.C.S. levels in severe depressive illness. Br. Med. J. 1968, 3, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Carroll, B.J.; Feinberg, M.; Greden, J.F.; Tarika, J.; Albala, A.A.; Haskett, R.F.; James, N.M.; Kronfol, Z.; Lohr, N.; Steiner, M.; et al. A specific laboratory test for the diagnosis of melancholia. Standardization, validation, and clinical utility. Arch. Gen. Psychiatry 1981, 38, 15–22. [Google Scholar] [CrossRef]
- Schlesser, M.A.; Winokur, G.; Sherman, B.M. Hypothalamic-pituitary-adrenal axis activity in depressive illness. Its relationship to classification. Arch. Gen. Psychiatry 1980, 37, 737–743. [Google Scholar] [CrossRef]
- Carroll, B.J. Dexamethasone suppression test: A review of contemporary confusion. J. Clin. Psychiatry 1985, 46 Pt 2, 13–24. [Google Scholar]
- Rush, A.J.; Giles, D.E.; Schlesser, M.A.; Orsulak, P.J.; Parker, C.R.; Weissenburger, J.E.; Crowley, G.T.; Khatami, M.; Vasavada, N. The dexamethasone suppression test in patients with mood disorders. J. Clin. Psychiatry 1996, 57, 470–484. [Google Scholar] [CrossRef]
- Schumacher, M.M.; Santambrogio, J. Cortisol and the dexamethasone suppression test as a biomarker for melancholic depression: A narrative review. J. Pers. Med. 2023, 13, 837. [Google Scholar] [CrossRef] [PubMed]
- Parker, G. Classifying depression: Should paradigms lost be regained? Am. J. Psychiatry 2000, 157, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Parker, G. Defining melancholia: The primacy of psychomotor disturbance. Acta Psychiatr Scand. 2007, 15 (Suppl. S433), 21–50. [Google Scholar] [CrossRef] [PubMed]
- Shorter, E. The doctrine of the two depressions in historical perspective. Acta Psychiatr. Scand. Suppl. 2007, 115, 5–13. [Google Scholar] [CrossRef]
- Fink, M.; Bolwig, T.G.; Parker, G.; Shorter, E. Melancholia: Restoration in psychiatric classification recommended. Acta Psychiatr. Scand. 2007, 115, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Fink, M.; Taylor, M.A. Resurrecting melancholia. Acta Psychiatr. Scand. 2007, 15 (Suppl. S433), 14–20. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 3rd ed.; American Psychiatric Association: Washington, DC, USA, 1980. [Google Scholar]
- Rasmussen, K.G. Electroconvulsive therapy and melancholia: Review of the literature and suggestions for further study. J. ECT 2011, 27, 315–322. [Google Scholar] [CrossRef]
- Mizushima, J.; Sakurai, H.; Mizuno, Y.; Shinfuku, M.; Tani, H.; Yoshida, K.; Ozawa, C.; Serizawa, A.; Kodashiro, N.; Koide, S.; et al. Melancholic and reactive depression: A reappraisal of old categories. BMC Psychiatry 2013, 13, 311. [Google Scholar] [CrossRef]
- Ross, E.D.; Rush, A.J. Diagnostic issues and neuroanatomical correlates of depression in brain damaged patients: Implications for a neurology of depression. Arch. Gen. Psychiatry 1981, 38, 344–1354. [Google Scholar] [CrossRef]
- Ross, E.D. Acute agitation and other behaviors associated with Wernicke aphasia and their possible neurological bases. Neuropsychiatry Neuropsychol. Behav. Neurol. 1993, 6, 9–18. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 2nd ed.; American Psychiatric Association: Washington, DC, USA, 1968. [Google Scholar]
- Ross, E.D.; Stewart, R. Pathological display of affect in patients with depression and right focal brain damage: An alternative mechanism. J. Nerv. Ment. Dis. 1978, 175, 165–172. [Google Scholar] [CrossRef]
- Gordon, W.A.; Hibbard, M.R.; Egelko, S.; Riley, E.; Simon, D.; Diller, L.; Ross, E.D.; Lieberman, A. Issues in the diagnosis of post-stroke depression. Rehab. Psychol. 1991, 36, 71–86. [Google Scholar] [CrossRef]
- Harney, J.; Fulton, C.; Ross, E.D.; Rush, A.J. Dexamethasone suppression test and onset of poststroke depression in patients with ischemic infarction. J. Clin. Psychiatry 1993, 54, 343–348. [Google Scholar] [PubMed]
- Buck, R.; Duffy, R. Nonverbal communication of affect in brain-damaged patients. Cortex 1980, 16, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Buck, R. Human Motivation and Emotion, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1988. [Google Scholar]
- Buck, R. The biological affects: A typology. Psychol. Rev. 1999, 106, 301–336. [Google Scholar] [CrossRef]
