
Within-Person Modulation of Neural Networks following Interoceptive Awareness Training through Mindful Awareness in Body-Oriented Therapy (MABT): A Pilot Study


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design Overview, Ethics Statement, and Experiment Power
2.2. Participants
2.3. Measures
2.4. Intervention: Mindful Awareness in Body-Oriented Therapy
2.5. Data Collection Procedures
2.6. The Interoceptive/Exteroceptive Attention Task
2.7. Interoceptive Awareness Meditation Task
2.8. Functional Imaging Data Acquisition
2.9. Image Processing
2.10. Preliminary Analyses
2.11. Neuroimaging Analysis
3. Results
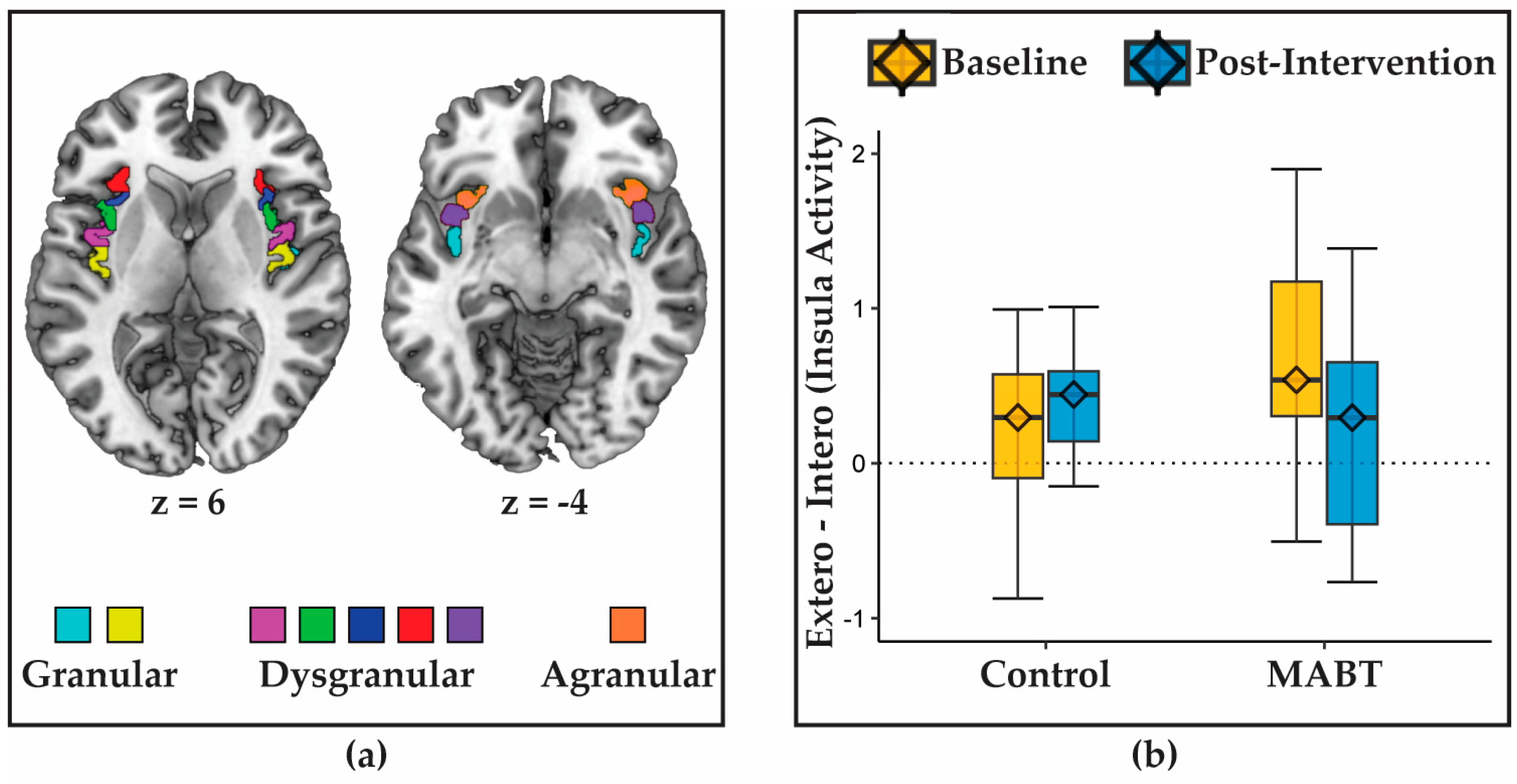
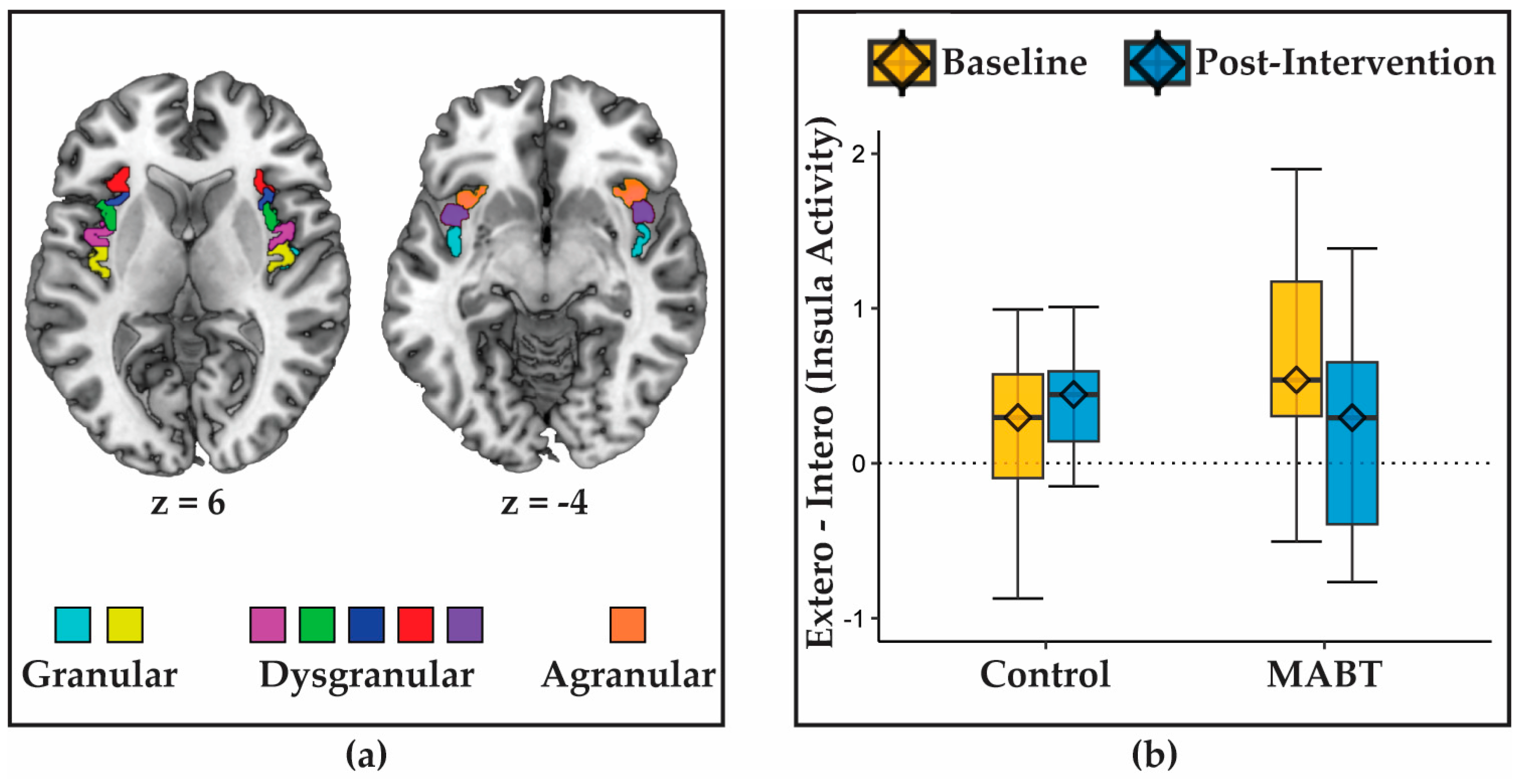
3.1. MABT Reduces Interoception-Related Insula Deactivation
3.2. MABT Increases Interoceptive Sensibility
3.3. MABT Increases Insula Connectivity with the Dorsal Attention Network
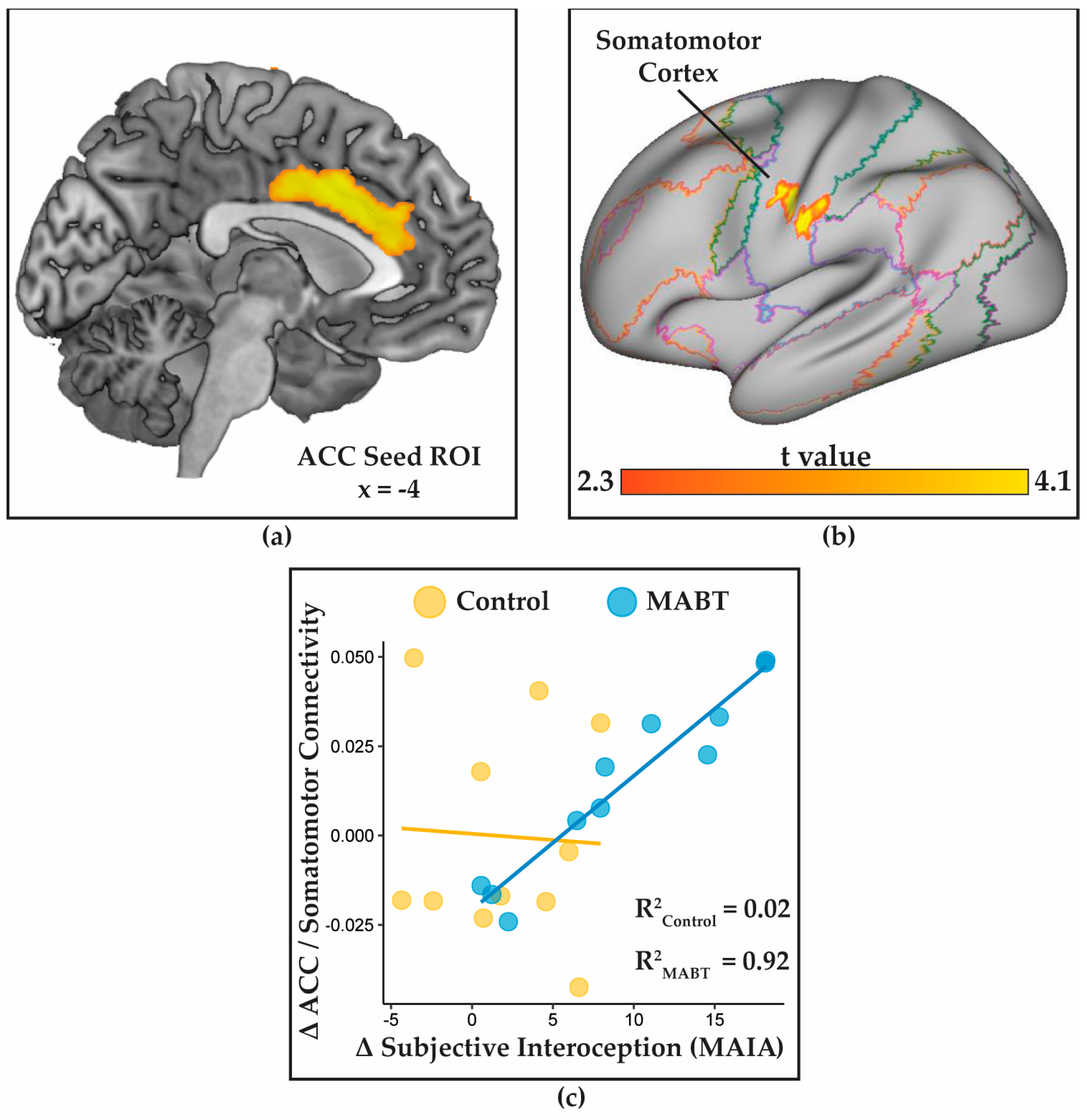
3.4. MABT Increases ACC Connectivity with the Somatosmotor Cortex
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Craig, A.D. Interoception: The Sense of the Physiological Condition of the Body. Curr. Opin. Neurobiol. 2003, 13, 500–505. [Google Scholar] [CrossRef]
- Rolls, A. Immunoception: The Insular Cortex Perspective. Cell Mol. Immunol. 2023, 1–7. [Google Scholar] [CrossRef]
- Hsueh, B.; Chen, R.; Jo, Y.; Tang, D.; Raffiee, M.; Kim, Y.S.; Inoue, M.; Randles, S.; Ramakrishnan, C.; Patel, S.; et al. Cardiogenic Control of Affective Behavioural State. Nature 2023, 615, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Livneh, Y.; Sugden, A.U.; Madara, J.C.; Essner, R.A.; Flores, V.I.; Sugden, L.A.; Resch, J.M.; Lowell, B.B.; Andermann, M.L. Estimation of Current and Future Physiological States in Insular Cortex. Neuron 2020, 105, 1094–1111.e10. [Google Scholar] [CrossRef]
- Verdejo-Garcia, A.; Clark, L.; Dunn, B.D. The Role of Interoception in Addiction: A Critical Review. Neurosci. Biobehav. Rev. 2012, 36, 1857–1869. [Google Scholar] [CrossRef]