- Ross, E.D.; Orbelo, D.M.; Cartwright, J.; Hansel, S.; Burgard, M.; Testa, J.A.; Buck, R. Affective-prosodic deficits in schizophrenia: Comparison to patients with brain damage and relation to schizophrenic symptoms [corrected]. J. Neurol. Neurosurg. Psychiatry 2001, 70, 597–604. [Google Scholar] [CrossRef]
- Hareli, S.; Parkinson, B. What’s social about social emotions? J. Theory Soc. Behav. 2008, 38, 131–156. [Google Scholar] [CrossRef]
- Leventhal, H. Emotions: A basic problem for social psychology. In Social Psychology: Classic and Contemporary Integrations; Nemeth, C., Ed.; Rand McNally: Chicago, IL, USA, 1974; pp. 1–51. [Google Scholar]
- Leventhal, H. A perceptual motor theory of emotion. Soc. Sci. Inf. 1982, 21, 819–845. [Google Scholar] [CrossRef]
- Leventhal, H.; Tomarken, A.J. Emotion: Todays’s problem. Ann. Rev. Psychol. 1986, 37, 565–610. [Google Scholar] [CrossRef]
- Wilson, S.A.K. Some problems in neurology. II: Pathological laughing and crying. J. Neurol. Psychopathol. 1924, 4, 299–333. [Google Scholar] [CrossRef]
- Black, D.W. Pathological laughter: A review of the literature. J. Nerv. Ment. Dis. 1982, 170, 67–71. [Google Scholar] [CrossRef]
- Fernald, A. Human maternal vocalizations to infants as biologically relevant signals: An evolutionary perspective. In The Adapted Mind: Evolutionary Psychology and the Generation of Culture; Barkow, J.H., Cosmides, L., Tooby, J., Eds.; Oxford University Press: New York, NY, USA, 1992; pp. 391–428. [Google Scholar]
- Monnot, M.; Foley, R.; Ross, E.D. Affective prosody: Whence motherese. Behav. Brain Sci. 2004, 27, 518–519. [Google Scholar] [CrossRef]
- Johnson, M.H. Subcortical face processing. Nat. Rev. Neurosci. 2005, 6, 766–774. [Google Scholar] [CrossRef]
- Andre, M.; Plenat, F.; Floquet, J.; Picard, L. Hydranencephalie. Lesions cerebrales majeures avec comportement neurologique neo-natal normal [Hydranencephaly. Major cerebral lesions with normal neonatal neurologic behavior]. Arch. Fr. Pediatr. 1975, 32, 915–924. [Google Scholar] [PubMed]
- Francis, P.L.; Self, P.A.; McCaffree, M.A. Behavioral assessment of a hydranencephalic neonate. Child Dev. 1984, 55, 262–266. [Google Scholar] [CrossRef]
- Merker, B. Consciousness without a cerebral cortex: A challenge for neuroscience and medicine. Behav. Brain Sci. 2007, 30, 63–81, discussion 81–134. [Google Scholar] [CrossRef]
- Beshkar, M. The presence of consciousness in the absence of the cerebral cortex. Synapse 2008, 62, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Feinman, S. Social referencing in infancy. Merrill Palmer Q. 1982, 28, 445–470. [Google Scholar]
- Walden, T.A.; Ogan, T.A. The development of social referencing. Child Dev. 1988, 59, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. The Expression of the Emotions in Man and Animals; reproduction of 1872 publication; Philosophical Library Edition: London, UK, 1955. [Google Scholar]
- Ekman, P.; Friesen, W.V. Constants across cultures in the face and emotion. J. Pers. Soc. Psychol. 1971, 17, 124–129. [Google Scholar] [CrossRef]
- Ekman, P.; Friesen, W.V.; Ellsworth, P. Emotion in the Human Face; Pergammon Press: Elmsford, NY, USA, 1972. [Google Scholar]
- Izard, C.E. Human Emotions; Plenum Press: New York, NY, USA, 1977. [Google Scholar]
- Panksepp, J. Affective Neuroscience: The Foundations of Human and Animal Emotions; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Ross, E.D. Cortical Representation of the Emotions. In Contemporary Behavioral Neurology; Trimble, M.R., Cummings, J.L., Eds.; Butterworth-Heineman: Woburn, MA, USA, 1997; pp. 107–126. [Google Scholar]
- Kraemer, G.W. A psychobiological theory of attachment. Behav. Brain Sci. 1992, 15, 467–493. [Google Scholar] [CrossRef]
- Harlow, H.F.; Mears, C.E. Emotional Sequences and Consequences. In Emotion: Theory, Research, and Experience, Vol. 2: Emotions in Early Development; Plutchik, R., Kellerman, H., Eds.; Academic Press: New York, NY, USA, 1983; Chapter 6; pp. 171–197. [Google Scholar]
- Buck, R.; Losow, J.I.; Murphy, M.M.; Costanzo, P. Social facilitation and inhibition of emotional expression and communication. J. Personal. Soc. Psychol. 1992, 63, 962–968. [Google Scholar] [CrossRef]
- Ekman, P.; Friesen, W.V. Unmasking the Face; Prentice-Hall: Engelwood Cliffs, NJ, USA, 1975. [Google Scholar]
- Saarni, C. Children’s understanding of display rules for expressive behavior. Dev. Psychol. 1979, 15, 424–429. [Google Scholar] [CrossRef]
- Saarni, C. An observational study of children’s attempts to monitor their expressive behavior. Child Dev. 1984, 55, 1504–1513. [Google Scholar] [CrossRef]
- Malatesta, C.Z.; Haviland, J.M. Learning display rules: The socialization of emotion expression in infancy. Child Dev. 1982, 53, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Malatesta, C.Z.; Kalnok, D. Emotional experience in younger and older adults. J. Gerontol. 1984, 39, 301–308. [Google Scholar] [CrossRef]
- Ekman, P.; Friesen, W.V. Felt, false and miserable smiles. J. Nonverbal Behav. 1982, 6, 238–252. [Google Scholar] [CrossRef]
- Ross, E.D.; Gupta, S.S.; Adnan, A.M.; Holden, T.L.; Havlicek, J.; Radhakrishnan, S. Neurophysiology of spontaneous facial expressions: I. Motor control of the upper and lower face is behaviorally independent in adults. Cortex 2016, 76, 28–42. [Google Scholar] [CrossRef]
- Ross, E.D. Hemispheric specialization for emotions, affective aspects of language and communication and the cognitive control of display behaviors in humans. Prog. Brain Res. 1996, 107, 583–594. [Google Scholar] [PubMed]
- Ross, E.D.; Gupta, S.S.; Adnan, A.M.; Holden, T.L.; Havlicek, J.; Radhakrishnan, S. Neurophysiology of spontaneous facial expressions: II. Motor control of the right and left face is partially independent in adults. Cortex 2019, 111, 164–182. [Google Scholar] [CrossRef]
- Ekman, P. Darwin, deception, and facial expressions. Ann. N. Y. Acad. Sci. 2003, 1000, 205–221. [Google Scholar] [CrossRef]
- Ekman, P.; Friesen, V.W. Nonverbal leakage and clues to deception. Psychiatry 1969, 32, 88–106. [Google Scholar] [CrossRef] [PubMed]
- Ekman, P.; Friesen, W.V.; O’Sullivan, M. Smiles when lying. J. Personal. Soc. Psychol. 1988, 54, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Ekman, P. Telling Lies: Clues to Deceit in the Marketplace, Politics and Marriage; Norton Press: New York, NY, USA, 1992. [Google Scholar]
- Schacter, S.C.; Devinsky, O. (Eds.) Behavioral Neurology and the Legacy of Norman Geschwind; Lippincott-Raven: Philadelphia, PA, USA, 1997. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| APHASIAS | Spontaneous Speech | Verbal Repetition | Verbal Comprehension | Verbal Naming |
| Motor | non-fluent | poor | good | poor § |
| Sensory | fluent * | poor | poor | poor |
| Conduction | fluent | poor | good | poor |
| Global | non-fluent | poor | poor | poor |
| Transcortical Motor | non-fluent | good | good | poor § |
| Transcortical Sensory | fluent | good | poor | poor |
| Mixed Transcortical | non-fluent | good | poor | poor |
| Anomic | fluent | good | good | poor |
| APROSODIAS | Spontaneous Affective Prosody | Affective Prosodic Repetition | Affective Prosodic Comprehension | Facial Expression Naming |
| Motor | poor | poor | good | good |
| Sensory | good | poor | poor | poor |
| Conduction | good | poor | good | poor |
| Global | poor | poor | poor | poor |
| Transcortical Motor | poor | good | good | good |
| Transcortical Sensory | good | good | poor | poor |
| Mixed Transcortical | poor | good | poor | poor |
| Agesic | good | good | good | poor |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ross, E.D. Affective Prosody and Its Impact on the Neurology of Language, Depression, Memory and Emotions. Brain Sci. 2023, 13, 1572. https://doi.org/10.3390/brainsci13111572
Ross ED. Affective Prosody and Its Impact on the Neurology of Language, Depression, Memory and Emotions. Brain Sciences. 2023; 13(11):1572. https://doi.org/10.3390/brainsci13111572
Chicago/Turabian StyleRoss, Elliott D. 2023. "Affective Prosody and Its Impact on the Neurology of Language, Depression, Memory and Emotions" Brain Sciences 13, no. 11: 1572. https://doi.org/10.3390/brainsci13111572
APA StyleRoss, E. D. (2023). Affective Prosody and Its Impact on the Neurology of Language, Depression, Memory and Emotions. Brain Sciences, 13(11), 1572. https://doi.org/10.3390/brainsci13111572