- Khalsa, S.S.; Adolphs, R.; Cameron, O.G.; Critchley, H.D.; Davenport, P.W.; Feinstein, J.S.; Feusner, J.D.; Garfinkel, S.N.; Lane, R.D.; Mehling, W.E.; et al. Interoception and Mental Health: A Roadmap. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 501–513. [Google Scholar] [CrossRef]
- Craig, A.D. How Do You Feel--Now? The Anterior Insula and Human Awareness. Nat. Rev. Neurosci. 2009, 10, 59–70. [Google Scholar] [CrossRef]
- Farb, N.A.; Segal, Z.V.; Anderson, A.K. Attentional Modulation of Primary Interoceptive and Exteroceptive Cortices. Cereb. Cortex 2013, 23, 114–126. [Google Scholar] [CrossRef]
- Kleckner, I.R.; Zhang, J.; Touroutoglou, A.; Chanes, L.; Xia, C.; Simmons, W.K.; Quigley, K.S.; Dickerson, B.C.; Feldman Barrett, L. Evidence for a Large-Scale Brain System Supporting Allostasis and Interoception in Humans. Nat. Hum. Behav. 2017, 1, 0069. [Google Scholar] [CrossRef]
- Quadt, L.; Critchley, H.D.; Garfinkel, S.N. The Neurobiology of Interoception in Health and Disease: Neuroscience of Interoception. Ann. N. Y. Acad. Sci. 2018, 1428, 112–128. [Google Scholar] [CrossRef]
- Noël, X.; Brevers, D.; Bechara, A. A Neurocognitive Approach to Understanding the Neurobiology of Addiction. Curr. Opin. Neurobiol. 2013, 23, 632–638. [Google Scholar] [CrossRef]
- Schulz, A. Interoception and Stress. Front. Psychol. 2015, 6, 993. [Google Scholar] [CrossRef]
- Seth, A.K. Interoceptive Inference, Emotion, and the Embodied Self. Trends Cogn. Sci. 2013, 17, 565–573. [Google Scholar] [CrossRef]
- Alvarez, R.P.; Kirlic, N.; Misaki, M.; Bodurka, J.; Rhudy, J.L.; Paulus, M.P.; Drevets, W.C. Increased Anterior Insula Activity in Anxious Individuals Is Linked to Diminished Perceived Control. Transl. Psychiatry 2015, 5, e591. [Google Scholar] [CrossRef]
- Simmons, A.; Matthews, S.C.; Stein, M.B.; Paulus, M.P. Anticipation of Emotionally Aversive Visual Stimuli Activates Right Insula. Neuroreport 2004, 15, 2261–2265. [Google Scholar] [CrossRef]
- Farb, N.A.; Anderson, A.K.; Mayberg, H.; Bean, J.; McKeon, D.; Segal, Z.V. Minding One’s Emotions: Mindfulness Training Alters the Neural Expression of Sadness. Emotion 2010, 10, 25–33. [Google Scholar] [CrossRef]
- Farb, N.A.S.; Desormeau, P.; Anderson, A.K.; Segal, Z.V. Static and Treatment-Responsive Brain Biomarkers of Depression Relapse Vulnerability Following Prophylactic Psychotherapy: Evidence from a Randomized Control Trial. NeuroImage Clin. 2022, 34, 102969. [Google Scholar] [CrossRef]
- Le Moal, M.; Koob, G.F. Drug Addiction: Pathways to the Disease and Pathophysiological Perspectives. Eur. Neuropsychopharmacol. 2007, 17, 377–393. [Google Scholar] [CrossRef]
- Medford, N.; Critchley, H.D. Conjoint Activity of Anterior Insular and Anterior Cingulate Cortex: Awareness and Response. Brain Struct. Funct. 2010, 214, 535–549. [Google Scholar] [CrossRef]
- Farb, N.A.S.; Daubenmier, J.; Price, C.J.; Gard, T.; Kerr, C.; Dunn, B.D.; Klein, A.C.; Paulus, M.P.; Mehling, W.E. Interoception, Contemplative Practice, and Health. Front. Psychol. 2015, 6, 763. [Google Scholar] [CrossRef]
- Vago, D.R.; Silbersweig, D.A. Self-Awareness, Self-Regulation, and Self-Transcendence (S-ART): A Framework for Understanding the Neurobiological Mechanisms of Mindfulness. Front. Hum. Neurosci. 2012, 6, 296. [Google Scholar] [CrossRef] [PubMed]
- Damasio, A.; Carvalho, G.B. The Nature of Feelings: Evolutionary and Neurobiological Origins. Nat. Rev. Neurosci. 2013, 14, 143–152. [Google Scholar] [CrossRef]
- Lane, R.D. Neural Substrates of Implicit and Explicit Emotional Processes: A Unifying Framework for Psychosomatic Medicine. Psychosom. Med. 2008, 70, 214–231. [Google Scholar] [CrossRef] [PubMed]
- Pais-Vieira, C.; Wing, E.A.; Cabeza, R. The Influence of Self-Awareness on Emotional Memory Formation: An FMRI Study. Social. Cogn. Affect. Neurosci. 2016, 11, 580–592. [Google Scholar] [CrossRef]
- Farb, N.A.; Segal, Z.V.; Anderson, A.K. Mindfulness Meditation Training Alters Cortical Representations of Interoceptive Attention. Social. Cogn. Affect. Neurosci. 2013, 8, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Sevinc, G.; Hölzel, B.K.; Greenberg, J.; Gard, T.; Brunsch, V.; Hashmi, J.A.; Vangel, M.; Orr, S.P.; Milad, M.R.; Lazar, S.W. Strengthened Hippocampal Circuits Underlie Enhanced Retrieval of Extinguished Fear Memories Following Mindfulness Training. Biol. Psychiatry 2019, 86, 693–702. [Google Scholar] [CrossRef]
- Paulus, M.P.; Stein, M.B. Interoception in Anxiety and Depression. Brain Struct. Funct. 2010, 214, 451–463. [Google Scholar] [CrossRef]
- Barrett, F.S.; Bradstreet, M.P.; Leoutsakos, J.-M.S.; Johnson, M.W.; Griffiths, R.R. The Challenging Experience Questionnaire: Characterization of Challenging Experiences with Psilocybin Mushrooms. J. Psychopharmacol. 2016, 30, 1279–1295. [Google Scholar] [CrossRef]
- Naqvi, N.H.; Bechara, A. The Insula and Drug Addiction: An Interoceptive View of Pleasure, Urges, and Decision-Making. Brain Struct. Funct. 2010, 214, 435–450. [Google Scholar] [CrossRef]
- Paulus, M.P.; Stewart, J.L. Interoception and Drug Addiction. Neuropharmacology 2014, 76, 342–350. [Google Scholar] [CrossRef]
- Kerr, K.L.; Moseman, S.E.; Avery, J.A.; Bodurka, J.; Zucker, N.L.; Simmons, W.K. Altered Insula Activity during Visceral Interoception in Weight-Restored Patients with Anorexia Nervosa. Neuropsychopharmacology 2016, 41, 521–528. [Google Scholar] [CrossRef]
- Mallorquí-Bagué, N.; Garfinkel, S.N.; Engels, M.; Eccles, J.A.; Pailhez, G.; Bulbena, A.; Critchley, H.D. Neuroimaging and Psychophysiological Investigation of the Link between Anxiety, Enhanced Affective Reactivity and Interoception in People with Joint Hypermobility. Front. Psychol. 2014, 5, 1162. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Wei, D.; Zhang, M.; Yang, J.; Jelinčić, V.; Qiu, J. The Role of Mid-Insula in the Relationship between Cardiac Interoceptive Attention and Anxiety: Evidence from an FMRI Study. Sci. Rep. 2018, 8, 17280. [Google Scholar] [CrossRef]
- Terasawa, Y.; Umeda, S. Neurocognitive Mechanisms of Social Anxiety and Interoception. In Island of Reil (Insula) in the Human Brain; Turgut, M., Yurttaş, C., Tubbs, R.S., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 113–120. ISBN 978-3-319-75467-3. [Google Scholar]
- Wiebking, C.; Northoff, G. Neural Activity during Interoceptive Awareness and Its Associations with Alexithymia—An FMRI Study in Major Depressive Disorder and Non-Psychiatric Controls. Front. Psychol. 2015, 6, 589. [Google Scholar] [CrossRef] [PubMed]
- Bornemann, B.; Herbert, B.M.; Mehling, W.E.; Singer, T. Differential Changes in Self-Reported Aspects of Interoceptive Awareness through 3 Months of Contemplative Training. Front. Psychol. 2014, 5, 1504. [Google Scholar] [CrossRef] [PubMed]
- de Jong, M.; Lazar, S.W.; Hug, K.; Mehling, W.E.; Hölzel, B.K.; Sack, A.T.; Peeters, F.; Ashih, H.; Mischoulon, D.; Gard, T. Effects of Mindfulness-Based Cognitive Therapy on Body Awareness in Patients with Chronic Pain and Comorbid Depression. Front. Psychol. 2016, 7, 967. [Google Scholar] [CrossRef]
- Mehling, W.E.; Acree, M.; Stewart, A.; Silas, J.; Jones, A. The Multidimensional Assessment of Interoceptive Awareness, Version 2 (MAIA-2). PLoS ONE 2018, 13, e0208034. [Google Scholar] [CrossRef]
- Price, C.J.; Wells, E.A.; Donovan, D.M.; Rue, T. Mindful Awareness in Body-Oriented Therapy as an Adjunct to Women’s Substance Use Disorder Treatment: A Pilot Feasibility Study. J. Subst. Abus. Treat. 2012, 43, 94–107. [Google Scholar] [CrossRef]
- Price, C.J.; Thompson, E.A.; Crowell, S.; Pike, K. Longitudinal Effects of Interoceptive Awareness Training through Mindful Awareness in Body-Oriented Therapy (MABT) as an Adjunct to Women’s Substance Use Disorder Treatment: A Randomized Controlled Trial. Drug Alcohol. Depend. 2019, 198, 140–149. [Google Scholar] [CrossRef]
- Price, C.J.; Hooven, C. Interoceptive Awareness Skills for Emotion Regulation: Theory and Approach of Mindful Awareness in Body-Oriented Therapy (MABT). Front. Psychol. 2018, 9, 798. [Google Scholar] [CrossRef]
- Price, C. Body-Oriented Therapy in Recovery from Child Sexual Abuse: An Efficacy Study. Altern. Ther. Health Med. 2005, 11, 46–57. [Google Scholar] [PubMed]
- Price, C. Dissociation Reduction in Body Therapy during Sexual Abuse Recovery. Complement. Ther. Clin. Pract. 2007, 13, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Price, C.J.; Smith-DiJulio, K. Interoceptive Awareness Is Important for Relapse Prevention: Perceptions of Women Who Received Mindful Body Awareness in Substance Use Disorder Treatment. J. Addict. Nurs. 2016, 27, 32–38. [Google Scholar] [CrossRef]
- Price, C.J.; Herting, J.R. Changes in Post Traumatic Stress Symptoms Among Women in Substance Use Disorder Treatment: The Mediating Role of Bodily Dissociation and Emotion Regulation. Subst. Abus. Res. Treat. 2013, 7, SART.S12426. [Google Scholar] [CrossRef] [PubMed]
- Farb, N.A.S.; Zuo, Z.; Price, C.J. Interoceptive Awareness of the Breath Preserves Attention and Language Networks amidst Widespread Cortical Deactivation: A Within-Participant Neuroimaging Study. eNeuro 2023, 10, ENEURO.0088-23.2023. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Kamarck, T.; Mermelstein, R. A Global Measure of Perceived Stress. J. Health Soc. Behav. 1983, 24, 385–396. [Google Scholar] [CrossRef]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-Mental State”. A Practical Method for Grading the Cognitive State of Patients for the Clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Mehling, W.E.; Price, C.; Daubenmier, J.J.; Acree, M.; Bartmess, E.; Stewart, A. The Multidimensional Assessment of Interoceptive Awareness (MAIA). PLoS ONE 2012, 7, e48230. [Google Scholar] [CrossRef]
- Kroenke, K.; Spitzer, R.L.; Williams, J.B.W.; Löwe, B. The Patient Health Questionnaire Somatic, Anxiety, and Depressive Symptom Scales: A Systematic Review. General. Hosp. Psychiatry 2010, 32, 345–359. [Google Scholar] [CrossRef]
- Kroenke, K.; Spitzer, R.L.; Williams, J.B.W. The PHQ-9: Validity of a Brief Depression Severity Measure. J. Gen. Intern. Med. 2001, 16, 606–613. [Google Scholar] [CrossRef]
- Spitzer, R.L.; Kroenke, K.; Williams, J.B.W.; Löwe, B. A Brief Measure for Assessing Generalized Anxiety Disorder: The GAD-7. Arch. Intern. Med. 2006, 166, 1092. [Google Scholar] [CrossRef]
- Kroenke, K.; Spitzer, R.L.; Williams, J.B.W. The PHQ-15: Validity of a New Measure for Evaluating the Severity of Somatic Symptoms. Psychosom. Med. 2002, 64, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Dinno, A. Paran: Horn’s Test of Principal Components/Factors; UTC: London, UK, 2012. [Google Scholar]
- Revelle, W. Psych: Procedures for Personality and Psychological Research. 2017. Available online: https://CRAN.R-project.org/package=psych (accessed on 4 December 2018).
- Fresco, D.M.; Moore, M.T.; van Dulmen, M.H.; Segal, Z.V.; Ma, S.H.; Teasdale, J.D.; Williams, J.M. Initial Psychometric Properties of the Experiences Questionnaire: Validation of a Self-Report Measure of Decentering. Behav. Ther. 2007, 38, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Baer, R.A.; Smith, G.T.; Lykins, E.; Button, D.; Krietemeyer, J.; Sauer, S.; Walsh, E.; Duggan, D.; Williams, J.M. Construct Validity of the Five Facet Mindfulness Questionnaire in Meditating and Nonmeditating Samples. Assessment 2008, 15, 329–342. [Google Scholar] [CrossRef]
- Esteban, O.; Markiewicz, C.J.; Blair, R.W.; Moodie, C.A.; Isik, A.I.; Erramuzpe, A.; Kent, J.D.; Goncalves, M.; DuPre, E.; Snyder, M.; et al. FMRIPrep: A Robust Preprocessing Pipeline for Functional MRI. Nat. Methods 2019, 16, 111–116. [Google Scholar] [CrossRef]
- Behzadi, Y.; Restom, K.; Liau, J.; Liu, T.T. A Component Based Noise Correction Method (CompCor) for BOLD and Perfusion Based FMRI. NeuroImage 2007, 37, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Seeley, W.W. The Salience Network: A Neural System for Perceiving and Responding to Homeostatic Demands. J. Neurosci. 2019, 39, 9878–9882. [Google Scholar] [CrossRef]
- Critchley, H.D.; Wiens, S.; Rotshtein, P.; Ohman, A.; Dolan, R.J. Neural Systems Supporting Interoceptive Awareness. Nat. Neurosci. 2004, 7, 189–195. [Google Scholar] [CrossRef]
- Pollatos, O.; Gramann, K.; Schandry, R. Neural Systems Connecting Interoceptive Awareness and Feelings. Hum. Brain Mapp. 2007, 28, 9–18. [Google Scholar] [CrossRef]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A Functional Connectivity Toolbox for Correlated and Anticorrelated Brain Networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Rorden, C.; Brett, M. Stereotaxic Display of Brain Lesions. Behav. Neurol. 2000, 12, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Marcus, D.S.; Harms, M.P.; Snyder, A.Z.; Jenkinson, M.; Wilson, J.A.; Glasser, M.F.; Barch, D.M.; Archie, K.A.; Burgess, G.C.; Ramaratnam, M.; et al. Human Connectome Project Informatics: Quality Control, Database Services, and Data Visualization. NeuroImage 2013, 80, 202–219. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis (Use R!), 2nd ed.; Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Roberts, R.L.; Ledermann, K.; Garland, E.L. Mindfulness-Oriented Recovery Enhancement Improves Negative Emotion Regulation among Opioid-Treated Chronic Pain Patients by Increasing Interoceptive Awareness. J. Psychosom. Res. 2022, 152, 110677. [Google Scholar] [CrossRef] [PubMed]
- Khoury, N.M.; Lutz, J.; Schuman-Olivier, Z. Interoception in Psychiatric Disorders: A Review of Randomized, Controlled Trials with Interoception-Based Interventions. Harv. Rev. Psychiatry 2018, 26, 250–263. [Google Scholar] [CrossRef] [PubMed]
- Datko, M.; Lutz, J.; Gawande, R.; Comeau, A.; To, M.N.; Desel, T.; Gan, J.; Desbordes, G.; Napadow, V.; Schuman-Olivier, Z. Increased Insula Response to Interoceptive Attention Following Mindfulness Training Is Associated with Increased Body Trusting among Patients with Depression. Psychiatry Res. Neuroimaging 2022, 327, 111559. [Google Scholar] [CrossRef]
- Price, C.J.; Merrill, J.O.; McCarty, R.L.; Pike, K.C.; Tsui, J.I. A Pilot Study of Mindful Body Awareness Training as an Adjunct to Office-Based Medication Treatment of Opioid Use Disorder. J. Subst. Abus. Treat. 2020, 108, 123–128. [Google Scholar] [CrossRef]
- Craig, A.D. How Do You Feel? Interoception: The Sense of the Physiological Condition of the Body. Nat. Rev. Neurosci. 2002, 3, 655–666. [Google Scholar] [CrossRef]
- Farb, N.A.; Segal, Z.V.; Mayberg, H.; Bean, J.; McKeon, D.; Fatima, Z.; Anderson, A.K. Attending to the Present: Mindfulness Meditation Reveals Distinct Neural Modes of Self-Reference. Soc. Cogn. Affect. Neurosci. 2007, 2, 313–322. [Google Scholar] [CrossRef]
- Khalsa, S.S.; Rudrauf, D.; Feinstein, J.S.; Tranel, D. The Pathways of Interoceptive Awareness. Nat. Neurosci. 2009, 12, 1494–1496. [Google Scholar] [CrossRef]
- Rolls, E.T.; Deco, G.; Huang, C.-C.; Feng, J. Prefrontal and Somatosensory-Motor Cortex Effective Connectivity in Humans. Cereb. Cortex 2023, 33, 4939–4963. [Google Scholar] [CrossRef]
- Fustos, J.; Gramann, K.; Herbert, B.M.; Pollatos, O. On the Embodiment of Emotion Regulation: Interoceptive Awareness Facilitates Reappraisal. Soc. Cogn. Affect. Neurosci. 2012, 8, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, L.A.; Suyenobu, B.Y.; Smith, S.R.; Bueller, J.A.; Goodman, T.; Creswell, J.D.; Tillisch, K.; Mayer, E.A.; Naliboff, B.D. Impact of Mindfulness-Based Stress Reduction Training on Intrinsic Brain Connectivity. Neuroimage 2011, 56, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-Y.; Tang, Y.; Tang, R.; Lewis-Peacock, J.A. Brief Mental Training Reorganizes Large-Scale Brain Networks. Front. Syst. Neurosci. 2017, 11, 6. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.B.; Craske, M.G. Treating Anxiety in 2017: Optimizing Care to Improve Outcomes. JAMA 2017, 318, 235. [Google Scholar] [CrossRef]
- Kaas, J.H.; Nelson, R.J.; Sur, M.; Lin, C.-S.; Merzenich, M.M. Multiple Representations of the Body Within the Primary Somatosensory Cortex of Primates. Science 1979, 204, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Price, C.J.; Weng, H.Y. Facilitating Adaptive Emotion Processing and Somatic Reappraisal via Sustained Mindful Interoceptive Attention. Front. Psychol. 2021, 12, 578827. [Google Scholar] [CrossRef]
- Dahl, C.J.; Lutz, A.; Davidson, R.J. Reconstructing and Deconstructing the Self: Cognitive Mechanisms in Meditation Practice. Trends Cogn. Sci. 2015, 19, 515–523. [Google Scholar] [CrossRef]
- Pinna, T.; Edwards, D.J. A Systematic Review of Associations Between Interoception, Vagal Tone, and Emotional Regulation: Potential Applications for Mental Health, Wellbeing, Psychological Flexibility, and Chronic Conditions. Front. Psychol. 2020, 11, 1792. [Google Scholar] [CrossRef]
- Buhle, J.T.; Silvers, J.A.; Wager, T.D.; Lopez, R.; Onyemekwu, C.; Kober, H.; Weber, J.; Ochsner, K.N. Cognitive Reappraisal of Emotion: A Meta-Analysis of Human Neuroimaging Studies. Cereb. Cortex 2014, 24, 2981–2990. [Google Scholar] [CrossRef]
- Ochsner, K.; Gross, J. The Cognitive Control of Emotion. Trends Cogn. Sci. 2005, 9, 242–249. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Price, C.J.; Sevinc, G.; Farb, N.A.S. Within-Person Modulation of Neural Networks following Interoceptive Awareness Training through Mindful Awareness in Body-Oriented Therapy (MABT): A Pilot Study. Brain Sci. 2023, 13, 1396. https://doi.org/10.3390/brainsci13101396
Price CJ, Sevinc G, Farb NAS. Within-Person Modulation of Neural Networks following Interoceptive Awareness Training through Mindful Awareness in Body-Oriented Therapy (MABT): A Pilot Study. Brain Sciences. 2023; 13(10):1396. https://doi.org/10.3390/brainsci13101396
Chicago/Turabian StylePrice, Cynthia J., Gunes Sevinc, and Norman A. S. Farb. 2023. "Within-Person Modulation of Neural Networks following Interoceptive Awareness Training through Mindful Awareness in Body-Oriented Therapy (MABT): A Pilot Study" Brain Sciences 13, no. 10: 1396. https://doi.org/10.3390/brainsci13101396
APA StylePrice, C. J., Sevinc, G., & Farb, N. A. S. (2023). Within-Person Modulation of Neural Networks following Interoceptive Awareness Training through Mindful Awareness in Body-Oriented Therapy (MABT): A Pilot Study. Brain Sciences, 13(10), 1396. https://doi.org/10.3390/brainsci13101396